
WNT Co-Receptor LRP6 Is Critical for Zygotic Genome Activation and Embryonic Developmental Potential by Interacting with Oviductal Paracrine Ligand WNT2

Abstract
1. Introduction
2. Results
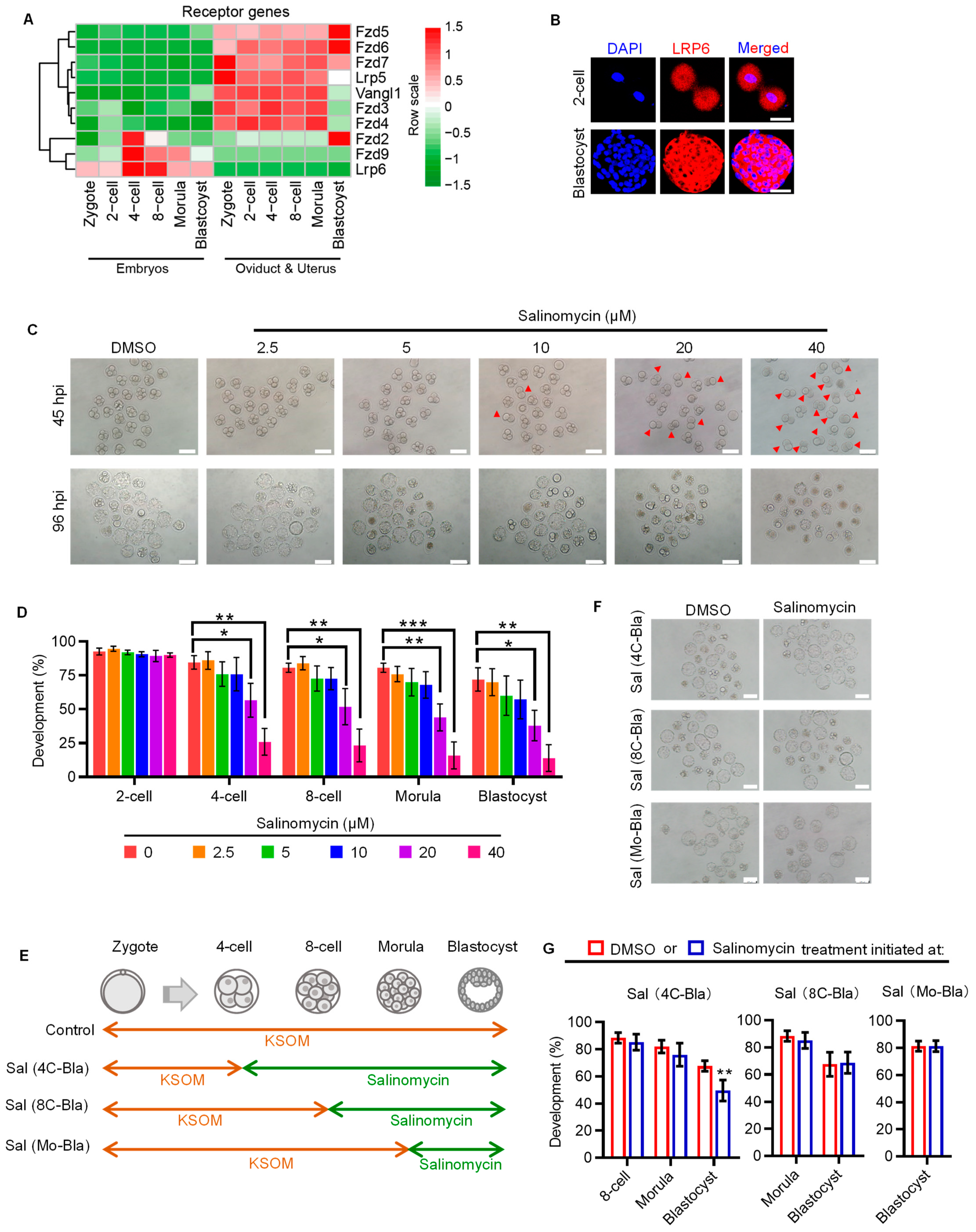
2.1. WNT Co-Receptor LRP6 Is Critical to Early Cleavage of Preimplantation Development
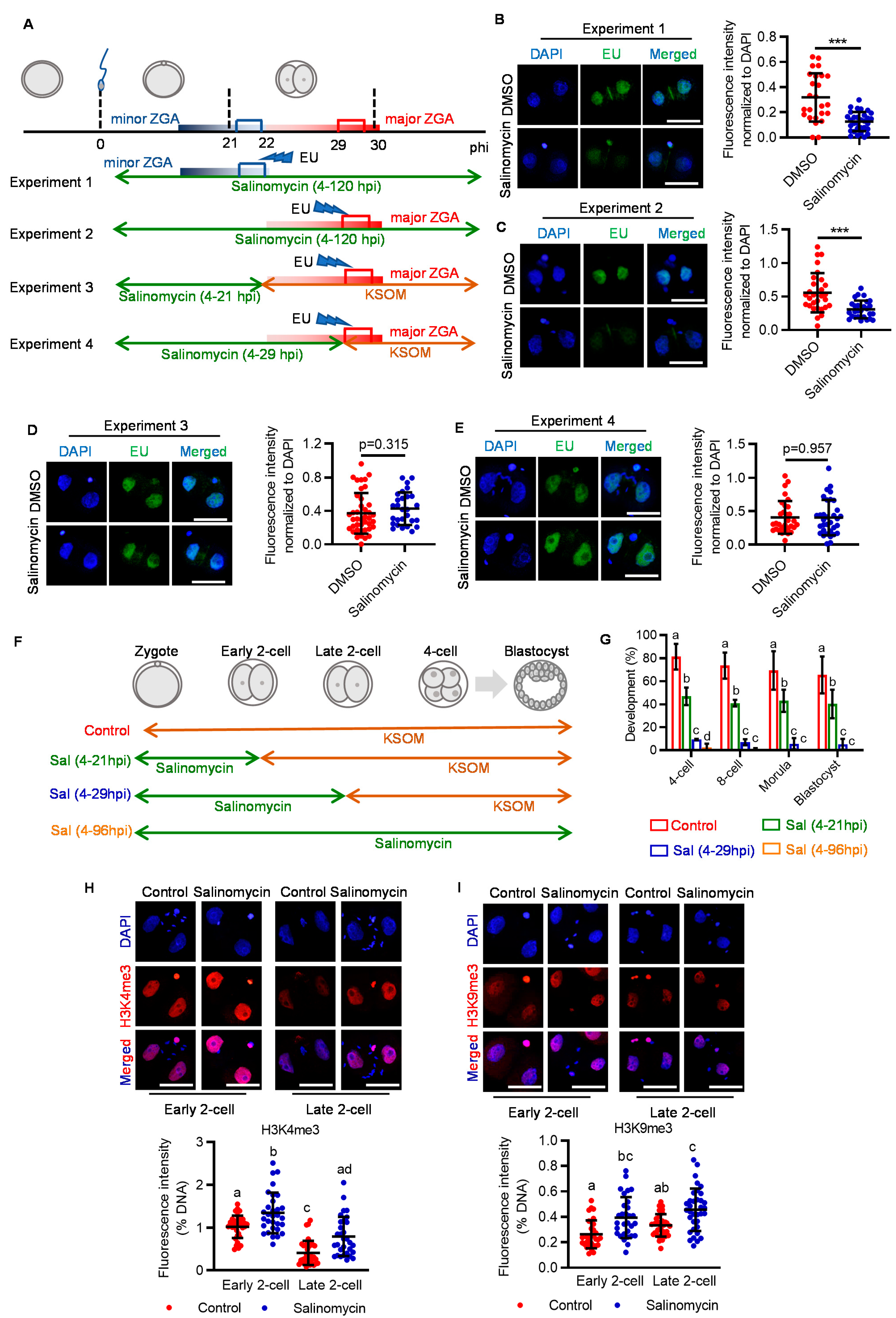
2.2. LRP6 Inhibition Impedes Zygotic Genome Activation and Disrupts Relevant Epigenetic Reprogramming
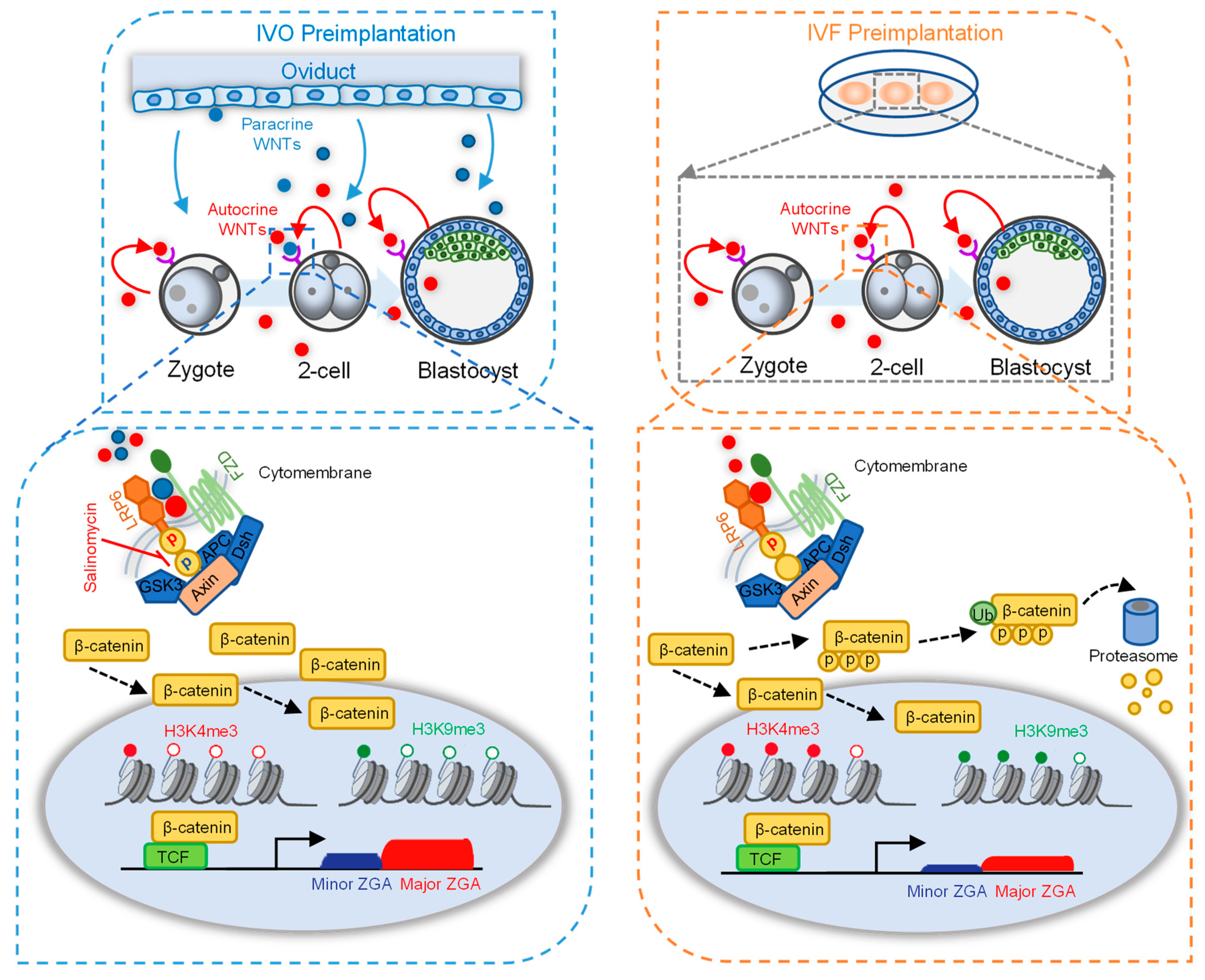
2.3. Oviductal Paracrine WNT2 Is Critical for Activating LRP6 and Prompting ZGA
2.4. Supplementation of Oviductal Paracrine WNT2 Improves Embryo Quality and Pregnancy Outcomes
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Oocyte Source, In Vitro Fertilization (IVF) and Embryo Culture
4.3. Analysis of Transcription Activity
4.4. Embryo Transfer
4.5. Blastocyst Outgrowth
4.6. Immunofluorescence Staining
4.7. GSVA and GSEA Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hardy, K.; Spanos, S. Growth factor expression and function in the human and mouse preimplantation embryo. J. Endocrinol. 2002, 172, 221–236. [Google Scholar] [CrossRef]
- Kaye, P.L. Preimplantation growth factor physiology. Rev. Reprod. 1997, 2, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; An, L.; Miao, K.; Ren, L.; Hou, Z.; Tao, L.; Zhang, Z.; Wang, X.; Xia, W.; Liu, J.; et al. Impaired imprinted X chromosome inactivation is responsible for the skewed sex ratio following in vitro fertilization. Proc. Natl. Acad. Sci. USA 2016, 113, 3197–3202. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Yue, Y.; Miao, K.; Xi, G.; Zhang, C.; Wang, W.; An, L.; Tian, J. Repression of FGF signaling is responsible for Dnmt3b inhibition and impaired de novo DNA methylation during early development of in vitro fertilized embryos. Int. J. Biol. Sci. 2020, 16, 3085–3099. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hao, Y.; Li, Z.; Zhou, J.; Zhu, H.; Bu, G.; Liu, Z.; Hou, X.; Zhang, X.; Miao, Y.L. Maternal Cytokines CXCL12, VEGFA, and WNT5A Promote Porcine Oocyte Maturation via MAPK Activation and Canonical WNT Inhibition. Front. Cell Dev. Biol. 2020, 8, 578. [Google Scholar] [CrossRef]
- Hiradate, Y.; Hara, K.; Tanemura, K. Effect of neurotensin on cultured mouse preimplantation embryos. J. Reprod. Dev. 2020, 66, 421–425. [Google Scholar] [CrossRef]
- Gambini, A.; Stein, P.; Savy, V.; Grow, E.J.; Papas, B.N.; Zhang, Y.; Kenan, A.C.; Padilla-Banks, E.; Cairns, B.R.; Williams, C.J. Developmentally Programmed Tankyrase Activity Upregulates beta-Catenin and Licenses Progression of Embryonic Genome Activation. Dev. Cell 2020, 53, 545–560. [Google Scholar] [CrossRef]
- Xie, H.; Tranguch, S.; Jia, X.; Zhang, H.; Das, S.K.; Dey, S.K.; Kuo, C.J.; Wang, H. Inactivation of nuclear Wnt-beta-catenin signaling limits blastocyst competency for implantation. Development 2008, 135, 717–727. [Google Scholar] [CrossRef]
- Albrecht, L.V.; Tejeda-Muñoz, N.; De Robertis, E.M. Cell Biology of Canonical Wnt Signaling. Annu. Rev. Cell Dev. Biol. 2021, 37, 369–389. [Google Scholar] [CrossRef]
- Gordon, M.D.; Nusse, R. Wnt signaling: Multiple pathways, multiple receptors, and multiple transcription factors. J. Biol. Chem. 2006, 281, 22429–22433. [Google Scholar] [CrossRef]
- Barker, N.; Hurlstone, A.; Musisi, H.; Miles, A.; Bienz, M.; Clevers, H. The chromatin remodelling factor Brg-1 interacts with beta-catenin to promote target gene activation. EMBO J. 2001, 20, 4935–4943. [Google Scholar] [CrossRef] [PubMed]
- Hecht, A.; Vleminckx, K.; Stemmler, M.P.; van Roy, F.; Kemler, R. The p300/CBP acetyltransferases function as transcriptional coactivators of beta-catenin in vertebrates. EMBO J. 2000, 19, 1839–1850. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, B.T.; Tamai, K.; He, X. Wnt/beta-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Harwood, B.N.; Cross, S.K.; Radford, E.E.; Haac, B.E.; De Vries, W.N. Members of the WNT signaling pathways are widely expressed in mouse ovaries, oocytes, and cleavage stage embryos. Dev. Dyn. 2008, 237, 1099–1111. [Google Scholar] [CrossRef]
- Wang, J.; Sinha, T.; Wynshaw-Boris, A. Wnt signaling in mammalian development: Lessons from mouse genetics. Cold Spring Harb. Perspect. Biol. 2012, 4, a007963. [Google Scholar] [CrossRef]
- Chu, M.; Yao, F.; Xi, G.; Yang, J.; Zhang, Z.; Yang, Q.; Tian, J.; An, L. Vitamin C Rescues in vitro Embryonic Development by Correcting Impaired Active DNA Demethylation. Front. Cell Dev. Biol. 2021, 9, 784244. [Google Scholar] [CrossRef]
- Lu, D.; Choi, M.Y.; Yu, J.; Castro, J.E.; Kipps, T.J.; Carson, D.A. Salinomycin inhibits Wnt signaling and selectively induces apoptosis in chronic lymphocytic leukemia cells. Proc. Natl. Acad. Sci. USA 2011, 108, 13253–13257. [Google Scholar] [CrossRef]
- Abe, K.I.; Funaya, S.; Tsukioka, D.; Kawamura, M.; Suzuki, Y.; Suzuki, M.G.; Schultz, R.M.; Aoki, F. Minor zygotic gene activation is essential for mouse preimplantation development. Proc. Natl. Acad. Sci. USA 2018, 115, E6780–E6788. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, Y. Loss of DUX causes minor defects in zygotic genome activation and is compatible with mouse development. Nat. Genet. 2019, 51, 947–951. [Google Scholar] [CrossRef]
- Jao, C.Y.; Salic, A. Exploring RNA transcription and turnover in vivo by using click chemistry. Proc. Natl. Acad. Sci. USA 2008, 105, 15779–15784. [Google Scholar] [CrossRef]
- Uribe-Etxebarria, V.; Garcia-Gallastegui, P.; Perez-Garrastachu, M.; Casado-Andres, M.; Irastorza, I.; Unda, F.; Ibarretxe, G.; Subiran, N. Wnt-3a Induces Epigenetic Remodeling in Human Dental Pulp Stem Cells. Cells 2020, 9, 652. [Google Scholar] [CrossRef] [PubMed]
- Dahl, J.A.; Jung, I.; Aanes, H.; Greggains, G.D.; Manaf, A.; Lerdrup, M.; Li, G.; Kuan, S.; Li, B.; Lee, A.Y.; et al. Broad histone H3K4me3 domains in mouse oocytes modulate maternal-to-zygotic transition. Nature 2016, 537, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Li, S.; Wu, Q.; Li, C.; Jiang, M.; Guo, L.; Chen, M.; Yang, L.; Dong, X.; Wang, H.; et al. Stage-specific H3K9me3 occupancy ensures retrotransposon silencing in human pre-implantation embryos. Cell Stem Cell 2022, 29, 1051–1066. [Google Scholar] [CrossRef] [PubMed]
- Chronopoulou, E.; Harper, J.C. IVF culture media: Past, present and future. Hum. Reprod. Update 2015, 21, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dey, S.K. Roadmap to embryo implantation: Clues from mouse models. Nat. Rev. Genet. 2006, 7, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mayernik, L.; Armant, D.R. Integrin signaling regulates blastocyst adhesion to fibronectin at implantation: Intracellular calcium transients and vesicle trafficking in primary trophoblast cells. Dev. Biol. 2002, 245, 270–279. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, Z.; Wu, J. Signaling pathways and preimplantation development of mammalian embryos. FEBS J. 2007, 274, 4349–4359. [Google Scholar] [CrossRef]
- Xia, W.; Xie, W. Rebooting the Epigenomes during Mammalian Early Embryogenesis. Stem Cell Rep. 2020, 15, 1158–1175. [Google Scholar] [CrossRef]
- Shin, S.W.; Tokoro, M.; Nishikawa, S.; Lee, H.H.; Hatanaka, Y.; Nishihara, T.; Amano, T.; Anzai, M.; Kato, H.; Mitani, T.; et al. Inhibition of the ubiquitin-proteasome system leads to delay of the onset of ZGA gene expression. J. Reprod. Dev. 2010, 56, 655–663. [Google Scholar] [CrossRef]
- Sims, R.R.; Reinberg, D. Stem cells: Escaping fates with open states. Nature 2009, 460, 802–803. [Google Scholar] [CrossRef]
- Liu, X.; Wang, C.; Liu, W.; Li, J.; Li, C.; Kou, X.; Chen, J.; Zhao, Y.; Gao, H.; Wang, H.; et al. Distinct features of H3K4me3 and H3K27me3 chromatin domains in pre-implantation embryos. Nature 2016, 537, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zheng, H.; Huang, B.; Li, W.; Xiang, Y.; Peng, X.; Ming, J.; Wu, X.; Zhang, Y.; Xu, Q.; et al. Allelic reprogramming of the histone modification H3K4me3 in early mammalian development. Nature 2016, 537, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Lindeman, L.C.; Andersen, I.S.; Reiner, A.H.; Li, N.; Aanes, H.; Ostrup, O.; Winata, C.; Mathavan, S.; Muller, F.; Alestrom, P.; et al. Prepatterning of developmental gene expression by modified histones before zygotic genome activation. Dev. Cell 2011, 21, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, R.; Gonzales-Cope, M.; Chronis, C.; Bonora, G.; Mckee, R.; Huang, C.; Patel, S.; Lopez, D.; Mishra, N.; Pellegrini, M.; et al. Proteomic and genomic approaches reveal critical functions of H3K9 methylation and heterochromatin protein-1gamma in reprogramming to pluripotency. Nat. Cell Biol. 2013, 15, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Sankar, A.; Lerdrup, M.; Manaf, A.; Johansen, J.V.; Gonzalez, J.M.; Borup, R.; Blanshard, R.; Klungland, A.; Hansen, K.; Andersen, C.Y.; et al. KDM4A regulates the maternal-to-zygotic transition by protecting broad H3K4me3 domains from H3K9me3 invasion in oocytes. Nat. Cell Biol. 2020, 22, 380–388. [Google Scholar] [CrossRef]
- Barrera, A.D.; Garcia, E.V.; Hamdi, M.; Sanchez-Calabuig, M.J.; Lopez-Cardona, A.P.; Balvis, N.F.; Rizos, D.; Gutierrez-Adan, A. Embryo culture in presence of oviductal fluid induces DNA methylation changes in bovine blastocysts. Reproduction 2017, 154, 1–12. [Google Scholar] [CrossRef]
- Canovas, S.; Ivanova, E.; Romar, R.; Garcia-Martinez, S.; Soriano-Ubeda, C.; Garcia-Vazquez, F.A.; Saadeh, H.; Andrews, S.; Kelsey, G.; Coy, P. DNA methylation and gene expression changes derived from assisted reproductive technologies can be decreased by reproductive fluids. Elife 2017, 6, e23670. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | WNT2 | |
|---|---|---|
| Recipients (n) | 31 | 18 |
| Implantations (%) | 66.13 ± 23.37 | 82.87 ± 15.33 * |
| Fetuses (% of transferred) | 30.11 ± 13.50 | 39.81 ± 17.25 * |
| Fetuses (% of implanted) | 50.56 ± 23.60 | 48.42 ± 19.74 |
| Fetal weight (g) | 1.54 ± 0.30 | 1.56 ± 0.27 |
| Occipito-frontal diameter (mm) | 6.79 ± 1.26 | 6.88 ± 1.38 |
| Crown rump length (mm) | 24.44 ± 2.22 | 24.80 ± 2.21 |
| Tail length (mm) | 12.61 ± 1.79 | 12.90 ± 1.25 |
| Placental weight (g) | 0.16 ± 0.03 | 0.15 ± 0.04 |
| Placental diameter (mm) | 8.89 ± 1.17 | 8.63 ± 1.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, F.; Hao, J.; Wang, Z.; Chu, M.; Zhang, J.; Xi, G.; Zhang, Z.; An, L.; Tian, J. WNT Co-Receptor LRP6 Is Critical for Zygotic Genome Activation and Embryonic Developmental Potential by Interacting with Oviductal Paracrine Ligand WNT2. Genes 2023, 14, 891. https://doi.org/10.3390/genes14040891
Yao F, Hao J, Wang Z, Chu M, Zhang J, Xi G, Zhang Z, An L, Tian J. WNT Co-Receptor LRP6 Is Critical for Zygotic Genome Activation and Embryonic Developmental Potential by Interacting with Oviductal Paracrine Ligand WNT2. Genes. 2023; 14(4):891. https://doi.org/10.3390/genes14040891
Chicago/Turabian StyleYao, Fusheng, Jia Hao, Zhaochen Wang, Meiqiang Chu, Jingyu Zhang, Guangyin Xi, Zhenni Zhang, Lei An, and Jianhui Tian. 2023. "WNT Co-Receptor LRP6 Is Critical for Zygotic Genome Activation and Embryonic Developmental Potential by Interacting with Oviductal Paracrine Ligand WNT2" Genes 14, no. 4: 891. https://doi.org/10.3390/genes14040891
APA StyleYao, F., Hao, J., Wang, Z., Chu, M., Zhang, J., Xi, G., Zhang, Z., An, L., & Tian, J. (2023). WNT Co-Receptor LRP6 Is Critical for Zygotic Genome Activation and Embryonic Developmental Potential by Interacting with Oviductal Paracrine Ligand WNT2. Genes, 14(4), 891. https://doi.org/10.3390/genes14040891