
MOSR and NDHA Genes Comprising G-Quadruplex as Promising Therapeutic Targets against Mycobacterium tuberculosis: Molecular Recognition by Mitoxantrone Suppresses Replication and Gene Regulation

Abstract
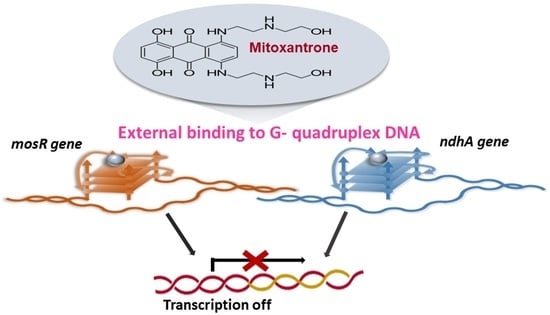
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Surface Plasmon Resonance (SPR)
2.3. Absorption Spectroscopy
2.4. Steady-State Fluorescence
2.5. Method of Continuous Variation (Job Plot)
2.6. Time-Resolved Fluorescence
2.7. Circular Dichroism
2.8. Thermal Profiling (Tm) Using Circular Dichroism (CD)
2.9. Thermal Profiling (Tm) Using Differential Scanning Calorimetry (DSC)
2.10. Taq Polymerase Stop Assay
2.11. Alamar Blue Assay (MIC Calculation)
2.12. Gene Expression by qRT-PCR
3. Results and Discussion
3.1. mosR/ndhA DNA Sequences Adopt Stable G4 DNA Conformation
3.2. Real-Time Binding of Mitoxantrone to mosR/ndhA G4 DNA
3.3. Binding Characteristics of Mitoxantrone-G4 DNA Complex
3.4. Ligand-Induced Conformational Changes in mosR/ndhA G4 DNA
3.5. Ligand-Induced Thermal Stabilization of mosR/ndhA G4 DNA
3.6. Stall Replication Machinery by Taq Polymerase Enzyme Assay
3.7. Mitoxantrone Regulates Gene Expression of mosR and ndhA
3.8. Ligand Binding-Induced Thermal Stabilization Relates to Gene Function
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knight, G.M.; McQuaid, C.F.; Dodd, P.J.; Houben, R.M.G.J. Global Burden of Latent Multidrug-Resistant Tuberculosis: Trends and Estimates Based on Mathematical Modelling. Lancet Infect. Dis. 2019, 19, 903–912. [Google Scholar] [CrossRef] [PubMed]
- De Cian, A.; Lacroix, L.; Douarre, C.; Temime-Smaali, N.; Trentesaux, C.; Riou, J.F.; Mergny, J.L. Targeting Telomeres and Telomerase. Biochimie 2008, 90, 131–155. [Google Scholar] [CrossRef] [PubMed]
- Sekaran, V.; Soares, J.; Jarstfer, M.B. Telomere Maintenance as a Target for Drug Discovery. J. Med. Chem. 2014, 57, 521–538. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-Quadruplexes in Gene Promoters: A Novel Anticancer Strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef]
- Perrone, R.; Lavezzo, E.; Riello, E.; Manganelli, R.; Palù, G.; Toppo, S.; Provvedi, R.; Richter, S.N. Mapping and Characterization of G-Quadruplexes in Mycobacterium Tuberculosis Gene Promoter Regions. Sci. Rep. 2017, 7, 5743. [Google Scholar] [CrossRef]
- Mishra, S.K.; Shankar, U.; Jain, N.; Sikri, K.; Tyagi, J.S.; Sharma, T.K.; Mergny, J.L.; Kumar, A. Characterization of G-Quadruplex Motifs in EspB, EspK, and Cyp51 Genes of Mycobacterium Tuberculosis as Potential Drug Targets. Mol. Ther.—Nucleic Acids 2019, 16, 698–706. [Google Scholar] [CrossRef]
- Harbut, M.B.; Yang, B.; Liu, R.; Yano, T.; Vilchèze, C.; Cheng, B.; Lockner, J.; Guo, H.; Yu, C.; Franzblau, S.G.; et al. Small Molecules Targeting Mycobacterium Tuberculosis Type II NADH Dehydrogenase Exhibit Antimycobacterial Activity. Angew. Chem.—Int. Ed. 2018, 57, 3478–3482. [Google Scholar] [CrossRef]
- Abomoelak, B.; Hoye, E.A.; Chi, J.; Marcus, S.A.; Laval, F.; Bannantine, J.P.; Ward, S.K.; Daffé, M.; Hong, D.L.; Talaat, A.M. MosR, a Novel Transcriptional Regulator of Hypoxia and Virulence in Mycobacterium Tuberculosis. J. Bacteriol. 2009, 191, 5941–5952. [Google Scholar] [CrossRef]
- Vilchèze, C.; Weinrick, B.; Leung, L.W.; Jacobs, W.R. Plasticity of Mycobacterium Tuberculosis NADH Dehydrogenases and Their Role in Virulence. Proc. Natl. Acad. Sci. USA 2018, 115, 1599–1604. [Google Scholar] [CrossRef]
- Kikin, O.; D’Antonio, L.; Bagga, P.S. QGRS Mapper: A Web-Based Server for Predicting G-Quadruplexes in Nucleotide Sequences. Nucleic Acids Res. 2006, 34, 676–682. [Google Scholar] [CrossRef]
- Puig Lombardi, E.; Londoño-Vallejo, A. A Guide to Computational Methods for G-Quadruplex Prediction. Nucleic Acids Res. 2020, 48, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C.; et al. Deciphering the Biology of Mycobacterium Tuberculosis from the Complete Genome Sequence. Nature 1998, 393, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Thakur, R.S.; Desingu, A.; Basavaraju, S.; Subramanya, S.; Rao, D.N.; Nagaraju, G. Mycobacterium Tuberculosis DinG Is a Structure-Specific Helicase That Unwinds G4 DNA: Implications for Targeting G4 DNA as a Novel Therapeutic Approach. J. Biol. Chem. 2014, 289, 25112–25136. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, E.; Richter, S.N. Survey and Summary G-Quadruplexes and G-Quadruplex Ligands: Targets and Tools in Antiviral Therapy. Nucleic Acids Res. 2018, 46, 3270–3283. [Google Scholar] [CrossRef] [PubMed]
- Vorlíčková, M.; Kejnovská, I.; Sagi, J.; Renčiuk, D.; Bednářová, K.; Motlová, J.; Kypr, J. Circular dichroism and guanine quadruplexes. Methods 2012, 57, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Kypr, J.; Kejnovská, I.; Renčiuk, D.; Vorlíčková, M. Circular dichroism and conformational polymorphism of DNA. Nucleic Acids Res. 2009, 37, 1713–1725. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Wu, G.; Wang, K.; Onel, B.; Sakai, S.; Shao, Y.; Yang, D. Molecular Recognition of the Hybrid-2 Human Telomeric G-Quadruplex by Epiberberine. Angew. Chem. 2018, 57, 10888–10893. [Google Scholar] [CrossRef] [PubMed]
- Phan, A.T.; Luu, K.N.; Patel, D.J. Different Loop Arrangements of Intramolecular Human Telomeric (3+1) G-Quadruplexes in K+ Solution. Nucleic Acids Res. 2006, 34, 5715–5719. [Google Scholar] [CrossRef]
- Largy, E.; Mergny, J.L.; Gabelica, V. Role of alkali metal ions in G-Quadruplex nucleic acid structure and stability. Met. Ions. Life Sci. 2016, 16, 203–258. [Google Scholar] [CrossRef]
- Dey, A.; Pandav, K.; Nath, M.; Barthwal, R.; Prasad, R. Molecular recognition of telomere DNA Sequence by 2, 6 anthraquinone derivatives leads to thermal stabilization and induces apoptosis in cancer cells. Int. J. Biol. Macromol. 2022, 221, 355–370. [Google Scholar] [CrossRef]
- Pradeep, T.P.; Barthwal, R. A 4:1 Stoichiometric Binding and Stabilization of Mitoxantrone-Parallel Stranded G-Quadruplex Complex Established by Spectroscopy Techniques. J. Photochem. Photobiol. B Biol. 2016, 162, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Anand, K.; Tripathi, A.; Shukla, K.; Malhotra, N. Redox Biology Mycobacterium Tuberculosis SufR Responds to Nitric Oxide via Its 4Fe–4S Cluster and Regulates Fe–S Cluster Biogenesis for Persistence in Mice. Redox Biol. 2021, 46, 102062. [Google Scholar] [CrossRef] [PubMed]
- Krugh, T.R.; Reinhardt, C.G. Evidence for Sequence Preferences in the Intercalative Binding of Ethidium Bromide to Dinucleoside Monophosphates. J. Mol. Biol. 1975, 1971, 133–162. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Wan, C.; Becker, H.C.; Zhong, D.; Zewail, A.H. The Anticancer Drug-DNA Complex: Femtosecond Primary Dynamics for Anthracycline Antibiotics Function. Proc. Natl. Acad. Sci. USA 2001, 98, 14212–14217. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Struve, W.S. Solvatochromism and Time-Resolved Fluorescence of the Antitumor Agent Mitoxantrone and Its Analogues in Solution and in DNA. J. Phys. Chem. 1991, 95, 2251–2256. [Google Scholar] [CrossRef]
- Wei, C.; Wang, L.; Jia, G.; Zhou, J.; Han, G.; Li, C. The Binding Mode of Porphyrins with Cation Side Arms to (TG4T)4 G-Quadruplex: Spectroscopic Evidence. Biophys. Chem. 2009, 143, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.; Pradeep, T.P.; Barthwal, R. Molecular Recognition of Parallel DNA Quadruplex d(TTAGGGT)4 by Mitoxantrone: Binding with 1:2 Stoichiometry Leading to Thermal Stabilization and Telomerase Inhibition. ChemBioChem 2016, 17, 554–560. [Google Scholar] [CrossRef] [PubMed]
- White, E.W.; Tanious, F.; Ismail, M.A.; Reszka, A.P.; Neidle, S.; Boykin, D.W.; Wilson, W.D. Structure-Specific Recognition of Quadruplex DNA by Organic Cations: Influence of Shape, Substituents and Charge. Biophys. Chem. 2007, 126, 140–153. [Google Scholar] [CrossRef] [PubMed]
- Rubio-Magnieto, J.; Di Meo, F.; Lo, M.; Delcourt, C.; Clément, S.; Norman, P.; Richeter, S.; Linares, M.; Surin, M. Binding Modes of a Core-Extended Metalloporphyrin to Human Telomeric DNA G-Quadruplexes. Org. Biomol. Chem. 2015, 13, 2453–2463. [Google Scholar] [CrossRef] [PubMed]
- Percivalle, C.; Sissi, C.; Greco, M.L.; Musetti, C.; Mariani, A.; Artese, A.; Costa, G.; Perrore, M.L.; Alcaro, S.; Freccero, M. Aryl Ethynyl Anthraquinones: A Useful Platform for Targeting Telomeric G-Quadruplex Structures. Org. Biomol. Chem. 2014, 12, 3744–3754. [Google Scholar] [CrossRef]
- Jain, A.K.; Reddy, V.V.; Paul, A.; Muniyappa, K.; Bhattacharya, S. Synthesis and Evaluation of a Novel Class of G-Quadruplex-Stabilizing Small Molecules Based on the 1,3-Phenylene-Bis(Piperazinyl Benzimidazole) System. Biochemistry 2009, 48, 10693–10704. [Google Scholar] [CrossRef] [PubMed]
- Lown, J.W.; Morgan, A.R.; Yen, S.F.; Wang, Y.H.; Wilson, W.D. Characteristics of the binding of the anticancer agents mitoxantrone and ametantrone and related structures to deoxyribonucleic acids. Biochemistry 1985, 24, 4028–4035. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Jangir, D.K.; Mehrotra, R. Spectroscopic Studies of the effects of anticancer drug mitoxantrone interaction with calf-thymus DNA. J. Photochem. Photobiol. B Biol. 2013, 120, 177–182. [Google Scholar] [CrossRef]
- Bhattacharyya, J.; Basu, A.; Suresh Kumar, G. Intercalative Interaction of the anticancer drug mitoxantrone with double stranded dna: A calorimetric characterization of the energetics. J. Chem. Thermodyn. 2014, 75, 45–51. [Google Scholar] [CrossRef]
- Dogra, S.; Awasthi, P.; Nair, M.; Barthwal, R. Interaction of anticancer drug mitoxantrone with DNA hexamer sequence D-(CTCGAG)2 by absorption, fluorescence and circular dichroism spectroscopy. J. Photochem. Photobiol. B Biol. 2013, 123, 48–54. [Google Scholar] [CrossRef]
- Barthwal, R.; Tariq, Z. Molecular Recognition of Parallel G-quadruplex [d-(TTGGGGT)]4 Containing Tetrahymena Telomeric DNA Sequence by Anticancer Drug Daunomycin: NMR-Based Structure and Thermal Stability. Molecules 2018, 23, 2266. [Google Scholar] [CrossRef]
- Tariq, Z.; Barthwal, R. Binding of anticancer drug daunomycin to parallel G-quadruplex DNA [d-(TTGGGGT)]4 leads to thermal stabilization: A multispectroscopic investigation. Int J Biol Macromol. 2018, 120, 1965–1974. [Google Scholar] [CrossRef]
- Hargrove, A.E.; Zhong, Z.; Sessler, J.L.; Anslyn, E.V. Algorithms for the determination of binding constants and enantiomeric excess in complex host: Guest equilibria using optical measurements. New J. Chem. 2010, 34, 348–354. [Google Scholar] [CrossRef]
- Padiadpu, J.; Baloni, P.; Anand, K.; Munshi, M.; Thakur, C.; Mohan, A.; Singh, A.; Chandra, N. Identifying and Tackling Emergent Vulnerability in Drug-Resistant Mycobacteria. ACS Infect. Dis. 2016, 2, 592–607. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Tm1 | ∆Tm1 | Tm2 | ∆Tm2 | Tm3 | ∆Tm3 | χ2/DOF |
|---|---|---|---|---|---|---|---|
| free mosR | 47.3 ± 0.6 | - | 58.3 ± 0.2 * | - | 71.4 ± 0.5 | - | 1.7 × 104 |
| mosR + MTX | 67.2 ± 1.6 | 19.9 | 83.6 ± 0.3 * | 25.3 | 100.2 ± 0.4 | 28.8 | 1.9 × 105 |
| free ndhA | 46.8 ± 0.5 | - | 64.0 ± 0.4 | - | 84.4 ± 0.3 * | - | 9.9 × 104 |
| ndhA + MTX | 46.5 ± 0.4 | - | 81.9 * ± 0.5 | 17.9 | 94.5 ± 0.3 | 10.1 | 1.4 × 105 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dey, A.; Anand, K.; Singh, A.; Prasad, R.; Barthwal, R. MOSR and NDHA Genes Comprising G-Quadruplex as Promising Therapeutic Targets against Mycobacterium tuberculosis: Molecular Recognition by Mitoxantrone Suppresses Replication and Gene Regulation. Genes 2023, 14, 978. https://doi.org/10.3390/genes14050978
Dey A, Anand K, Singh A, Prasad R, Barthwal R. MOSR and NDHA Genes Comprising G-Quadruplex as Promising Therapeutic Targets against Mycobacterium tuberculosis: Molecular Recognition by Mitoxantrone Suppresses Replication and Gene Regulation. Genes. 2023; 14(5):978. https://doi.org/10.3390/genes14050978
Chicago/Turabian StyleDey, Arpita, Kushi Anand, Amit Singh, Ramasare Prasad, and Ritu Barthwal. 2023. "MOSR and NDHA Genes Comprising G-Quadruplex as Promising Therapeutic Targets against Mycobacterium tuberculosis: Molecular Recognition by Mitoxantrone Suppresses Replication and Gene Regulation" Genes 14, no. 5: 978. https://doi.org/10.3390/genes14050978
APA StyleDey, A., Anand, K., Singh, A., Prasad, R., & Barthwal, R. (2023). MOSR and NDHA Genes Comprising G-Quadruplex as Promising Therapeutic Targets against Mycobacterium tuberculosis: Molecular Recognition by Mitoxantrone Suppresses Replication and Gene Regulation. Genes, 14(5), 978. https://doi.org/10.3390/genes14050978