
Integrated Transcriptomics and Metabolomics Analysis of the Fructan Metabolism Response to Low-Temperature Stress in Garlic
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatment
2.2. RNA Extraction and cDNA Synthesis
2.3. Quantitative Real-Time PCR (qRT–PCR) Analysis
2.4. Construction and Detection of the cDNA Library
2.5. Original Data and Filtering and Transcript Splicing
2.6. Gene Function Annotation and CDS Prediction
2.7. Functional Annotation and Enrichment Analysis of Differentially Expressed Genes (DEGs)
2.8. Sample Preparation and Extraction
2.9. UPLC Conditions
2.10. ESI-Q TRAP-MS/MS
2.11. Screening and Functional Annotation of Differential Expression Metabolites
2.12. Integration of Transcriptome and Metabolome
2.13. Data Statistics and Analysis
3. Results
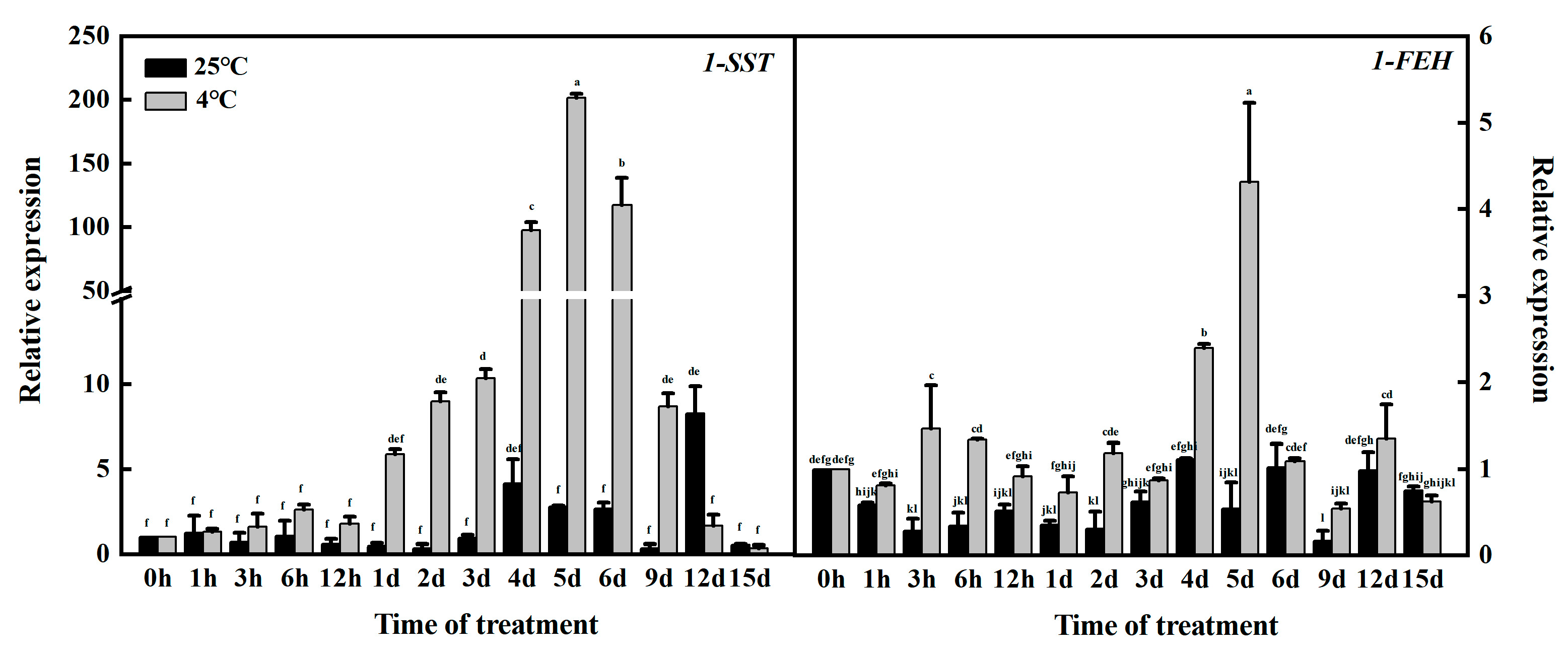
3.1. Selection of Key Periods of Garlic Fructan Gene Expression
3.2. Transcriptome Analysis Results
3.2.1. Sequencing and De Novo Assembly of the Garlic Transcriptome
3.2.2. Functional Annotation and Classification of Unigenes
3.2.3. Identification of DEGs
3.2.4. KEGG Enrichment Analysis of Differentially Expressed Carbohydrate Genes Produced by Low-Temperature Stress
3.2.5. qRT–PCR Verification
3.3. Metabolite Analysis Results
3.3.1. Detection of Metabolites and Screening of Differential Expression Metabolites
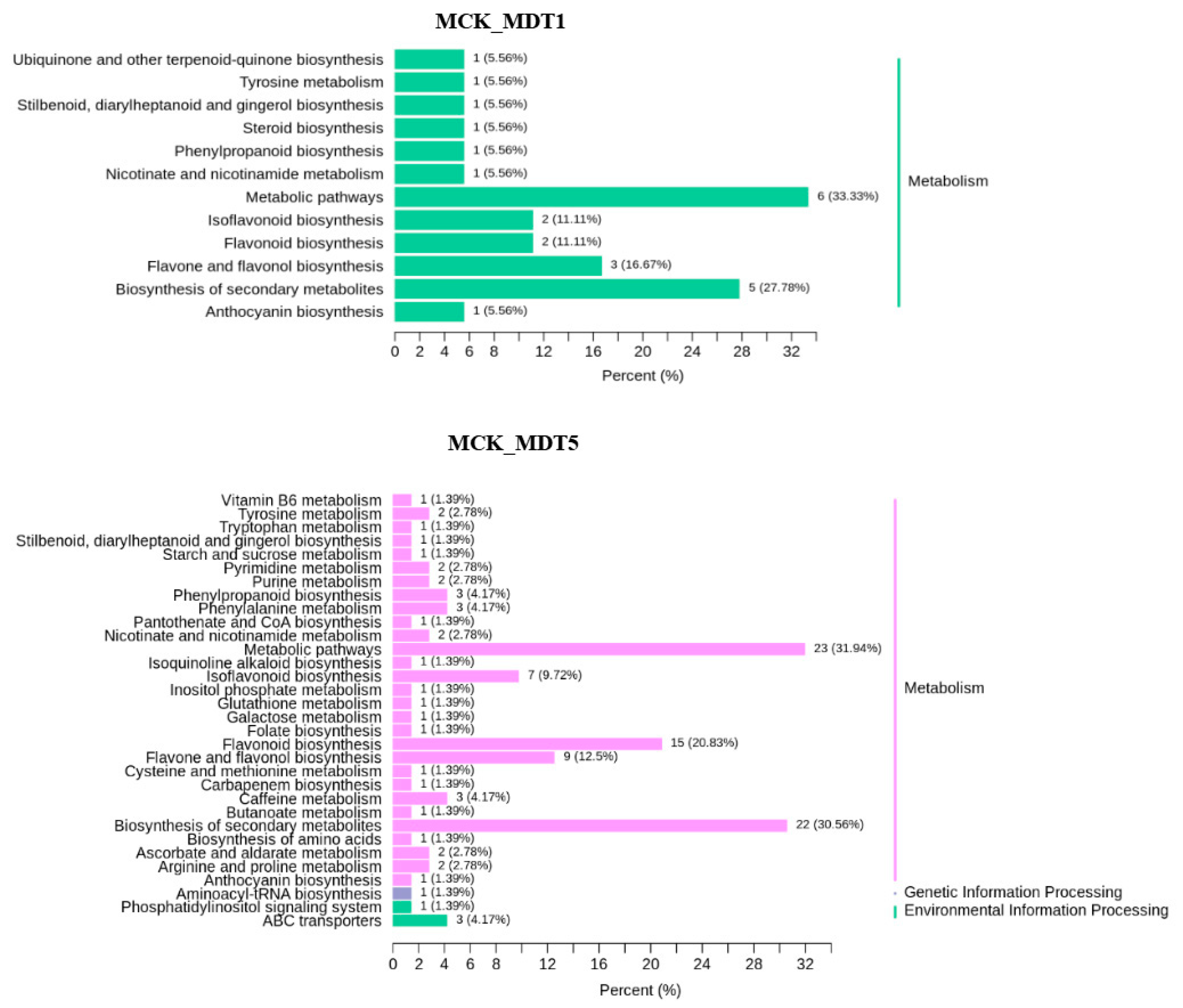
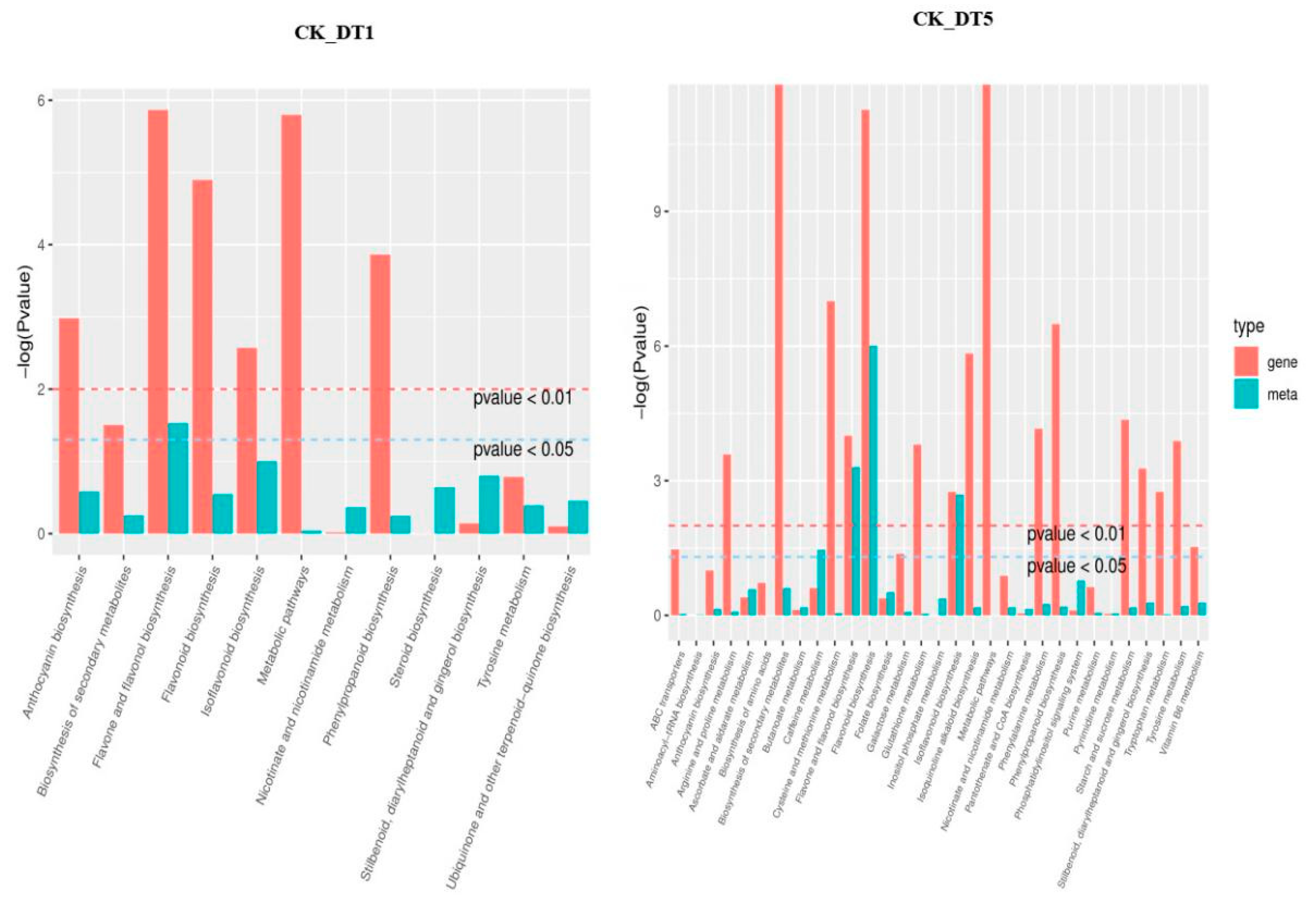
3.3.2. KEGG Enrichment Analysis of Differential Expression Metabolites under Low-Temperature Stress
3.3.3. Screening of Differential Expression Carbohydrate Metabolites
3.4. Correlation Analysis of the Transcriptiome and Metabolome
3.4.1. KEGG Enrichment Analysis of Differential Expression Genes and Differential Expression Metabolites
3.4.2. Weighted Gene Co-Expression Network Analysis
3.4.3. Correlation Analysis and Hub Genes Screening of Key Genes in Fructan Metabolism and Carbohydrate Metabolites
3.4.4. Analysis of Sugar Metabolism Pathway under Low-Temperature Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guevara-Figueroa, T.; López-Hernández, L.; Lopez, M.G.; Hurtado, M.D.D.; Vázquez-Barrios, M.E.; Guevara-Olvera, L.; González, R.G.G.; Rivera-Pastrana, D.M.; Torres-Robles, H.; Mercado-Silva, E.M. Conditioning garlic “seed” cloves at low temperature modifies plant growth, sugar, fructan content, and sucrose sucrose fructosyl transferase (1-SST) expression. Sci. Hortic. 2015, 189, 150–158. [Google Scholar] [CrossRef]
- Etoh, T.; Simon, P.W. Diversity, fertility and seed production of garlic. In Allium Crop Science: Recent Advances; CABI Publishing: Wallingford, UK, 2002. [Google Scholar]
- Ried, K.; Frank, O.R.; Stocks, N.P. Aged garlic extract reduces blood pressure in hypertensives: A dose-responsedose–response trial. Eur. J. Clin. Nutr. 2013, 67, 64–70. [Google Scholar] [CrossRef]
- Santhosha, S.G.; Jamuna, P.; Prabhavathi, S.N. Bioactive components of garlic and their physiological role in health maintenance: A review. Food Biosci. 2013, 3, 59–74. [Google Scholar] [CrossRef]
- Mathew, D.; Forer, Y.; Rabinowitch, H.D.; Kamenetsky, R. Effect of long photoperiod on the reproductive and bulbing processes in garlic (Allium sativum L.) genotypes. Environ. Exp. Bot. 2011, 71, 166–173. [Google Scholar] [CrossRef]
- Thomashow, F.M. Plant Cold Acclimation: Freezing Tolerance Genes and Regulatory Mechanisms. Annu. Rev. Plant Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Miao, Y.; Shao, L.; Hu, T.; Yang, P. De novo transcriptome sequencing and gene expression profiling of Elymus nutans under cold stress. BMC Genom. 2016, 17, 870. [Google Scholar] [CrossRef]
- Hirayama, T. Research on plant abiotic stress responses in the post-genomepostgenome era: Past, present and future. Plant J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef]
- Chen, S.; Huang, X.; Yan, X.; Liang, Y.; Wang, Y.; Li, X.; Peng, X.; Ma, X.; Zhang, L.; Cai, Y.; et al. Transcriptome Analysis in Sheepgrass (Leymus chinensis): A Dominant Perennial Grass of the Eurasian Steppe. PLoS ONE 2013, 8, e67974. [Google Scholar] [CrossRef]
- Chai, F.; Liu, W.; Xiang, Y.; Meng, X.; Sun, X.; Cheng, C.; Liu, G.; Duan, L.; Xin, H.; Li, S. Comparative metabolic profiling of Vitis amurensis and Vitis vinifera during cold acclimation. Hortic. Res. 2019, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhang, H.; Zhang, J.; Liu, P.; Chen, X.; Li, Z.; Xu, Y.; Lu, P.; Cao, P. Integrated transcriptomics and metabolomics analysis to characterize cold stress responses in Nicotiana tabacum. BMC Genom. 2017, 18, 496. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Li, N.; Zhao, R.; Zhao, M.; Cui, X.; Xu, Y.; Qiao, X. In vitro Prebiotic Properties of Garlic Polysaccharides and Its Oligosaccharide Mixtures Obtained by Acid Hydrolysis. Front. Nutr. 2021, 8, 798450. [Google Scholar] [CrossRef]
- Roberfroid, M.B. Inulin-Type Fructans: Functional Food Ingredients. Br. J. Nutr. 2007, 137, 2493S–2502S. [Google Scholar] [CrossRef]
- Valluru, R.; Ende, W. Plant fructans in stress environments: Emerging concepts and future prospects. J. Exp. Bot. 2008, 59, 2905–2916. [Google Scholar] [CrossRef]
- Tognetti, J.A.; Calderón, P.L.; Pontis, H.G. Fructan metabolism: Reversal of cold acclimation. J. Plant Physiol. 1989, 134, 232–236. [Google Scholar] [CrossRef]
- Kawakami, A.; Yoshida, M. Molecular characterization of sucrose sucrose1-fructosyltransferase and sucrose: Fructan 6-fructosyltransferase associated with fructan accumulation in winter wheat during cold hardening. Biosci. Biotechnol. Biochem. 2002, 66, 2297–2305. [Google Scholar] [CrossRef]
- Hendry, G. Evolutionary origins and natural functions of fructans -a climatological, biogeographic and mechanistic appraisal. New Phytol. 2006, 123, 3–14. [Google Scholar] [CrossRef]
- Shalini, R.; Krishna, J.; Sankaranarayanan, M.; Antony, U. Enhancement of fructan extraction from garlic and fructooligosaccharide purification using an activated charcoal column. LWT-Food Sci. Technol. 2021, 148, 111703. [Google Scholar]
- Chen, J.; Cheong, K.L.; Song, Z.; Shi, Y.; Huang, X. Structure and protective effect on UVB-induced keratinocyte damage of fructan from white garlic. Carbohydr. Polym. 2013, 92, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wu, Z.; Jiang, F. Selection and validation of garlic reference genes for quantitative real-time PCR normalization. Plant Cell Tissue Organ Cult. 2015, 122, 435–444. [Google Scholar] [CrossRef]
- Wang, R.; Shu, P.; Zhang, C.; Zhang, J.; Chen, Y.; Zhang, Y.; Du, K.; Xie, Y.; Li, M.; Ma, T.; et al. Integrative analyses of metabolome and genome-wide transcriptome reveal the regulatory network governing flavor formation in kiwifruit (Actinidia chinensis). New Phytol. 2022, 233, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Li, Q.; Liu, G.; Xu, N.; Yang, Y.; Zeng, W.; Chen, A.; Wang, S. Integrated analysis of transcriptomic and metabolomic data reveals critical metabolic pathways involved in polyphenol biosynthesis in Nicotiana tabacum under chilling stress. Funct. Plant Biol. FPB 2018, 46, 30–43. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593. [Google Scholar] [CrossRef]
- Nägele, T.; Heyer, A.G. Approximating subcellular organisationorganization of carbohydrate metabolism during cold acclimation in different natural accessions of Arabidopsis thaliana. New Phytol. 2013, 198, 777–787. [Google Scholar] [CrossRef]
- Rao, K.M.; Åshild, E.; Arne, R.O. Freezing tolerance revisited-effects of variable temperatures on gene regulation in temperate grasses and legumes. Curr. Opin. Plant Biol. 2016, 33, 140–146. [Google Scholar]
- Livingston, D.P.; Henson, C.A. Apoplastic Sugars, Fructans, Fructan Exohydrolase, and Invertase in Winter Oat: Responses to Second-Phase Cold Hardening. Plant Physiol. 1998, 116, 403–408. [Google Scholar] [CrossRef]
- Janská, A.; Aprile, A.; Zámečník, J.; Cattivelli, L.; Ovesná, J. Transcriptional responses of winter barley to cold indicate nucleosome remodelling as a specific feature of crown tissues. Funct. Integr. Genom. 2011, 11, 307–325. [Google Scholar] [CrossRef]
- Viso, F.D. Molecular characterization of a putative sucrose:fructan 6-fructosyltransferase (6-SFT) of the cold-resistant Patagonian grass Bromus pictus associated with fructan accumulation under low temperatures. Plant Cell Physiol. 2009, 50, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Hincha, D.K.; Hellwege, E.M.; Heyer, A.G.; Crowe, J.H. Plant fructans stabilize phosphatidylcholine liposomes during freeze-drying. Eur. J. Biochem. 2000, 267, 535–540. [Google Scholar] [CrossRef]
- Li, H.J.; Yang, A.F.; Zhang, X.C.; Gao, F.; Zhang, J.R. Improving freezing tolerance of transgenic tobacco expressing sucrose: Sucrose 1-fructosyltransferase gene from Lactuca sativa. Plant Cell Tissue Organ Cult. 2007, 89, 37–48. [Google Scholar] [CrossRef]
- Parvanova, D.; Ivanov, S.; Konstantinova, T.; Karanov, E.; Atanassov, A.; Tsvetkov, T.; Alexieva, V.; Djilianov, D. Transgenic tobacco plants accumulating osmolytes show reduced oxidative damage under freezing stress. Plant Physiol. Biochem. 2004, 42, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Parvanova, D.; Popova, A.; Zaharieva, I.; Lambrev, P.; Konstantinova, T.; Taneva, S.; Atanassov, A.; Goltsev, V.; Djilianov, D. Low temperature tolerance of tobacco plants transformed to accumulate proline, fructans, or glycine betaine. Variable chlorophyll fluorescence evidence. Photosynthetica 2003, 42, 179–185. [Google Scholar] [CrossRef]
- Lee, B.; Henderson, D.A.; Zhu, J. The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell 2005, 17, 3155–3175. [Google Scholar] [CrossRef] [PubMed]
- Mark, S.; Ronan, S.; Joost, K. Metabolic networks: How to identify key components in the regulation of metabolism and growth. Plant Physiol. 2010, 152, 428–444. [Google Scholar]
- Baumgartner, S.; Dax, T.G.; Praznik, W.; Falk, H. Characterisation of the high-molecular weight fructan isolated from garlic (Allium sativum L.). Carbohydr. Res. 2000, 328, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Ernst, M.K.; Chatterton, N.J.; Harrison, P.A.; Matitschka, G. Characterization of fructan oligomers from species of the genus Allium L. J. Plant Physiol. 1998, 153, 53–60. [Google Scholar] [CrossRef]
- Yoshida, M. Fructan Structure and Metabolism in Overwintering Plants. Plants 2021, 10, 933. [Google Scholar] [CrossRef]
- Pego, J.V.; Kortstee, A.J.; Huijser, C.; Smeekens, S.C. Photosynthesis, sugars and the regulation of gene expression. J. Exp. Bot. 2000, 51, 407–416. [Google Scholar] [CrossRef]
- Filip, R.; Elena, B.; Jen, S. Sugar sensing and signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar]
- Lundmark, M.; Cavaco, A.M.; Trevanion, S.; Hurry, V. Carbon partitioning and export in transgenic Arabidopsis thaliana with altered capacity for sucrose synthesis grown at low temperature: A role for metabolite transporters. Plant Cell Environ. 2006, 29, 1703–1714. [Google Scholar] [CrossRef]
- De Coninck, B.; Le Roy, K.; Francis, I.; Clerens, S.; Vergauwen, R.; Halliday, A.M.; Smith, S.M.; Van Laere, A.; Van Den Ende, W. Arabidopsis AtcwINV3 and 6 are not invertases but are fructan exohydrolases (FEHs) with different substrate specificities. Plant Cell Environ. 2010, 28, 432–443. [Google Scholar] [CrossRef]
- Lasseur, B.; Schroeven, L.; Lammens, W.; Le Roy, K.; Spangenberg, G.; Manduzio, H.; Vergauwen, R.; Lothier, J.; Prud, M.-P.; Van den Ende, W. Transforming a Fructan: Fructan 6G-Fructosyltransferase from Perennial Ryegrass into a Sucrose: Sucrose 1-Fructosyltransferase. Plant Physiol. 2009, 149, 327. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Compounds | ko_ID | Regulated | |
|---|---|---|---|---|
| MCK_vs_MDT1 | MCK_vs_MDT5 | |||
| pma0134 | D(-)-Threose | - | - | - |
| pmb0786 | Glucosamine | ko00520, ko01100 | - | - |
| pmb2653 | D(+)-Melezitose O-rhamnoside | - | - | - |
| pmb2858 | Maltotetraose | - | - | up |
| pmb3088 | Trehalose 6-phosphate | ko00500, ko01100 | up | up |
| pme0519 | D-(+)-Sucrose | - | - | up |
| pme1684 | D-(+)-Glucono-1,5-lactone | ko00030, ko01100, ko01110, ko01200 | - | - |
| pme1846 | D(+)-Glucose | ko00010, ko00030, ko00052, ko00500, ko00520, ko00901, ko01100, ko01110 | - | down |
| pme2019 | DL-Arabinose | - | - | down |
| pme2755 | N-Acetyl-D-glucosamine | ko00520, ko01100 | - | - |
| pme3160 | D-Glucose 6-phosphate | ko00500, ko00562, ko00999, ko01100 | - | - |
| pme3163 | D-Sedoheptuiose 7-phosphate | - | - | down |
| pme3313 | D-Fructose 6-phosphate | ko00052, ko00500, ko00710, ko01100, ko01200 | - | - |
| pmf0032 | Galactinol | - | up | - |
| pmf0035 | Glucose-1-phosphate | - | - | - |
| pmf0138 | D-(+)-Mannose | ko00051, ko00052, ko00520, ko01100 | - | down |
| pmf0139 | D-(+)-Galactose | ko00052, ko01100 | - | down |
| pmf0220 | D-Fructose 6-phosphate-disodium salt | - | - | - |
| pmf0282 | Melibiose | ko00052 | - | - |
| pmf0574 | Sucralose | - | - | - |
| Gene Name | Gene ID | KEGG Annotation | TCK_vs_TDT1 | TCK_vs_TDT5 | ||
|---|---|---|---|---|---|---|
| Log2(FC) | Regulated | Log2(FC) | Regulated | |||
| 1-SST (sucrose: sucrose 1-fructosyltransferase gene) | Cluster-4573.149147 | K21351 sucrose:sucrose fructosyltransferase [EC:2.4.1.99] | 2.870591819 | - | 4.205341664 | up |
| Cluster-4573.161561 | K21351 sucrose:sucrose fructosyltransferase [EC:2.4.1.99] | 3.722296577 | up | 4.685898781 | up | |
| Cluster-4573.168322 | K21351 sucrose:sucrose fructosyltransferase [EC:2.4.1.99] | 3.194678677 | up | 4.483060574 | up | |
| Cluster-4573.170978 | K21351 sucrose:sucrose fructosyltransferase [EC:2.4.1.99] | 3.126650449 | up | 3.692679624 | up | |
| 6G-FFT (fructan: fructan 6G-fructosyltransferase gene) | Cluster-4573.161559 | K21352 6(G)-fructosyltransferase [EC:2.4.1.243] | 4.409936024 | up | 5.733601046 | up |
| Cluster-4573.139958 | K21352 6(G)-fructosyltransferase [EC:2.4.1.243] | 3.320464074 | - | 4.081188018 | up | |
| 1-FEH (fructan 1-exohydrolase gene) | Cluster-4573.153574 | K20848 fructan beta-(2,1)-fructosidase [EC:3.2.1.153] | −0.658755386 | - | −1.628771042 | down |
| Cluster-4573.153578 | K20848 fructan beta-(2,1)-fructosidase [EC:3.2.1.153] | - | - | 2.790164693 | up | |
| Cluster-4573.132188 | K20848 fructan beta-(2,1)-fructosidase [EC:3.2.1.153] | - | - | 8.74315447 | up | |
| Cluster-4573.144699 | K20848 fructan beta-(2,1)-fructosidase [EC:3.2.1.153] | - | - | 4.53163043 | up | |
| Cluster-4573.155551 | K20848 fructan beta-(2,1)-fructosidase [EC:3.2.1.153] | −0.481771581 | - | −1.077228392 | down | |
| Cluster-4573.179376 | K20848 fructan beta-(2,1)-fructosidase [EC:3.2.1.153] | 0.484182905 | - | 1.386712343 | up | |
| Gene Name | Gene ID | Module Colors | kWithin |
|---|---|---|---|
| 1-SST | Cluster-4573.149147 | blue | 3970.90 |
| Cluster-4573.161561 | blue | 5022.95 | |
| Cluster-4573.168322 | blue | 4904.94 | |
| 6G-FFT | Cluster-4573.161559 | blue | 5233.82 |
| Cluster-4573.139958 | blue | 3655.29 | |
| 1-FEH | Cluster-4573.153578 | blue | 2584.97 |
| Cluster-4573.132188 | blue | 1437.04 | |
| Cluster-4573.144699 | blue | 3038.59 | |
| Cluster-4573.179376 | blue | 4277.31 | |
| Cluster-4573.153574 | turquoise | 7742.60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bian, H.; Zhou, Q.; Du, Z.; Zhang, G.; Han, R.; Chen, L.; Tian, J.; Li, Y. Integrated Transcriptomics and Metabolomics Analysis of the Fructan Metabolism Response to Low-Temperature Stress in Garlic. Genes 2023, 14, 1290. https://doi.org/10.3390/genes14061290
Bian H, Zhou Q, Du Z, Zhang G, Han R, Chen L, Tian J, Li Y. Integrated Transcriptomics and Metabolomics Analysis of the Fructan Metabolism Response to Low-Temperature Stress in Garlic. Genes. 2023; 14(6):1290. https://doi.org/10.3390/genes14061290
Chicago/Turabian StyleBian, Haiyan, Qianyi Zhou, Zhongping Du, Guangnan Zhang, Rui Han, Laisheng Chen, Jie Tian, and Yi Li. 2023. "Integrated Transcriptomics and Metabolomics Analysis of the Fructan Metabolism Response to Low-Temperature Stress in Garlic" Genes 14, no. 6: 1290. https://doi.org/10.3390/genes14061290
APA StyleBian, H., Zhou, Q., Du, Z., Zhang, G., Han, R., Chen, L., Tian, J., & Li, Y. (2023). Integrated Transcriptomics and Metabolomics Analysis of the Fructan Metabolism Response to Low-Temperature Stress in Garlic. Genes, 14(6), 1290. https://doi.org/10.3390/genes14061290