
Genome-Wide Association Studies and Runs of Homozygosity to Identify Reproduction-Related Genes in Yorkshire Pig Population
 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Genotyping and Quality Control
2.3. Phenotypic Correlation Analysis
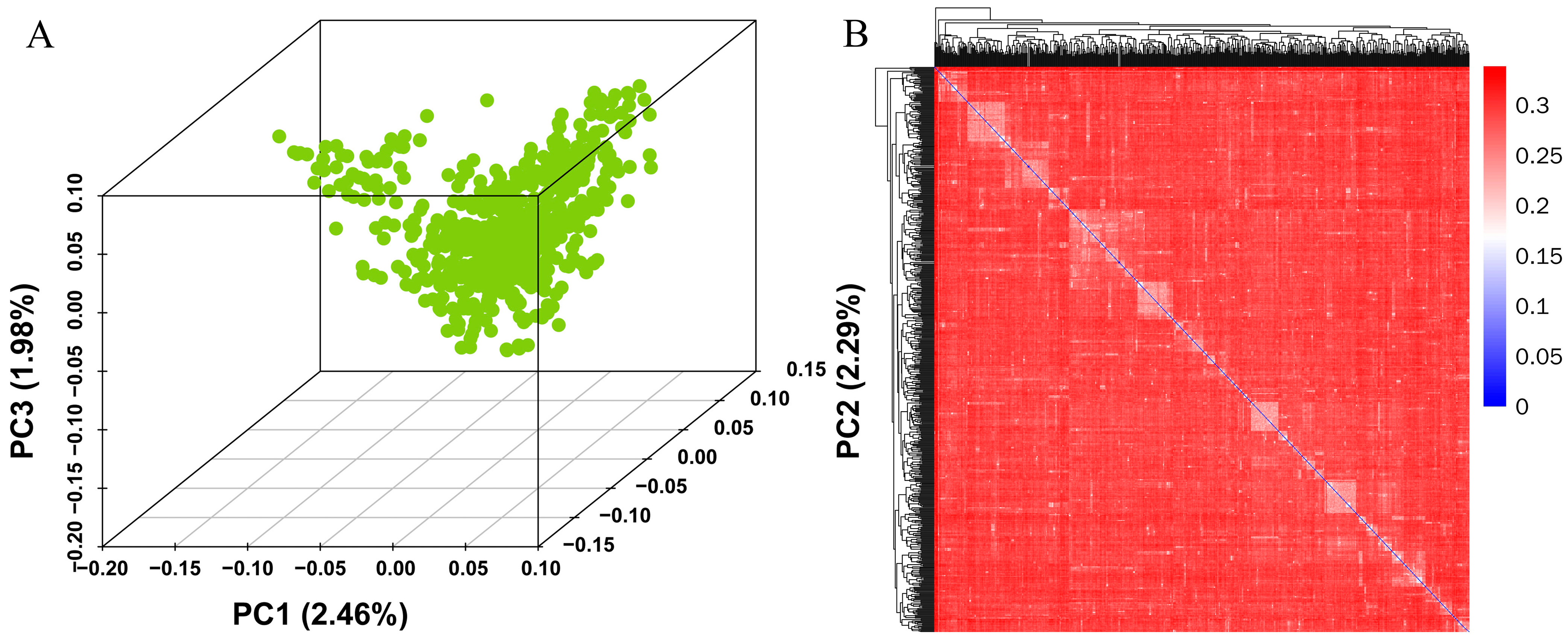
2.4. Principal Component Analysis (PCA) and Relationship Analysis
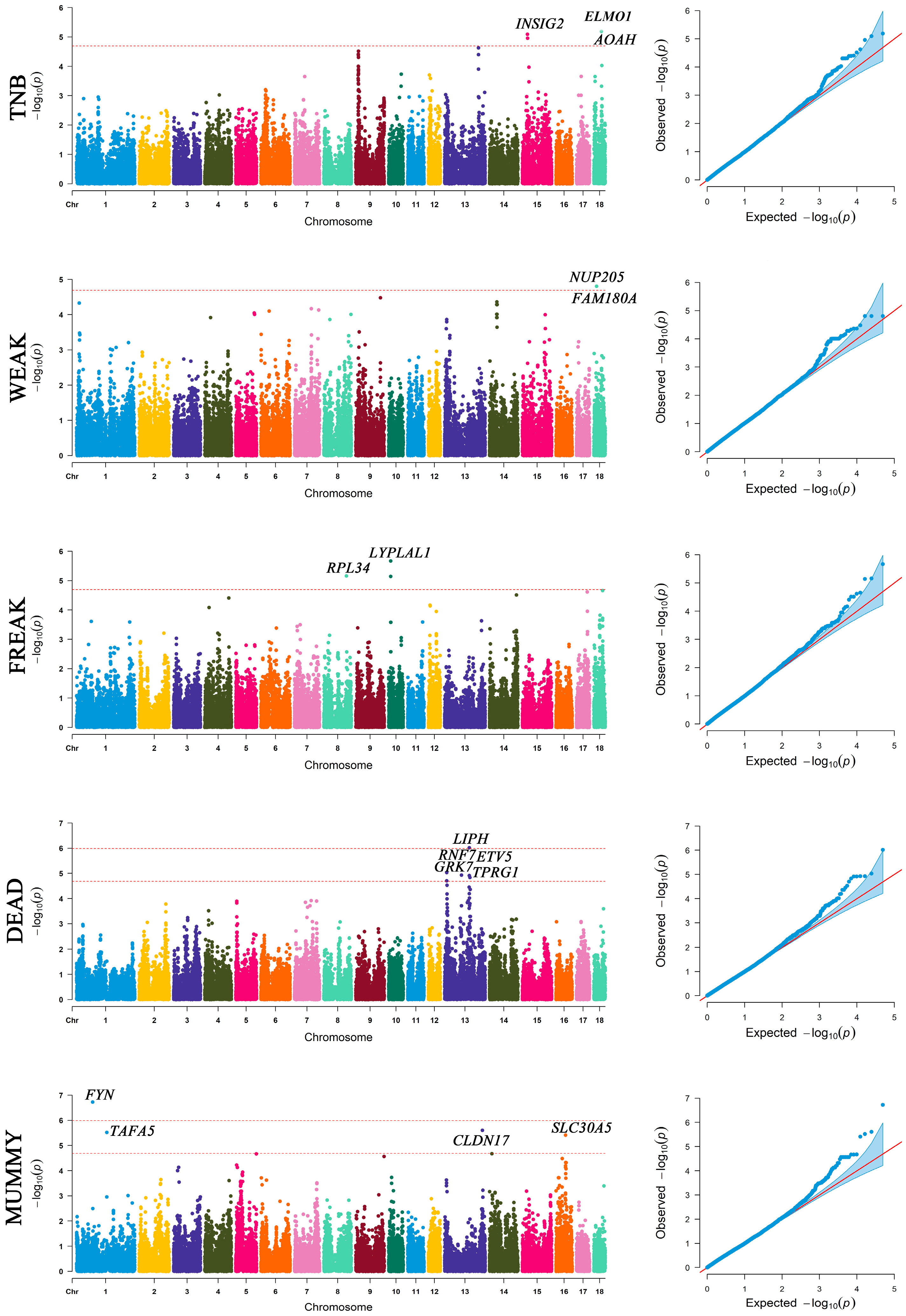
2.5. Genome-Wide Association Study
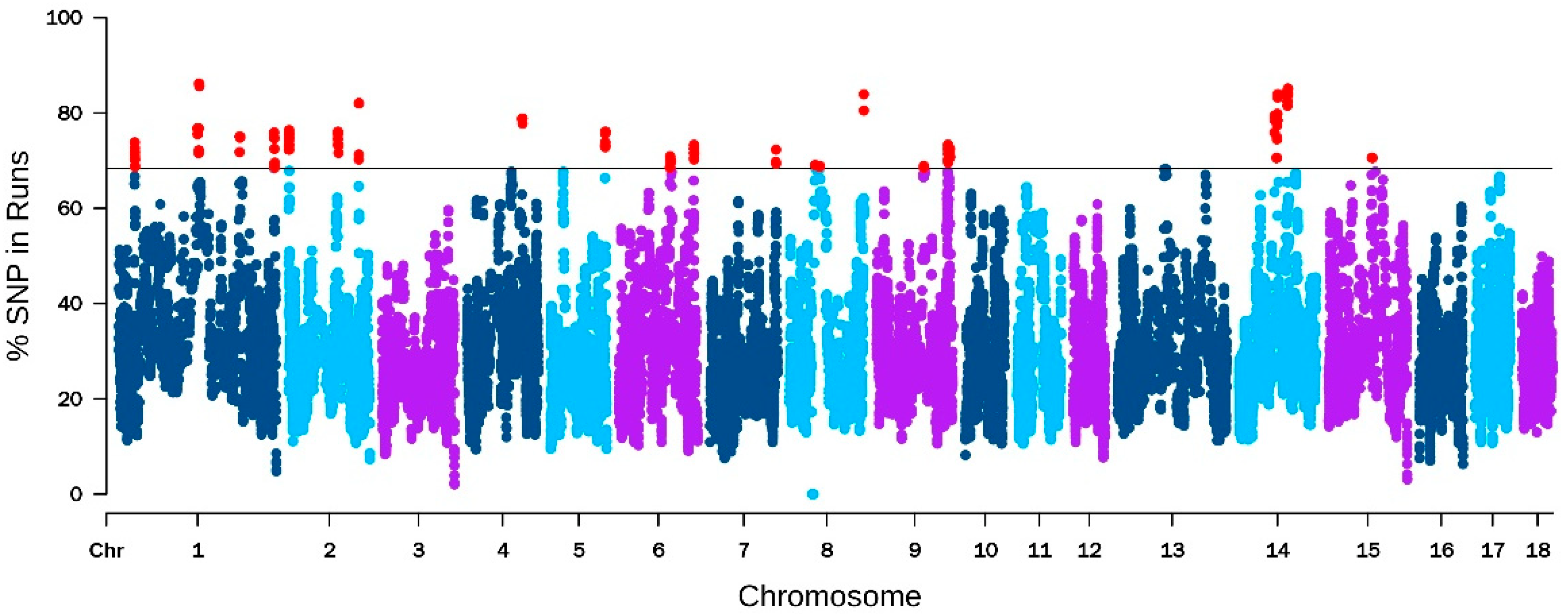
2.6. Detection of ROH Segments and Common Runs of Homozygosity
3. Results and Discussion
3.1. Correlation Analysis between Phenotypes
3.2. PCA and Kinship Analysis
3.3. The Significant SNPs and Genes from GWAS
3.4. ROH Detection and Candidate Genes within Runs of Homozygosity Islands
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Onteru, S.K.; Fan, B.; Du, Z.Q.; Garrick, D.J.; Stalder, K.J.; Rothschild, M.F. A whole-genome association study for pig reproductive traits. Anim. Genet. 2012, 43, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Wang, K.; Zhou, J.; Chen, D.; Yang, Q.; Yang, X.; Liu, Y.; Feng, B.; Jiang, A.; Shen, L.; et al. GWAS on Imputed Whole-Genome Resequencing From Genotyping-by-Sequencing Data for Farrowing Interval of Different Parities in Pigs. Front. Genet. 2019, 10, 1012. [Google Scholar] [CrossRef] [PubMed]
- Melo, T.; Fortes, M.; Bresolin, T.; Mota, L.; Albuquerque, L.; Carvalheiro, R. Multitrait meta-analysis identified genomic regions associated with sexual precocity in tropical beef cattle. J. Anim. Sci. 2018, 96, 4087–4099. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Rempel, L.; Rohrer, G. Genome-wide association study of swine farrowing traits. Part I: Genetic and genomic parameter estimates. J. Anim. Sci. 2012, 90, 3353–3359. [Google Scholar] [CrossRef] [PubMed]
- Leman, A. Optimizing farrowing rate and litter size and minimizing nonproductive sow days. Vet. Clin. N. Am. Food Anim. Pract. 1992, 8, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Vashi, Y.; Magotra, A.; Kalita, D.; Banik, S.; Sahoo, N.; Gupta, S.; Naskar, S. Evaluation of candidate genes related to litter traits in Indian pig breeds. Reprod. Domest. Anim. Zuchthyg. 2021, 56, 577–585. [Google Scholar] [CrossRef]
- Ding, R.; Qiu, Y.; Zhuang, Z.; Ruan, D.; Wu, J.; Zhou, S.; Ye, J.; Cao, L.; Hong, L.; Xu, Z.; et al. Genome-wide association studies reveals polygenic genetic architecture of litter traits in Duroc pigs. Theriogenology 2021, 173, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Wang, K.; Yang, Q.; Zhou, J.; Chen, D.; Ma, J.; Tang, Q.; Jin, L.; Xiao, W.; Jiang, A.; et al. Identifying SNPs and candidate genes for three litter traits using single-step GWAS across six parities in Landrace and Large White pigs. Physiol. Genom. 2018, 50, 1026–1035. [Google Scholar] [CrossRef]
- Schiavo, G.; Bovo, S.; Tinarelli, S.; Bertolini, F.; Dall’Olio, S.; Gallo, M.; Fontanesi, L. Genome-wide association analyses for several exterior traits in the autochthonous Casertana pig breed. Livest. Sci. 2019, 230, 103842. [Google Scholar] [CrossRef]
- Tam, V.; Patel, N.; Turcotte, M.; Bosse, Y.; Pare, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef]
- Zhuang, Z.; Ding, R.; Peng, L.; Wu, J.; Ye, Y.; Zhou, S.; Wang, X.; Quan, J.; Zheng, E.; Cai, G.; et al. Genome-wide association analyses identify known and novel loci for teat number in Duroc pigs using single-locus and multi-locus models. BMC Genom. 2020, 21, 344. [Google Scholar] [CrossRef]
- Guo, X.; Su, G.; Christensen, O.F.; Janss, L.; Lund, M.S. Genome-wide association analyses using a Bayesian approach for litter size and piglet mortality in Danish Landrace and Yorkshire pigs. BMC Genom. 2016, 17, 468. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ding, X.; Tan, Z.; Xing, K.; Yang, T.; Wang, Y.; Sun, D.; Wang, C. Genome-wide association study for reproductive traits in a Large White pig population. Anim. Genet. 2018, 49, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ye, S.; Teng, J.; Diao, S.; Yuan, X.; Chen, Z.; Zhang, H.; Li, J.; Zhang, Z. Genome-wide association studies for the number of animals born alive and dead in duroc pigs. Theriogenology 2019, 139, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wilkinson, J.; Wang, Z.; Ladinig, A.; Harding, J.; Plastow, G. A genome-wide association study of fetal response to type 2 porcine reproductive and respiratory syndrome virus challenge. Sci. Rep. 2016, 6, 20305. [Google Scholar] [CrossRef]
- Ceballos, F.; Joshi, P.; Clark, D.; Ramsay, M.; Wilson, J. Runs of homozygosity: Windows into population history and trait architecture. Nat. Rev. Genet. 2018, 19, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Purfield, D.; Berry, D.; McParland, S.; Bradley, D. Runs of homozygosity and population history in cattle. BMC Genet. 2012, 13, 70. [Google Scholar] [CrossRef]
- Peripolli, E.; Munari, D.; Silva, M.; Lima, A.; Irgang, R.; Baldi, F. Runs of homozygosity: Current knowledge and applications in livestock. Anim. Genet. 2017, 48, 255–271. [Google Scholar] [CrossRef]
- Xu, Z.; Mei, S.; Zhou, J.; Zhang, Y.; Qiao, M.; Sun, H.; Li, Z.; Li, L.; Dong, B.; Oyelami, F.; et al. Genome-Wide Assessment of Runs of Homozygosity and Estimates of Genomic Inbreeding in a Chinese Composite Pig Breed. Front. Genet. 2021, 12, 720081. [Google Scholar] [CrossRef]
- Peripolli, E.; Stafuzza, N.; Munari, D.; Lima, A.; Irgang, R.; Machado, M.; Panetto, J.; Ventura, R.; Baldi, F.; da Silva, M. Assessment of runs of homozygosity islands and estimates of genomic inbreeding in Gyr (Bos indicus) dairy cattle. BMC Genom. 2018, 19, 34. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, Q.; Xiao, Q.; Sun, H.; Gao, H.; Yang, Y.; Chen, J.; Li, Z.; Xue, M.; Ma, P.; et al. Distribution of runs of homozygosity in Chinese and Western pig breeds evaluated by reduced-representation sequencing data. Anim. Genet. 2018, 49, 579–591. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, J.B.; Xue, Y.; Peng, Y.L.; Chen, G.; Fang, M.Y. Differential expression of CYB5A in Chinese and European pig breeds due to genetic variations in the promoter region. Anim. Genet. 2015, 46, 16–22. [Google Scholar] [CrossRef]
- Meng, Q.; Wang, K.; Liu, X.; Zhou, H.; Xu, L.; Wang, Z.; Fang, M. Identification of growth trait related genes in a Yorkshire purebred pig population by genome-wide association studies. Asian-Australas. J. Anim. Sci. 2017, 30, 462–469. [Google Scholar] [CrossRef]
- Price, A.; Patterson, N.; Plenge, R.; Weinblatt, M.; Shadick, N.; Reich, D. Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 2006, 38, 904–909. [Google Scholar] [CrossRef]
- Listgarten, J.; Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLOS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Fang, Y.; Hao, X.; Xu, Z.; Sun, H.; Zhao, Q.; Cao, R.; Zhang, Z.; Ma, P.; Sun, Y.; Qi, Z.; et al. Genome-Wide Detection of Runs of Homozygosity in Laiwu Pigs Revealed by Sequencing Data. Front. Genet. 2021, 12, 629966. [Google Scholar] [CrossRef]
- Pemberton, T.; Absher, D.; Feldman, M.; Myers, R.; Rosenberg, N.; Li, J. Genomic patterns of homozygosity in worldwide human populations. Am. J. Hum. Genet. 2012, 91, 275–292. [Google Scholar] [CrossRef]
- Dixit, S.; Singh, S.; Ganguly, I.; Bhatia, A.; Sharma, A.; Kumar, N.; Dang, A.; Jayakumar, S. Bos indicusGenome-Wide Runs of Homozygosity Revealed Selection Signatures in. Front. Genet. 2020, 11, 92. [Google Scholar] [CrossRef]
- Liu, L.; Zhou, J.; Chen, C.; Zhang, J.; Wen, W.; Tian, J.; Zhang, Z.; Gu, Y. GWAS-Based Identification of New Loci for Milk Yield, Fat, and Protein in Holstein Cattle. Animals 2020, 10, 2048. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, H.; Chen, X.; Wang, Y.; Huang, S.; Chen, B.; Zhang, C.; Hou, B. Identification of LIPH as an unfavorable biomarkers correlated with immune suppression or evasion in pancreatic cancer based on RNA-seq. Cancer Immunol. Immunother. CII 2022, 71, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, C.; Chen, H.; Li, R.; Chong, Q.; Xiao, H.; Chen, S. Genome-wide scan of selection signatures in Dehong humped cattle for heat tolerance and disease resistance. Anim. Genet. 2020, 51, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Qu, K.; Ma, X.; Hanif, Q.; Zhang, J.; Liu, J.; Chen, N.; Suolang, Q.; Lei, C.; Huang, B. Whole-Genome Analyses Reveal Genomic Characteristics and Selection Signatures of Lincang Humped Cattle at the China-Myanmar Border. Front. Genet. 2022, 13, 833503. [Google Scholar] [CrossRef]
- Orozco-terWengel, P.; Barbato, M.; Nicolazzi, E.; Biscarini, F.; Milanesi, M.; Davies, W.; Williams, D.; Stella, A.; Ajmone-Marsan, P.; Bruford, M. Revisiting demographic processes in cattle with genome-wide population genetic analysis. Front. Genet. 2015, 6, 191. [Google Scholar] [CrossRef]
- Littauer, E.; Skountzou, I. Hormonal Regulation of Physiology, Innate Immunity and Antibody Response to H1N1 Influenza Virus Infection During Pregnancy. Front. Immunol. 2018, 9, 2455. [Google Scholar] [CrossRef]
- McGinnis, L.; Albertini, D.; Kinsey, W. Localized activation of Src-family protein kinases in the mouse egg. Dev. Biol. 2007, 306, 241–254. [Google Scholar] [CrossRef]
- Luo, J.; Gupta, V.; Kern, B.; Tash, J.; Sanchez, G.; Blanco, G.; Kinsey, W. Role of FYN kinase in spermatogenesis: Defects characteristic of Fyn-null sperm in mice. Biol. Reprod. 2012, 86, 1–8. [Google Scholar] [CrossRef]
- Qiu, Y.; Yang, H.; Li, C.; Xu, C. Progress in Research on Sperm DNA Fragmentation. Med. Sci. Monit. 2020, 26, e918746. [Google Scholar] [CrossRef]
- Zini, A.; Libman, J. Sperm DNA damage: Clinical significance in the era of assisted reproduction. CMAJ 2006, 175, 495–500. [Google Scholar] [CrossRef][Green Version]
- Elliott, M.; Zheng, S.; Park, D.; Woodson, R.; Reardon, M.; Juncadella, I.; Kinchen, J.; Zhang, J.; Lysiak, J.; Ravichandran, K. Unexpected requirement for ELMO1 in clearance of apoptotic germ cells in vivo. Nature 2010, 467, 333–337. [Google Scholar] [CrossRef]
- Elliott, M.; Ravichandran, K. ELMO1 signaling in apoptotic germ cell clearance and spermatogenesis. Ann. N. Y. Acad. Sci. 2010, 1209, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Klaffenbach, D.; Friedrich, D.; Strick, R.; Strissel, P.L.; Beckmann, M.W.; Rascher, W.; Gessner, A.; Dotsch, J.; Meissner, U.; Schnare, M. Contribution of different placental cells to the expression and stimulation of antimicrobial proteins (AMPs). Placenta 2011, 32, 830–837. [Google Scholar] [CrossRef] [PubMed]
- Vanwong, N.; Sukasem, C.; Unaharassamee, W.; Jiratjintana, N.; Na Nakorn, C.; Hongkaew, Y.; Puangpetch, A. SREBF2Associations of the Gene and Polymorphisms with Obesity and Dyslipidemia in Thai Psychotic Disorder Patients Treated with Risperidone. J. Pers. Med. 2021, 11, 943. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, Y.; Qiu, L.; Miao, Y. Negative effect of insulin-induced gene 2 on milk fat synthesis in buffalo mammary epithelial cells. J. Dairy Res. 2021, 88, 401–406. [Google Scholar] [CrossRef]
- Wang, F.; Zhou, Z.; Ren, X.; Wang, Y.; Yang, R.; Luo, J.; Strappe, P. Effect of Ganoderma lucidum spores intervention on glucose and lipid metabolism gene expression profiles in type 2 diabetic rats. Lipids Health Dis. 2015, 14, 49. [Google Scholar] [CrossRef]
- Tint, G.; Yu, H.; Shang, Q.; Xu, G.; Patel, S. The use of the Dhcr7 knockout mouse to accurately determine the origin of fetal sterols. J. Lipid Res. 2006, 47, 1535–1541. [Google Scholar] [CrossRef]
- Shehu, A.; Mao, J.; Gibori, G.; Halperin, J.; Le, J.; Devi, Y.; Merrill, B.; Kiyokawa, H.; Gibori, G. Prolactin receptor-associated protein/17beta-hydroxysteroid dehydrogenase type 7 gene (Hsd17b7) plays a crucial role in embryonic development and fetal survival. Mol. Endocrinol. 2008, 22, 2268–2277. [Google Scholar] [CrossRef]
- Herrera, E. Metabolic adaptations in pregnancy and their implications for the availability of substrates to the fetus. Eur. J. Clin. Nutr. 2000, 54 (Suppl. 1), S47–S51. [Google Scholar] [CrossRef]
- Woollett, L. Review: Transport of maternal cholesterol to the fetal circulation. Placenta 2011, 32, S218–S221. [Google Scholar] [CrossRef]
- Anand-Ivell, R.; Dai, Y.; Ivell, R. Neohormones as biomarkers of reproductive health. Fertil. Steril. 2013, 99, 1153–1160. [Google Scholar] [CrossRef]
- Burnicka-Turek, O.; Shirneshan, K.; Paprotta, I.; Grzmil, P.; Meinhardt, A.; Engel, W.; Adham, I. Inactivation of insulin-like factor 6 disrupts the progression of spermatogenesis at late meiotic prophase. Endocrinology 2009, 150, 4348–4357. [Google Scholar] [CrossRef]
- Zhang, Q.; Ji, S.; Busayavalasa, K.; Shao, J.; Yu, C. Meiosis I progression in spermatogenesis requires a type of testis-specific 20S core proteasome. Nat. Commun. 2019, 10, 3387. [Google Scholar] [CrossRef]
- Gómez-H, L.; Felipe-Medina, N.; Condezo, Y.; Garcia-Valiente, R.; Ramos, I.; Suja, J.; Barbero, J.; Roig, I.; Sánchez-Martín, M.; de Rooij, D.; et al. The PSMA8 subunit of the spermatoproteasome is essential for proper meiotic exit and mouse fertility. PLoS Genet. 2019, 15, e1008316. [Google Scholar] [CrossRef]
- Yuan, S.; Stratton, C.; Bao, J.; Zheng, H.; Bhetwal, B.; Yanagimachi, R.; Yan, W. Spata6 is required for normal assembly of the sperm connecting piece and tight head-tail conjunction. Proc. Natl. Acad. Sci. USA 2015, 112, E430–E439. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, E.; Seymour, K.; Sigrist, K.; Rooij, D.; Freiman, R. ZFP628 Is a TAF4b-Interacting Transcription Factor Required for Mouse Spermiogenesis. Mol. Cell. Biol. 2020, 40, e00228-19. [Google Scholar] [CrossRef]
- Cao, Q.; Chen, J.; Zhu, L.; Liu, Y.; Zhou, Z.; Sha, J.; Wang, S.; Li, J. A testis-specific and testis developmentally regulated tumor protein D52 (TPD52)-like protein TPD52L3/hD55 interacts with TPD52 family proteins. Biochem. Biophys. Res. Commun. 2006, 344, 798–806. [Google Scholar] [CrossRef]
- Li, S.; Lan, K.; Hung, H.; Huang, W.; Lai, Y.; Cheng, H.; Tsai, C.; Huang, K.; You, H.; Hsu, T. HSPA4 Is a Biomarker of Placenta Accreta and Enhances the Angiogenesis Ability of Vessel Endothelial Cells. Int. J. Mol. Sci. 2022, 23, 5682. [Google Scholar] [CrossRef]
- Ye, B.; Liu, B.; Hao, L.; Zhu, X.; Yang, L.; Wang, S.; Xia, P.; Du, Y.; Meng, S.; Huang, G.; et al. Klf4 glutamylation is required for cell reprogramming and early embryonic development in mice. Nat. Commun. 2018, 9, 1261. [Google Scholar] [CrossRef]
- Lancaster, M.; Gopal, D.; Kim, J.; Saleem, S.; Silhavy, J.; Louie, C.; Thacker, B.; Williams, Y.; Zaki, M.; Gleeson, J. Defective Wnt-dependent cerebellar midline fusion in a mouse model of Joubert syndrome. Nat. Med. 2011, 17, 726–731. [Google Scholar] [CrossRef]
- Ouseph, M.; Li, J.; Chen, H.; Pécot, T.; Wenzel, P.; Thompson, J.; Comstock, G.; Chokshi, V.; Byrne, M.; Forde, B.; et al. Atypical E2F repressors and activators coordinate placental development. Dev. Cell 2012, 22, 849–862. [Google Scholar] [CrossRef]
- Li, J.; Ran, C.; Li, E.; Gordon, F.; Comstock, G.; Siddiqui, H.; Cleghorn, W.; Chen, H.; Kornacker, K.; Liu, C.; et al. Synergistic function of E2F7 and E2F8 is essential for cell survival and embryonic development. Dev. Cell 2008, 14, 62–75. [Google Scholar] [CrossRef]
- Fujioka, K.; Nishida, K.; Ashina, M.; Abe, S.; Fukushima, S.; Ikuta, T.; Ohyama, S.; Morioka, I.; Iijima, K. DNA methylation of the Rtl1 promoter in the placentas with fetal growth restriction. Pediatr. Neonatol. 2019, 60, 512–516. [Google Scholar] [CrossRef]
- Suntharalingham, J.; Ishida, M.; Buonocore, F.; Del Valle, I.; Solanky, N.; Demetriou, C.; Regan, L.; Moore, G.; Achermann, J. CDKN1CAnalysis of in fetal growth restriction and pregnancy loss. F1000Research 2019, 8, 90. [Google Scholar] [CrossRef]
- Belli, M.; Shimasaki, S. Molecular Aspects and Clinical Relevance of GDF9 and BMP15 in Ovarian Function. Vitam. Horm. 2018, 107, 317–348. [Google Scholar] [CrossRef]
- Hobeika, E.; Armouti, M.; Fierro, M.; Winston, N.; Scoccia, H.; Zamah, A.; Stocco, C. Regulation of Insulin-Like Growth Factor 2 by Oocyte-Secreted Factors in Primary Human Granulosa Cells. J. Clin. Endocrinol. Metab. 2020, 105, 327–335. [Google Scholar] [CrossRef]
- Labade, A.S.; Karmodiya, K.; Sengupta, K. HOXA repression is mediated by nucleoporin Nup93 assisted by its interactors Nup188 and Nup205. Epigenetics Chromatin 2016, 9, 54. [Google Scholar] [CrossRef]
- Broere-Brown, Z.; Schalekamp-Timmermans, S.; Jaddoe, V.; Steegers, E. Deceleration of fetal growth rate as alternative predictor for childhood outcomes: A birth cohort study. BMC Pregnancy Childbirth 2019, 19, 216. [Google Scholar] [CrossRef]
- Bansal, S.; Gupta, N.; Sankhwar, S.; Rajender, S. Differential Genes Expression between Fertile and Infertile Spermatozoa Revealed by Transcriptome Analysis. PLoS ONE 2015, 10, e0127007. [Google Scholar] [CrossRef]
- Lei, X.; Callaway, M.; Zhou, H.; Yang, Y.; Chen, W. Obesity associated Lyplal1 gene is regulated in diet induced obesity but not required for adipocyte differentiation. Mol. Cell. Endocrinol. 2015, 411, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Bellver, J.; Pellicer, A.; García-Velasco, J.; Ballesteros, A.; Remohí, J.; Meseguer, M. Obesity reduces uterine receptivity: Clinical experience from 9,587 first cycles of ovum donation with normal weight donors. Fertil. Steril. 2013, 100, 1050–1058. [Google Scholar] [CrossRef]
- Campos, P.; Silva, B.; Donzele, J.; Oliveira, R.; Knol, E. Effects of sow nutrition during gestation on within-litter birth weight variation: A review. Animal 2012, 6, 797–806. [Google Scholar] [CrossRef]
- Tenenbaum-Gavish, K.; Hod, M. Impact of maternal obesity on fetal health. Fetal Diagn. Ther. 2013, 34, 1–7. [Google Scholar] [CrossRef]
- Dong, R.; Gao, S.; Shan, M. Identification of the similarly expressed genes in patients with polycystic ovary syndrome and transsexuals. Medicine 2021, 100, e26990. [Google Scholar] [CrossRef]
- Tachibanaki, S.; Arinobu, D.; Shimauchi-Matsukawa, Y.; Tsushima, S.; Kawamura, S. Highly effective phosphorylation by G protein-coupled receptor kinase 7 of light-activated visual pigment in cones. Proc. Natl. Acad. Sci. USA 2005, 102, 9329–9334. [Google Scholar] [CrossRef]
- Colas, E.; Pedrola, N.; Devis, L.; Ertekin, T.; Campoy, I.; Martinez, E.; Llaurado, M.; Rigau, M.; Olivan, M.; Garcia, M.; et al. The EMT signaling pathways in endometrial carcinoma. Clin. Transl. Oncol. 2012, 14, 715–720. [Google Scholar] [CrossRef]
- Jamsai, D.; Clark, B.J.; Smith, S.J.; Whittle, B.; Goodnow, C.C.; Ormandy, C.J.; O’Bryan, M.K. A missense mutation in the transcription factor ETV5 leads to sterility, increased embryonic and perinatal death, postnatal growth restriction, renal asymmetry and polydactyly in the mouse. PLoS ONE 2013, 8, e77311. [Google Scholar] [CrossRef]
- Tyagi, G.; Carnes, K.; Morrow, C.; Kostereva, N.; Ekman, G.; Meling, D.; Hostetler, C.; Griswold, M.; Murphy, K.; Hess, R.; et al. Loss of Etv5 decreases proliferation and RET levels in neonatal mouse testicular germ cells and causes an abnormal first wave of spermatogenesis. Biol. Reprod. 2009, 81, 258–266. [Google Scholar] [CrossRef]
- Malavolta, M.; Costarelli, L.; Giacconi, R.; Basso, A.; Piacenza, F.; Pierpaoli, E.; Provinciali, M.; Ogo, O.A.; Ford, D. Changes in Zn homeostasis during long term culture of primary endothelial cells and effects of Zn on endothelial cell senescence. Exp. Gerontol. 2017, 99, 35–45. [Google Scholar] [CrossRef]
- Kumar, L.; Michalczyk, A.; McKay, J.; Ford, D.; Kambe, T.; Hudek, L.; Varigios, G.; Taylor, P.E.; Ackland, M.L. Altered expression of two zinc transporters, SLC30A5 and SLC30A6, underlies a mammary gland disorder of reduced zinc secretion into milk. Genes Nutr. 2015, 10, 487. [Google Scholar] [CrossRef]
- Brion, L.; Heyne, R.; Lair, C. Role of zinc in neonatal growth and brain growth: Review and scoping review. Pediatr. Res. 2021, 89, 1627–1640. [Google Scholar] [CrossRef] [PubMed]
- Staub, E.; Evers, K.; Askie, L. Enteral zinc supplementation for prevention of morbidity and mortality in preterm neonates. Cochrane Database Syst. Rev. 2021, 3, CD012797. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, S.; Zhan, F.; Song, M.; Shang, P.; Zhu, F.; Li, J.; Yang, F.; Li, X.; Qiao, R.; et al. Population Genetic Analysis of Six Chinese Indigenous Pig Meta-Populations Based on Geographically Isolated Regions. Animals 2023, 13, 1396. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Shi, L.; Liu, J.; Deng, T.; Wang, L.; Liu, Y.; Zhao, F. Genome-Wide Scan for Runs of Homozygosity Identifies Candidate Genes in Three Pig Breeds. Animals 2019, 9, 518. [Google Scholar] [CrossRef] [PubMed]
- Lovasco, L.; Gustafson, E.; Seymour, K.; de Rooij, D.; Freiman, R. TAF4b is required for mouse spermatogonial stem cell development. Stem Cells 2015, 33, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Falender, A.; Freiman, R.; Geles, K.; Lo, K.; Hwang, K.; Lamb, D.; Morris, P.; Tjian, R.; Richards, J. Maintenance of spermatogenesis requires TAF4b, a gonad-specific subunit of TFIID. Genes Dev. 2005, 19, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Al-Mutar, H.; Younis, L. Effect of Point Mutation in the Growth Differentiation Factor 9 Gene of Oocytes on the Sterility and Fertility of Awassi Sheep. Arch. Razi Inst. 2020, 75, 101–108. [Google Scholar] [CrossRef]
- Wang, F.; Chu, M.; Pan, L.; Wang, X.; He, X.; Zhang, R.; Tao, L.; La, Y.; Ma, L.; Di, R. Polymorphism Detection of GDF9 Gene and Its Association with Litter Size in Luzhong Mutton Sheep (Ovis aries). Animals 2021, 11, 571. [Google Scholar] [CrossRef]
- Das, A.; Shaha, M.; Gupta, M.; Dutta, A.; Miazi, O. Polymorphism of fecundity genes (BMP15 and GDF9) and their association with litter size in Bangladeshi prolific Black Bengal goat. Trop. Anim. Health Prod. 2021, 53, 230. [Google Scholar] [CrossRef]
- Li, Y.; Jin, W.; Wang, Y.; Zhang, J.; Meng, C.; Wang, H.; Qian, Y.; Li, Q.; Cao, S. GDF9 Three Complete Linkage SNPs of Gene Affect the Litter Size Probably Mediated by OCT1 in Hu Sheep. DNA Cell Biol. 2020, 39, 563–571. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | SNP | Ref SNP | SSC | p Value | Candidate Genes | Distance (bp) |
|---|---|---|---|---|---|---|
| TNB | ASGA0079603 | rs81468948 | 18 | 6.49 × 10−6 | ELMO1 AOAH | Within 343,972 |
| TNB | DRGA0015019 | rs81301323 | 15 | 8.10 × 10−6 | INSIG2 | Within |
| TNB | ALGA0084417 | rs81451913 | 15 | 1.10 × 10−5 | INSIG2 | 203,311 |
| WEAK | ALGA0097098 | rs81471717 | 18 | 1.57 × 10−5 | NUP205 | 16,550 |
| WEAK | ASGA0078964 | rs81471771 | 18 | 1.57 × 10−5 | FAM180A | 24,824 |
| WEAK | H3GA0050399 | rs81471732 | 18 | 1.57 × 10−5 | FAM180A | 26,006 |
| FREAK | ALGA0056905 | rs81428907 | 10 | 2.15 × 10−6 | LYPLAL1 | 252,956 |
| FREAK | WU_10.2_8_121864282 | rs328492172 | 8 | 6.98 × 10−6 | RPL34 | 154,848 |
| FREAK | DRGA0010254 | rs81302520 | 10 | 7.24 × 10−6 | LYPLAL1 | 219,860 |
| DEAD | ALGA0071870 | rs80920882 | 13 | 9.56 × 10−7 | LIPH ENSSSCG00000051486 | Within 73,460 |
| DEAD | ASGA0100324 | rs81321047 | 13 | 9.27 × 10−6 | ENSSSCG00000047980 | 248,227 |
| DEAD | ASGA0058420 | rs80872186 | 13 | 1.18 × 10−5 | RNF7 | 41,373 |
| DEAD | ASGA0058424 | rs81447229 | 13 | 1.18 × 10−5 | GRK7 | Within |
| DEAD | ASGA0099314 | rs81319768 | 13 | 1.23 × 10−5 | ETV5 | Within |
| DEAD | MARC0004732 | rs81232688 | 13 | 1.23 × 10−5 | ETV5 | Within |
| DEAD | SIRI0000314 | rs329411938 | 13 | 1.47 × 10−5 | TPRG1 | 26,006 |
| MUMMY | MARC0022221 | rs81290729 | 1 | 1.88 × 10−7 | FYN | Within |
| MUMMY | WU_10.2_13_204127273 | rs324169348 | 13 | 2.48 × 10−6 | CLDN17 | 9458 |
| MUMMY | ASGA0004838 | rs81348947 | 1 | 3.02 × 10−6 | TAFA5 | 92,417 |
| MUMMY | MARC0028125 | rs81223068 | 16 | 3.90 × 10−6 | SLC30A5 | 147,756 |
| CHR | Start (bp) | End (bp) | Gene Symbol | SNP % | Function |
|---|---|---|---|---|---|
| 1 | 216,792,785 | 216,820,197 | INSL6 | 75.04 | Ensures sperm motility [49,50] |
| 6 | 110,799,065 | 110,836,937 | PSMA8 | 69.91 | Component of the sperm proteasome [51,52] |
| 6 | 163,278,954 | 163,410,174 | SPATA6 | 71.62 | Connects the sperm head and tail during spermatogenesis [53] |
| 6 | 110,855,636 | 110,972,798 | TAF4B | 69.91 | Plays a role in spermatogenesis and oogenesis [54] |
| 1 | 215,841,463 | 215,842,606 | TPD52L3 | 71.79 | Potential role in testis development and spermatogenesis [55] |
| 14 | 71,126,100 | 71,269,517 | HERC4 | 83.93 | Necessary for sperm maturation [56] |
| 6 | 161,953,495 | 163,216,282 | AGBL4 | 73.33 | Regulates embryogenesis [57] |
| 1 | 28,566,279 | 28,760,471 | AHI1 | 71.79 | Plays a crucial role in ciliary signaling during cerebellum embryonic development [58] |
| 5 | 104,126,924 | 104,163,582 | E2F7 | 76.07 | Critical for development of the fetus [59,60] |
| 7 | 121,706,955 | 121,720,472 | RTL1 | 69.40 | Maintains fetal and maternal interface and placental development [61] |
| 2 | 2,019,391 | 2,022,092 | CDKN1C | 72.99 | Causes fetal dysplasia and miscarriage [62] |
| 2 | 135,181,494 | 135,184,451 | GDF9 | 82.05 | Regulates the quality and developmental ability of eggs [63,64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Zhang, S.; Yuan, M.; Zhan, F.; Song, M.; Shang, P.; Yang, F.; Li, X.; Qiao, R.; Han, X.; et al. Genome-Wide Association Studies and Runs of Homozygosity to Identify Reproduction-Related Genes in Yorkshire Pig Population. Genes 2023, 14, 2133. https://doi.org/10.3390/genes14122133
Zhang L, Zhang S, Yuan M, Zhan F, Song M, Shang P, Yang F, Li X, Qiao R, Han X, et al. Genome-Wide Association Studies and Runs of Homozygosity to Identify Reproduction-Related Genes in Yorkshire Pig Population. Genes. 2023; 14(12):2133. https://doi.org/10.3390/genes14122133
Chicago/Turabian StyleZhang, Lige, Songyuan Zhang, Meng Yuan, Fengting Zhan, Mingkun Song, Peng Shang, Feng Yang, Xiuling Li, Ruimin Qiao, Xuelei Han, and et al. 2023. "Genome-Wide Association Studies and Runs of Homozygosity to Identify Reproduction-Related Genes in Yorkshire Pig Population" Genes 14, no. 12: 2133. https://doi.org/10.3390/genes14122133
APA StyleZhang, L., Zhang, S., Yuan, M., Zhan, F., Song, M., Shang, P., Yang, F., Li, X., Qiao, R., Han, X., Li, X., Fang, M., & Wang, K. (2023). Genome-Wide Association Studies and Runs of Homozygosity to Identify Reproduction-Related Genes in Yorkshire Pig Population. Genes, 14(12), 2133. https://doi.org/10.3390/genes14122133