
Exploring the Application Potential of Aquaculture Sewage Treatment of Pseudomonas chengduensis Strain WD211 Based on Its Complete Genome

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sewage Treatment Capacity
2.2. Antibiotic Susceptibility Tests
2.3. pH and Salt Tolerance
2.4. Genome Sequencing and Annotation
2.5. Whole-Genome Sequence Alignments
3. Results
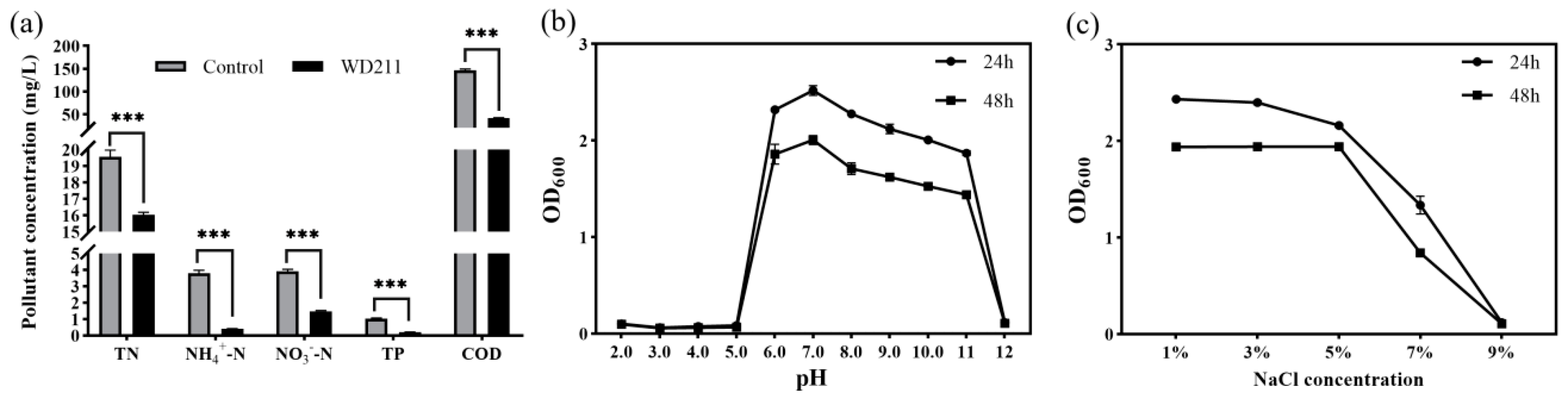
3.1. Effect of Strain WD211 on Removing Pollutants from Fish-Pond Aquaculture Wastewater
3.2. pH and Salt Tolerance Ranges of Strain WD211
3.3. Antibiotic Susceptibility of Strain WD211
3.4. General Features of Genome
3.5. Genomic Insights into Sewage Treatment
3.5.1. Genes Related to Nitrogen and Phosphorus Removal
3.5.2. Genes Related to Organic Matter Degradation
3.6. Genomic Insight into Stress Resistance
3.6.1. Genes Related to Antibiotic Resistance
3.6.2. Genes Related to Salt Resistance
3.6.3. Genes Related to the Two-Component System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, Y.; Ciais, P.; Goll, D.S.; Sardans, J.; Peñuelas, J.; Cresto-Aleina, F.; Zhang, H. The shift of phosphorus transfers in global fisheries and aquaculture. Nat. Commun. 2020, 11, 355. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Lee, J.W.; Chandran, K.; Kim, S.; Khanal, S.K. Nitrous oxide (N2O) emission from aquaculture: A review. Environ. Sci. Technol. 2012, 46, 6470–6480. [Google Scholar] [CrossRef] [PubMed]
- van Rijn, J. Waste treatment in recirculating aquaculture systems. Aquac. Eng. 2013, 53, 49–56. [Google Scholar] [CrossRef]
- Chai, Y.; Peng, R.; Jiang, M.; Jiang, X.; Han, Q.; Han, Z. Effects of ammonia nitrogen stress on the blood cell immunity and liver antioxidant function of Sepia pharaonis. Aquaculture 2022, 546, 737417. [Google Scholar] [CrossRef]
- Zainab, S.M.; Junaid, M.; Xu, N.; Malik, R.N. Antibiotics and antibiotic resistant genes (ARGs) in groundwater: A global review on dissemination, sources, interactions, environmental and human health risks. Water Res. 2020, 187, 116455. [Google Scholar] [CrossRef]
- Liang, D.H.; Hu, Y.Y.; Cheng, J.H.; Chen, Y.C. Simultaneous sulfamethoxazole biodegradation and nitrogen conversion in low C/N ratio pharmaceutical wastewater by Achromobacter sp. JL9. Sci. Total Environ. 2020, 703, 135586. [Google Scholar] [CrossRef]
- Jin, P.; Chen, Y.Y.; Xu, T.; Cui, Z.W.; Zheng, Z.W. Efficient nitrogen removal by simultaneous heterotrophic nitrifying-aerobic denitrifying bacterium in a purification tank bioreactor amended with two-stage dissolved oxygen control. Bioresour. Technol. 2019, 281, 392–400. [Google Scholar] [CrossRef]
- Chen, J.; Sun, R.; Pan, C.; Sun, Y.; Mai, B.; Li, Q.X. Antibiotics and Food Safety in Aquaculture. J. Agric. Food Chem. 2020, 68, 11908–11919. [Google Scholar] [CrossRef]
- Wu, L.; Wei, Q.; Zhang, Y.; Fan, Y.; Li, M.; Rong, L.; Xiao, X.; Huang, X.; Zou, X. Effects of antibiotics on enhanced biological phosphorus removal and its mechanisms. Sci. Total Environ. 2021, 774, 145571. [Google Scholar] [CrossRef]
- Peix, A.; Ramírez-Bahena, M.H.; Velázquez, E. The current status on the taxonomy of Pseudomonas revisited: An update. Infect. Genet. Evol. 2018, 57, 106–116. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [PubMed]
- Saati-Santamaría, Z.; Baroncelli, R.; Rivas, R.; García-Fraile, P. Comparative Genomics of the Genus Pseudomonas Reveals Host- and Environment-Specific Evolution. Microbiol. Spectr. 2022, 10, e0237022. [Google Scholar] [CrossRef] [PubMed]
- Kchouk, M.; Gibrat, J.F.; Elloumi, M. Generations of sequencing technologies: From first to next generation. Biol. Med. 2017, 9, 395. [Google Scholar] [CrossRef]
- Tao, Y.; Zhou, Y.; He, X.; Hu, X.; Li, D. Pseudomonas chengduensis sp. nov., isolated from landfill leachate. Int. J. Syst. Evol. Microbiol. 2014, 64, 95–100. [Google Scholar] [CrossRef]
- Shi, Z.; Yu, X.L.; Duan, J.; Guo, W. The complete genome sequence of Pseudomonas chengduensis BC1815 for genome mining of PET degrading enzymes. Mar. Genom. 2023, 67, 101008. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, D.; Gao, P.; Gu, W.; He, X.; Yang, W.; Tang, W. Analysis of biosorption and biotransformation mechanism of Pseudomonas chengduensis strain MBR under Cd(II) stress from genomic perspective. Ecotoxicol. Environ. Saf. 2020, 198, 110655. [Google Scholar] [CrossRef]
- Yi, M.; Wang, H.; Ma, X.; Wang, C.; Wang, M.; Liu, Z.; Lu, M.; Cao, J.; Ke, X. Efficient nitrogen removal of a novel Pseudomonas chengduensis strain BF6 mainly through assimilation in the recirculating aquaculture systems. Bioresour. Technol. 2023, 379, 129036. [Google Scholar] [CrossRef]
- State Environmental Protection Administration. Methods for Monitoring and Analysis of Water and Wastewater; Chinese Environmental Science Press: Beijing, China, 2002. [Google Scholar]
- Darling, A.E.; Tritt, A.; Eisen, J.A.; Facciotti, M.T. Mauve assembly metrics. Bioinformatics 2011, 27, 2756–2757. [Google Scholar] [CrossRef]
- Dai, H.; Sun, Y.; Wan, D.; Abbasi, H.N.; Guo, Z.; Geng, H.; Wang, X.; Chen, Y. Simultaneous denitrification and phosphorus removal: A review on the functional strains and activated sludge processes. Sci. Total Environ. 2022, 835, 155409. [Google Scholar] [CrossRef]
- Kumari, S.; Das, S. Bacterial enzymatic degradation of recalcitrant organic pollutants: Catabolic pathways and genetic regulations. Environ. Sci. Pollut. Res. Int. 2023, 30, 79676–79705. [Google Scholar] [CrossRef]
- Gangola, S.; Joshi, S.; Kumar, S.; Pandey, S.C. Comparative analysis of fungal and bacterial enzymes in biodegradation of xenobiotic compounds. In Smart Bioremediation Technologies, 1st ed.; Bhatt, P., Ed.; Academic Press: Cambridge, UK, 2019; pp. 169–189. [Google Scholar]
- Padan, E.; Schuldiner, S. Molecular physiology of the Na+/H+ antiporter in Escherichia coli. J. Exp. Biol. 1994, 196, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, H.; Pu, Q.; Zhang, C.; Chen, Y.; Lin, Z.; Hu, X.; Li, O. Complete genome of Sphingomonas paucimobilis ZJSH1, an endophytic bacterium from Dendrobium officinale with stress resistance and growth promotion potential. Arch. Microbiol. 2023, 205, 132. [Google Scholar] [CrossRef] [PubMed]
- Cayley, S.; Lewis, B.A.; Record, M.T., Jr. Origins of the osmoprotective properties of betaine and proline in Escherichia coli K-12. J. Bacteriol. 1992, 174, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Iturriaga, G.; Suárez, R.; Nova-Franco, B. Trehalose metabolism: From osmoprotection to signaling. Int. J. Mol. Sci. 2009, 10, 3793–3810. [Google Scholar] [CrossRef]
- Tierney, A.R.; Rather, P.N. Roles of two-component regulatory systems in antibiotic resistance. Future Microbiol. 2019, 14, 533–552. [Google Scholar] [CrossRef]
- Pahel, G.; Rothstein, D.M.; Magasanik, B. Complex glnA-glnL-glnG operon of Escherichia coli. J. Bacteriol. 1982, 150, 202–213. [Google Scholar] [CrossRef]
- Baek, J.H.; Kang, Y.J.; Lee, S.Y. Transcript and protein level analyses of the interactions among PhoB, PhoR, PhoU and CreC in response to phosphate starvation in Escherichia coli. FEMS Microbiol. Lett. 2007, 277, 254–259. [Google Scholar] [CrossRef]
- Heermann, R.; Jung, K. The complexity of the ‘simple’ two-component system KdpD/KdpE in Escherichia coli. FEMS Microbiol. Lett. 2010, 304, 97–106. [Google Scholar] [CrossRef]
- Groisman, E.A.; Duprey, A.; Choi, J. How the PhoP/PhoQ System Controls Virulence and Mg2+ Homeostasis: Lessons in Signal Transduction, Pathogenesis, Physiology, and Evolution. Microbiol. Mol. Biol. Rev. 2021, 85, e0017620. [Google Scholar] [CrossRef]
- Yang, B.; Liu, C.; Pan, X.; Fu, W.; Fan, Z.; Jin, Y.; Bai, F.; Cheng, Z.; Wu, W. Identification of Novel PhoP-PhoQ Regulated Genes That Contribute to Polymyxin B Tolerance in Pseudomonas aeruginosa. Microorganisms 2021, 9, 344. [Google Scholar] [CrossRef]
- Chen, D.; Zhao, Y.; Qiu, Y.; Xiao, L.; He, H.; Zheng, D.; Li, X.; Yu, X.; Xu, N.; Hu, X.; et al. CusS-CusR Two-Component System Mediates Tigecycline Resistance in Carbapenem-Resistant Klebsiella pneumoniae. Front. Microbiol. 2020, 10, 3159. [Google Scholar] [CrossRef] [PubMed]
- Hesse, C.; Schulz, F.; Bull, C.T.; Shaffer, B.T.; Yan, Q.; Shapiro, N.; Hassan, K.A.; Varghese, N.; Elbourne, L.D.H.; Paulsen, I.T.; et al. Genome-based evolutionary history of Pseudomonas spp. Environ. Microbiol. 2018, 20, 2142–2159. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, H.; Yang, Z.; Cheng, X.; Zhao, Y.; Qin, L.; Bisseling, T.; Cao, Q.; Willemsen, V. Plant growth-promoting rhizobacterium Pseudomonas sp. CM11 specifically induces lateral roots. New Phytol. 2022, 235, 1575–1588. [Google Scholar] [CrossRef]
- Khare, E.; Arora, N.K. Biosurfactant based formulation of Pseudomonas guariconensis LE3 with multifarious plant growth promoting traits controls charcoal rot disease in Helianthus annus. World J. Microbiol. Biotechnol. 2021, 37, 55. [Google Scholar] [CrossRef] [PubMed]
- Yesankar, P.J.; Patil, A.; Kapley, A.; Qureshi, A. Catalytic resilience of multicomponent aromatic ring-hydroxylating dioxygenases in Pseudomonas for degradation of polycyclic aromatic hydrocarbons. World J. Microbiol. Biotechnol. 2023, 39, 166. [Google Scholar] [CrossRef]
- Li, Z.; Li, L.; Sun, H.; Wang, W.; Yang, Y.; Qi, Z.; Liu, X. Ammonia assimilation: A double-edged sword influencing denitrification of Rhodobacter azotoformans and for nitrogen removal of aquaculture wastewater. Bioresour. Technol. 2022, 345, 126495. [Google Scholar] [CrossRef]
- Zheng, F.; Wang, J.; Xiao, R.; Chai, W.; Xing, D.; Lu, H. Dissolved organic nitrogen in wastewater treatment processes: Transformation, biosynthesis and ecological impacts. Environ. Pollut. 2021, 273, 116436. [Google Scholar] [CrossRef]
- Zhang, M.; Pan, L.; Su, C.; Liu, L.; Dou, L. Simultaneous aerobic removal of phosphorus and nitrogen by a novel salt-tolerant phosphate-accumulating organism and the application potential in treatment of domestic sewage and aquaculture sewage. Sci. Total Environ. 2021, 758, 143580. [Google Scholar] [CrossRef]
- Zhu, B.T.; Chen, S.C.; Zhao, C.G.; Zhong, W.H.; Zeng, R.Y.; Yang, S.P. Effects of Marichromatium gracile YL28 on the nitrogen management in the aquaculture pond water. Bioresour. Technol. 2019, 292, 121917. [Google Scholar] [CrossRef]
- Uygur, A.; Kargi, F. Salt inhibition on biological nutrient removal from saline wastewater in a sequencing batch reactor. Enzym. Microb. Technol. 2004, 34, 313–318. [Google Scholar] [CrossRef]
- Intrasungkha, N.; Keller, J.; Blackall, L.L. Biological nutrient removal efficiency in treatment of saline wastewater. Water Sci. Technol. 1999, 39, 183–190. [Google Scholar] [CrossRef]
- Adenaya, A.; Berger, M.; Brinkhoff, T.; Ribas-Ribas, M.; Wurl, O. Usage of antibiotics in aquaculture and the impact on coastal waters. Mar. Pollut. Bull. 2023, 188, 114645. [Google Scholar] [CrossRef] [PubMed]
- Hatosy, S.M.; Martiny, A.C. The Ocean as a Global Reservoir of Antibiotic Resistance Genes. Appl. Environ. Microbiol. 2015, 81, 7593–7599. [Google Scholar] [CrossRef]
- Minerdi, D.; Zgrablic, I.; Castrignanò, S.; Catucci, G.; Medana, C.; Terlizzi, M.E.; Gribaudo, G.; Gilardi, G.; Sadeghi, S.J. Escherichia coli Overexpressing a Baeyer-Villiger Monooxygenase from Acinetobacter radioresistens Becomes Resistant to Imipenem. Antimicrob. Agents Chemother. 2015, 60, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.K.; Abdelwahab, H.; Del Campo, J.S.M.; Mehra-Chaudhary, R.; Sobrado, P.; Tanner, J.J. The Structure of the Antibiotic Deactivating, N-hydroxylating Rifampicin Monooxygenase. J. Biol. Chem. 2016, 291, 21553–21562. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, D.F.; Istvan, P.; Noronha, E.F.; Quirino, B.F.; Krüger, R.H. New dioxygenase from metagenomic library from Brazilian soil: Insights into antibiotic resistance and bioremediation. Biotechnol. Lett. 2015, 37, 1809–1817. [Google Scholar] [CrossRef]
- Battle, S.E.; Rello, J.; Hauser, A.R. Genomic islands of Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2009, 290, 70–78. [Google Scholar] [CrossRef]
- Bellanger, X.; Payot, S.; Leblond-Bourget, N.; Guédon, G. Conjugative and mobilizable genomic islands in bacteria: Evolution and diversity. FEMS Microbiol. Rev. 2014, 38, 720–760. [Google Scholar] [CrossRef]
- Hall, R.M. Salmonella genomic islands and antibiotic resistance in Salmonella enterica. Future Microbiol. 2010, 5, 1525–1538. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic | Concentration | Inhibition Zone Diameter (mm) | Antibiotic | Concentration | Inhibition Zone Diameter (mm) |
|---|---|---|---|---|---|
| Penicillin | 10 U/piece | 0 | Neomycin | 30 μg/piece | 0 |
| Oxacillin | 1 μg/piece | 8 | Tetracycline | 30 μg/piece | 0 |
| Ampicillin | 10 μg/piece | 8 | Doxycycline | 30 μg/piece | 0 |
| Carbenicillin | 100 μg/piece | 0 | Minocycline | 30 μg/piece | 0 |
| Piperacillin | 100 μg/piece | 28 | Erythrocin | 15 μg/piece | 0 |
| Cefalexin | 30 μg/piece | 0 | Medemycin | 30 μg/piece | 0 |
| Cefazolin | 30 μg/piece | 0 | Norfloxacin | 10 μg/piece | 16 |
| Cefradine | 30 μg/piece | 0 | Ofloxacin | 5 μg/piece | 23 |
| Cefuroxime | 30 μg/piece | 8 | Ciprofloxacin | 5 μg/piece | 15 |
| Ceftazidime | 30 μg/piece | 0 | Vancomycin | 30 μg/piece | 0 |
| Ceftriaxone | 30 μg/piece | 0 | Selectrin | 23.75 μg/piece | 0 |
| Cefoperazone | 75 μg/piece | 0 | Furazolidone | 300 μg/piece | 8 |
| Amikacin | 30 μg/piece | 23 | Chloramphenico | 30 μg/piece | 9 |
| Gentamicin | 30 μg/piece | 0 | Clindamycin | 2 μg/piece | 8 |
| Kanamycin | 30 μg/piece | 0 |
| Features | Chromosome |
|---|---|
| Genome total size(bp) | 5,208,067 |
| G + C content | 62.75% |
| Coding genes | 4734 |
| Plasmid | 0 |
| rRNAs | 12 |
| tRNAs | 67 |
| Pseudogene | 1 |
| CRISPRs | 2 |
| Genomic islands | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, H.; Wu, H.; Gu, W.; Lu, Y.; Qin, H.; You, Y.; Zhou, D.; Wang, D.; Sun, L.; Zhou, C.; et al. Exploring the Application Potential of Aquaculture Sewage Treatment of Pseudomonas chengduensis Strain WD211 Based on Its Complete Genome. Genes 2023, 14, 2107. https://doi.org/10.3390/genes14122107
Peng H, Wu H, Gu W, Lu Y, Qin H, You Y, Zhou D, Wang D, Sun L, Zhou C, et al. Exploring the Application Potential of Aquaculture Sewage Treatment of Pseudomonas chengduensis Strain WD211 Based on Its Complete Genome. Genes. 2023; 14(12):2107. https://doi.org/10.3390/genes14122107
Chicago/Turabian StylePeng, Huanlong, Hangtao Wu, Wenjie Gu, Yusheng Lu, Hongjie Qin, Yi You, Donglai Zhou, Dan Wang, Lili Sun, Changmin Zhou, and et al. 2023. "Exploring the Application Potential of Aquaculture Sewage Treatment of Pseudomonas chengduensis Strain WD211 Based on Its Complete Genome" Genes 14, no. 12: 2107. https://doi.org/10.3390/genes14122107
APA StylePeng, H., Wu, H., Gu, W., Lu, Y., Qin, H., You, Y., Zhou, D., Wang, D., Sun, L., Zhou, C., & Zheng, Y. (2023). Exploring the Application Potential of Aquaculture Sewage Treatment of Pseudomonas chengduensis Strain WD211 Based on Its Complete Genome. Genes, 14(12), 2107. https://doi.org/10.3390/genes14122107