DNMT3B rs2424913 as a Risk Factor for Congenital Heart Defects in Down Syndrome

 ,
, 
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. DNA Isolation and Genotyping
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, W.; He, J.; Shao, X. Incidence and mortality trend of congenital heart disease at the global, regional, and national level, 1990–2017. Medicine 2020, 99, e20593. [Google Scholar] [CrossRef]
- GBD 2017 Congenital Heart Disease Collaborators. Global, regional, and national burden of congenital heart disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Child Adolesc. Health 2020, 4, 185–200. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, S.; Zühlke, L.; Black, G.C.; Choy, M.K.; Li, N.; Keavney, B.D. Global birth prevalence of congenital heart defects 1970–2017: Updated systematic review and meta-analysis of 260 studies. Int. J. Epidemiol. 2019, 48, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Lalani, S.R.; Belmont, J.W. Genetic basis of congenital cardiovascular malformations. Eur. J. Med. Genet. 2014, 57, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Landis, B.J.; Cooper, D.S.; Hinton, R.B. CHD associated with syndromic diagnoses: Peri-Operative risk factors and early outcomes. Cardiol. Young 2016, 26, 30–52. [Google Scholar] [CrossRef] [PubMed]
- Pfitzer, C.; Helm, P.C.; Rosenthal, L.M.; Berger, F.; Bauer, U.M.M.; Schmitt, K.R. Dynamics in prevalence of Down syndrome in children with congenital heart disease. Eur. J. Pediatr. 2018, 177, 107–115. [Google Scholar] [CrossRef]
- Ackerman, C.; Locke, A.E.; Feingold, E.; Reshey, B.; Espana, K.; Thusberg, J.; Mooney, S.; Bean, L.J.; Dooley, K.J.; Cua, C.L.; et al. An excess of deleterious variants in VEGF-A pathway genes in Down-syndrome-associated atrioventricular septal defects. Am. J. Hum. Genet. 2012, 91, 646–659. [Google Scholar] [CrossRef]
- Benhaourech, S.; Drighil, A.; Hammiri, A.E. Congenital heart disease and Down syndrome: Various aspects of a confirmed association. Cardiovasc. J. Afr. 2016, 27, 287–290. [Google Scholar] [CrossRef]
- Zaidi, S.; Brueckner, M. Genetics and Genomics of Congenital Heart Disease. Circ. Res. 2017, 120, 923–940. [Google Scholar] [CrossRef]
- Antonarakis, S.E.; Lyle, R.; Dermitzakis, E.T.; Reymond, A.; Deutsch, S. Chromosome 21 and Down syndrome: From genomics to pathophysiology. Nat. Rev. Genet. 2004, 5, 725–738. [Google Scholar] [CrossRef]
- Letourneau, A.; Santoni, F.A.; Bonilla, X.; Sailani, M.R.; Gonzalez, D.; Kind, J.; Chevalier, C.; Thurman, R.; Sandstrom, R.S.; Hibaoui, Y.; et al. Domains of genome-wide gene expression dysregulation in Down’s syndrome. Nature 2014, 508, 345–350. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, L.; Tian, J. Molecular mechanisms of congenital heart disease in down syndrome. Genes Dis. 2019, 6, 372–377. [Google Scholar] [CrossRef]
- Asim, A.; Agarwal, S. Congenital heart defects among Down’s syndrome cases: An updated review from basic research to an emerging diagnostics technology and genetic counselling. J. Genet. 2021, 100, 45. [Google Scholar] [CrossRef] [PubMed]
- Coppedè, F. The genetics of folate metabolism and maternal risk of birth of a child with Down syndrome and associated congenital heart defects. Front. Genet. 2015, 6, 223. [Google Scholar] [CrossRef] [PubMed]
- Coppedè, F.; Stoccoro, A.; Tannorella, P.; Gallo, R.; Nicolì, V.; Migliore, L. Association of Polymorphisms in Genes Involved in One-Carbon Metabolism with MTHFR Methylation Levels. Int. J. Mol. Sci. 2019, 20, 3754. [Google Scholar] [CrossRef] [PubMed]
- Villicaña, S.; Bell, J.T. Genetic impacts on DNA methylation: Research findings and future perspectives. Genome Biol. 2021, 22, 127. [Google Scholar] [CrossRef]
- Jin, Z.; Liu, Y. DNA methylation in human diseases. Genes Dis. 2018, 5, 1–8. [Google Scholar] [CrossRef]
- Moore, L.; Le, T.; Fan, G. DNA Methylation and Its Basic Function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Wang, H.; Lou, D.; Wang, Z. Crosstalk of Genetic Variants, Allele-Specific DNA Methylation, and Environmental Factors for Complex Disease Risk. Front. Genet. 2019, 9, 695. [Google Scholar] [CrossRef]
- Coppedè, F. Risk factors for Down syndrome. Arch. Toxicol. 2016, 90, 2917–2929. [Google Scholar] [CrossRef]
- Chowdhury, S.; Cleves, M.A.; MacLeod, S.L.; James, S.J.; Zhao, W.; Hobbs, C.A. Maternal DNA hypomethylation and congenital heart defects. Birth Defects Res. A Clin. Mol. Teratol. 2011, 91, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wu, Q.; Huang, Y.; Wang, L.; Su, Z.; Ye, H. The role of DNA methylation in syndromic and non-syndromic congenital heart disease. Clin. Epigenet. 2021, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhou, S.; Wang, Y.; Wang, L.; Zhou, J.; Wang, H.; Li, C.; Chang, M. Association of DNMT1 Gene Polymorphisms with Congenital Heart Disease in Child Patients. Pediatr. Cardiol. 2015, 36, 906–911. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Wang, W.; Jiang, X. The roles of MTRR and MTHFR gene polymorphisms in congenital heart diseases: A me-ta-analysis. Biosci. Rep. 2018, 38, BSR20181160. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Zhuang, Z.; Wen, Z.; Zang, X.; Mo, X. MTHFR A1298C polymorphisms reduce the risk of congenital heart defects: A meta-analysis from 16 case-control studies. Ital. J. Pediatr. 2017, 43, 108. [Google Scholar] [CrossRef] [PubMed]
- Castiglia, P.; Sanna, V.; Azara, A.; De Miglio, M.R.; Murgia, L.; Pira, G.; Sanges, F.; Fancellu, A.; Carru, C.; Bisail, M.; et al. Methylenetetrahydrofolate reductase (MTHFR) C677T and A1298C polymorphisms in breast cancer: A Sardinian preliminary case-control study. Int. J. Med. Sci. 2019, 16, 1089–1095. [Google Scholar] [CrossRef]
- Cai, B.; Zhang, T.; Zhong, R.; Zou, L.; Zhu, B.; Chen, W.; Shen, N.; Ke, J.; Lou, J.; Wang, Z.; et al. Genetic variant in MTRR, but not MTR, is associated with risk of congenital heart disease: An integrated meta-analysis. PLoS ONE 2014, 9, e89609. [Google Scholar] [CrossRef]
- Sheng, W.; Qian, Y.; Wang, H.; Ma, X.; Zhang, P.; Chen, L.; Ma, D.; Huang, G. Association between mRNA levels of DNMT1, DNMT3A, DNMT3B, MBD2 and LINE-1 methylation status in infants with tetralogy of Fallot. Int. J. Mol. Med. 2013, 32, 694–702. [Google Scholar] [CrossRef]
- Lei, L.; Lin, H.; Zhong, S.; Zhang, Z.; Chen, J.; Yu, X.; Liu, X.; Zhang, C.; Nie, Z.; Zhuang, J. DNA methyltransferase 1 rs16999593 genetic polymorphism decreases risk in patients with transposition of great arteries. Gene 2017, 615, 50–56. [Google Scholar] [CrossRef]
- Zhu, S.; Zhang, H.; Tang, Y.; Liu, P.; Wang, J. DNMT3B polymorphisms and cancer risk: A meta analysis of 24 case-control studies. Mol. Biol. Rep. 2012, 39, 4429–4437. [Google Scholar] [CrossRef]
- Shen, H.; Wang, L.; Spitz, M.R.; Hong, W.K.; Mao, L.; Wei, Q. A novel polymorphism in human cytosine DNA-methyltransferase-3B promoter is associated with an increased risk of lung cancer. Cancer Res. 2002, 62, 4992–4995. [Google Scholar]
- Xiao, Y.; Word, B.; Hammons, G.; Lyn-Cook, B. Transcriptional activity of DNMT3B in pancreatic cancer cells: Effects of −149 (C→T) promoter polymorphism. Biochem. Biophys. Res. Commun. 2011, 415, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Saradalekshmi, K.R.; Neetha, N.V.; Sathyan, S.; Nair, I.V.; Nair, C.M.; Banerjee, M. DNA methyl transferase (DNMT) gene polymorphisms could be a primary event in epigenetic susceptibility to schizophrenia. PLoS ONE 2014, 9, e98182. [Google Scholar] [CrossRef] [PubMed]
- Coppedè, F.; Boco, P.; Tannorella, P.; Romano, C.; Antonucci, I.; Stuppia, L.; Romano, C.; Migliore, L. DNMT3B promoter polymorphisms and maternal risk of birth of a child with Down syndrome. Hum. Reprod. 2013, 28, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, A.A.; Lin, M.; Lister, R.L.; Maslov, A.A.; Wang, Y.; Suzuki, M.; Wu, B.; Greally, J.M.; Zheng, D.; Zhou, B. DNA methylation is developmentally regulated for genes essential for cardiogenesis. J. Am. Heart Assoc. 2014, 3, e000976. [Google Scholar] [CrossRef]
- Fang, X.; Poulsen, R.R.; Wang-Hu, J.; Shi, O.; Calvo, N.S.; Simmons, C.S.; Rivkees, S.A.; Wendler, C.C. Knockdown of DNA methyltransferase 3a alters gene expression and inhibits function of embryonic cardiomyocytes. FASEB J. 2016, 30, 3238–3255. [Google Scholar] [CrossRef]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef]
- Gilsbach, R.; Preissl, S.; Grüning, B.A.; Schnick, T.; Burger, L.; Benes, V.; Würch, A.; Bönisch, U.; Günther, S.; Backofen, R.; et al. Dynamic DNA methylation orchestrates cardiomyocyte development, maturation and disease. Nat. Commun. 2014, 5, 5288. [Google Scholar] [CrossRef]
- Zhang, D.; Wu, B.; Wang, P.; Wang, Y.; Lu, P.; Nechiporuk, T.; Floss, T.; Greally, J.M.; Zheng, D.; Zhou, B. Non-CpG methylation by DNMT3B facilitates REST binding and gene silencing in developing mouse hearts. Nucleic Acids Res. 2017, 45, 3102–3115. [Google Scholar] [CrossRef]
- Gaunt, T.R.; Shihab, H.A.; Hemani, G.; Min, J.L.; Woodward, G.; Lyttleton, O.; Zheng, J.; Duggirala, A.; McArdle, W.L.; Ho, K.; et al. Systematic identification of genetic influences on methylation across the human life course. Genome Biol. 2016, 17, 61. [Google Scholar] [CrossRef]
- Tao, H.; Yang, J.J.; Chen, Z.W.; Xu, S.S.; Zhou, X.; Zhan, H.; Shi, K.H. DNMT3A silencing RASSF1A promotes cardiac fibrosis through upregulation of ERK1/2. Toxicology 2014, 323, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Poulsen, R.; Zhao, L.; Wang, J.; Rivkees, S.A.; Wendler, C.C. Knockdown of DNA methyltransferase 1 reduces DNA methylation and alters expression patterns of cardiac genes in embryonic cardiomyocytes. FEBS Open Bio. 2021, 11, 2364–2382. [Google Scholar] [CrossRef] [PubMed]
- Junker, R.; Kotthoff, S.; Vielhaber, H.; Halimeh, S.; Kosch, A.; Koch, H.G.; Kassenböhmer, R.; Heineking, B.; Nowak-Göttl, U. Infant methylenetetrahydrofolate reductase 677TT genotype is a risk factor for congenital heart disease. Cardiovasc. Res. 2001, 51, 251–254. [Google Scholar] [CrossRef] [PubMed]
- van Beynum, I.M.; den Heijer, M.; Blom, H.J.; Kapusta, L. The MTHFR 677C->T polymorphism and the risk of congenital heart defects: A literature review and meta-analysis. QJM 2007, 100, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.N.; Wang, H.D.; Tie, L.Z.; Li, T.; Xiao, H.; Long, J.G.; Liao, S.X. Parental Genetic Variants, MTHFR 677C>T and MTRR 66A>G, Associated Differently with Fetal Congenital Heart Defect. Biomed. Res. Int. 2017, 2017, 3043476. [Google Scholar] [CrossRef]
- Asim, A.; Agarwal, S.; Panigrahi, I.; Saiyed, N.; Bakshi, S. MTHFR promoter hypermethylation may lead to congenital heart defects in Down syndrome. Intractable Rare Dis. Res. 2017, 6, 295–298. [Google Scholar] [CrossRef]
- Božović, I.B.; Vraneković, J.; Cizmarević, N.S.; Mahulja-Stamenković, V.; Prpić, I.; Brajenović-Milić, B. MTHFR C677T and A1298C polymorphisms as a risk factor for congenital heart defects in Down syndrome. Pediatr. Int. 2011, 53, 546–550. [Google Scholar] [CrossRef]
- Vraneković, J.; Slivšek, G.; Majstorović, D. Methyltetrahydrofolate-homocysteine methyltransferase reductase gene and congenital heart defects in Down syndrome. Genet. Appl. 2020, 41, 12–17. [Google Scholar] [CrossRef]
- Ganguly, A.; Halder, P.; Pal, U.; Sarkar, S.; Datta, S.; Pati, S.; Ghosh, S. Risk of Atrioventricular Septal Defects in Down syndrome: Association of MTHFR C677T and RFC1 A80G polymorphisms in Indian Bengali cohort. J. Hum. Genet. Genom. 2021, 5, 1–12. [Google Scholar] [CrossRef]
- Khatami, F.; Noorinayer, B.; Ghiasi, S.; Mohebi, R.; Hashemi, M.; Zali, M.R. Lack of Effects of Single Nucleotide Polymorphisms of the DNA Methyltransferase 1 Gene on Gastric Cancer in Iranian Patients: A Case Control Study. Asian Pac. J. Cancer Prev. 2009, 10, 1177–1182. [Google Scholar]
- Fan, H.; Liu, D.; Qiu, X.; Qiao, F.; Wu, Q.; Su, X.; Zhang, F.; Song, Y.; Zhao, Z.; Xie, W. A functional polymorphism in the DNA methyltransferase-3A promoter modifies the susceptibility in gastric cancer but not in esophageal carcinoma. BMC Med. 2010, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Zhang, F.; Hu, J.; Liu, D.; Zhao, Z. Promoter polymorphisms of DNMT3B and the risk of colorectal cancer in Chinese: A case-control study. J. Exp. Clin. Cancer Res. CR 2008, 27, 24. [Google Scholar] [CrossRef] [PubMed]
- Mostowska, A.; Sajdak, S.; Pawlik, P.; Lianeri, M.; Jagodzinski, P.P. DNMT1, DNMT3A and DNMT3B gene variants in relation to ovarian cancer risk in the Polish population. Mol. Biol. Rep. 2013, 40, 4893–4899. [Google Scholar] [CrossRef] [PubMed]
- Coppedè, F.; Marini, G.; Bargagna, S.; Stuppia, L.; Minichilli, F.; Fontana, I.; Colognato, R.; Astrea, G.; Palka, G.; Migliore, L. Folate gene polymorphisms and the risk of Down syndrome pregnancies in young Italian women. Am. J. Med. Genet. Part A 2006, 140, 1083–1091. [Google Scholar] [CrossRef]
- Jacques, P.F.; Bostom, A.G.; Selhub, J.; Rich, S.; Ellison, R.C.; Eckfeldt, J.H.; Gravel, R.A.; Rozen, R.; National Heart, Lung and Blood Institute; National Institutes of Health. Effects of polymorphisms of methionine synthase and methionine synthase reductase on total plasma homocysteine in the NHLBI Family Heart Study. Atherosclerosis 2003, 66, 49–55. [Google Scholar] [CrossRef]


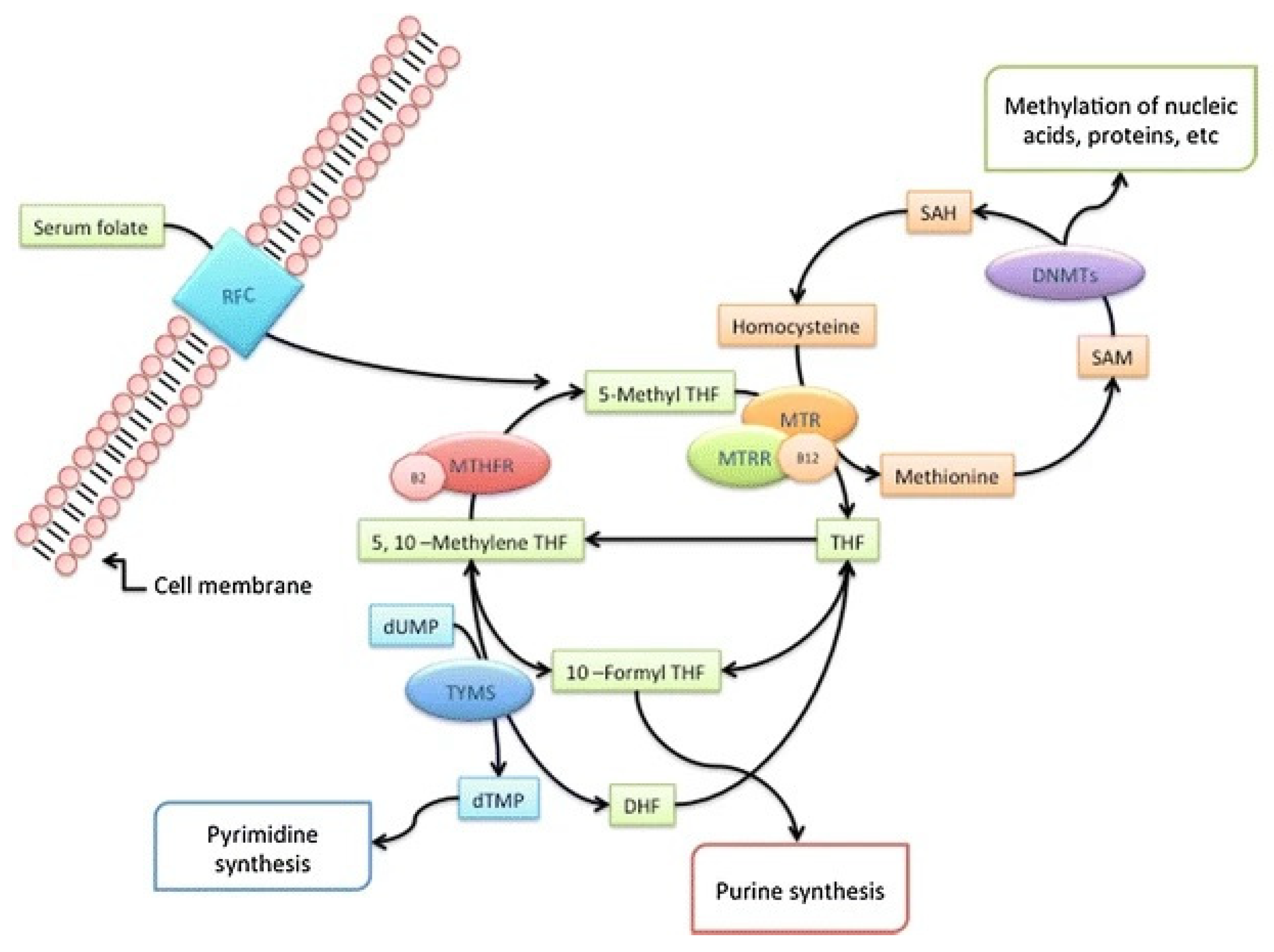{kind=link}
| DSCHD+ | DSCHD− | CHD | |
|---|---|---|---|
| No. of cases (%) | 134 | 124 | 92 |
| Gender | |||
| Male | 69 () (51.5) | 71 () (57.3) | 55 (59.8) |
| Female | 65 () (48.5) | 53 () (42.7) | 37 (40.2) |
| Age | Median [range] | ||
| 2 [0–27] | 2.5 [0–55] | 7.5 [0–32] | |
| DSCHD+ N (%) | DSCHD− N (%) | CHD N (%) | χ2 DSCHD+ DSCHD− | p DSCHD+ DSCHD− | χ2 DSCHD+ CHD | p DSCHD+ CHD | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| DNMT1 | rs2228611 | Genotype | AA AG GG | 31 (24.0) 80 (62.0) 18 (14.0) | 38 (31.7) 66 (55.0) 16 (13.3) | 27 (29.3) 46 (50.0) 19 (20.7) | 1.81 1.26 0.02 | 0.180 0.262 0.887 | 0.78 3.16 1.73 | 0.377 0.076 0.191 |
| Allele | A G | 142 (55.0) 116 (45.0) | 142 (59.2) 98 (40.8) | 100 (54.3) 84 (45.7) | 0.87 | 0.353 | 0.02 | 0.886 | ||
| DNMT3A | rs1550117 | Genotype | AA AG GG | 1 (0.8) 21 (16.7) 104 (82.5) | 1 (0.8) 17 (14.3) 101 (84.9) | 1 (1.1) 15 (16.5) 75 (82.4) | 0.00 0.27 0.24 | 0.737 0.607 0.622 | 0.05 0.00 0.00 | 0.664 0.972 0.981 |
| Allele | A G | 23 (10.0) 229 (90.0) | 19 (8.0) 219 (92.0) | 17 (9.3) 165 (90.7) | 0.20 | 0.652 | 0.04 | 0.485 | ||
| DNMT3B | rs1569686 | Genotype | GG TG TT | 50 (38.5) 63 (48.5) 17 (13.0) | 45 (36.3) 65 (52.4) 14 (11.3) | 32 (34.8) 38 (41.3) 22 (23.9) | 0.13 0.40 0.19 | 0.721 0.528 0.664 | 0.31 1.11 4.37 | 0.576 0.292 0.039 |
| Allele | G T | 163 (63) 97 (37) | 155 (63) 93 (38) | 102 (55) 82 (45) | 0.00 | 0.964 | 2.36 | 0.125 | ||
| rs2424913 | Genotype | CC CT TT | 41 (33.1) 52 (41.9) 31 (25.0) | 35 (30.2) 66 (56.9) 15 (12.9) | 29 (32.2) 37 (41.1) 24 (26.7) | 0.23 5.37 5.63 | 0.630 0.021 0.019 | 0.02 0.01 0.08 | 0.897 0.904 0.783 | |
| Allele | C T | 134 (54) 114 (46) | 136 (59) 96 (41) | 95 (53) 85 (47) | 1.03 | 0.311 | 0.07 | 0.246 | ||
| MTHFR | rs1801133 | Genotype | CC CT TT | 51 (38.1) 68 (50.7) 15 (11.2) | 58 (46.7) 55 (44.4) 11 (8.9) | 45 (48.5) 37 (40.2) 10 (10.9) | 2.01 1.06 0.38 | 0.157 0.305 0.537 | 2.64 2.43 0.01 | 0.106 0.120 0.939 |
| Allele | C T | 170 (63.4) 98 (36.6) | 171 (68.9) 77 (31.1) | 127 (69.0) 57 (31.0) | 1.75 | 0.186 | 1.51 | 0.219 | ||
| rs1801131 | Genotype | AA AC CC | 76 (56.7) 55 (41.1) 3 (2.2) | 55 (44.4) 65 (52.4) 4 (3.2) | 41 (44.6) 46 (50.0) 5 (5.4) | 3.94 3.35 0.24 | 0.048 0.068 0.457 | 3.23 1.77 1.63 | 0.073 0.184 0.181 | |
| Allele | A C | 207 (77.2) 61 (22.8) | 175 (70.6) 73 (29.4) | 128 (69.6) 56 (30.4) | 2.99 | 0.085 | 3.35 | 0.068 | ||
| MTRR | rs1801394 | Genotype | AA AG GG | 32 (24.1) 70 (52.6) 31 (23.3) | 23 (18.5) 72 (58.1) 29 (23.4) | 16 (17.4) 59 (64.1) 17 (18.5) | 1.09 0.77 0.00 | 0.297 0.382 0.988 | 1.44 2.94 0.76 | 0.232 0.087 0.386 |
| Allele | A G | 134 (50.4) 132 (49.6) | 118 (47.6) 130 (52.4) | 91 (49.5) 93 (50.5) | 0.40 | 0.527 | 0.04 | 0.848 | ||
| Genetic Model | DSCHD+ vs. DSCHD− | DSCHD+ vs. CHD | |||
|---|---|---|---|---|---|
| OR (95% CI) | p | OR (95% CI) | p | ||
| DNMT1 rs2228611 | |||||
| Dominant | AA + AG vs. GG | 1.05 (0.51–2.17) | 0.886 | 1.60 (0.78–3.26) | 0.191 |
| Recessive | AA vs. AG + GG | 1.46 (0.83–2.55) | 0.179 | 0.76 (0.41–1.39) | 0.376 |
| Codominant | AA vs. GG | 1.37 (0.60–3.14) | 0.444 | 1.21 (0.53–2.76) | 0.648 |
| Codominant | AA vs. AG | 1.48 (0.83–2.64) | 0.177 | 0.66 (0.35–1.24) | 0.196 |
| Codominant | GG vs. AG | 1.07 (0.50–2.27) | 0.845 | 0.54 (0.26–1.14) | 0.107 |
| DNMT3A rs1550117 | |||||
| Dominant | AA + AG vs. GG | 0.84 (0.42–1.66) | 0.621 | 0.99 (0.48–2.01) | 0.981 |
| Recessive | AA vs. AG + GG | 1.05 (0.06–17.13) | 0.736 | 0.72 (0.04–11.66) | 0.664 |
| Codominant | AA vs. GG | 1.02 (0.06–16.68) | 0.743 | 0.72 (0.04–11.71) | 0.664 |
| Codominant | AA vs. AG | 1.23 (0.07–21.24) | 0.703 | 0.71 (0.04–12.34) | 0.671 |
| Codominant | GG vs. AG | 1.19 (0.59–2.40) | 0.608 | 0.99 (0.47–2.04) | 0.979 |
| DNMT3B rs1569686 | |||||
| Dominant | GG + TG vs. TT | 1.18 (0.55–2.51) | 0.664 | 2.08 (1.03–4.20) | 0.039 |
| Recessive | GG vs. TG + TT | 0.91 (0.54–1.51) | 0.720 | 1.17 (0.67–2.04) | 0.576 |
| Codominant | GG vs. TT | 1.09 (0.48–2.46) | 0.830 | 2.02 (0.93–4.38) | 0.074 |
| Codominant | GG vs. TG | 0.87 (0.51–1.48) | 0.614 | 0.94 (0.51–1.71) | 0.846 |
| Codominant | TT vs. TG | 0.79 (0.36–1.75) | 0.574 | 0.46 (0.22–0.98) | 0.046 |
| DNMT3B rs2424913 | |||||
| Dominant | CC + CT vs. TT | 2.24 (1.13–4.42) | 0.019 | 1.09 (0.58–2.02) | 0.783 |
| Recessive | CC vs. CT + TT | 0.87 (0.50–1.50) | 0.630 | 1.20 (0.68–2.14) | 0.514 |
| Codominant | CC vs. TT | 1.76 (0.82–3.78) | 0.145 | 1.09 (0.53–2.23) | 0.804 |
| Codominant | CC vs. CT | 0.67 (0.37–1.20) | 0.179 | 1.00 (0.53–1.89) | 0.985 |
| Codominant | TT vs. CT | 0.38 (0.18–0.77) | 0.008 | 0.91 (0.46–1.81) | 0.807 |
| MTHFR rs1801133 | |||||
| Dominant | CC + CT vs. TT | 1.29 (0.57–2.93) | 0.536 | 0.96 (0.41–2.25) | 0.939 |
| Recessive | CC vs. CT + TT | 1.43 (0.87–2.34) | 0.157 | 0.64 (0.37–1.09) | 0.105 |
| Codominant | CC vs. TT | 1.55 (0.65–3.68) | 0.319 | 0.75 (0.30–1.84) | 0.539 |
| Codominant | CC vs. CT | 1.40 (0.83–2.35) | 0.196 | 0.61 (0.34–1.08) | 0.094 |
| Codominant | TT vs. CT | 0.90 (0.38–2.13) | 0.822 | 0.81 (0.33–1.99) | 0.656 |
| MTHFR rs1801131 | |||||
| Dominant | AA + AC vs. CC | 0.68 (0.15–3.13) | 0.457 | 2.50 (0.58–10.77) | 0.180 |
| Recessive | AA vs. AC + CC | 0.60 (0.37–0.99) | 0.047 | 1.62 (0.95–2.78) | 0.073 |
| Codominant | AA vs. CC | 0.54 (0.11–2.52) | 0.341 | 3.08 (0.70–13.58) | 0.120 |
| Codominant | AA vs. AC | 0.61 (0.37–1.00) | 0.054 | 1.55 (0.89–2.67) | 0.115 |
| Codominant | CC vs. AC | 1.12 (0.24–5.25) | 0.596 | 0.50 (0.11–2.21) | 0.288 |
| MTRR rs1801394 | |||||
| Dominant | AA + AG vs. GG | 0.99 (0.55–1.77) | 0.988 | 0.74 (0.38–1.44) | 0.385 |
| Recessive | AA vs. AG + GG | 0.71 (0.39–1.31) | 0.282 | 1.50 (0.77–2.94) | 0.231 |
| Codominant | AA vs.GG | 0.76 (0.36–1.60) | 0.483 | 1.09 (0.47–2.54) | 0.829 |
| Codominant | AA vs. AG | 0.69 (0.37–1.31) | 0.263 | 1.68 (0.84–3.37) | 0.139 |
| Codominant | GG vs. AG | 0.90 (0.49–1.66) | 0.758 | 1.53 (0.77–3.05) | 0.219 |
| ASD N (%) | CHD N (%) | χ2 | OR (95% Cl) | p-Value | |
|---|---|---|---|---|---|
| CC | 11 (24.4) | 30 (38.0) | 2.37 | 0.53 (0.23–1.20) | 0.126 |
| CT | 18 (40.0) | 35 (44.3) | 0.22 | 0.84 (0.40–1.76) | 0.642 |
| TT | 16 (35.6) | 14 (17.7) | 4.97 | 2.56 (1.11–5.93) | 0.028 |
| CC vs. CT + TT | Recessive | 2.37 | 0.53 (0.23–1.20) | 0.126 | |
| CC + CT vs. TT | Dominant | 4.97 | 0.39 (0.17–0.90) | 0.028 | |
| Allele | 5.69 | 0.53 (0.31–0.90) | 0.018 | ||
| C | 40 (44.4) | 95 (60.1) | |||
| T | 50 (55.6) | 63 (39.9) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majstorović, D.; Barišić, A.; Božović, I.B.; Čače, I.B.; Čače, N.; Štifanić, M.; Vraneković, J. DNMT3B rs2424913 as a Risk Factor for Congenital Heart Defects in Down Syndrome. Genes 2023, 14, 576. https://doi.org/10.3390/genes14030576
Majstorović D, Barišić A, Božović IB, Čače IB, Čače N, Štifanić M, Vraneković J. DNMT3B rs2424913 as a Risk Factor for Congenital Heart Defects in Down Syndrome. Genes. 2023; 14(3):576. https://doi.org/10.3390/genes14030576
Chicago/Turabian StyleMajstorović, Dijana, Anita Barišić, Ivana Babić Božović, Iva Bilić Čače, Neven Čače, Mauro Štifanić, and Jadranka Vraneković. 2023. "DNMT3B rs2424913 as a Risk Factor for Congenital Heart Defects in Down Syndrome" Genes 14, no. 3: 576. https://doi.org/10.3390/genes14030576
APA StyleMajstorović, D., Barišić, A., Božović, I. B., Čače, I. B., Čače, N., Štifanić, M., & Vraneković, J. (2023). DNMT3B rs2424913 as a Risk Factor for Congenital Heart Defects in Down Syndrome. Genes, 14(3), 576. https://doi.org/10.3390/genes14030576