

Metabarcoding of Antarctic Lichens from Areas with Different Deglaciation Times Reveals a High Diversity of Lichen-Associated Communities


Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. Sample Preparation, Lysis, and DNA Extraction
2.3. DNA Amplification and Metabarcoding
2.4. Barcode Sequence Analysis, Processing, and Amplicon Sequencing Variant (ASV) Identification
2.5. Sample Pooling, Data Exclusion, and Plausibility Control
2.6. KRONA Metagenomic Visualisation
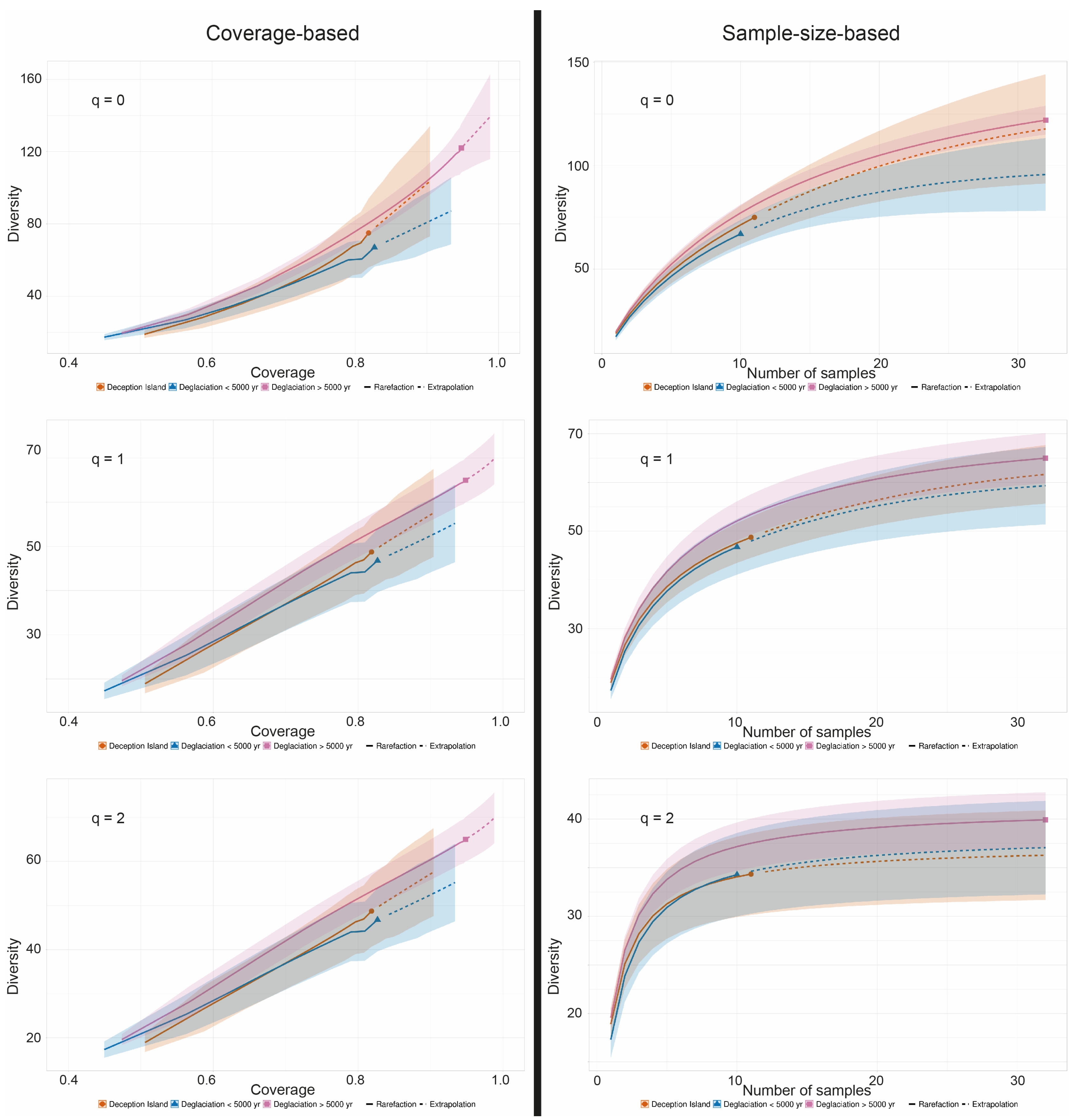
2.7. Diversity Estimations
3. Results and Discussion
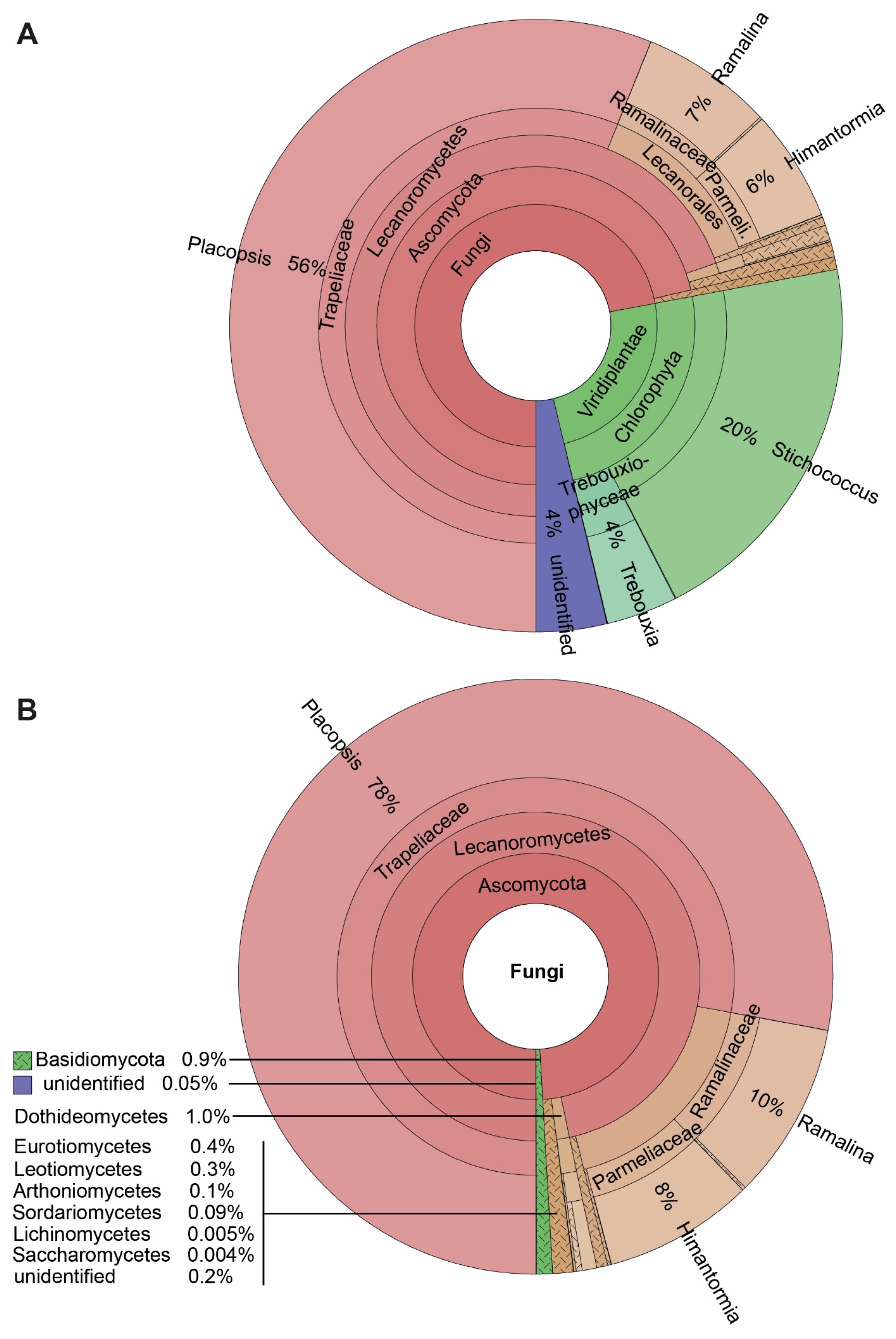
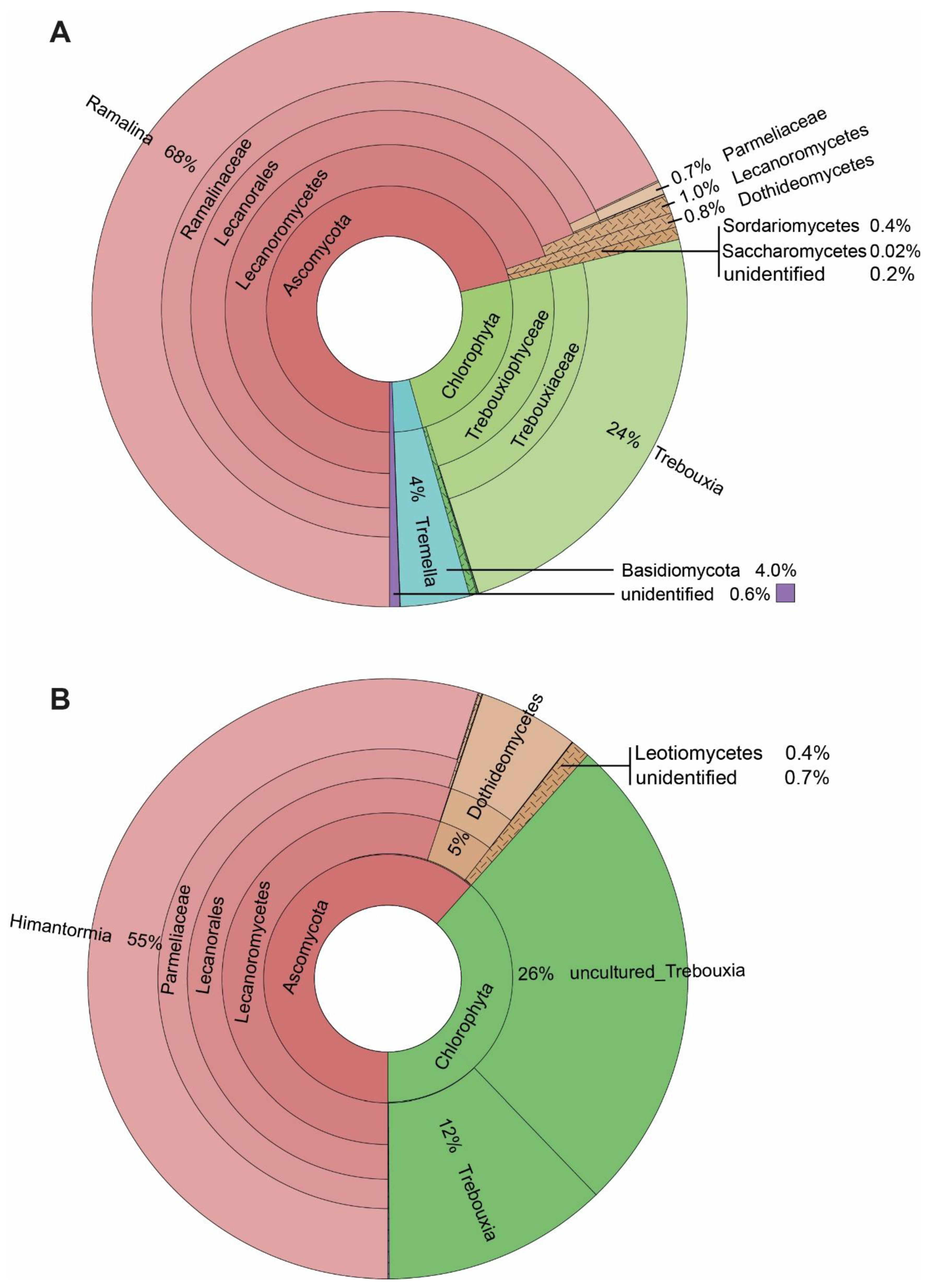
3.1. Eukaryotes Associated with the Lichens Himantormia, Placopsis, and Ramalina
3.2. ASVs Associated with the Fruticose Lichens Ramalina terebrata and Himantormia lugubris
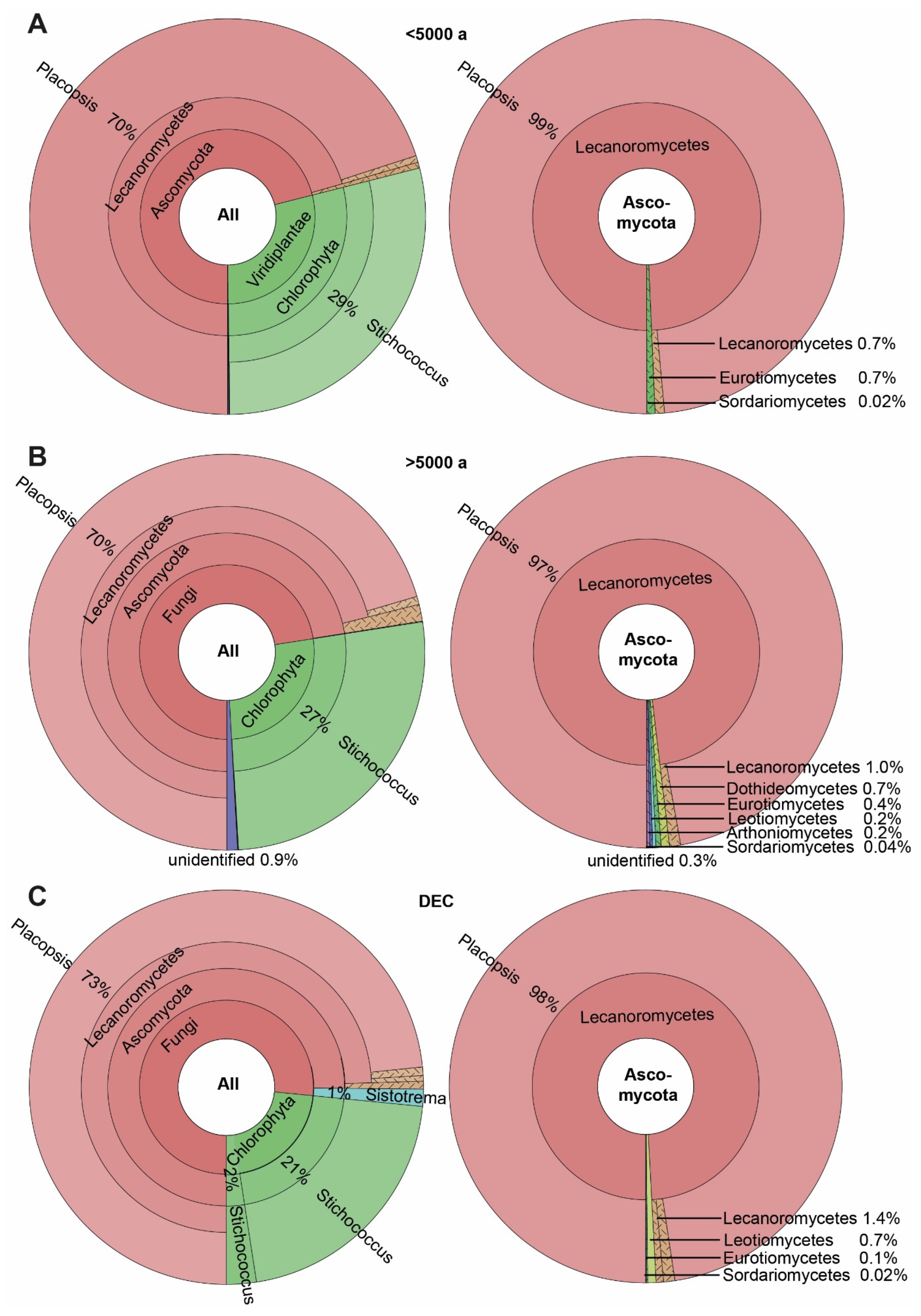
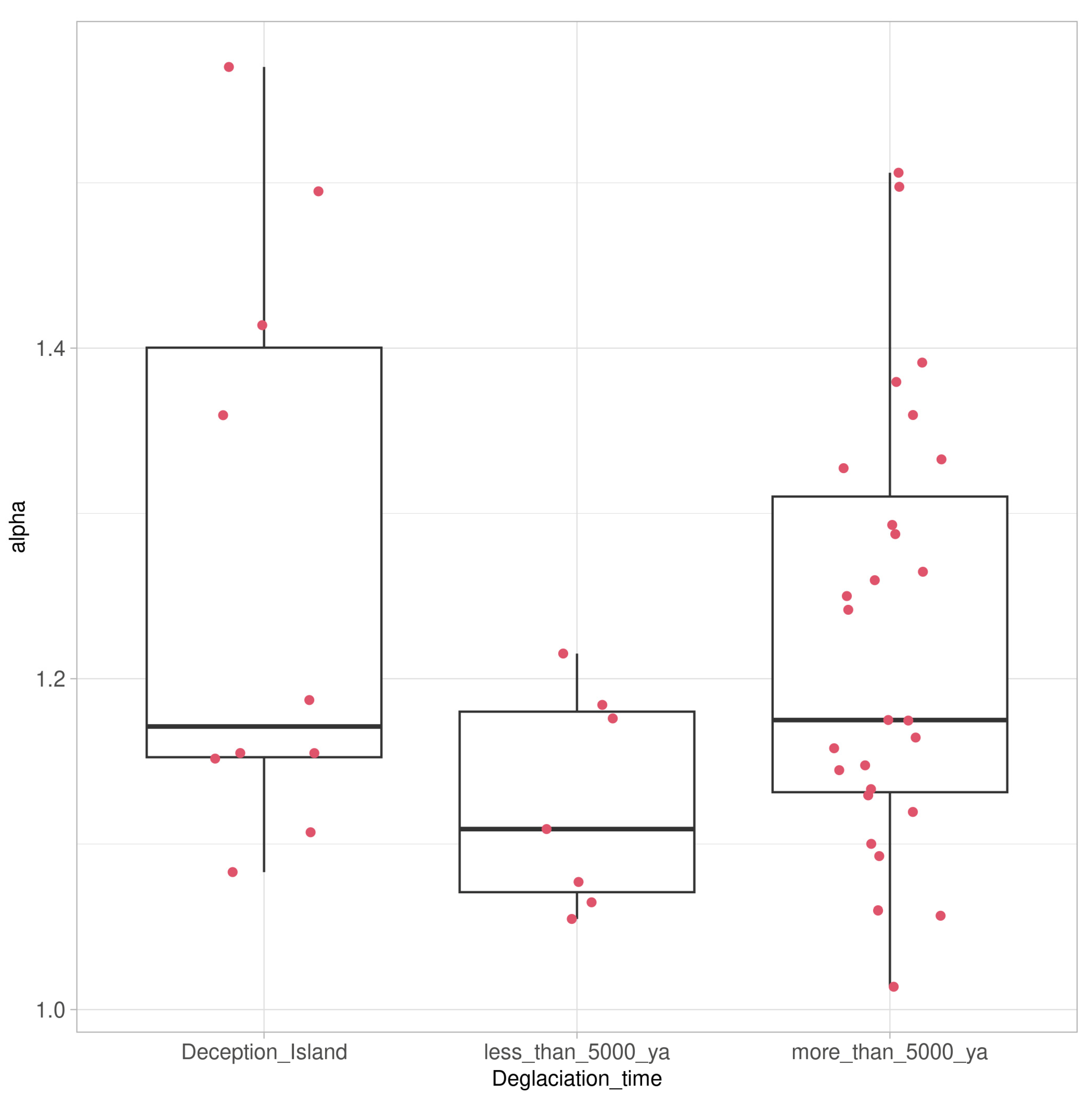
3.3. ASVs Associated with the Crustose Lichen Genus Placopsis Growing on Soils with Different Deglaciation Times
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Øvstedal, D.O.; Lewis Smith, R.I. Lichens of Antarctica and South Georgia: A Guide to Their Identification and Ecology; Taylor & Francis: Cambridge, UK, 2001; ISBN 9780521662413. [Google Scholar]
- Elbert, W.; Weber, B.; Burrows, S.; Steinkamp, J.; Büdel, B.; Andreae, M.O.; Pöschl, U. Contribution of Cryptogamic Covers to the Global Cycles of Carbon and Nitrogen. Nat. Geosci. 2012, 5, 459–462. [Google Scholar] [CrossRef]
- Lindsay, D.C. The Role of Lichens in Antarctic Ecosystems. Bryologist 1978, 81, 268–276. [Google Scholar] [CrossRef]
- Green, T.G.A.; Schroeter, B.; Sancho, L.G. Plants in Antarctica. In Handbook of Functional Plant Ecology; Marcel Dekker: New York, NY, USA, 2007; pp. 389–434. ISBN 978-0-429-12247-7. [Google Scholar]
- Poelking, E.L.; Schaefer, C.E.R.; Fernandes Filho, E.I.; de Andrade, A.M.; Spielmann, A.A. Soil–Landform–Plant-Community Relationships of a Periglacial Landscape on Potter Peninsula, Maritime Antarctica. Solid Earth 2015, 6, 583–594. [Google Scholar] [CrossRef]
- Nash, I.; Thomas, H. Lichen Biology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2008; ISBN 978-0-521-87162-4. [Google Scholar]
- Honegger, R. Plant Relationships; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Farrar, J.F. Lichen as an Ecosystem: Observation and Experiment. In Lichenology: Progress and Problems; Proceedings of an International Symposium; Academic Press: London, UK, 1976. [Google Scholar]
- Rambold, G.; Triebel, D. The Inter-Lecanoralean Associations; Bibliotheca Lichenologica: Stuttgart, Germany, 1992; ISBN 978-3-443-58027-8. [Google Scholar]
- Diederich, P.; Lawrey, J.D.; Ertz, D. The 2018 Classification and Checklist of Lichenicolous Fungi, with 2000 Non-Lichenised, Obligately Lichenicolous Taxa. Bryologist 2018, 121, 340–425. [Google Scholar] [CrossRef]
- Honegger, R. Functional Aspects of the Lichen Symbiosis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 553–578. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Grube, M. Lichens Redefined as Complex Ecosystems. New Phytol. 2020, 227, 1281–1283. [Google Scholar] [CrossRef] [PubMed]
- Spribille, T.; Tuovinen, V.; Resl, P.; Vanderpool, D.; Wolinski, H.; Aime, M.C.; Schneider, K.; Stabentheiner, E.; Toome-Heller, M.; Thor, G.; et al. Basidiomycete Yeasts in the Cortex of Ascomycete Macrolichens. Science 2016, 353, 488–492. [Google Scholar] [CrossRef]
- Cernava, T.; Müller, H.; Aschenbrenner, I.A.; Grube, M.; Berg, G. Analysing the Antagonistic Potential of the Lichen Microbiome against Pathogens by Bridging Metagenomic with Culture Studies. Front. Microbiol. 2015, 6, 620. [Google Scholar] [CrossRef]
- Cernava, T.; Erlacher, A.; Aschenbrenner, I.A.; Krug, L.; Lassek, C.; Riedel, K.; Grube, M.; Berg, G. Deciphering Functional Diversification within the Lichen Microbiota by Meta-Omics. Microbiome 2017, 5, 82. [Google Scholar] [CrossRef]
- Grube, M.; Cernava, T.; Soh, J.; Fuchs, S.; Aschenbrenner, I.; Lassek, C.; Wegner, U.; Becher, D.; Riedel, K.; Sensen, C.W.; et al. Exploring Functional Contexts of Symbiotic Sustain within Lichen-Associated Bacteria by Comparative Omics. ISME J. 2015, 9, 412–424. [Google Scholar] [CrossRef]
- Parks, D.H.; Rinke, C.; Chuvochina, M.; Chaumeil, P.-A.; Woodcroft, B.J.; Evans, P.N.; Hugenholtz, P.; Tyson, G.W. Recovery of Nearly 8,000 Metagenome-Assembled Genomes Substantially Expands the Tree of Life. Nat. Microbiol. 2017, 2, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.T.; Cropsey, G.W.; Caporaso, J.G.; Knight, R.; Fierer, N. Bacterial Communities Associated with the Lichen Symbiosis. Appl. Environ. Microbiol. 2011, 77, 1309–1314. [Google Scholar] [CrossRef] [PubMed]
- Bemm, F.; Weiß, C.L.; Schultz, J.; Förster, F. Genome of a Tardigrade: Horizontal Gene Transfer or Bacterial Contamination? Proc. Natl. Acad. Sci. USA 2016, 113, E3054–E3056. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, I.; Yamamoto, Y.; Nakano, T.; Finnie, J. Isolation and Culture of Lichen Photobionts and Mycobionts. In Protocols in Lichenology: Culturing, Biochemistry, Ecophysiology and Use in Biomonitoring; Kranner, I.C., Beckett, R.P., Varma, A.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 3–33. ISBN 978-3-642-56359-1. [Google Scholar]
- Abdel-Hameed, M.; Bertrand, R.L.; Piercey-Normore, M.D.; Sorensen, J.L. Putative Identification of the Usnic Acid Biosynthetic Gene Cluster by de Novo Whole-Genome Sequencing of a Lichen-Forming Fungus. Fungal Biol. 2016, 120, 306–316. [Google Scholar] [CrossRef]
- Dal Grande, F.; Meiser, A.; Greshake Tzovaras, B.; Otte, J.; Ebersberger, I.; Schmitt, I. The Draft Genome of the Lichen-Forming Fungus Lasallia hispanica (Frey) Sancho & A. Crespo. Lichenologist 2018, 50, 329–340. [Google Scholar] [CrossRef]
- Park, C.H.; Kim, K.M.; Elvebakk, A.; Kim, O.-S.; Jeong, G.; Hong, S.G. Algal and Fungal Diversity in Antarctic Lichens. J. Eukaryot. Microbiol. 2015, 62, 196–205. [Google Scholar] [CrossRef]
- Tuovinen, V.; Ekman, S.; Thor, G.; Vanderpool, D.; Spribille, T.; Johannesson, H. Two Basidiomycete Fungi in the Cortex of Wolf Lichens. Curr. Biol. 2019, 29, 476–483.e5. [Google Scholar] [CrossRef]
- Lendemer, J.C.; Keepers, K.G.; Tripp, E.A.; Pogoda, C.S.; McCain, C.M.; Kane, N.C. A Taxonomically Broad Metagenomic Survey of 339 Species Spanning 57 Families Suggests Cystobasidiomycete Yeasts Are Not Ubiquitous across All Lichens. Am. J. Bot. 2019, 106, 1090–1095. [Google Scholar] [CrossRef]
- Casano, L.M.; del Campo, E.M.; García-Breijo, F.J.; Reig-Armiñana, J.; Gasulla, F.; del Hoyo, A.; Guéra, A.; Barreno, E. Two Trebouxia Algae with Different Physiological Performances Are Ever-Present in Lichen Thalli of Ramalina farinacea. Coexistence versus Competition? Environ. Microbiol. 2011, 13, 806–818. [Google Scholar] [CrossRef]
- Černajová, I.; Steinová, J.; Škvorová, Z.; Škaloud, P. The Curious Case of Cladonia luteoalba: No Support for Its Distinction. Lichenologist 2022, 54, 345–354. [Google Scholar] [CrossRef]
- Robison, A.; Baugh, M.; Muggia, L.; Leavitt, S.D. Fruticose Lichen Communities at the Edge: Distribution and Diversity in a Desert Sky Island on the Colorado Plateau. Conservation 2022, 2, 550–565. [Google Scholar] [CrossRef]
- Cometto, A.; Leavitt, S.D.; Millanes, A.M.; Wedin, M.; Grube, M.; Muggia, L. The Yeast Lichenosphere: High Diversity of Basidiomycetes from the Lichens Tephromela atra and Rhizoplaca melanophthalma. Fungal Biol. 2022, 126, 587–608. [Google Scholar] [CrossRef]
- Zhang, T.; Yan, D.; Ji, Z.; Chen, X.; Yu, L. A Comprehensive Assessment of Fungal Communities in Various Habitats from an Ice-Free Area of Maritime Antarctica: Diversity, Distribution, and Ecological Trait. Environ. Microbiome 2022, 17, 54. [Google Scholar] [CrossRef]
- Coleine, C.; Zucconi, L.; Onofri, S.; Pombubpa, N.; Stajich, J.E.; Selbmann, L. Sun Exposure Shapes Functional Grouping of Fungi in Cryptoendolithic Antarctic Communities. Life 2018, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.J.; Hollingsworth, P.M.; Coppins, B.J.; Ellis, C.J.; Harrold, P.; Tosh, J.; Yahr, R. DNA Barcoding of Lichenized Fungi Demonstrates High Identification Success in a Floristic Context. New Phytol. 2011, 191, 288–300. [Google Scholar] [CrossRef]
- Flück, D. DNA Barcoding of Swiss Epiphytic Crustose Lichens—A Feasibility Study. Master’s Thesis, University of Bern, Bern, Switzerland, 2012. [Google Scholar]
- Divakar, P.K.; Leavitt, S.D.; Molina, M.C.; Del-Prado, R.; Lumbsch, H.T.; Crespo, A. A DNA Barcoding Approach for Identification of Hidden Diversity in Parmeliaceae (Ascomycota): Parmelia Sensu Stricto as a Case Study. Bot. J. Linn. Soc. 2016, 180, 21–29. [Google Scholar] [CrossRef]
- Mark, K.; Laanisto, L.; Bueno, C.G.; Niinemets, Ü.; Keller, C.; Scheidegger, C. Contrasting Co-Occurrence Patterns of Photobiont and Cystobasidiomycete Yeast Associated with Common Epiphytic Lichen Species. New Phytol. 2020, 227, 1362–1375. [Google Scholar] [CrossRef]
- Banchi, E.; Stankovic, D.; Fernández-Mendoza, F.; Gionechetti, F.; Pallavicini, A.; Muggia, L. ITS2 Metabarcoding Analysis Complements Lichen Mycobiome Diversity Data. Mycol. Prog. 2018, 17, 1049–1066. [Google Scholar] [CrossRef] [PubMed]
- Spielmann, A.A.; Pereira, A.B. Lichens on the Maritime Antarctica. A Small Field Guide for Some Common Species. Glal 2012, 4, 1–28. [Google Scholar]
- Beck, A.; Bechteler, J.; Casanova-Katny, A.; Dzhilyanova, I. The Pioneer Lichen Placopsis in Maritime Antarctica: Genetic Diversity of Their Mycobionts and Green Algal Symbionts, and Their Correlation with Deglaciation Time. Symbiosis 2019, 79, 1–24. [Google Scholar] [CrossRef]
- Beck, A.; Mayr, C. Nitrogen and Carbon Isotope Variability in the Green-Algal Lichen Xanthoria parietina and Their Implications on Mycobiont–Photobiont Interactions. Ecol. Evol. 2012, 2, 3132–3144. [Google Scholar] [CrossRef]
- Ihrmark, K.; Bödeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandström-Durling, M.; Clemmensen, K.E.; et al. New Primers to Amplify the Fungal ITS2 Region—Evaluation by 454-Sequencing of Artificial and Natural Communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Morinière, J.; de Araujo, B.C.; Lam, A.W.; Hausmann, A.; Balke, M.; Schmidt, S.; Hendrich, L.; Doczkal, D.; Fartmann, B.; Arvidsson, S.; et al. Species Identification in Malaise Trap Samples by DNA Barcoding Based on NGS Technologies and a Scoring Matrix. PLoS ONE 2016, 11, e0155497. [Google Scholar] [CrossRef] [PubMed]
- Deagle, B.E.; Thomas, A.C.; Shaffer, A.K.; Trites, A.W.; Jarman, S.N. Quantifying Sequence Proportions in a DNA-Based Diet Study Using Ion Torrent Amplicon Sequencing: Which Counts Count? Mol. Ecol. Resour. 2013, 13, 620–633. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive Metagenomic Visualization in a Web Browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef]
- Hsieh, T.; Ma, K.; Chao, C. INEXT: An R Package for Rarefaction and Extrapolation of Species Diversity (Hill Numbers). Methods Ecol. Evol 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and Extrapolation with Hill Numbers: A Framework for Sampling and Estimation in Species Diversity Studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and Estimators Linking Individual-Based and Sample-Based Rarefaction, Extrapolation and Comparison of Assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.; Simpson, G.; Solymos, P. Vegan: Community Ecology Package (Version 2.5-6). The Comprehensive R Archive Network. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 April 2023).
- De Mendiburu, F. Agricolae Tutorial (Version 1.3-5); Universidad Nacional Agraria: La Molina, Peru, 2021. [Google Scholar]
- Wickham, H.; Chang, W.; Wickham, M.H. Package ‘Ggplot2’. Create Elegant Data Visualisations Using the Grammar of Graphics; Springer: New York, NY, USA, 2016; Volume 2, pp. 1–189. [Google Scholar]
- Sancho, L.; de los Ríos, A.; Pintado, A.; Colesie, C.; Raggio, J.; Ascaso, C.; Green, A. Himantormia lugubris, an Antarctic Endemic on the Edge of the Lichen Symbiosis. Symbiosis 2020, 82, 49–58. [Google Scholar] [CrossRef]
- Gálvez, F.E.; Saldarriaga-Córdoba, M.; Huovinen, P.; Silva, A.X.; Gómez, I. Revealing the Characteristics of the Antarctic Snow Alga Chlorominima collina Gen. et Sp. Nov. Through Taxonomy, Physiology, and Transcriptomics. Front. Plant Sci. 2021, 12, 1050. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global Diversity and Geography of Soil Fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef]
- Lawrey, J.D.; Sikaroodi, M.; Gillevet, P.M.; Diederich, P. A New Species of Bulbil-Forming Lichenicolous Fungi Represents an Isolated Clade in the Cantharellales. Bryologist 2020, 123, 155–162. [Google Scholar] [CrossRef]
- Kondratyuk, S.Y.; Upreti, D.K.; Mishra, G.K.; Nayaka, S.; Ingle, K.K.; Orlov, O.O.; Kondratiuk, A.S.; Lőkös, L.; Farkas, E.; Woo, J.-J.; et al. New and Noteworthy Lichen-Forming and Lichenicolous Fungi 10. Acta Bot. Hung. 2020, 62, 69–108. [Google Scholar] [CrossRef]
- Dal Grande, F.; Rolshausen, G.; Divakar, P.K.; Crespo, A.; Otte, J.; Schleuning, M.; Schmitt, I. Environment and Host Identity Structure Communities of Green Algal Symbionts in Lichens. New Phytol. 2018, 217, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Rolshausen, G.; Dal Grande, F.; Otte, J.; Schmitt, I. Lichen Holobionts Show Compositional Structure along Elevation. Mol. Ecol. 2022, 1–12. [Google Scholar] [CrossRef]
- Lee, J.R.; Raymond, B.; Bracegirdle, T.J.; Chadès, I.; Fuller, R.A.; Shaw, J.D.; Terauds, A. Climate Change Drives Expansion of Antarctic Ice-Free Habitat. Nature 2017, 547, 49–54. [Google Scholar] [CrossRef]
- Lee, J.R.; Waterman, M.J.; Shaw, J.D.; Bergstrom, D.M.; Lynch, H.J.; Wall, D.H.; Robinson, S.A. Islands in the Ice: Potential Impacts of Habitat Transformation on Antarctic Biodiversity. Glob. Chang. Biol. 2022, 28, 5865–5880. [Google Scholar] [CrossRef]
- Gyeong, H.; Hyun, C.-U.; Kim, S.C.; Tripathi, B.M.; Yun, J.; Kim, J.; Kang, H.; Kim, J.H.; Kim, S.; Kim, M. Contrasting Early Successional Dynamics of Bacterial and Fungal Communities in Recently Deglaciated Soils of the Maritime Antarctic. Mol. Ecol. 2021, 30, 4231–4244. [Google Scholar] [CrossRef]
- Beck, A.; Peršoh, D.; Rambold, G. First Evidence for Seasonal Fluctuations in Lichen- and Bark-Colonising Fungal Communities. Folia Microbiol. 2014, 59, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Kappen, L. Field Measurements of Carbon Dioxide Exchange of the Antarctic Lichen Usnea sphacelata in the Frozen State. Antarct. Sci. 1989, 1, 31–34. [Google Scholar] [CrossRef]
- Schroeter, B.; Green, T.G.A.; Kappen, L.; Seepelt, R.D. Carbon Dioxide Exchange at Subzero Temperatures. Field Measurements on Umbilicaria aprina in Antarctica. Cryptogam. Bot. 1994, 4, 233–241. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa Used in This Study | Voucher Information | Deglaciation Time | Herbarium Number/Collection Number | Sample Number |
|---|---|---|---|---|
| Himantormia lugubris | Antarctica, King George Island, Ardley. S 62°12.622′, W 58°55.848′; 10 m asl. | >5000 a | M-0019715/140116b | B1583 |
| H. lugubris | Antarctica, King George Island, Fildes Peninsula, Co Gemelos. S 62°11.780′, W 58°59.602′; 33 m asl. | >5000 a | M-0019714/140111 | B1582 |
| H. lugubris | Antarctica, King George Island, Fildes Peninsula, Meseta La Cruz. S 62°12.360′, W 58°57.239′; 49 m asl. | >5000 a | M-0019719/140109 | B1587 |
| H. lugubris | Antarctica, King George Island, Fildes Peninsula, Meseta La Cruz. S 62°12.360′, W 58°57.239′; 49 m asl. | >5000 a | M-0019708 | B1666 |
| H. lugubris | Antarctica, King George Island, Fildes Peninsula, Meseta La Cruz. S 62°12.360′, W 58°57.239′; 49 m asl. | >5000 a | M-0019710/140109 | B1668 |
| H. lugubris | Antarctica, Livingston Island, Byers Peninsula. S 62°38.345′, W 61°05.380′; 94 m asl./LI_By-C | <5000 a | M-0019716/140129c | B1584 |
| H. lugubris | Antarctica, Livingston Island, Byers Peninsula. S 62°38.345′, W 61°05.380′; 94 m asl./LI_By-C | <5000 a | M-0019711 | B1669 |
| Placopsis antarctica | Antarctica, Deception Island. S 62°59.211′, W 60°40.534′; 100 m asl./DI-SW | M-0019706/ Dec 15/05aI | B1664 | |
| P. antarctica | Antarctica, Deception Island. S 62°59.211′, W 60°40.534′; 100 m asl./DI-SW | M-0019704/ Dec 15/05aVI | B1661 | |
| P. antarctica | Antarctica, Deception Island. S 62°59.211′, W 60°40.534′; 100 m asl./DI-SW | M-0019673/ Dec 15/05cV | B1618 | |
| P. antarctica | Antarctica, Deception Island. S 62°57.945′, W 60°42.912′; 20 m asl./DI-NW | M-0019700/ Dec 15/07aIV | B1645 | |
| P. antarctica | Antarctica, Deception Island. S 62°57.945′, W 60°42.912′; 20 m asl./DI-NW | M-0019701/ Dec 15/07bV | B1646 | |
| P. antarctica | Antarctica, Deception Island. S 62°57.945′, W 60°42.912′; 20 m asl./DI-NW | M-0019664/ Dec 15/07cII | B1609 | |
| P. antarctica | Antarctica, Deception Island. S 62°56.330′, W 60°35.648′; 30 m asl./DI-NE | M-0019698/ Dec 15/08bII | B1643 | |
| P. antarctica | Antarctica, Deception Island. S 62°56.330′, W 60°35.648′; 30 m asl./DI-NE | M-0019699/ Dec 15/08cI | B1644 | |
| P. antarctica | Antarctica, Deception Island. S 62°59.309′, W 60°32.915′; 120 m asl./DI-SE | M-0019672/ DEC 15/04bII | B1617 | |
| P. antarctica | Antarctica, Deception Island. S 62°59.312′, W 60°32.928′; 115 m asl./DI-SE | M-0019730/ 150202a4_DecWB | B1598 | |
| P. antarctica | Antarctica, King George Island, Ardley. S 62°12.645′, W 58°56.139′; 33 m asl./KGI-Ar | >5000 a | M-0019686/ ARD 15/03bV | B1631 |
| P. antarctica | Antarctica, King George Island, Ardley. S 62°12.645′, W 58°56.139′; 33 m asl./KGI-Ar | >5000 a | M-0019687/ ARD 15/03aIII | B1632 |
| P. antarctica | Antarctica, King George Island, Ardley. S 62°12.645′, W 58°56.139′; 33 m asl./KGI-Ar | >5000 a | M-0019655/ ARD 15/03bVI | B1600 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula, Co Gemelos. S 62°11.780′, W 58°59.602′; 33 m asl./KGI-Fi-C | >5000 a | M-0019685/ LGEM 15/01aIII | B1630 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula, Co Gemelos. S 62°11.780′, W 58°59.602′; 33 m asl./KGI-Fi-C | >5000 a | M-0019658/ LGEM 15/01cVII | B1603 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula, Co Gemelos. S 62°11.780′, W 58°59.602′; 33 m asl./KGI-Fi-C | >5000 a | M-0019659/ LGEM 15/01dIV | B1604 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula, Collins/Green Point. S 62°10.164′, W 58°51.373′; 79 m asl./KGI-Fi-NE | >5000 a | M-0019693/ 140118d_GreP | B1638 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°13.382′, W 59°01.137′; 49 m asl./KGI-Fi-S | >5000 a | M-0019689/ FiSo 15/01bIII | B1634 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°13.382′, W 59°01.137′; 49 m asl./KGI-Fi-S | >5000 a | M-0019690/ FiSo 15/01dIV | B1635 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°13.382′, W 59°01.137′; 49 m asl./KGI-Fi-S | >5000 a | M-0019663/ FiSo 15/01aV | B1608 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula, Meseta La Cruz. S 62°12.360′, W 58°57.239′; 49 m asl./KGI-Fi-C | >5000 a | M-0019656/ MCRU 15/01aII | B1601 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°10.026′, W 58°56.667′; 42 m asl./KGI-Fi-NW | <5000 a | M-0019688/ 140115a_VALK | B1633 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°12.811′, W 59°00.567′; 197 m asl./KGI-Fi-S | >5000 a | M-0019676/ 140121b_ProSch | B1621 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°13.733′, W 58°58.385′; 47 m asl./KGI-Fi-S | >5000 a | M-0019682/ FiSo 15/03fVI | B1627 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°13.733′, W 58°58.385′; 47 m asl./KGI-Fi-S | >5000 a | M-0019691/ FiSo 15/03cVI | B1636 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°13.733′, W 58°58.385′; 47 m asl./KGI-Fi-S | >5000 a | M-0019692/ FiSo 15/03eI | B1637 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°13.733′, W 58°58.385′; 47 m asl./KGI-Fi-S | >5000 a | M-0019665/ FiSo 15/03bV | B1610 |
| P. antarctica | Antarctica, King George Island, Potter Peninsula. S 62°14.887′, W 58°40.378′; 68 m asl./KGI-Po | >5000 a | M-0019675/ 150117c_Potter | B1620 |
| P. antarctica | Antarctica, King George Island. Northern part of Fildes Peninsula. S 62°8.727′, W 58°54.341′; 45 m asl./KGI-Fi-NW | <5000 a | M-0019684/ VALK 15/02dII | B1629 |
| P. antarctica | Antarctica, King George Island. Northern part of Fildes Peninsula. S 62°8.727′, W 58°54.341′; 45 m asl./KGI-Fi-NW | <5000 a | M-0019661/ VALK 15/02bII | B1606 |
| P. antarctica | Antarctica, King George Island. Northern part of Fildes Peninsula, Green Point. S 62°10.165′, W 58°51.294′; 50 m asl./KGI-Fi-NE | <5000 a | M-0019666/ GreP 15/01eIV | B1611 |
| P. antarctica | Antarctica, King George Island. Northern part of Fildes Peninsula, Green Point. S 62°10.165′, W 58°51.294′; 50 m asl./KGI-Fi-NE | <5000 a | M-0019667/ GreP 15/01dI | B1612 |
| P. antarctica | Antarctica, King George Island. Northern part of Fildes Peninsula. S 62°10.628′, W 58°58.048′; 50 m asl./KGI-Fi-C | >5000 a | M-0019662/ VALK 15/04bI | B1607 |
| P. antarctica | Antarctica, King George Island. Potter Peninsula. S 62°14.570′, W 58°39.675′; 64 m asl./KGI-Po | >5000 a | M-0019694/ Carl 15/01cI | B1639 |
| P. antarctica | Antarctica, King George Island. Potter Peninsula. S 62°14.570′, W 58°39.675′; 64 m asl./KGI-Po | >5000 a | M-0019695/ Carl 15/01eII | B1640 |
| P. antarctica | Antarctica, King George Island. Potter Peninsula. S 62°14.570′, W 58°39.675′; 64 m asl./KGI-Po | >5000 a | M-0019668/ Carl 15/01aI | B1613 |
| P. antarctica | Antarctica, Livingston Island, Byers Peninsula. S 62°39.210′, W 61°00.444′; 60 m asl./LI_By-E | <5000 a | M-0019678/ 140128e_CeNegro | B1623 |
| P. antarctica | Antarctica, Livingston Island, Byers Peninsula. S 62°38.345′, W 61°05.380′; 94 m asl./LI_By-C | <5000 a | M-0019731/ 140129c_CeChester | B1599 |
| P. antarctica | Antarctica, Livingston Island, Byers Peninsula, Cerro Smellie. S 62°39.112′, W 61°08.761′; 15 m asl./LI-By-W | >5000 a | M-0019696/ 140130c_CeSmellie | B1641 |
| P. antarctica | Antarctica, Livingston Island, Byers Peninsula. S 62°40.065′, W 60°54.739′; 130 m asl./LI-By-E | >5000 a | M-0019697/ Byer 15/06bIII | B1642 |
| P. antarctica | Antarctica, Livingston Island, Byers Peninsula. S 62°40.065′, W 60°54.739′; 130 m asl./LI-By-E | >5000 a | M-0019671/ Byer 15/06dIII | B1616 |
| P. antarctica | Antarctica, Livingston Island, Byers Peninsula. S 62°40.144′, W 61°10.333′; 70 m asl./LI-By-W | >5000 a | M-0019670/ Byer 15/01aII | B1615 |
| P. antarctica | Antarctica, Robert Island, Coppermine Peninsula. S 62°22,311′, W 59°43.024′; 150 m asl./RI-Co | >5000 a | M-0019707/ Robe 15/01cVI | B1665 |
| P. antarctica | Antarctica, Robert Island, Coppermine Peninsula. S 62°22,311′, W 59°43.024′; 150 m asl./RI-Co | >5000 a | M-0019669/ Robe 15/01aII | B1614 |
| P. antarctica | Antarctica, King George Island, Fildes Peninsula. S 62°12.243′, W 58°57.590′; ca. 70 m asl./KGI-Fi-C | >5000 a | M-0019729/ 140109_MCRU | B1597 |
| P. contortuplicata | Antarctica, Deception Island. S 62°56.330′, W 60°35.648′; 30 m asl./DI-NE | M-0019705/ Dec 15/08aVI | B1663 | |
| P. contortuplicata | Antarctica, King George Island, Fildes Peninsula. S 62°12.360′, W 58°57.239′; 49 m asl./KGI-Fi-C | >5000 a | M-0019657/ MCRU 15/01cII | B1602 |
| P. contortuplicata | Antarctica, King George Island, Fildes Peninsula. S 62°12.360′, W 58°57.239′; 49 m asl./KGI-Fi-C | >5000 a | M-0019679/ MCRU 15/01bV | B1624 |
| P. contortuplicata | Antarctica, King George Island. Northern part of Fildes Peninsula. S 62°11.780′, W 58°59.602′; 33 m asl./KGI-Fi-NW | <5000 a | M-0019660/ VALK 15/01cVI | B1605 |
| P. contortuplicata | Antarctica, King George Island. Northern part of Fildes Peninsula. S 62°11.780′, W 58°59.602′; 33 m asl./KGI-Fi-NW | <5000 a | M-0019680/ VALK 15/01aIII | B1625 |
| P. contortuplicata | Antarctica, Livingston Island, Byers Peninsula. S 62°40.065′, W 60°54.739′; 130 m asl./LI-By-E | >5000 a | M-0019683/ Byer 15/06fIII | B1628 |
| P. contortuplicata | Antarctica, Livingston Island, Byers Peninsula. S 62°40.220′, W 61°06.776′; 73 m asl./LI-By-C | >5000 a | M-0019677/ 140127d_ByCeCol | B1622 |
| P. contortuplicata | Antarctica, King George Island. Northern part of Fildes Peninsula. S 62°11.780′, W 58°59.602′; 33 m asl./KGI-Fi-NW | <5000 a | M-0019681/ VALK 15/01eI | B1626 |
| Ramalina terebrata | Antarctica, Deception Island. S 62°59.272′, W 60°33.006′; 59 m asl. | M-0019724/140124e | B1592 | |
| R. terebrata | Antarctica, King George Island, Ardley. S 62°12.675′, W 58°55.461′; 18 m asl. | >5000 a | M-0019728/140116d | B1596 |
| R. terebrata | Antarctica, King George Island, Ardley. S 62°12.55′, W 58°56.416′; 25 m asl. | >5000 a | M-0019727/140116f | B1595 |
| R. terebrata | Antarctica, King George Island. Fildes Peninsula, Nebles Point. S 62°11.033′, W 58°51.544′; 9 m asl. | >5000 a | M-0019725/140118c | B1593 |
| R. terebrata | Antarctica, King George Island, Potter Peninsula. S 62°14.834′, W 58°40.760′; 20 m asl. | >5000 a | M-0019726/140117b2 | B1594 |
| R. terebrata | Antarctica, Livingston Island, Byers peninsula, Playa los Presidentes. S 62°39.325′, W 61°08.792′; 2 m asl. | >5000 a | M-0019722/140130d | B1590 |
| R. terebrata | Antarctica, Livingston Island, Byers peninsula, Punta Petreles. S 62°40.496′, W 61°05.807′; 13 m asl. | >5000 a | M-0019723/140127a | B1591 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beck, A.; Casanova-Katny, A.; Gerasimova, J. Metabarcoding of Antarctic Lichens from Areas with Different Deglaciation Times Reveals a High Diversity of Lichen-Associated Communities. Genes 2023, 14, 1019. https://doi.org/10.3390/genes14051019
Beck A, Casanova-Katny A, Gerasimova J. Metabarcoding of Antarctic Lichens from Areas with Different Deglaciation Times Reveals a High Diversity of Lichen-Associated Communities. Genes. 2023; 14(5):1019. https://doi.org/10.3390/genes14051019
Chicago/Turabian StyleBeck, Andreas, Angélica Casanova-Katny, and Julia Gerasimova. 2023. "Metabarcoding of Antarctic Lichens from Areas with Different Deglaciation Times Reveals a High Diversity of Lichen-Associated Communities" Genes 14, no. 5: 1019. https://doi.org/10.3390/genes14051019
APA StyleBeck, A., Casanova-Katny, A., & Gerasimova, J. (2023). Metabarcoding of Antarctic Lichens from Areas with Different Deglaciation Times Reveals a High Diversity of Lichen-Associated Communities. Genes, 14(5), 1019. https://doi.org/10.3390/genes14051019