
The Potential Regulation of A-to-I RNA Editing on Genes in Parkinson’s Disease

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
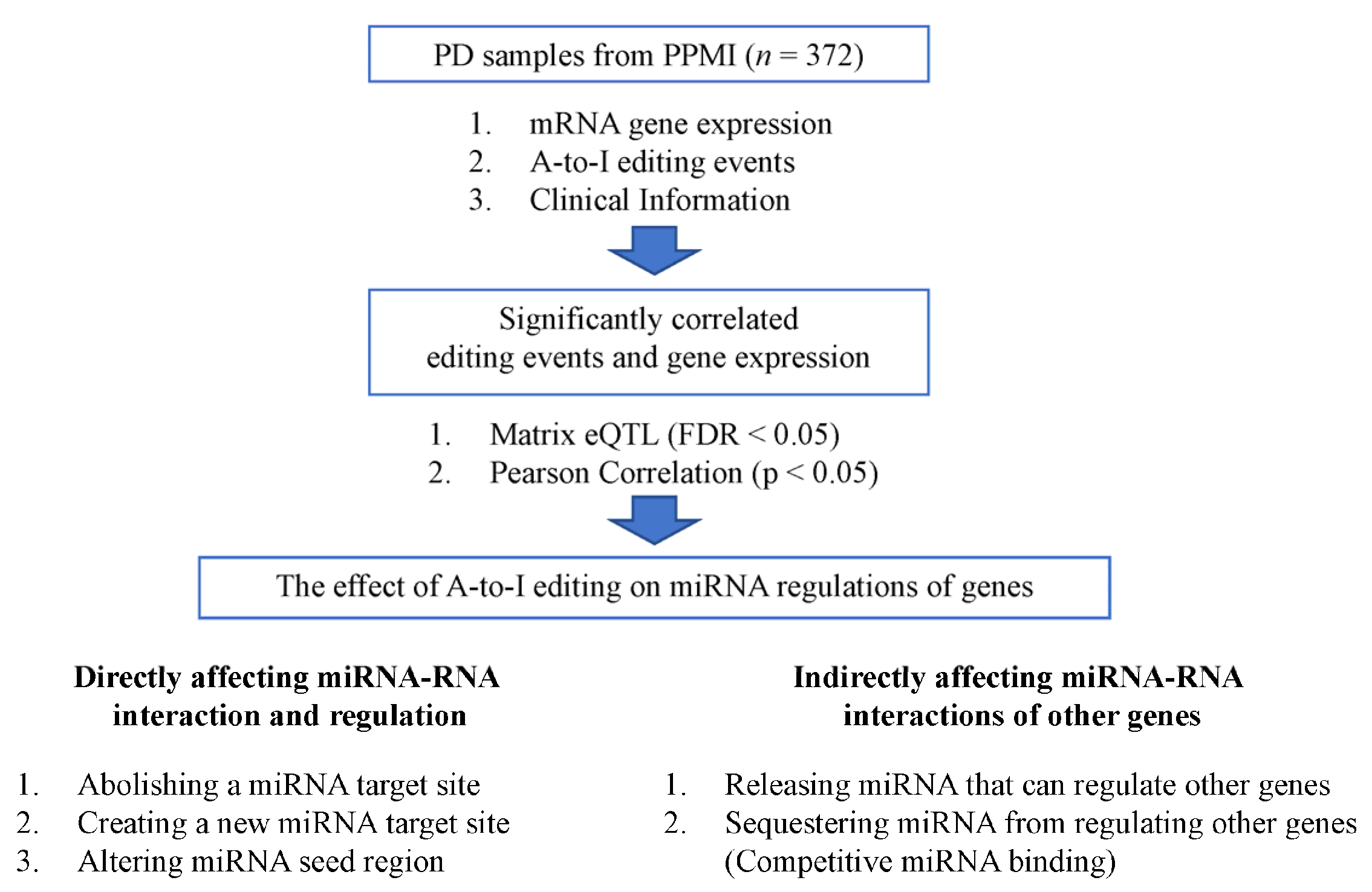
2. Materials and Methods
2.1. Samples Involved in This Study
2.2. A-to-I RNA Editing Detection
2.3. Gene Expression Quantification
2.4. Correlation Analysis between A-to-I RNA Editing Events and Genes
2.5. Analysis of RNA Editing Effects on Genes via Potential miRNA Regulation Mechanisms
3. Results
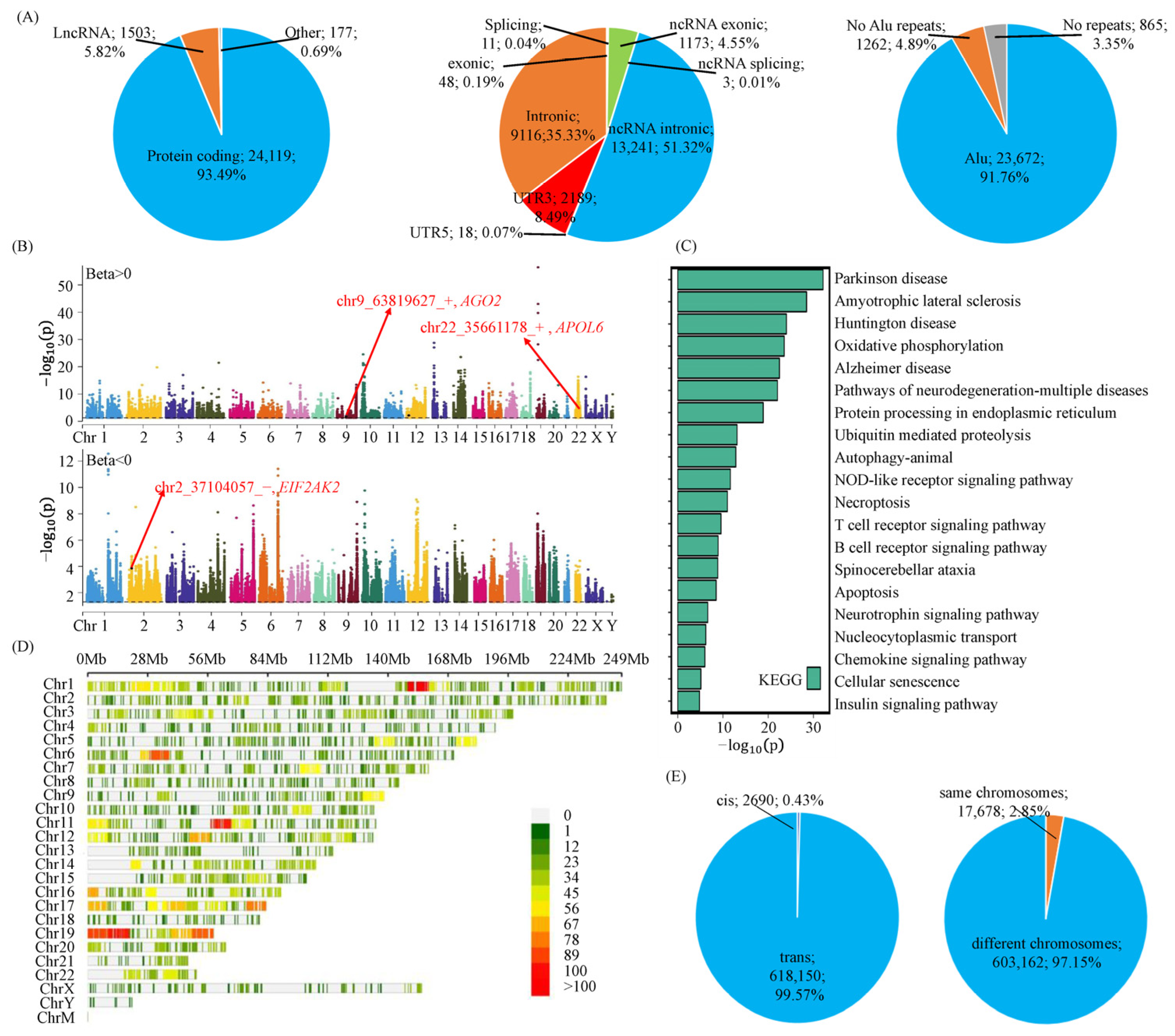
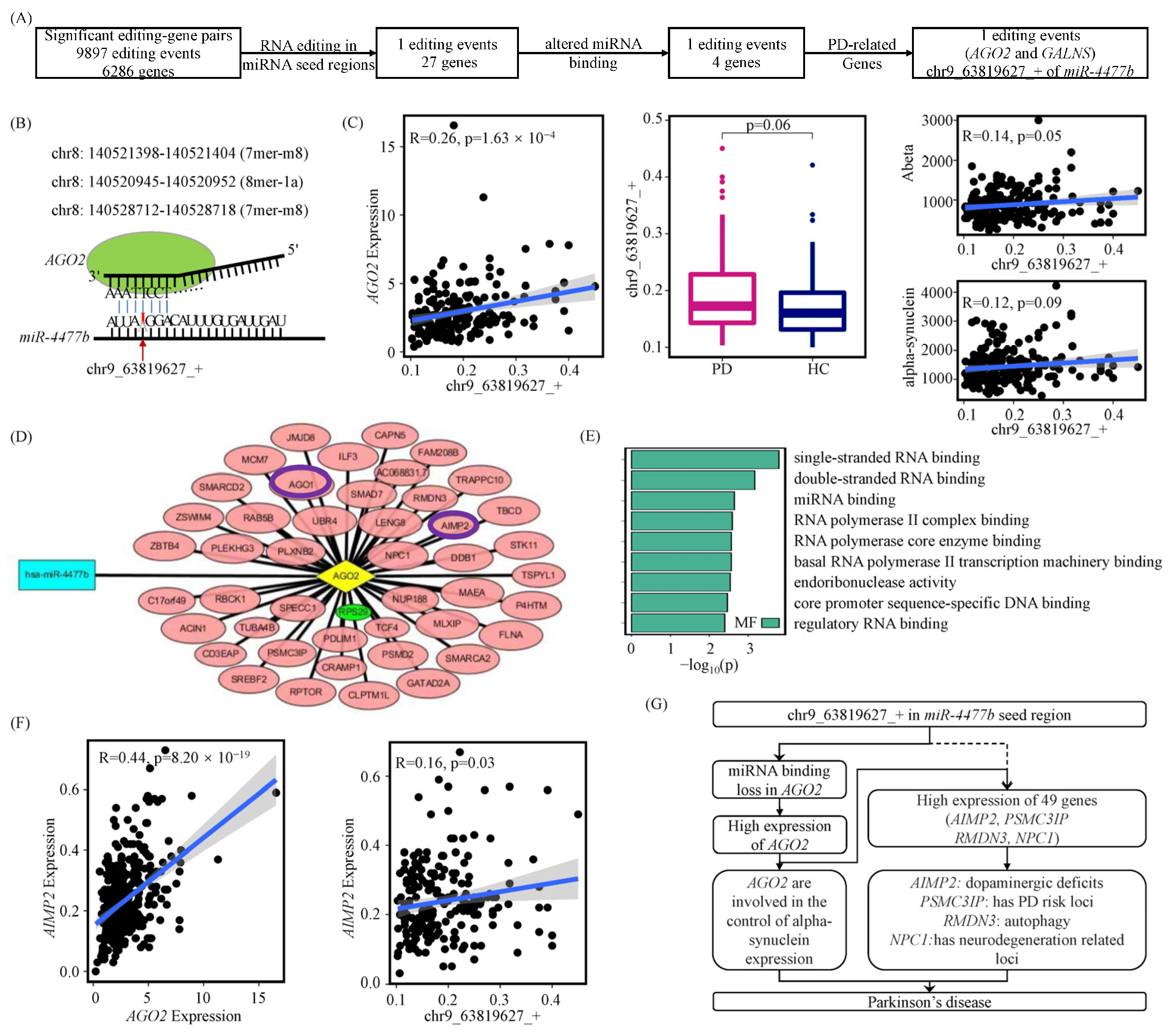
3.1. A-to-I RNA Editing Events Are Involved in Parkinson’s Disease via Their Effects on Gene Expressions
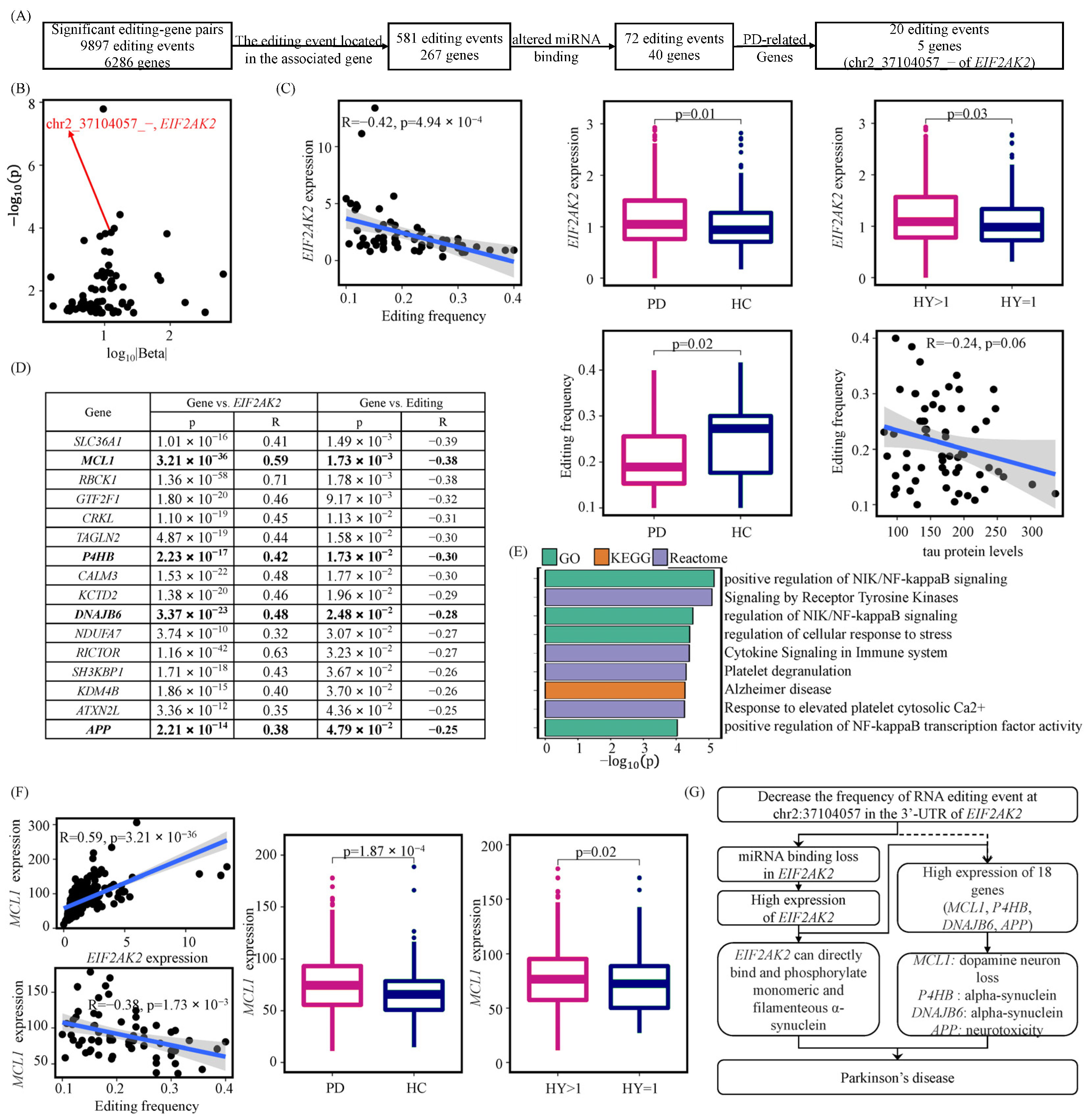
3.2. A-to-I RNA Editing Events May Affect miRNA Regulations of Their Host Genes
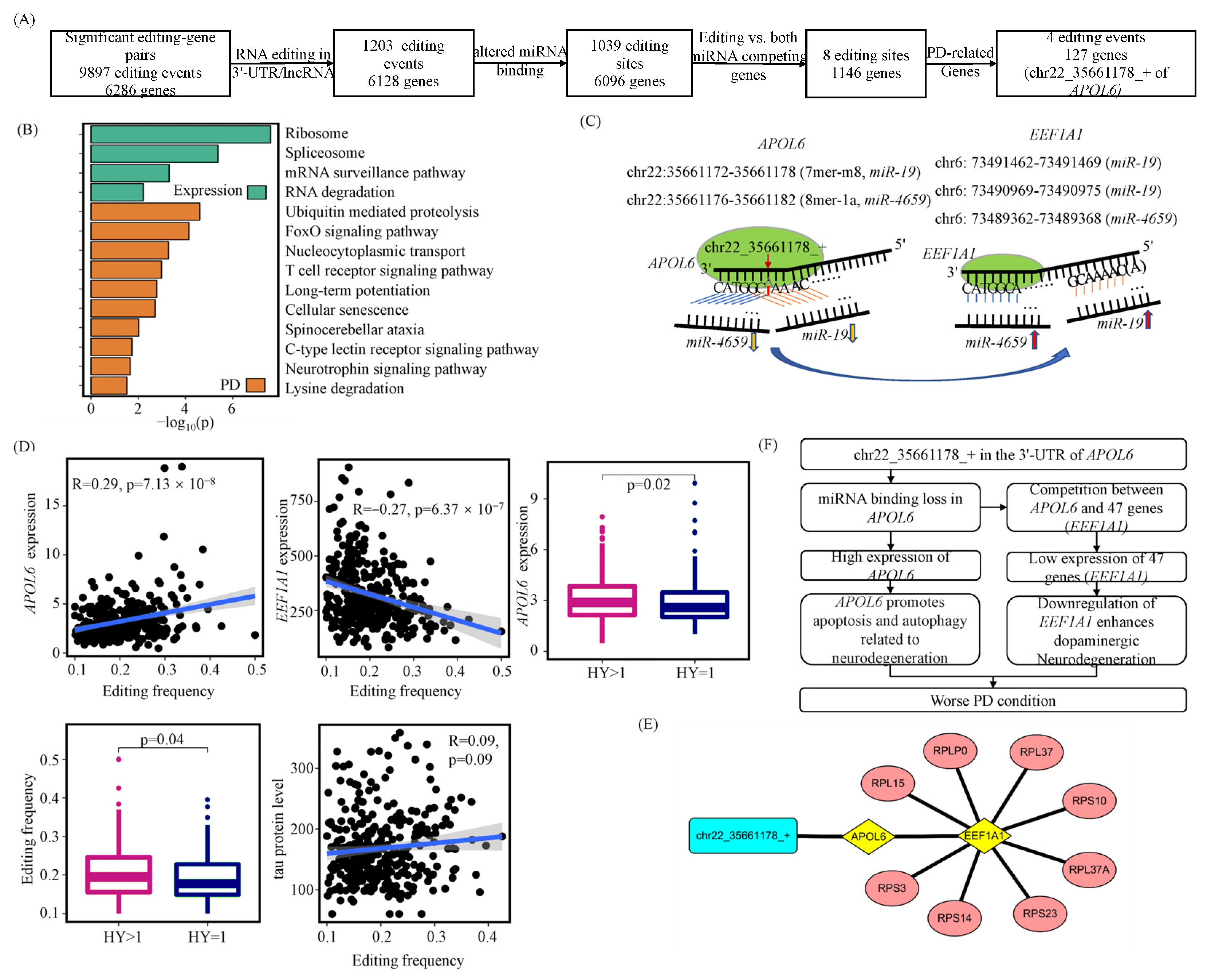
3.3. A-to-I RNA Editing Events May Alter miRNA Competitions between Their Host Genes and Other Genes
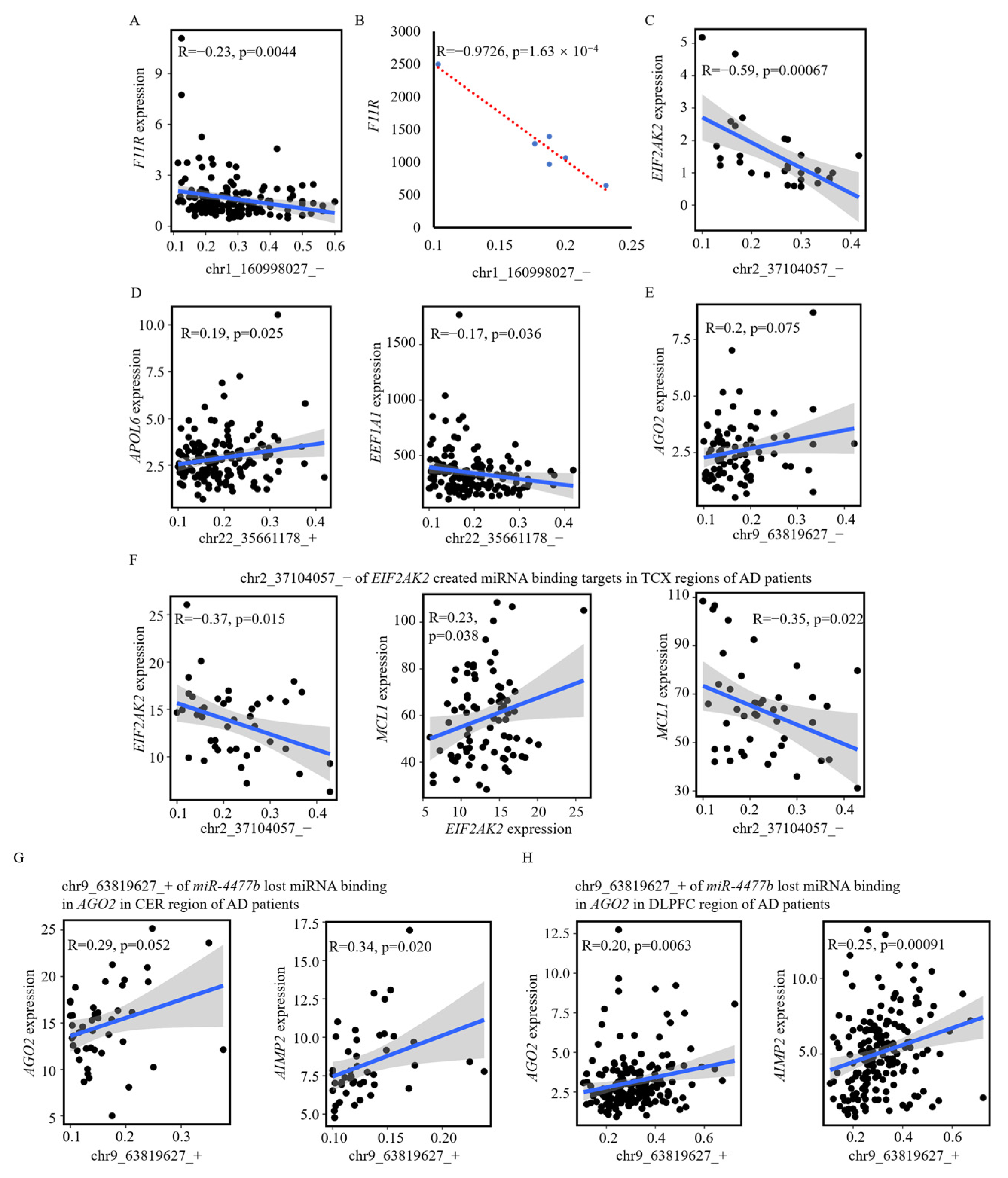
3.4. A-to-I RNA Editing Events May Modify miRNA Seed Regions to Disturb Their Regulations
3.5. A-to-I RNA Editing Effects on Genes via Disturbing miRNA Regulations in Other Datasets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef] [PubMed]
- Aarsland, D.; Batzu, L.; Halliday, G.M.; Geurtsen, G.J.; Ballard, C.; Ray Chaudhuri, K.; Weintraub, D. Parkinson disease-associated cognitive impairment. Nat. Rev. Dis. Prim. 2021, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, M.J.; Okun, M.S. Diagnosis and treatment of Parkinson disease: A review. JAMA 2020, 323, 548–560. [Google Scholar] [CrossRef]
- Schapira, A.H.; Chaudhuri, K.; Jenner, P. Non-motor features of Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 435–450. [Google Scholar] [CrossRef]
- Dionísio, P.; Amaral, J.; Rodrigues, C. Oxidative stress and regulated cell death in Parkinson’s disease. Ageing Res. Rev. 2021, 67, 101263. [Google Scholar] [CrossRef] [PubMed]
- Tansey, M.G.; Wallings, R.L.; Houser, M.C.; Herrick, M.K.; Keating, C.E.; Joers, V. Inflammation and immune dysfunction in Parkinson disease. Nat. Rev. Immunol. 2022, 22, 657–673. [Google Scholar] [CrossRef] [PubMed]
- Malpartida, A.B.; Williamson, M.; Narendra, D.P.; Wade-Martins, R.; Ryan, B.J. Mitochondrial Dysfunction and Mitophagy in Parkinson’s Disease: From Mechanism to Therapy. Trends Biochem. Sci. 2021, 46, 329–343. [Google Scholar] [CrossRef]
- Hindle, J.V. Ageing, neurodegeneration and Parkinson’s disease. Age Ageing 2010, 39, 156–161. [Google Scholar] [CrossRef]
- Marras, C.; Canning, C.G.; Goldman, S.M. Environment, lifestyle, and Parkinson’s disease: Implications for prevention in the next decade. Mov. Disord. 2019, 34, 801–811. [Google Scholar] [CrossRef]
- Polymeropoulos, M.H.; Lavedan, C.; Leroy, E.; Ide, S.E.; Dehejia, A.; Dutra, A.; Pike, B.; Root, H.; Rubenstein, J.; Boyer, R. Mutation in the α-synuclein gene identified in families with Parkinson’s disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef]
- Mata, I.F.; Shi, M.; Agarwal, P.; Chung, K.A.; Edwards, K.L.; Factor, S.A.; Galasko, D.R.; Ginghina, C.; Griffith, A.; Higgins, D.S. SNCA Variant Associated with Parkinson Disease and Plasma α-Synuclein Level. Arch. Neurol. 2010, 67, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, S.M.; Tikhonova, E.B.; Baca, K.R.; Zhao, F.; Zhu, X.; Karamyshev, A.L. Unexpected Implication of SRP and AGO2 in Parkinson’s Disease: Involvement in Alpha-Synuclein Biogenesis. Cells 2021, 10, 2792. [Google Scholar] [CrossRef] [PubMed]
- Reimer, L.; Vesterager, L.B.; Betzer, C.; Zheng, J.; Nielsen, L.D.; Kofoed, R.H.; Lassen, L.B.; Bølcho, U.; Paludan, S.R.; Fog, K. Inflammation kinase PKR phosphorylates α-synuclein and causes α-synuclein-dependent cell death. Neurobiol. Dis. 2018, 115, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Reimer, L.; Gram, H.; Jensen, N.M.; Betzer, C.; Yang, L.; Jin, L.; Shi, M.; Boudeffa, D.; Fusco, G.; De Simone, A.; et al. Protein kinase R dependent phosphorylation of α-synuclein regulates its membrane binding and aggregation. PNAS Nexus 2022, 1, pgac259. [Google Scholar] [CrossRef]
- Chalorak, P.; Dharmasaroja, P.; Meemon, K. Downregulation of eEF1A/EFT3-4 Enhances Dopaminergic Neurodegeneration After 6-OHDA Exposure in C. elegans Model. Front. Neurosci. 2020, 14, 303. [Google Scholar] [CrossRef]
- Robinson, E.J.; Aguiar, S.; Smidt, M.P.; Van der Heide, L.P. MCL1 as a Therapeutic Target in Parkinson’s Disease? Trends Mol. Med. 2019, 25, 1056–1065. [Google Scholar] [CrossRef]
- Aman, Y. Selective loss among dopaminergic neurons in Parkinson’s disease. Nat. Aging 2022, 2, 462. [Google Scholar] [CrossRef]
- Lee, Y.; Karuppagounder, S.S.; Shin, J.-H.; Lee, Y.-I.; Ko, H.S.; Swing, D.; Jiang, H.; Kang, S.-U.; Lee, B.D.; Kang, H.C.; et al. Parthanatos mediates AIMP2-activated age-dependent dopaminergic neuronal loss. Nat. Neurosci. 2013, 16, 1392–1400. [Google Scholar] [CrossRef]
- Ygland Rödström, E.; Mattsson-Carlgren, N.; Janelidze, S.; Hansson, O.; Puschmann, A. Serum Neurofilament Light Chain as a Marker of Progression in Parkinson’s Disease: Long-Term Observation and Implications of Clinical Subtypes. J. Park. Dis. 2022, 12, 571–584. [Google Scholar] [CrossRef]
- Kern, F.; Fehlmann, T.; Violich, I.; Alsop, E.; Hutchins, E.; Kahraman, M.; Grammes, N.L.; Guimarães, P.; Backes, C.; Poston, K.L.; et al. Deep sequencing of sncRNAs reveals hallmarks and regulatory modules of the transcriptome during Parkinson’s disease progression. Nat. Aging 2021, 1, 309–322. [Google Scholar] [CrossRef]
- Karpenko, M.N.; Vasilishina, A.A.; Gromova, E.A.; Muruzheva, Z.M.; Miliukhina, I.V.; Bernadotte, A. Interleukin-1β, interleukin-1 receptor antagonist, interleukin-6, interleukin-10, and tumor necrosis factor-α levels in CSF and serum in relation to the clinical diversity of Parkinson’s disease. Cell. Immunol. 2018, 327, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, L.; Yang, Y.; Zhang, M.; Wong, G. Identification of potential blood biomarkers for Parkinson’s disease by gene expression and DNA methylation data integration analysis. Clin. Epigenetics 2019, 11, 24. [Google Scholar] [CrossRef]
- Zhang, W.; Shen, J.; Wang, Y.; Cai, K.; Zhang, Q.; Cao, M. Blood SSR1: A Possible Biomarker for Early Prediction of Parkinson’s Disease. Front. Mol. Neurosci. 2022, 15, 762544. [Google Scholar] [CrossRef]
- Gui, Y.; Liu, H.; Zhang, L.; Lv, W.; Hu, X. Altered microRNA profiles in cerebrospinal fluid exosome in Parkinson disease and Alzheimer disease. Oncotarget 2015, 6, 37043–37053. [Google Scholar] [CrossRef] [PubMed]
- Marek, K.; Jennings, D.; Lasch, S.; Siderowf, A.; Tanner, C.; Simuni, T.; Coffey, C.; Kieburtz, K.; Flagg, E.; Chowdhury, S. The Parkinson Progression Marker Initiative (PPMI). Prog. Neurobiol. 2011, 95, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Marek, K.; Chowdhury, S.; Siderowf, A.; Lasch, S.; Coffey, C.S.; Caspell-Garcia, C.; Simuni, T.; Jennings, D.; Tanner, C.M.; Trojanowski, J.Q. The Parkinson’s progression markers initiative (PPMI)—Establishing a PD biomarker cohort. Ann. Clin. Transl. Neurol. 2018, 5, 1460–1477. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. A-to-I editing of coding and non-coding RNAs by ADARs. Nat. Rev. Mol. Cell Biol. 2016, 17, 83–96. [Google Scholar] [CrossRef]
- Eisenberg, E.; Levanon, E.Y. A-to-I RNA editing—Immune protector and transcriptome diversifier. Nat. Rev. Genet. 2018, 19, 473–490. [Google Scholar] [CrossRef]
- Konen, L.M.; Wright, A.L.; Royle, G.A.; Morris, G.P.; Lau, B.K.; Seow, P.W.; Zinn, R.; Milham, L.T.; Vaughan, C.W.; Vissel, B. A new mouse line with reduced GluA2 Q/R site RNA editing exhibits loss of dendritic spines, hippocampal CA1-neuron loss, learning and memory impairments and NMDA receptor-independent seizure vulnerability. Mol. Brain 2020, 13, 27. [Google Scholar] [CrossRef]
- Pozdyshev, D.V.; Zharikova, A.A.; Medvedeva, M.V.; Muronetz, V.I. Differential Analysis of A-to-I mRNA Edited Sites in Parkinson’s Disease. Genes 2021, 13, 14. [Google Scholar] [CrossRef]
- Costa Cruz, P.H.; Kawahara, Y. RNA Editing in Neurological and Neurodegenerative Disorders. RNA Ed. 2021, 2181, 309–330. [Google Scholar] [CrossRef]
- Thele, A.; Gardner, O.; Vance, J.; Griswold, A.; Young, J.; Pericak-Vance, M.; Scott, W.; Martin, E.; Wang, W.; Dykxhoorn, D. RNA Editing in Parkinson’s Disease Shows Changes in Mitochondrial and Manganese-Binding Pathways in Multiple Brain Regions. Mov. Disord. 2021, 36, S342–S343. [Google Scholar]
- Wu, S.; Yang, M.; Kim, P.; Zhou, X. ADeditome provides the genomic landscape of A-to-I RNA editing in Alzheimer’s disease. Briefings Bioinform. 2021, 22, bbaa384. [Google Scholar] [CrossRef] [PubMed]
- Brümmer, A.; Yang, Y.; Chan, T.W.; Xiao, X. Structure-mediated modulation of mRNA abundance by A-to-I editing. Nat. Commun. 2017, 8, 1255. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, Z. MicroRNAs: Game changers in the regulation of α-synuclein in Parkinson’s disease. Park. Dis. 2019, 2019, 1743183. [Google Scholar] [CrossRef] [PubMed]
- Goetz, C.G.; Poewe, W.; Rascol, O.; Sampaio, C.; Stebbins, G.T.; Counsell, C.; Giladi, N.; Holloway, R.G.; Moore, C.G.; Wenning, G.K. Movement Disorder Society Task Force report on the Hoehn and Yahr staging scale: Status and recommendations the Movement Disorder Society Task Force on rating scales for Parkinson’s disease. Mov. Disord. 2004, 19, 1020–1028. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.W.; Aarsland, D.; Ffytche, D.; Taddei, R.N.; van Wamelen, D.J.; Wan, Y.-M.; Tan, E.K.; Ray Chaudhuri, K. Amyloid-β and Parkinson’s disease. J. Neurol. 2019, 266, 2605–2619. [Google Scholar] [CrossRef]
- Pan, L.; Meng, L.; He, M.; Zhang, Z. Tau in the Pathophysiology of Parkinson’s Disease. J. Mol. Neurosci. 2021, 71, 2179–2191. [Google Scholar] [CrossRef]
- Makkos, A.; Kovács, M.; Aschermann, Z.; Harmat, M.; Janszky, J.; Karádi, K.; Kovács, N. Are the MDS-UPDRS–Based Composite Scores Clinically Applicable? Mov. Disord. 2018, 33, 835–839. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Picardi, E.; Pesole, G. REDItools: High-throughput RNA editing detection made easy. Bioinformatics 2013, 29, 1813–1814. [Google Scholar] [CrossRef] [PubMed]
- Mansi, L.; Tangaro, M.A.; Lo Giudice, C.; Flati, T.; Kopel, E.; Schaffer, A.A.; Castrignanò, T.; Chillemi, G.; Pesole, G.; Picardi, E. REDIportal: Millions of novel A-to-I RNA editing events from thousands of RNAseq experiments. Nucleic Acids Res. 2021, 49, D1012–D1019. [Google Scholar] [CrossRef] [PubMed]
- Sherry, S.T.; Ward, M.-H.; Kholodov, M.; Baker, J.; Phan, L.; Smigielski, E.M.; Sirotkin, K. dbSNP: The NCBI database of genetic variation. Nucleic Acids Res. 2001, 29, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, M.; Hakonarson, H. ANNOVAR: Functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010, 38, e164. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2019, 48, D845–D855. [Google Scholar] [CrossRef] [PubMed]
- Rappaport, N.; Twik, M.; Plaschkes, I.; Nudel, R.; Iny Stein, T.; Levitt, J.; Gershoni, M.; Morrey, C.P.; Safran, M.; Lancet, D. MalaCards: An amalgamated human disease compendium with diverse clinical and genetic annotation and structured search. Nucleic Acids Res. 2016, 45, D877–D887. [Google Scholar] [CrossRef]
- Yu, W.; Clyne, M.; Khoury, M.J.; Gwinn, M. Phenopedia and Genopedia: Disease-centered and gene-centered views of the evolving knowledge of human genetic associations. Bioinformatics 2010, 26, 145–146. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef]
- Shabalin, A.A. Matrix eQTL: Ultra fast eQTL analysis via large matrix operations. Bioinformatics 2012, 28, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.M.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The biochemical basis of microRNA targeting efficacy. Science 2019, 366, 6472. [Google Scholar] [CrossRef] [PubMed]
- Enright, A.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D. MicroRNA targets in Drosophila. Genome Biol. 2003, 4, P8. [Google Scholar] [CrossRef]
- Li, J.-H.; Liu, S.; Zhou, H.; Qu, L.-H.; Yang, J.-H. starBase v2. 0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein–RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Chigaev, M.; Yu, H.; Samuels, D.C.; Sheng, Q.; Oyebamiji, O.; Ness, S.; Yue, W.; Zhao, Y.-Y.; Guo, Y. Genomic positional dissection of RNA editomes in tumor and normal samples. Front. Genet. 2019, 10, 211. [Google Scholar] [CrossRef]
- Gerstberger, S.; Hafner, M.; Tuschl, T. A census of human RNA-binding proteins. Nat. Rev. Genet. 2014, 15, 829–845. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, T.; Cui, T.; Wang, Z.; Zhang, Y.; Tan, P.; Huang, Y.; Yu, J.; Wang, D. RNAInter in 2020: RNA interactome repository with increased coverage and annotation. Nucleic Acids Res. 2020, 48, D189–D197. [Google Scholar] [CrossRef]
- Lang, B.; Armaos, A.; Tartaglia, G.G. RNAct: Protein–RNA interaction predictions for model organisms with supporting experimental data. Nucleic Acids Res. 2019, 47, D601–D606. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, T.G. Role of nuclear factor kappa B (NF-κB) signalling in neurodegenerative diseases: An mechanistic approach. Curr. Neuropharm. 2020, 18, 918–935. [Google Scholar] [CrossRef] [PubMed]
- Medinas, D.B.; Rozas, P.; Hetz, C. Critical roles of protein disulfide isomerases in balancing proteostasis in the nervous system. J. Biol. Chem. 2022, 298, 102087. [Google Scholar] [CrossRef] [PubMed]
- Arkan, S.; Ljungberg, M.; Kirik, D.; Hansen, C. DNAJB6 suppresses alpha-synuclein induced pathology in an animal model of Parkinson’s disease. Neurobiol. Dis. 2021, 158, 105477. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-C.; Zhang, W.; Chua, L.-L.; Chai, C.; Li, R.; Lin, L.; Cao, Z.; Angeles, D.C.; Stanton, L.W.; Peng, J.-H. Phosphorylation of amyloid precursor protein by mutant LRRK2 promotes AICD activity and neurotoxicity in Parkinson’s disease. Sci. Signal. 2017, 10, 488. [Google Scholar] [CrossRef]
- Zhaorigetu, S.; Yang, Z.; Toma, I.; McCaffrey, T.A.; Hu, C.-A.A. Apolipoprotein L6, Induced in Atherosclerotic Lesions, Promotes Apoptosis and Blocks Beclin 1-dependent Autophagy in Atherosclerotic Cells. J. Biol. Chem. 2011, 286, 27389–27398. [Google Scholar] [CrossRef]
- Liu, Z.; Lu, H.; Jiang, Z.; Pastuszyn, A.; Hu, C.-A.A. Apolipoprotein L6, a Novel Proapoptotic Bcl-2 Homology 3–Only Protein, Induces Mitochondria-Mediated Apoptosis in Cancer Cells. Mol. Cancer Res. 2005, 3, 21–31. [Google Scholar] [CrossRef]
- Stoker, T.B.; Greenland, J.C. Parkinson’s Disease: Pathogenesis and Clinical Aspects; Codon Publications: Brisbane, Australia, 2018. [Google Scholar]
- Chang, D.; Nalls, M.A.; Hallgrímsdóttir, I.B.; Hunkapiller, J.; Van Der Brug, M.; Cai, F.; Kerchner, G.A.; Ayalon, G.; Bingol, B.; Sheng, M. A meta-analysis of genome-wide association studies identifies 17 new Parkinson’s disease risk loci. Nat. Genet. 2017, 49, 1511–1516. [Google Scholar] [CrossRef]
- Kluenemann, H.H.; Nutt, J.G.; Davis, M.Y.; Bird, T.D. Parkinsonism syndrome in heterozygotes for Niemann–Pick C1. J. Neurol. Sci. 2013, 335, 219–220. [Google Scholar] [CrossRef]
- Gomez-Suaga, P.; Paillusson, S.; Miller, C.C. ER-mitochondria signaling regulates autophagy. Autophagy 2017, 13, 1250–1251. [Google Scholar] [CrossRef]
- Lu, C.; Ren, S.; Zhao, Z.; Wu, X.; Suo, A.; Zhou, N.; Yang, J.; Wu, S.; Li, T.; Peng, C. Characterizing relevant microRNA editing sites in Parkinson’s disease. Cells 2020, 12, 75. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Fan, Z.; Kim, P.; Huang, L.; Zhou, X. The Integrative Studies on the Functional A-to-I RNA Editing Events in Human Cancers. Genom. Proteom. Bioinform. 2023; in press. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Xue, Q.; Yang, M.; Wang, Y.; Kim, P.; Huang, L. Genetic control of RNA editing in Neurodegenerative disease. bioRxiv 2022. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Xue, Q.; Qin, X.; Wu, X.; Kim, P.; Chyr, J.; Zhou, X.; Huang, L. The Potential Regulation of A-to-I RNA Editing on Genes in Parkinson’s Disease. Genes 2023, 14, 919. https://doi.org/10.3390/genes14040919
Wu S, Xue Q, Qin X, Wu X, Kim P, Chyr J, Zhou X, Huang L. The Potential Regulation of A-to-I RNA Editing on Genes in Parkinson’s Disease. Genes. 2023; 14(4):919. https://doi.org/10.3390/genes14040919
Chicago/Turabian StyleWu, Sijia, Qiuping Xue, Xinyu Qin, Xiaoming Wu, Pora Kim, Jacqueline Chyr, Xiaobo Zhou, and Liyu Huang. 2023. "The Potential Regulation of A-to-I RNA Editing on Genes in Parkinson’s Disease" Genes 14, no. 4: 919. https://doi.org/10.3390/genes14040919
APA StyleWu, S., Xue, Q., Qin, X., Wu, X., Kim, P., Chyr, J., Zhou, X., & Huang, L. (2023). The Potential Regulation of A-to-I RNA Editing on Genes in Parkinson’s Disease. Genes, 14(4), 919. https://doi.org/10.3390/genes14040919