1. Introduction
Friedreich ataxia (FRDA) is the most common inherited recessive ataxia, a progressive and degenerative neurological disease. This ataxia is due to a mutation in the frataxin gene (FXN), which is, in most cases, an expansion in a GAA repeat in the first intron [
1] leading to a reduced expression of the frataxin protein. The reduction of frataxin depends on the number of GAA repeats [
2,
3] and the protein concentration is drastically reduced in human patients [
4]. This small protein is implicated in the iron transport and metabolism in mitochondria. Frataxin allows the acceleration of a sulfur transfer step, which limits the synthesis rate of 2Fe-2S clusters in the Fe-S assembly complex. Therefore, its decrease leads first to mitochondrial dysfunction and oxidative stress and finally to the death of cells including those in the cardiac and the nervous system [
5,
6]. Physical symptoms mean age of onset in classical FRDA is during childhood (6–16 years old) [
7,
8]. Patients suffering from Friedreich ataxia have difficulties with the coordination of their movements and their balance because of the loss of neurons in the central and peripheral nervous system. The premature death of patients is due principally to the cardiac damage associated with the disease [
7,
9,
10].
Currently, there is no effective treatment for Friedreich ataxia. There are multiple potential therapies that are being explored, focusing on frataxin directly, such as the administration of frataxin protein fused with a cell-penetrating peptide [
11], the induction of frataxin expression by the TALE protein fused with VP64 [
12,
13], and the administration of an AAV coding for the frataxin gene [
14,
15,
16,
17]. Another gene therapy approach is the excision of the GAA repeat with zinc finger nucleases (ZFNs) or with the CRISPR/Cas9 technologies [
18,
19,
20]. These gene therapies are using an AAV to deliver the CRISPR system in mice. A Phase 1/2 clinical trial using an AAVrh10 to deliver the frataxin gene (LX2006) [
21] to reduce FRDA cardiac symptoms has recently been approved by the Food and Drug Administration (FDA) [
17].
Salami et al. [
17] used a new mouse model, called αMyhc, created to reproduce the cardiomyopathy of the FRDA patients. The mouse model is of primary importance to obtain pre-clinical results. Different mouse models have been developed to investigate potential treatments, but so far, none of these models is close to the real phenotype of Friedreich ataxia. Puccio and coworkers in 2001 [
22] developed the MCK-Cre cardiac mouse model expressing the Cre recombinase under the muscle creatine kinase (MCK) promoter permitting to knock out the FNX gene leading to cardiomyopathy after 5 weeks of age [
23]. That group also developed the NSE-Cre nervous system mouse model expressing the Cre recombinase under the neuron-specific promoter enolase (NSE) [
24]. The NSE-Cre model showed not only neurological deficits due to frataxin absence but also cardiac deficits leading to symptoms in both systems [
22]. These mouse models were previously used in our laboratory, but the symptoms were too severe compared to the disease in humans. Indeed, their death occurred between 30 to 90 days for NSE-Cre and MCK-Cre respectively [
15,
25]. Recently, Puccio and co-workers generated a new conditional model called Pvalbtm1(Cre)Arbr/J using the parvalbumin promotor to target proprioceptive afferent sensory neurons such as the DRG, cerebellar Purkinje cells, and interneurons in the brain, to focus on the neuropathy associated with FRDA. These mice mimic the neuropathophysiology observed in FRDA patients but with a more rapid and severe course [
16]. The KIKO model (Fxn(tm1Mkn/J) has an expansion of 230 GAA in one of the mouse Fxn genes and a deletion of the exon 4 in the other Fxn gene resulting in a robust neurobehavioral phenotype [
26]. In 2017, Chandran and coworkers [
27] created an inducible and reversible FRDAkd mouse model. This mouse model includes a transgene containing an shRNA against mouse frataxin under an H1 promotor, which was doxycycline-inducible. This permitted a significant frataxin protein reduction in all tissues. The strong decrease in mouse frataxin protein observed under doxycycline treatment was associated with a severe phenotype. The mice showed decreased locomotor activity and less coordination and were weaker and globally a phenotype, which is more similar to the Friedreich ataxia patients. However, this mouse model is inappropriate to test certain therapies targeting the human FXN gene such as deletion of the GAA repeats by CRISPR/Cas9, but the KIKO model could be used for this application.
Each mouse model has some advantages and some disadvantages, the goal is to choose the best one for the type of treatment to be investigated. The group of Pook [
28] developed two Friedreich ataxia mouse models: the YG8R and the YG8sR in which both mouse frataxin genes are knockout and a human frataxin transgene derived from a Friedreich patient is added, which is advantageous for the investigations of some potential treatments. The original YG8R contained a human FXN transgene with two consecutive GAA repeats (one containing 90 and the other 190 GAA) while the Y47R control mouse had only 9 GAA repeats [
28]. The improved YG8sR contained a human FXN transgene with only one repeat containing 250–300 GAAs [
29]. However, even this YG8sR model expressed only a weak phenotype.
We, therefore, assumed that the weak phenotype in YG8sR mice was due to an insufficient reduction of frataxin. We decided to accentuate the severity of the YG8sR phenotype by using a shRNA against human frataxin mRNA, an approach similar to the one used by Chandran et al., for the mouse frataxin gene [
27]. We initially selected an adequate shRNA by testing the effectiveness of different shRNAs to reduce frataxin in a culture of HeLa cells. The best shRNA was inserted in an AAV-PHP.B virus, which was injected intravenously, and the mice developed a more severe phenotype. The co-injection of two AAV-PHP.B codes for an anti-FXN shRNA and the frataxin gene used as a treatment prevented the development of a more severe FRDA phenotype.
In parallel, we compared this model to the new YG8-800 developed by Jackson Laboratory from the YG8sR descendants containing 800 GAA repeats instead of 300. The hypothesis here was that since a longer GAA expansion in humans correlates with a more severe phenotype [
8], the same trend would be observed in mice.
2. Materials and Methods
2.1. Plasmids Construction (Coding for FXN Gene and Anti-FXN shRNA)
Adeno-associated viruses, serotype PHP.B (AAV-PHP.B) [
30], were used to deliver either a plasmid coding for anti-FXN shRNA to increase the FRDA phenotype in YG8sR mice or the frataxin gene as a treatment in YG8sR mice. This specific serotype is known to drive expression in the central nervous system with an ability to cross the blood-brain barrier (BBB).
The previously described scAAV plasmid [
15] was modified to change the CAG promoter for the chicken β-actin hybrid (CBh) promoter to express the human frataxin gene ubiquitously (scAAV-CBh-FXN). To synthetize the scAAV-hSyn-FXN, a specific plasmid to drive expression in the central nervous system, the scAAV-CBh-FXN plasmid was digested with AvrII and AgeI to remove the CBh promoter. The human synapsin 1 (hSyn) promoter was excised from plasmid AAV synap FZ (Addgene #60231) by XbaI and AgeI and ligated into the plasmid to create scAAV-hSyn-FXN. Both constructions were used to compare the efficiency of both promoters to deliver the FXN gene as a potential treatment.
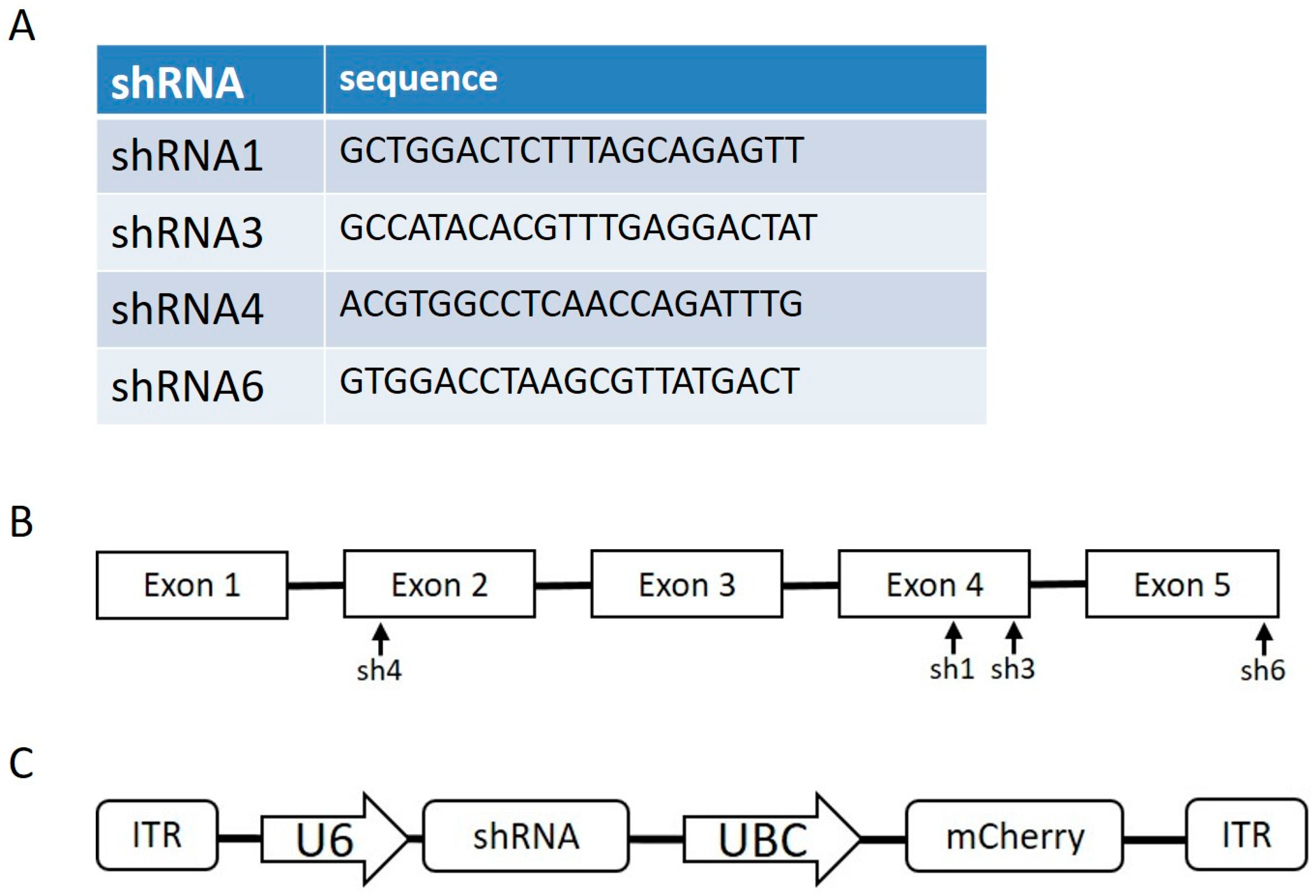
For the construction of anti-FXN shRNA plasmids to increase the YG8sR phenotype, scAAV-hSyn-FXN was digested with BamHI and MluI, and U6-shRE and UBC-mCherry were ligated to the plasmid using NEBuilders. The resulting plasmid was digested with AgeI and NheI, and shFXN oligonucleotides were annealed and ligated to the plasmid to create scAAV-shRNAFXN (1, 3, 4 or 6)-UBC-mCherry plasmids. The shRNA sequences were derived from predesigned Sigma sequences: TRCN0000006138 (shRNA1), TRCN0000380594 (shRNA3), TRCN0000010996 (shRNA4), TRCN0000350511 (shRNA6). The sequences and sites targeted by the various shRNAs are illustrated in
Figure 1A,B. The AAV plasmid containing the shRNA sequence is illustrated in
Figure 1C.
2.2. In Vitro Testing of the shRNAs
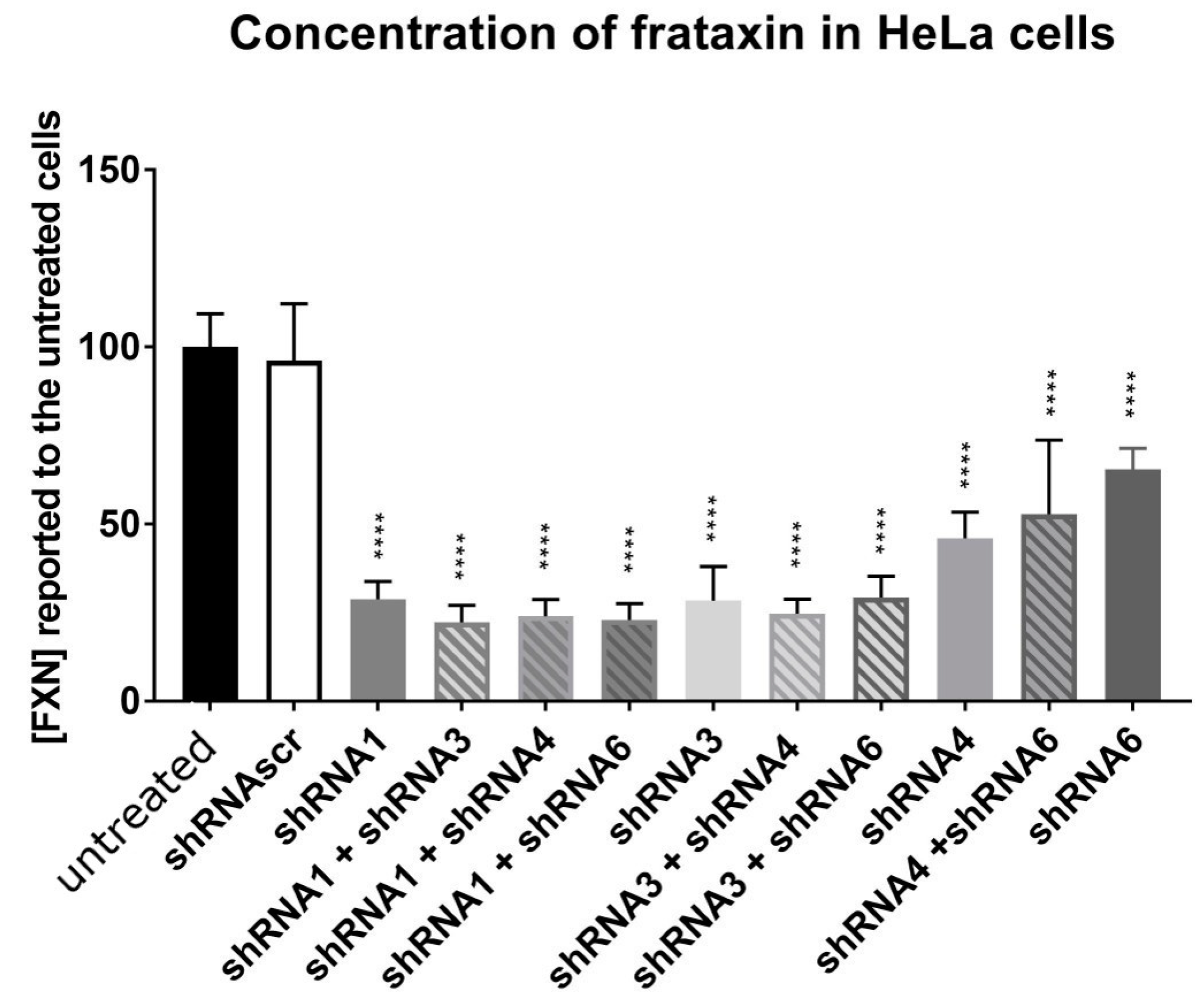
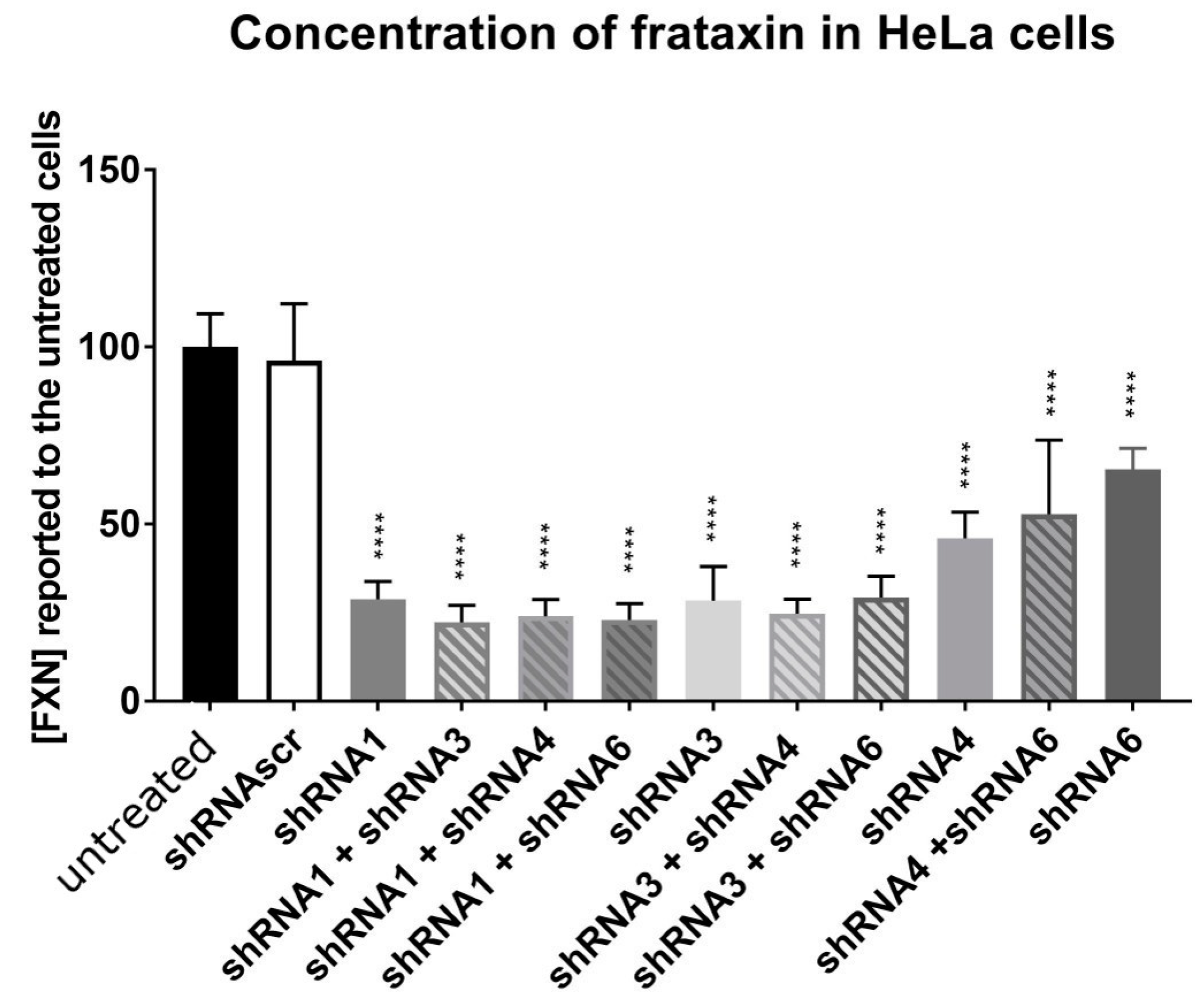
HeLa cells were transfected using Lipofectamine 2000 with the different plasmids coding for a shRNA and mCherry (scAAV-shFXN-mCherry) alone or in combination for a total of 2 µg of plasmids. The fluorescence of mCherry was used to assess the transfection success. The cells were collected after 72 h and the proteins were extracted. The frataxin protein concentration was measured by an ELISA test (Ab109881, Abcam, Cambridge, UK). These in vitro tests allowed us to select the best shRNA likely to reduce frataxin in vivo.
2.3. AAV Production
For in vivo testing, different promotors were used to control the expression of the shRNA and the FXN gene. The shRNA was placed under a U6 promotor and the mCherry gene under a UBC (
Figure 1C). In the AAV coding for the frataxin gene, two promoters were tested and compared, hSyn for specific delivery to the nervous system and CBh for a ubiquitous delivery.
Adeno-associated viruses, serotype PHP.B (AAV-PHP.B) [
30], were used to deliver the shRNA or the frataxin gene in vivo in YG8sR mice. The AAV-PHP.B vectors were produced by the Plateforme d’outils moléculaires, Centre de recherche CERVO (Québec, QC, Canada).
2.4. AAV Injection
Mice (approximately 2 months old) were injected in the tail vein (IV) with 0.9% saline solution, with an AAV coding for shRNAscr (scramble shRNA sequence, not targeting frataxin) for control mice or with an AAV coding for anti-FXN shRNA. Different doses of AAVs were used from 0.5× (i.e., 3 × 1011 viral copies) to 4× (i.e., 2.4 × 1012 viral copies) depending on the effect on the mice. To treat the 2 months old mice, an AAV coding for human frataxin (FXN) gene was administrated alone or mixed with the AAV coding for a shRNA.
2.5. YG8sR, YG8-800 and Y47R Mice
The YG8sR, YG8-800, and Y47R mice were obtained from Jackson Laboratories Inc and reproduced in our facilities. To summarize, YG8sR (Fxntm1Mkn Tg(FXN)YG8Pook/2J;
https://www.jax.org/strain/024113) (accessed on 10 June 2023) mice, YG8-800 (Fxnem2.1Lutzy Tg(FXN)YG8Pook/800J;
https://www.jax.org/strain/030395) (accessed on 10 June 2023) mice and Y47R (Fxntm1Mkn Tg(FXN)Y47Pook/J;
https://www.jax.org/strain/024097) (accessed on 10 June 2023) mice [
29] all have mouse FXN genes knockout but they have, respectively, a transgene containing the human FXN gene with a 250–300 GAA triplet, an 800 GAA triplet, or a 9 GAA triplet. C57Bl6J (subsequently called C57) mice were used as control mice, containing no human transgene. The C57 mice group was made of 15 mice (9 females and 6 males), the Y47R group of 21 mice (13 females and 8 males), the YG8sR group of 19 mice (11 females and 8 males), and the YG8-800 group of 15 mice (7 males and 8 females) initially at 2 months age. The housing conditions were 2–5 mice per standard ventilated cage under a 12-h light/dark cycle with access to food and water ad libitum. Neslet and Aspen’s shaving are given for nidification. All experiments involving animals were approved by the Comité de Protection des Animaux de l’Université Laval (CPAUL3) (Québec, QC, Canada).
The first mice group injected with an AAV were euthanized at 3.5 months after the behavior tests, 5 weeks after the IV injection, and perfused with 0.9% saline to remove the blood from tissues. Different tissues (Tibialis anterior muscle, heart, liver, cerebrum, cerebellum, and dorsal root ganglions (DRG)) were then collected and stored at −80 °C. The subsequent group made of 15 C57 mice (9 females and 6 males), 21 Y47R mice (13 females and 8 males), 19 YG8sR mice (11 females and 8 males), and 15 YG8-800 mice (7 males and 8 females) were euthanized after 11 to 14 months with behavior tests every 3 weeks starting at 8 weeks age. The same tissues were also collected and stored at −80 °C. The euthanasia method was cardiac perfusion under isoflurane anesthesia.
2.6. Behavior Tests
The physical performances of YG8sR and YG8-800 mice were compared with those of Y47R mice and also with those of C57Bl6J (C57) mice, which are control mice with their natural mouse frataxin genes. All mice were evaluated every 3 months starting at 2 months and up to 11 months. For statistics, the females and males were pooled together because there were no significant differences in the preliminary analysis. The C57 mice group was made of 15 mice (9 females and 6 males), the Y47R group of 21 mice (13 females and 8 males), the YG8sR group of 19 mice (11 females and 8 males), and the YG8-800 group of 15 mice (7 males and 8 females).
2.6.1. Parallel Rod Floor
The mice were placed in the parallel rod floor apparatus [
31] for 8 min and followed with a camera. The apparatus is a plexiglass cage with metallic parallel rods 1 cm above the metallic plate floor. The apparatus detected foot faults when the foot passed through the grid and touched the metallic plate at the bottom. Different parameters (i.e., the distance traveled, the average speed, the time spent immobile, and the foot fault number) were recorded with the ANY-maze software version 7.20 connected to the camera and floor foot detector.
2.6.2. Hanging Test
The mice were weighed before the test. This is a four-limbs hanging test, the mouse grasped a wire grid as it is inverted. The test was based on the Kondziela’s inverted screen test [
32,
33]. The time of sustained limb tension to oppose the mouse weight was measured. The chronometer was started as soon as the grid was inverted over the cage. The time the mouse remained hanging (before the mouse fell) was noted, with a maximum of 300 s for the test. Three sets of hanging tests were conducted with a minimum of 2 min between each test. The longest time the mouse remained hanging on the screen was noted.
2.6.3. Notched Beam Test
Our test was based on the Di Bonito [
34] notched beam test. We used a one-meter Plexiglass beam engraved with 18 squares of 2 cm wide/deep/long and spaced every 2 cm. The locomotion was assessed by the time required for the mouse to cross the beam over the squares. We also counted the foot faults manually with 2 observers (i.e., each time the foot touched the bottom of the square). The experiment was made in triplicates and the average number of foot faults was used.
2.6.4. Inverted T Beam/Balance Beam Test
This test used a Plexiglass balance beam, which was 1 m long and 2 cm wide with a central platform of 0.66 cm wide and 0.66 cm high based on the balance beam test of Luong and co-workers [
35]. The mouse walks on the 0.66 cm top beam and steps on the bottom parts of the platform (inverted T shape) when making a foot fault. The time to cross the beam was also recorded as well as every foot fault (mouse foot slipped off the beam), which were manually counted by 2 people. The experiment was made in triplicates and the average number of foot faults was used.
2.7. Imaging of mCherry Fluorescence in Mouse Organs
The integrated imaging station IVIS Lumina LT Series III (PerkinElmer, Waltham, USA) was used to evaluate the distribution of mCherry protein in macroscopic organs using its intrinsic orange-red fluorescence. The apparatus was set in fluorescence mode with filters at 535 nm (excitation) and 620 nm (emission), small binning, and 1–30 s range of exposure time. Under the dedicated Living Image software 4.5.4, the fluorescent signal intensity was calibrated as radiant efficiency and normalized to allow intensity comparisons. No significant auto-fluorescent signal was detected. The imaging was achieved by the Plateforme Imagerie animale par bioluminescence/fluorescence, Centre de recherche CHU (Québec, QC, Canada).
2.8. DNA Extraction from Tissues
DNA was extracted from different tissues (muscle, liver, heart, brain, and dorsal root ganglions). Briefly, a part of the tissue was recovered and incubated with 50 mL proteinase K (10 mg/mL) in a lysis buffer (1.82 g TrisBase, 5 g SDS, 10 mL EDTA 0.5 M completed with distilled water to 500 mL, pH 8.0) at 56 °C until the solution became clear. Digested tissues were then mixed with a 500 µL solution of phenol/chloroform/isoamyl alcohol (25:24:1; BioShop Canada Inc., Burlington, ON, Canada) and centrifuged for 3 min at 12,000 rpm. The upper solution was recovered and mixed with the same volume of chloroform and centrifuged again. The upper solution was recovered, and 50 µL of 5 M sodium chloride was added before the addition of 1 mL 100% ethanol. After centrifugation for 8 min at 12,000 rpm, the pellets were washed in 70% alcohol before another centrifugation. Pellets were dried before DNA suspension in sterile water. The detection of the AAV content was accomplished by PCR, with a band of 410 pb and 435 pb respectively for the plasmid delivered by AAV expressing the shRNA (AAV-shRNA) or the frataxin gene (AAV-FXN) (
Supplementary Figure S1). The shRNA plasmid contained a 410 pb mCherry section (
Figure 1C) and the frataxin gene DNA amplified fragment was 435 pb.
2.9. Quantification of the Viral Particles by qPCR
The viral particle concentration in organs was measured by qPCR using primers that target the mCherry sequence present with the shRNA sequence in the AAV. 50 ng of the previously extracted gDNA was used per well with 6 µL Advanced Universal SYBR Green supermix, 0.5 µL 10 mM Forward mCherry primer, 0.5 µL 10 mM Reverse mCherry primer and completed to 10 µL with DNase free water. Bio-Rad CFX96 was set at SYBR only with a 3-min 98 °C polymerase activation and DNA denaturation phase, then 40 cycles of 10 s 98 °C denaturation and 20 s 60 °C annealing and extension with a melt curve analysis from 65 °C to 95 °C increasing the temperature by 0.5 °C at every 5 s. The standard curve was made with 8 samples from 101 to 108 particles. Quantified organs were TA muscle, liver, heart, cerebrum, cerebellum, and DRG. All samples were duplicated. This qPCR used primers in the mCherry and in the HPRT reference gene (
Supplementary Table S1).
2.10. Quantification of mRNA Expression by qRT-PCR
Frataxin mRNA expression in tissues was compared by qRT-PCR. The RNA was first extracted by adding 1 mL of Trizol to the finely crushed tissue slice. A total of 200 µL chloroform was then added to separate the phases during 15 min centrifugation at 12,000 rpm. The supernatant was collected and precipitated in 500 µL cold (−20 °C) isopropanol centrifuged at 12,000 rpm for 10 min. The liquid was then removed from the tube to keep only the pellet to be washed twice with 1 mL 75% cold (−20 °C) ethanol and centrifuged at 10,000 rpm for 5 min. The pellet was resuspended in 20–60 µL RNAse free water. The RNA concentration was measured with a spectrophotometer and the migration on agarose gel 1% permitted to verify that the RNA was not degraded. The sample was then treated with DNAse (NEB Inc.) incubated for 10 min at 37 °C, then the DNAse was deactivated with EDTA 2.5 mM at 75 °C for 10 min. The RNA was then put in the presence of Reverse Transcriptase with dNTPs and random primers in the buffer provided by Thermo Fischer Scientific Inc. (Waltham, MA, USA) in the ‘High Capacity cDNA Reverse Transcription kit’. A qPCR was then performed with the complementary DNA obtained. This qRT-PCR used primers for frataxin as well as for HPRT, a reference gene (
Supplementary Table S1).
2.11. Frataxin Protein Quantification
The proteins were extracted using 300 µL of the protein extraction buffer provided by AbCam in the Human frataxin Elisa kit (ab176112, Abcam, Cambridge, MA, USA) per tissue slice (0.5 mm). The frataxin protein concentration was estimated using the PierceTM BCA Protein Assay Kit (Thermo Fischer Scientific Inc.). This kit uses a bovine serum albumin (BSA) standard of 0 mg/mL, 0.25 mg/mL, 0.625 mg/mL, 1.25 mg/mL, 1.875 mg/mL, 2.5 mg/mL and 5 mg/mL. All samples were conducted in duplicate and their optical density (OD) at 562 nm was compared to the standard curve to estimate their protein concentration.
The human frataxin protein was quantified using the Human frataxin Elisa kit (ab176112, Abcam, Cambridge, MA, USA) for in vitro and for mouse tissues. A standard curve was also made at the same time with recombinant human frataxin protein to quantify the frataxin protein in the tissues in nanogram/microgram total protein. All samples were carried out in duplicate and compared to the standard curve using their OD at 450 nm.
2.12. Statistical Analysis
The results are presented as mean ± SD. Statistical analyses were performed using GraphPad Prism7 software version 9.0 (GraphPad Inc., La Jolla, CA, USA) and detailed in each figure legend. The results were analyzed in two ways ANOVA test Tukey’s multiple comparisons test (* p value ˂ 0.05, ** p value ˂ 0.005, *** p value ˂ 0.0002 and **** ˂ 0.0001) for behavior tests. For qPCR the statistics were conducted by the software for the qPCR Biorad CFX Maestro. The p values are indicated in each figure. Statistics: one-way ANOVA was used at 0.05 (95% confidence interval); * p < 0.05, ** p < 0.003, *** p < 0.0003, and **** p < 0.0001.
4. Discussion
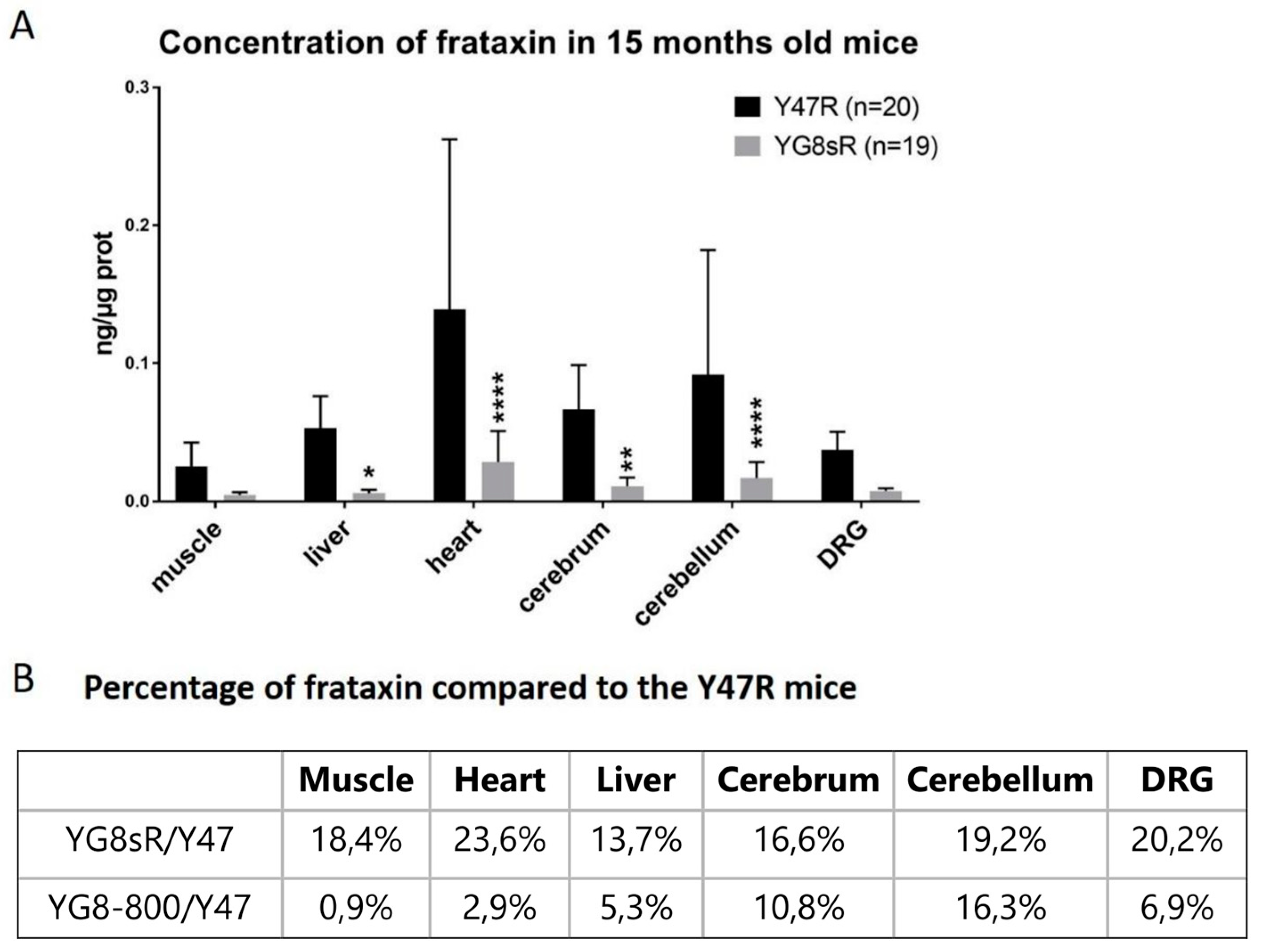
Initially, Virmouni and co-workers developed a new mouse model containing a human FNX transgene with about 200 GAA repeats, here called YG8sR(200) for an easier discussion [
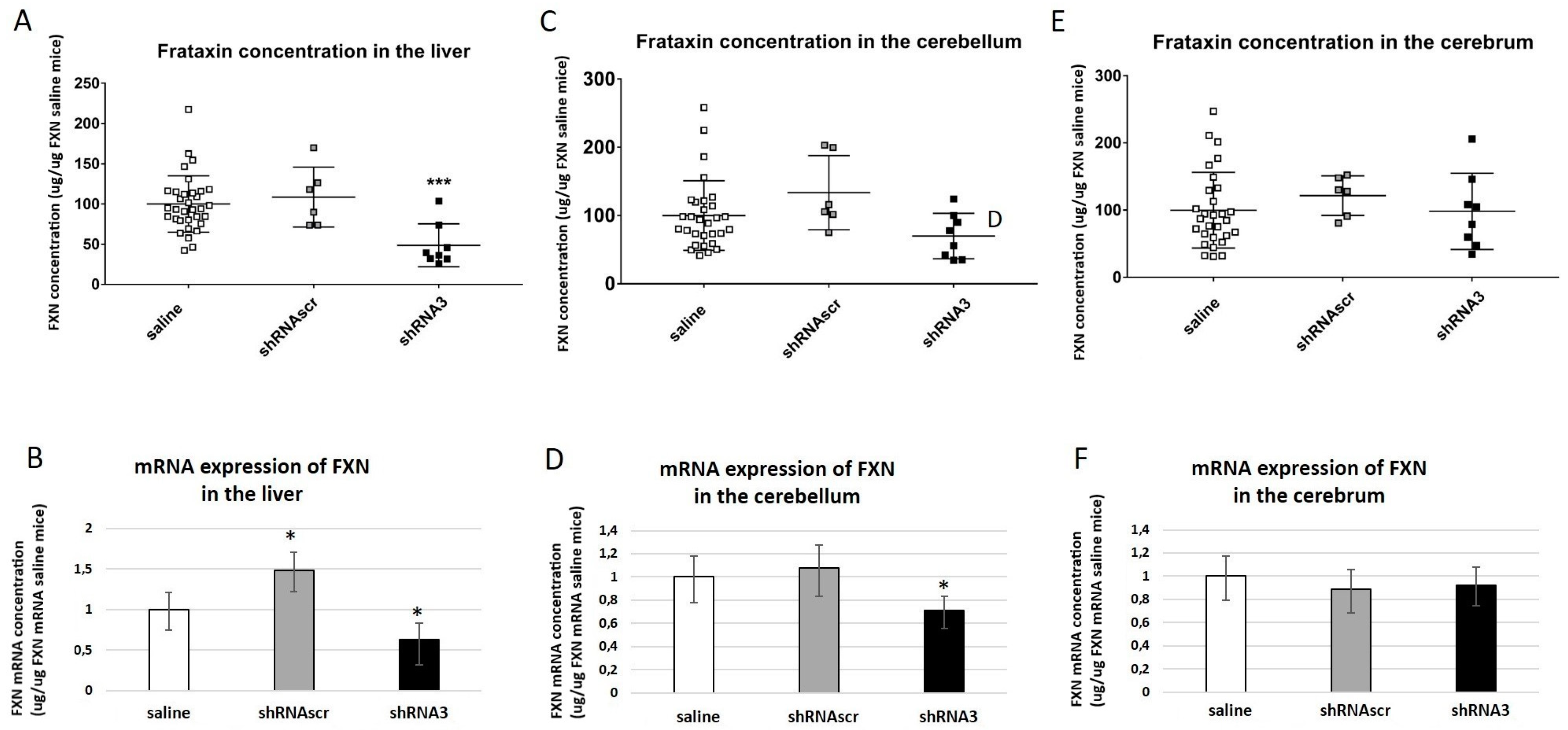
29]. Even if the behavior of these YG8sR(200) mice on the rotarod was significantly different from C57 and Y47R mice, by pooling males and females together, the differences were not as clear as the results obtained when the sexes were analyzed separately. Moreover, the YG8sR(200) mice showed reduced locomotion activity over time but the most significant differences were observed with more specific tests such as the beam tests, the hang-wire test, and the grip test at 12 months. Indeed, at that age, the YG8sR(200) mice took more time to cross the two beams, fell faster on the hang-wire test, and showed less grip strength compared to the Y47R or C57 mice. The human frataxin mRNA, as well as the human frataxin protein concentrations in YG8sR(200) mice, were also decreased in the brain, the cerebellum, and the liver at 12 months compared to Y47R and C57 mice [
29]. We have purchased the YG8sR mice from Jackson Laboratories Inc. and those that we have used contained between 250 and 300 GAA repeats (
https://www.jax.org/strain/024113) (accessed on 10 June 2023). In our studies, the YG8sR females performed better than the YG8sR males in the behavior tests. Nevertheless, no great difference was observed compared to Y47R mice in these tests. The concentration of human frataxin protein was reduced in YG8sR in all tissues tested compared to Y47R and even more than in the YG8sR(200) for the cerebellum (i.e., 70% in the YG8sR(200) and 19% in YG8sR). Considering that YG8sR mice have a human frataxin transgene containing 250–300 GAA repeats responsible for low human frataxin expression, these mice remain a good model to study the effects of frataxin reduction [
36], or to try to increase the frataxin expression by some treatments [
37,
38] or even through genetic modifications using the CRISPR-Cas9 system to delete the GAA repeat [
19,
20]. All these studies are essential to understanding the mechanism and interaction between the frataxin protein and other proteins or enzymes but at some point, it becomes essential to have a good mouse model that recreates the symptoms of Friedreich ataxia and allows for the detection of functional improvements with treatment.
The use of shRNAs is a method used to decrease the expression of a protein and can be tissue-specific by using a specific promoter. For example, by using a member of the cadherin family that is exclusively expressed in the renal tubular epithelial cells, i.e., Ksp-cadherin, Xu et al. [
39] observed 2 weeks after IV injection of an AAV9-shRNA-ALDH2, a decrease of the aldehyde dehydrogenase 2 protein (ALDH2) only in the kidney although this protein is also expressed in the liver and the heart [
39]. Chandran and co-workers in 2017 [
27] used shRNAs to decrease the mouse frataxin and create a mouse model for FRDA called FRDAkd. In their study, different shRNAs against frataxin were tested and they decided to use one, which induced the strongest reduction of frataxin in vitro. Among the different shRNAs that they tested, the one decreasing the most the frataxin expression was also targeting exon 4, as in our case for our sh1 and sh3 [
27].
Piras et al., in 2013 suggested that gene knockdown is generally less predictable than gene over-expression [
40]. In our study, the concentration of viral particles in the brain (cerebrum and cerebellum) was as high as in the liver but the frataxin concentration was significantly reduced only in the liver. In our case, the AAV serotype PHP.B was used. In the first study on AAV-PHP.B, the bio-distribution after IV injection in mice was compared to AAV9 and was similar between the two AAVs in the liver, the heart, and the skeletal muscle but the expression of GFP was lower with the AAV-PHP.B than with AAV9, even in the liver, where viral particles were as high as in the nervous system [
30].
Chandran and co-workers also created a new mouse model for Friedreich Ataxia. They incorporated in a defined genomic locus, a single copy of a doxycycline-inducible anti-FXN shRNA to silence frataxin. Different behavior tests were carried out at 12 and 24 weeks of age with doxycycline treatment and they observed neurological deficits. After 20 weeks of age, the mice showed a drastic reduction of frataxin protein in the eight organs tested (liver, heart, kidney, brain, muscle, pancreas, lung and spinal cord) [
27]. In our experiments, the effect of shRNA was faster and the frataxin mRNAs and protein concentrations were estimated only 5 weeks after the AAV-shRNA injection. However, after 5 weeks, a reduction of frataxin protein was observed in the liver but not in the other organs. Nevertheless, the frataxin mRNA was significantly reduced in the liver but also in the cerebellum. It is probable that 5 weeks is too short to observe an effect on the frataxin concentration on other organs than the liver because in the Chandran et al. study, the investigators showed a decrease of the frataxin protein concentration only for the liver in a time series starting at 3 weeks of age, but in other organs after 20 weeks [
27].
The important question is whether we can reverse a phenotype even in patients who have lived several years with the disease by increasing frataxin expression. Many researchers tried to answer this question. Chandran et al. [
27] with their inducible mouse model FRDAkd were able to stop the treatment with the doxycycline to increase again the expression of frataxin. After 13 weeks of doxycycline treatment followed by 12 weeks without this treatment, the mice recovered a healthy phenotype close to control mice [
27]. Therefore, it appears that the coordination impairment is reversible when frataxin is increased. Another method to increase the concentration of frataxin in mice is to inject an AAV coding for the frataxin gene. We have worked on the delivery of the frataxin gene by AAVs since 2014 [
15]. We initially used an AAV9 for the delivery in NSE-cre and MCK-cre mice, i.e., the mouse models created by Puccio H. in 2001 [
22]. We found a great improvement in heart function and behavior tests by intraperitoneal administration of AAV9-FXN at an early age, between 5 and 9 days of age [
15]. In the present study, our goal was to focus on the neuronal dysfunction, therefore, the AAV-PHP.B was chosen to target the nervous system with the shRNA. A co-injection (AAV-shRNA and AAV-FXN) was necessary to avoid a hyperacute immune response that could be provoked by a second AAV injection [
41]. In our study, the co-injection of AAV-PHP.B-CBh-FXN with an AAV-PHP.B-antiFXNshRNA improved the behavior of mice compared to those treated with the AAV-PHP.B shRNA against frataxin alone. Puccio and co-workers created in 2018 a mouse model to specifically target cells expressing parvalbumin, called parvalbumin conditional knockout mouse model (Pvalb cKO mouse) [
16]. The parvalbumin promoter is specific to proprioceptive neurons. These mice reflected the FRDA neuropathophysiology. They injected intravenously into 3.5 weeks old mice an AAV9-CAG-FXN and obtained a significant coordination improvement in the notched beam but not reaching the phenotype of their control mice (Fxn+/L3). Moreover, in the wire-hanging test, the treated Pvalb cKO mice were similar to the WT. Our aim was to have a comparable coordination impairment to this model in a mouse model containing the human gene with an expanded GAA to evaluate the impact of a genetic treatment on the coordination of mice. We used AAV. PHP.B to cross the BBB, but tissue analysis revealed not only the efficiency of the AAV9 on the sensory neuropathy but also a non-homogenous distribution of the AAVrh10 in the cerebellum leading to a partial rescue of Purkinje cells [
16]. AAV8 was also tested in the FRDAkd mouse model (described above) in 2022 [
42]. Ten weeks after dox treatment the mice received an intravenous injection of AAV8 coding for frataxin. Unfortunately, restoration of frataxin with this AAV failed to improve the behavior, metabolic and histological features of this mouse model [
42].
The different articles using an AAV coding for frataxin were often using a strong promoter such as CMV or CBh which led to overexpression of frataxin in different organs. A recent review by Sivakumar and Cherqui in 2022 highlighted the controversial effects of frataxin overexpression [
43]. In the past, in our studies, even if we obtained overexpression of frataxin in the tissues of the mouse models (and especially in the liver), we never found clear evidence that overexpression was harmful to the mice. This may depend on the frataxin concentration which was obtained. Indeed, Puccio’s group published in 2020 [
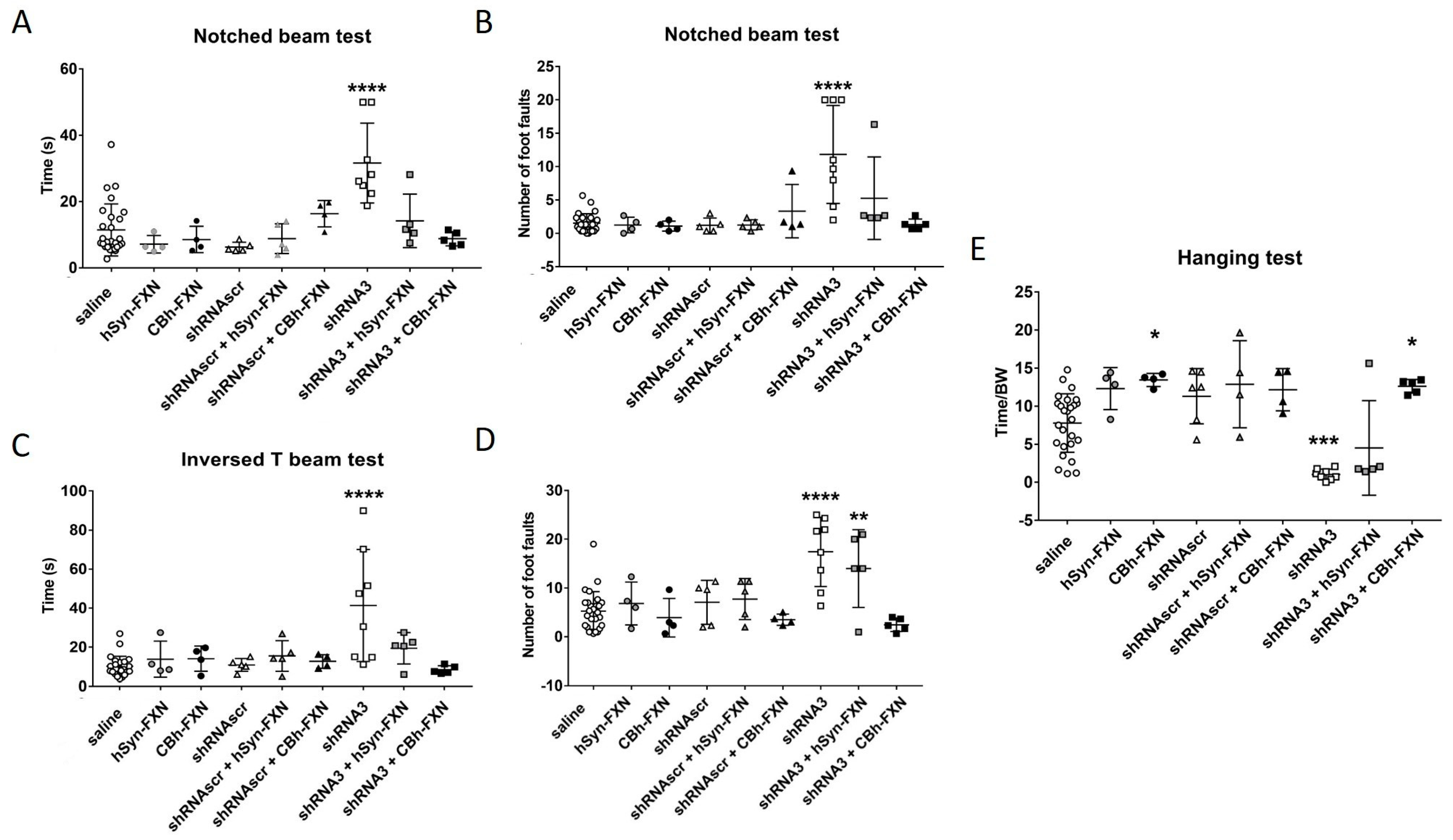
44] a study showing that a dose-dependent harmful effect was observed in mouse heart function started after a 20-fold increase in frataxin concentration. However, the harmful effect was tissue-specific since a 90-fold increase of frataxin in the liver of mice, did not induce toxicity. In the present study, we tested two promoters, a general and strong (CBh) one and a weak one more specific for the nervous system (hSyn). The frataxin expression under a CBh promoter showed at 5 weeks post injection a significant improvement in the behavior tests compared to the hSyn promoter. Nevertheless, the period of testing was just 5 weeks post-injection and it is possible that this follow-up period was too short to observe an improvement in the behavior with the hSyn promoter. Moreover, the hSyn promoter, due to its specificity, did not induce any increase of frataxin in the liver even if the number of viral particles was high.
It is important to find an appropriate mouse model of FRDA for what is being tested, e.g., to measure the impact of treatment on the behavioral phenotype. Mercado-Ayón et al., (2022) [
42], demonstrated that the symptoms obtained in their FRDAkd mouse model are not only the result of the reduction of frataxin expression but that the doxorubicin treatment has also an impact on the symptoms. In our study, the shRNA was provided by an AAV and no additional treatment was necessary. The only disadvantage of our approach is that we had to co-inject the treatment and the shRNA because the same AAV serotype was used for both and thus a delayed administration of the AAV was not possible because of the immune response induced by the first administration.
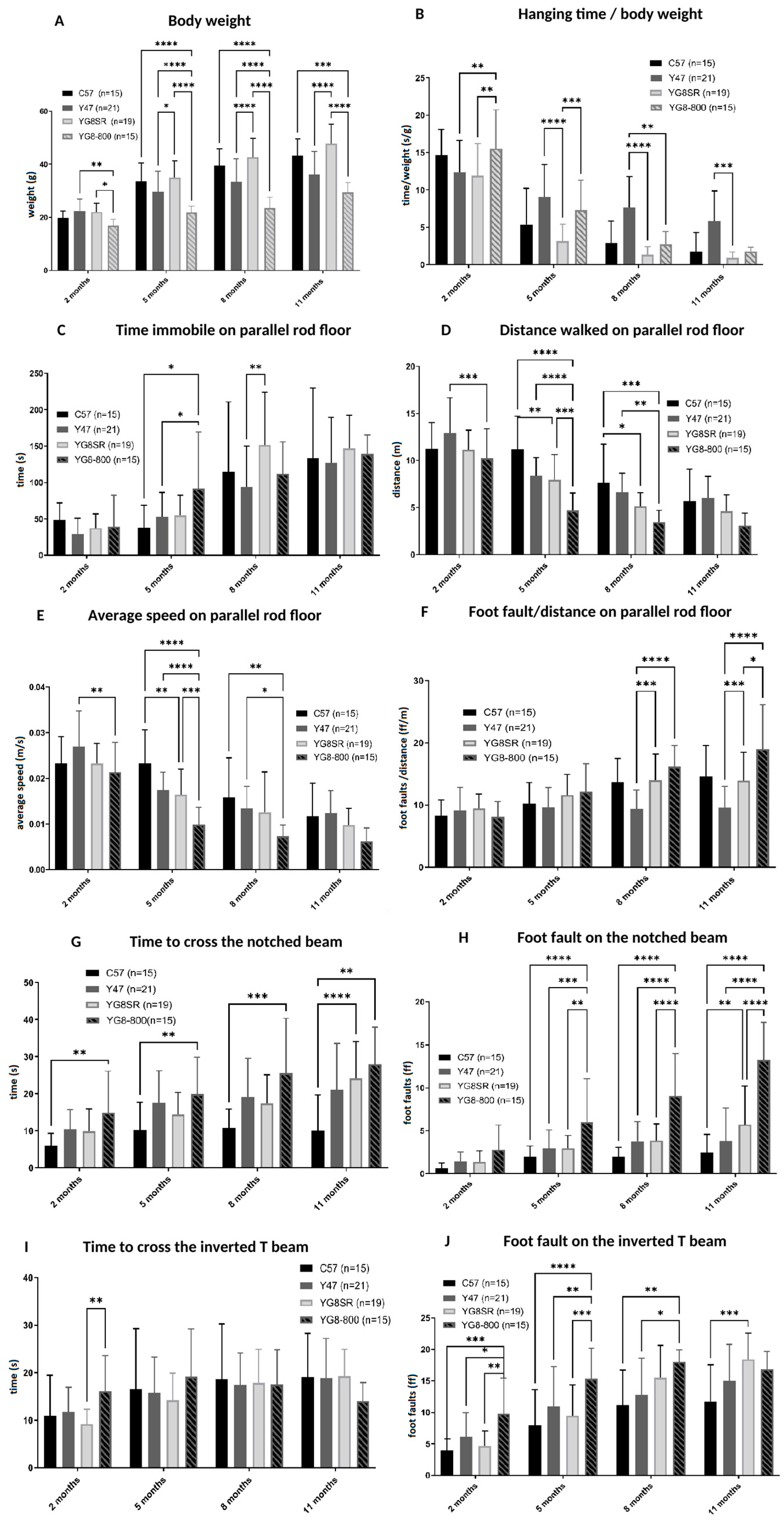
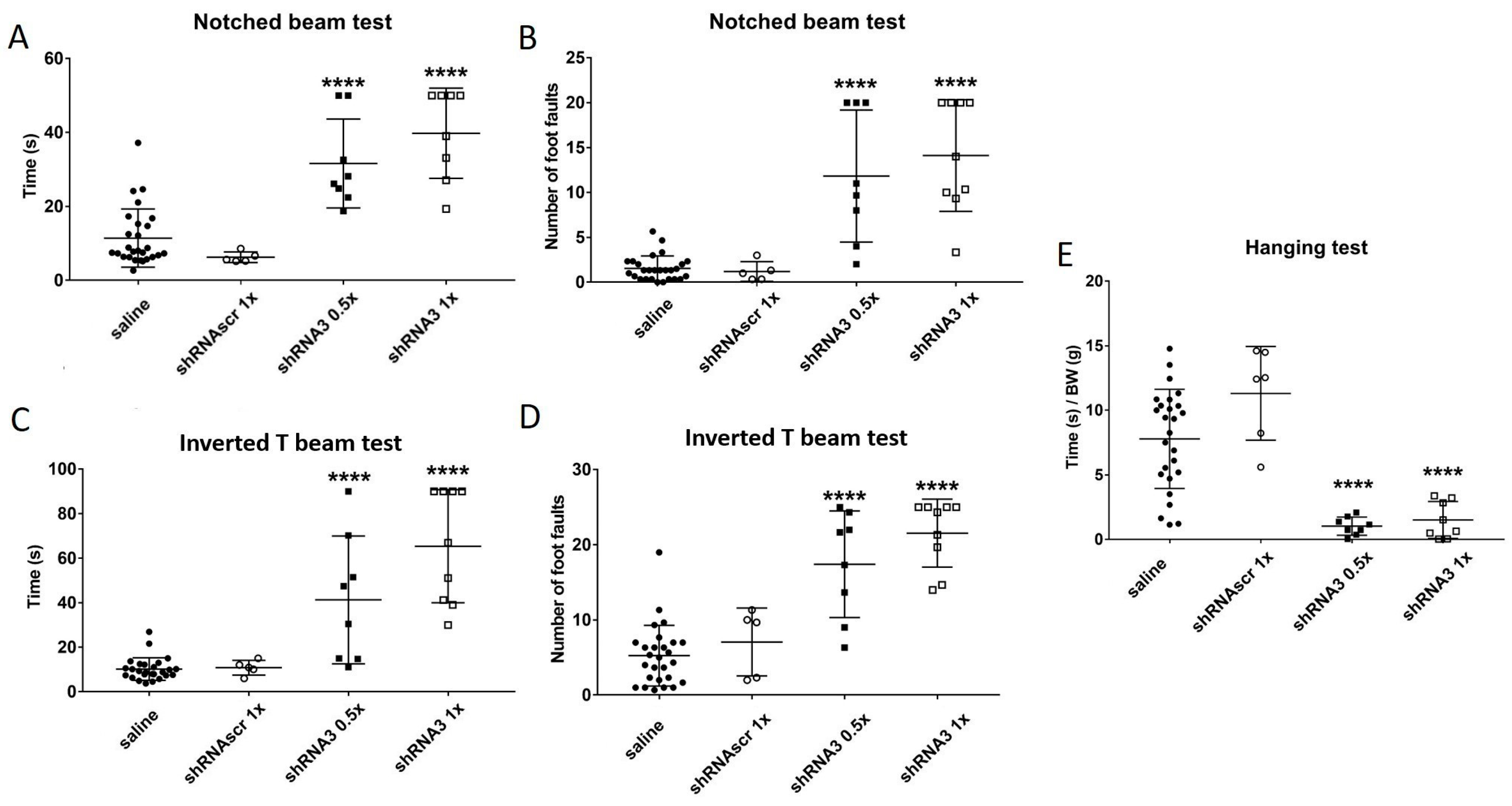
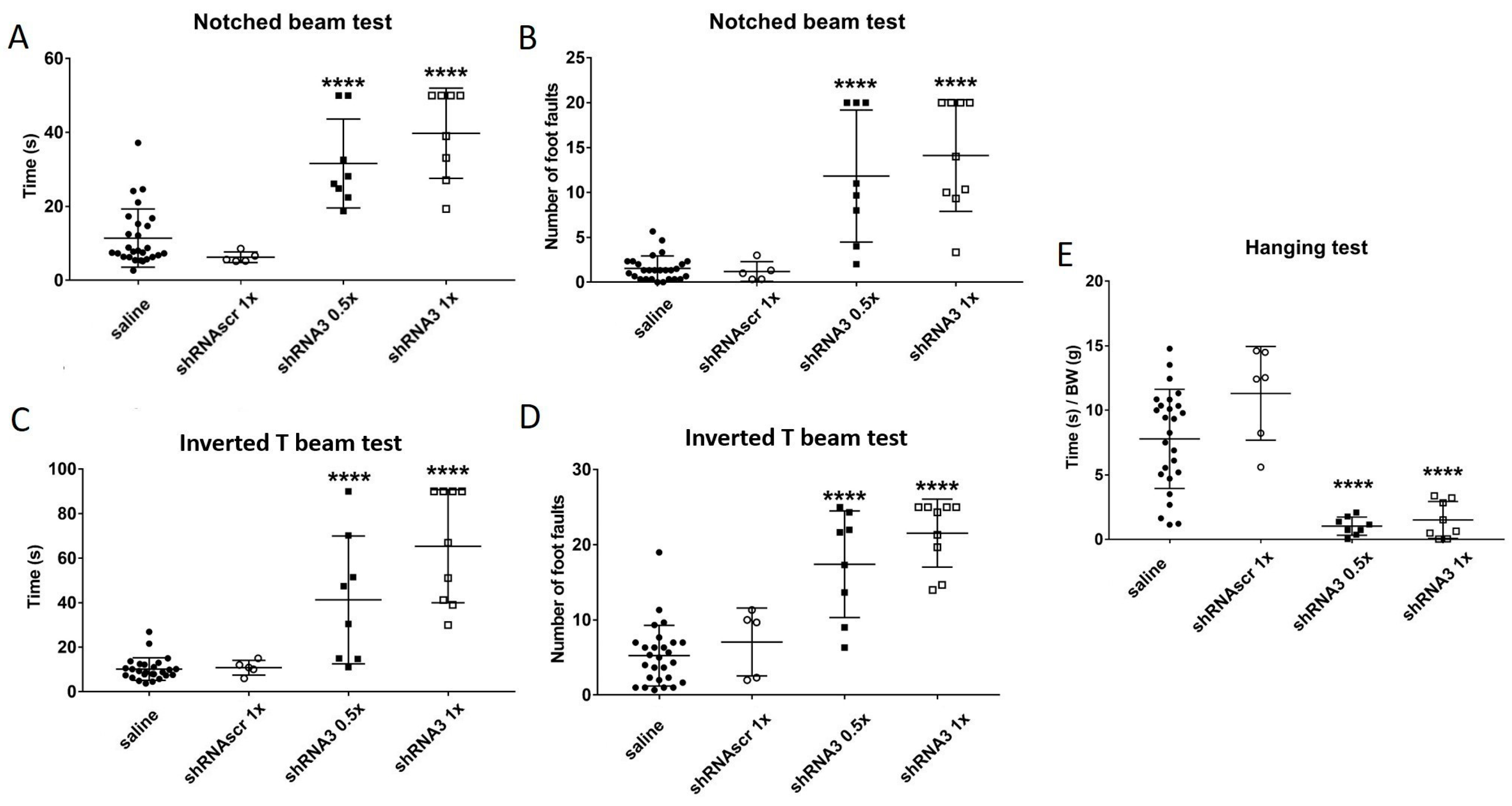
The aim of this study was to decrease the frataxin in YG8sR mice to increase the severity of their phenotype. Therefore, both YG8sR injected with an shRNA and YG8-800 mice phenotypes were compared. The results have shown that shRNA targeting frataxin does decrease its expression and increases the ataxic phenotype of YG8sR mice. The question was if this method is now the optimal one knowing that the YG8-800 mouse model is available. The complete characterization of this model has been carried out in our laboratory [
45] showing that the phenotype of the YG8-800 mice is comparable to the human patients since the movement coordination progressively decreases and a cardiac hypertrophy is noticed. When compared, the YG8sR mice injected with the shRNA3 (
Figure 5) have a phenotype comparable to the YG8-800 mice (
Figure 2G–J). In fact, at 3–4 months, the YG8sR +shRNA3 need an average of 35 s to cross the notched beam and make 12-foot faults, which is slightly more severe than the YG8-800 mice 20 s and 7-foot faults at 5 months (saline control needing in average 10 s and making around 2.5-foot faults). The same trend is noticed for the inverted T beam, on which the shRNA-treated mice need on average 57 s and make 19-foot faults, whereas YG8-800 mice need 18 s and make 14-foot faults (saline needs 9 s and make 6-foot faults). In conclusion, both YG8sR injected with shRNA3 and YG8-800 offer a phenotype that can be compared to human patients. The YG8-800 mouse model is recent and has only a few articles about it. In September 2022, our team published the characterization of this mouse model including the cardiac hypertrophy which is also part of the human phenotype [
45]. In January 2023, Kalef-Ezra et al. confirmed the impaired coordination and reduced frataxin levels of the mouse model [
46].
We also studied the possibility of using AAV.PHP.B to deliver the frataxin gene. Our results on YG8sR mice were promising since they reestablished the coordination and more tests will be conducted in YG8-800 mice without needing to co-inject with an anti-FXN shRNA.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}