
Linking New Alleles at the Oscillator Loci to Flowering and Expansion of Asian Rice

Abstract
1. Introduction
2. Materials and Methods
2.1. Assessing Allelic Diversity across Oscillator Loci
2.1.1. Defining Alleles from Genomic Data
2.1.2. Analysis of Ten Diverse Nuclear Genomes of O. sativa
2.1.3. Tests of Selection
2.1.4. Additional Surveys of Varieties of O. sativa to Validate Allelic Sequences
2.1.5. Parallel Analysis on Ten Nuclear Genomes of A. thaliana
2.2. Variation of Allelic Expressions in the Paddy Field
2.2.1. Preparation of Populations from RILs
2.2.2. Field Transcriptions of OsCCA1, OsPRR1, and OsPRR37 in a 48-h Interlude
2.2.3. Fixed-Time Transcriptions of OsCCA1 and OsPRR1 over Six Days Prior to Flowering
2.3. Functional Analysis on Mutations/Alleles of Oscillator Genes
2.3.1. Effects of Natural Temperature and Genetic Background on Allelic Expression
2.3.2. Effect of 5′-Deletion in OsPRR1 on Binding Capacity of OsCCA1 via Electrophoretic Mobility Shift Assays (EMSA)
2.3.3. Compatibility Tests of Os Alleles in Mutants (cca1-1 and toc1-1) of A. thaliana
2.4. Identifying Alleles Associated with Flowering Times and Yields
2.5. Distributions of Oscillator Alleles in Traditional Cultivation Regions of O. sativa
2.6. Statistical Analysis
3. Results
3.1. Rebuild of Circadian Clock Revealed by Allelic Diversities at Oscillator Loci of O. sativa
3.1.1. Genomic Survey of Five Oscillator Genes
3.1.2. Validations of Alleles at Loci of OsCCA1, OsPRR37, and OsPRR1
3.2. Tests of Locus-Specific Selection
3.3. Functional Analyses on OsCCA1, OsPRR37, and OsPRR1 Alleles
3.3.1. Two-day Transcription Patterns of Oscillator Genes in the Paddy Field
3.3.2. Allelic Expression Frequently Varies among Daily Environments and Genetic Backgrounds
3.3.3. Mutations in 5′ Regions Can Alter Expression Levels of Alleles
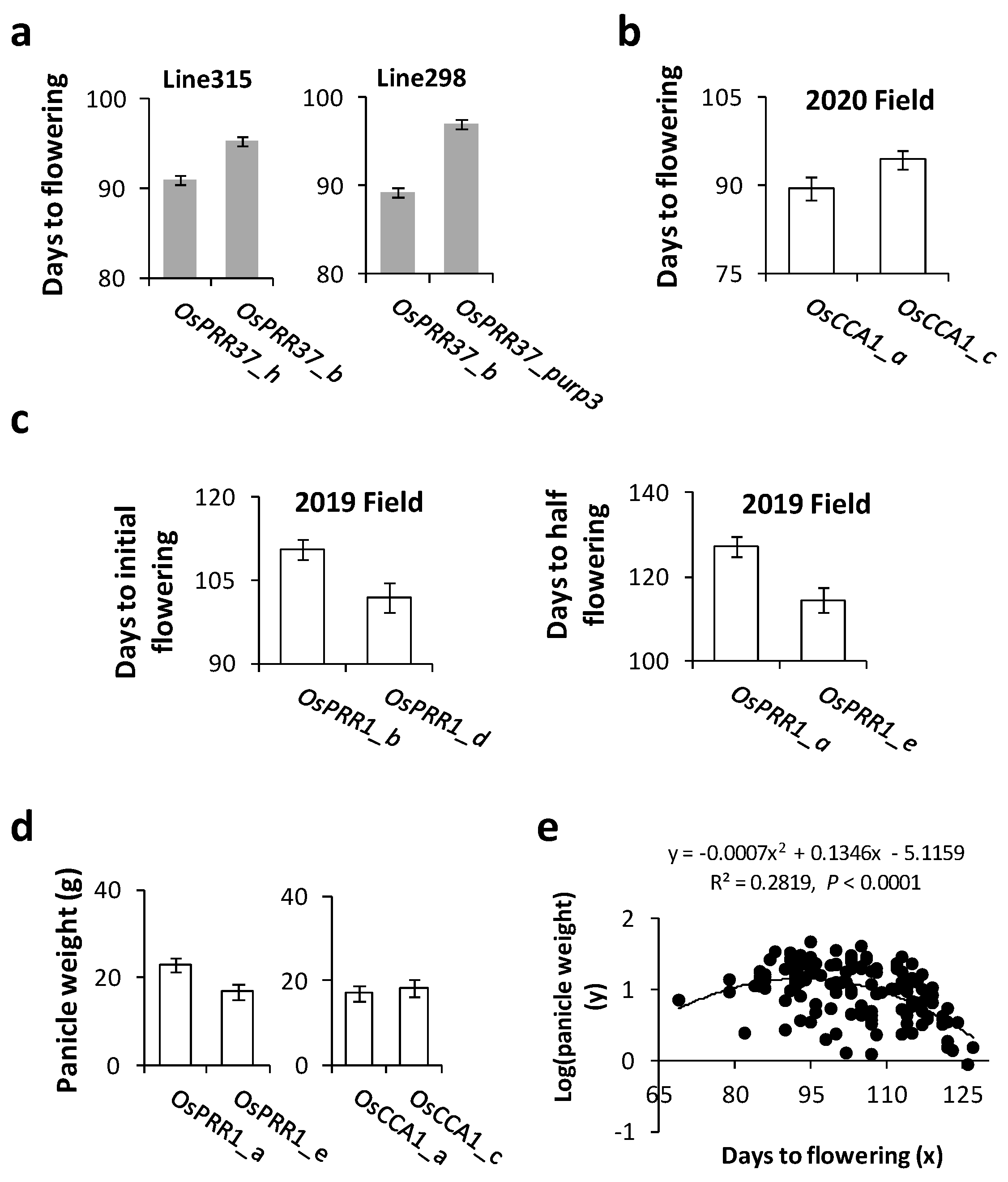
3.4. Associations of Oscillator Alleles with Rice Heading Time and Yield in the Field
3.5. Divergent Patterns of Oscillator Genes between Oryza and Arabidopsis
3.6. Compatibilities of OsCCA1 and OsPRR1 to Their Orthologs in A. thaliana
3.6.1. Effects of OsCCA1 alleles in A. thaliana
3.6.2. Effects of OsPRR1_a in A. thaliana
3.7. Linking Allelic Diversity of Oscillator Loci to Geographic Expansion of Rice Cultivation
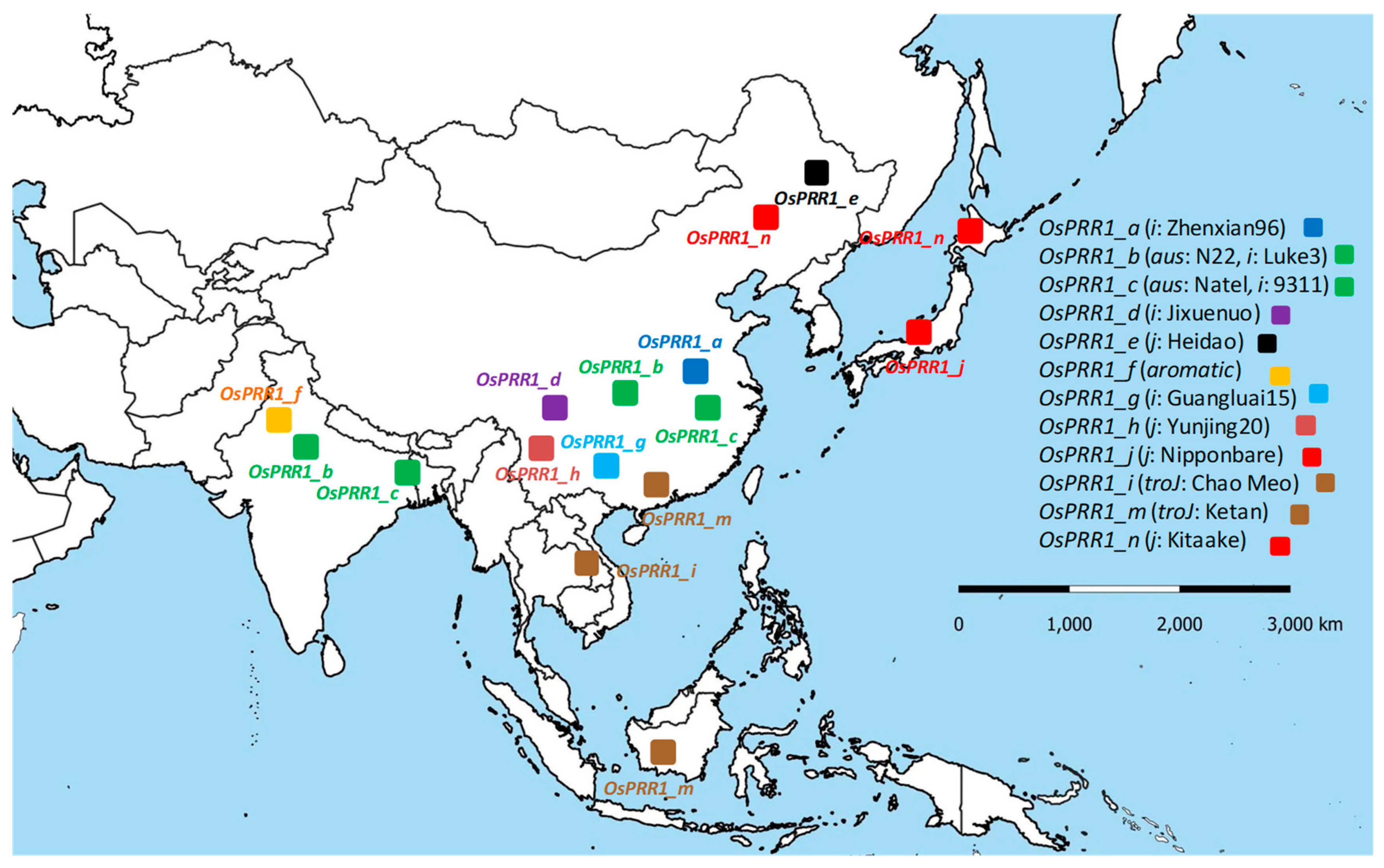
3.7.1. Geographic Distributions of Oscillator Alleles at OsPRR1 and OsPRR37 across Main Regions of Rice Cultivation
3.7.2. A Better Fitting of Allelic Distribution at OsPRR1 Than at OsPRR37 to Main Regions of Rice Cultivation
4. Discussion
4.1. Human Selection as a Major Force for Evolution of Oscillator in a Crop
4.2. Regulations of Circadian Clock in Quantity and Quality via Allelic Series
4.3. Birth of a New Allele Can Initiate at Either 5′ or the Coding Regions
4.4. Gene-Phenotype Associations
4.5. Implications for Future Breeding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alfoldi, J.; Wang, Q.B.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef]
- Gerstung, M.; Jolly, C.; Leshchiner, I.; Dentro, S.C.; Gonzalez, S.; Rosebrock, D.; Mitchell, T.J.; Rubanova, Y.; Anur, P.; Yu, K.X.; et al. The evolutionary history of 2658 cancers. Nature 2020, 578, 122–128. [Google Scholar] [CrossRef]
- Keane, T.M.; Goodstadt, L.; Danecek, P.; White, M.A.; Wong, K.; Yalcin, B.; Heger, A.; Agam, A.; Slater, G.; Goodson, M.; et al. Mouse genomic variation and its effect on phenotypes and gene regulation. Nature 2011, 477, 289–294. [Google Scholar] [CrossRef]
- Simon, M.M.; Greenaway, S.; White, J.K.; Fuchs, H.; Gailus-Durner, V.; Wells, S.; Sorg, T.; Wong, K.; Bedu, E.; Cartwright, E.J.; et al. A comparative phenotypic and genomic analysis of C57BL/6J and C57BL/6N mouse strains. Genome Biol. 2013, 14, R82. [Google Scholar] [CrossRef]
- Bogue, M.A.; Ball, R.L.; Philip, V.M.; Walton, D.O.; Dunn, M.H.; Kolishovski, G.; Lamoureux, A.; Gerring, M.; Liang, H.P.; Emerson, J.; et al. Mouse phenome database: Towards a more FAIR-compliant and TRUST-worthy data repository and tool suite for phenotypes and genotypes. Nucleic Acids Res. 2022, 51, D1067–D1074. [Google Scholar] [CrossRef]
- Andersen, E.C.; Gerke, J.P.; Shapiro, J.A.; Crissman, J.R.; Ghosh, R.; Bloom, J.S.; Felix, M.A.; Kruglyak, L. Chromosome-scale selective sweeps shape Caenorhabditis elegans genomic diversity. Nat. Genet. 2012, 44, 285–290. [Google Scholar] [CrossRef]
- Cook, D.E.; Zdraljevic, S.; Tanny, R.E.; Seo, B.; Riccardi, D.D.; Noble, L.M.; Rockman, M.V.; Alkema, M.J.; Braendle, C.; Kammenga, J.E.; et al. The genetic basis of natural variation in Caenorhabditis elegans telomere length. Genetics 2016, 204, 371–383. [Google Scholar] [CrossRef]
- Bank, C.; Hietpas, R.T.; Jensen, J.D.; Bolon, D.N.A. A systematic survey of an intragenic epistatic landscape. Mol. Biol. Evol. 2015, 32, 229–238. [Google Scholar] [CrossRef]
- Krach, E.K.; Skaro, M.; Wu, Y.; Arnold, J. Characterizing the gene-environment interaction underlying natural morphological variation in Neurospora crassa conidiophores using high-throughput phenomics and transcriptomics. G3-Genes Genomes Genet. 2022, 12, 14. [Google Scholar] [CrossRef]
- Alonso-Blanco, C.; Andrade, J.; Becker, C.; Bemm, F.; Bergelson, J.; Borgwardt, K.M.; Cao, J.; Chae, E.; Dezwaan, T.M.; Ding, W.; et al. 1,135 genomes reveal the global pattern of polymorphism in Arabidopsis thaliana. Cell 2016, 166, 481–491. [Google Scholar] [CrossRef]
- Kawakatsu, T.; Huang, S.S.C.; Jupe, F.; Sasaki, E.; Schmitz, R.J.; Urich, M.A.; Castanon, R.; Nery, J.R.; Barragan, C.; He, Y.P.; et al. Epigenomic diversity in a global collection of Arabidopsis thaliana accessions. Cell 2016, 166, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Rees, H.; Joynson, R.; Brown, J.K.M.; Hall, A. Naturally occurring circadian rhythm variation associated with clock gene loci in Swedish Arabidopsis accessions. Plant Cell Environ. 2021, 44, 807–820. [Google Scholar] [CrossRef] [PubMed]
- Migicovsky, Z.; Gardner, K.M.; Money, D.; Sawler, J.; Bloom, J.S.; Moffett, P.; Chao, C.T.; Schwaninger, H.; Fazio, G.; Zhong, G.Y.; et al. Genome to phenome mapping in apple using historical data. Plant Genome 2016, 9, 1–15. [Google Scholar] [CrossRef]
- Lu, Y. Functional significance of genetic polymorphisms. Front. Biol. China 2009, 4, 266–270. [Google Scholar] [CrossRef]
- Wright, S. Evolution and Genetics of Population; University of Chicago: Chicago, IL, USA, 1968; Volume 1, pp. 21–22. [Google Scholar]
- Slatkin, M.; Rannala, B. Estimating allele age. Annu. Rev. Genom. Hum. Genet. 2000, 1, 225–249. [Google Scholar] [CrossRef]
- Bradley, R.D.; Bull, J.J.; Johnson, A.D.; Hillis, D.M. Origin of a novel allele in a mammalian hybrid zone. Proc. Natl. Acad. Sci. USA 1993, 90, 8939–8941. [Google Scholar] [CrossRef]
- Bhave, M.; Morris, C.F. Molecular genetics of puroindolines and related genes: Allelic diversity in wheat and other grasses. Plant Mol.Biol. 2008, 66, 205–219. [Google Scholar] [CrossRef]
- Pin, P.A.; Zhang, W.Y.; Vogt, S.H.; Dally, N.; Buttner, B.; Schulze-Buxloh, G.; Jelly, N.S.; Chia, T.Y.P.; Mutasa-Gottgens, E.S.; Dohm, J.C.; et al. The role of a pseudo-response regulator gene in life cycle adaptation and domestication of beet. Curr. Biol. 2012, 22, 1095–1101. [Google Scholar] [CrossRef]
- Lu, Y.Q.; Xu, Y.Z.; Li, N. Early domestication history of asian rice revealed by mutations and genome-wide analysis of gene genealogies. Rice 2022, 15, 11. [Google Scholar] [CrossRef]
- Lee, W.; Tillo, D.; Bray, N.; Morse, R.H.; Davis, R.W.; Hughes, T.R.; Nislow, C. A high-resolution atlas of nucleosome occupancy in yeast. Nat. Genet. 2007, 39, 1235–1244. [Google Scholar] [CrossRef]
- Thurman, R.E.; Rynes, E.; Humbert, R.; Vierstra, J.; Maurano, M.T.; Haugen, E.; Sheffield, N.C.; Stergachis, A.B.; Wang, H.; Vernot, B.; et al. The accessible chromatin landscape of the human genome. Nature 2012, 489, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Pelechano, V.; Wei, W.; Steinmetz, L.M. Extensive transcriptional heterogeneity revealed by isoform profiling. Nature 2013, 497, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Matsushika, A.; Ashikari, M.; Yamashino, T.; Mizuno, T. Circadian-associated rice pseudo response regulators (OsPRRs): Insight into the control of flowering time. Biosci. Biotechnol. Biochem. 2005, 69, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Tago, Y.; Yamashino, T.; Mizuno, T. Comparative overviews of clock-associated genes of Arabidopsis thaliana and Oryza sativa. Plant Cell Physiol. 2007, 48, 110–121. [Google Scholar] [CrossRef]
- Murakami, M.; Tago, Y.; Yamashino, T.; Mizuno, T. Characterization of the rice circadian clock-associated pseudo-response regulators in Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2007, 71, 1107–1110. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, H.Y.; Qi, F.X.; Zhang, Z.Y.; Li, Q.P.; Han, Z.M.; Xing, Y.Z. Genetic interactions among Ghd7, Ghd8, OsPRR37 and Hd1 contribute to large variation in heading date in rice. Rice 2019, 12, 48. [Google Scholar] [CrossRef]
- Yano, M.; Katayose, Y.; Ashikari, M.; Yamanouchi, U.; Monna, L.; Fuse, T.; Baba, T.; Yamamoto, K.; Umehara, Y.; Nagamura, Y.; et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell 2000, 12, 2473–2483. [Google Scholar] [CrossRef]
- Xue, W.Y.; Xing, Y.Z.; Weng, X.Y.; Zhao, Y.; Tang, W.J.; Wang, L.; Zhou, H.J.; Yu, S.B.; Xu, C.G.; Li, X.H.; et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef]
- Wei, X.J.; Xu, J.F.; Guo, H.N.; Jiang, L.; Chen, S.H.; Yu, C.Y.; Zhou, Z.L.; Hu, P.S.; Zhai, H.Q.; Wan, J.M. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol. 2010, 153, 1747–1758. [Google Scholar] [CrossRef]
- Qiu, L.L.; Wu, Q.Q.; Wang, X.Y.; Han, J.P.; Zhuang, G.; Wang, H.; Shang, Z.Y.; Tian, W.; Chen, Z.; Lin, Z.C.; et al. Forecasting rice latitude adaptation through a daylength-sensing-based environment adaptation simulator. Nat. Food 2021, 2, 348–362. [Google Scholar] [CrossRef]
- Wang, Y.P.; Wu, F.Q.; Zhou, S.R.; Chen, W.W.; Li, C.Y.; Duan, E.R.; Wang, J.C.; Cheng, Z.J.; Zhang, X.; Lin, Q.B.; et al. Clock component OsPRR59 delays heading date by repressing transcription of Ehd3 in rice. Crop J. 2022, 10, 1570–1579. [Google Scholar] [CrossRef]
- Matsubara, K.; Yamanouchi, U.; Nonoue, Y.; Sugimoto, K.; Wang, Z.X.; Minobe, Y.; Yano, M. Ehd3, encoding a plant homeodomain finger-containing protein, is a critical promoter of rice flowering. Plant J. 2011, 66, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Q.; Tian, X.Y.; Li, J.; Bai, S.; Zhang, Z.Y.; Li, Y.; Cao, H.R.; Chen, Z.C. Two central circadian oscillators OsPRR59 and OsPRR95 modulate magnesium homeostasis and carbon fixation in rice. Mol. Plant. 2022, 15, 1602–1614. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.X.; Knowles, S.M.; Andronis, C.; Ong, M.S.; Tobin, E.M. CIRCADIAN CLOCK ASSOCIATED1 and LATE ELONGATED HYPOCOTYL function synergistically in the circadian clock of Arabidopsis. Plant Physiol. 2009, 150, 834–843. [Google Scholar] [CrossRef]
- Yakir, E.; Hilman, D.; Kron, I.; Hassidim, M.; Melamed-Book, N.; Green, R.M. Posttranslational Regulation of CIRCADIAN CLOCK ASSOCIATED1 in the circadian oscillator of Arabidopsis. Plant Physiol. 2009, 150, 844–857. [Google Scholar] [CrossRef]
- Lee, S.J.; Kang, K.; Lim, J.H.; Paek, N.C. Natural alleles of CIRCADIAN CLOCK ASSOCIATED1 contribute to rice cultivation by fine-tuning flowering time. Plant Physiol. 2022, 190, 640–656. [Google Scholar] [CrossRef]
- Wang, F.; Han, T.W.; Song, Q.X.; Ye, W.X.; Song, X.G.; Chu, J.F.; Li, J.Y.; Chen, Z.J. The rice circadian clock regulates tiller growth and panicle development through strigolactone signaling and sugar sensing. Plant Cell 2020, 32, 3124–3138. [Google Scholar] [CrossRef]
- Wei, H.; Xu, H.; Su, C.; Wang, X.L.; Wang, L. Rice CIRCADIAN CLOCK ASSOCIATED 1 transcriptionally regulates ABA signaling to confer multiple abiotic stress tolerance. Plant Physiol. 2022, 190, 1057–1073. [Google Scholar] [CrossRef]
- Matsuzaki, J.; Kawahara, Y.; Izawa, T. Punctual transcriptional regulation by the rice circadian clock under fluctuating field conditions. Plant Cell 2015, 27, 633–648. [Google Scholar] [CrossRef]
- Maeda, A.E.; Nakamichi, N. Plant clock modifications for adapting flowering time to local environments. Plant Physiol. 2022, 190, 952–967. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Tobin, E.M. Constitutive expression of the CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) gene disrupts circadian rhythms and suppresses its own expression. Cell 1998, 93, 1207–1217. [Google Scholar] [CrossRef] [PubMed]
- Matsushika, A.; Makino, S.; Kojima, M.; Mizuno, T. Circadian waves of expression of the APRR1/TOC1 family of pseudo-response regulators in Arabidopsis thaliana: Insight into the plant circadian clock. Plant Cell Physiol. 2000, 41, 1002–1012. [Google Scholar] [CrossRef] [PubMed]
- Kaczorowski, K.A.; Quail, P.H. Arabidopsis PSEUDO-RESPONSE REGULATOR7 is a signaling intermediate in phytochrome-regulated seedling deetiolation and phasing of the circadian clock. Plant Cell 2003, 15, 2654–2665. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.Y.; Harmer, S.L. Circadian phase has profound effects on differential expression analysis. PLoS ONE 2012, 7, e49853. [Google Scholar] [CrossRef] [PubMed]
- Kiba, T.; Henriques, R.; Sakakibara, H.; Chua, N.H. Targeted degradation of PSEUDO-RESPONSE REGULATOR5 by an SCFZTL complex regulates clock function and photomorphogenesis in Arabidopsis thaliana. Plant Cell 2007, 19, 2516–2530. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, M.; Li, G.; Yuan, L.; Xie, Y.; Wei, H.; Ma, X.; Li, Q.; Devlin, P.F.; Xu, X.; et al. Transcription factors FHY3 and FAR1 regulate light-induced CIRCADIAN CLOCK ASSOCIATED1 gene expression in Arabidopsis. Plant Cell 2020, 32, 1464–1478. [Google Scholar] [CrossRef]
- Kamioka, M.; Takao, S.; Suzuki, T.; Taki, K.; Higashiyama, T.; Kinoshita, T.; Nakamichi, N. Direct repression of evening genes by CIRCADIAN CLOCK-ASSOCIATED1 in the Arabidopsis circadian clock. Plant Cell 2016, 28, 696–711. [Google Scholar] [CrossRef]
- Nakamichi, N.; Kiba, T.; Henriques, R.; Mizuno, T.; Chua, N.H.; Sakakibara, H. PSEUDO-RESPONSE REGULATORS 9, 7, and 5 are transcriptional repressors in the Arabidopsis circadian clock. Plant Cell 2010, 22, 594–605. [Google Scholar] [CrossRef]
- Sanchez, S.E.; Rugnone, M.L.; Kay, S.A. Light perception: A matter of time. Mol. Plant. 2020, 13, 363–385. [Google Scholar] [CrossRef]
- Somers, D.E.; Webb, A.A.R.; Pearson, M.; Kay, S.A. The short-period mutant, toc1-1, alters circadian clock regulation of multiple outputs throughout development in Arabidopsis thaliana. Development 1998, 125, 485–494. [Google Scholar] [CrossRef]
- Strayer, C.; Oyama, T.; Schultz, T.F.; Raman, R.; Somers, D.E.; Mas, P.; Panda, S.; Kreps, J.A.; Kay, S.A. Cloning of the Arabidopsis clock gene TOC1, an autoregulatory response regulator homolog. Science 2000, 289, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Mas, P.; Alabadi, D.; Yanovsky, M.J.; Oyama, T.; Kay, S.A. Dual role of TOC1 in the control of circadian and photomorphogenic responses in Arabidopsis. Plant Cell 2003, 15, 223–236. [Google Scholar] [CrossRef]
- Hayama, R.; Yokoi, S.; Tamaki, S.; Yano, M.; Shimamoto, K. Adaptation of photoperiodic control pathways produces short-day flowering in rice. Nature 2003, 422, 719–722. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.; Rovira, A.; Veciana, N.; Soy, J.; Toledo-Ortiz, G.; Gommers, C.M.M.; Boix, M.; Henriques, R.; Minguet, E.G.; Alabadi, D.; et al. Circadian waves of transcriptional repression shape PIF-regulated photoperiod-responsive growth in Arabidopsis. Curr. Biol. 2018, 28, 311–318. [Google Scholar] [CrossRef] [PubMed]
- McClung, C.R. The plant circadian oscillator. Biology 2019, 8, 14. [Google Scholar] [CrossRef]
- Kim, H.; Kim, H.J.; Vu, Q.T.; Jung, S.; McClung, C.R.; Hong, S.; Nam, H.G. Circadian control of ORE1 by PRR9 positively regulates leaf senescence in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, 8448–8453. [Google Scholar] [CrossRef]
- Zhang, Y.; Pfeiffer, A.; Tepperman, J.M.; Dalton-Roesler, J.; Leivar, P.; Grandio, E.G.; Quail, P.H. Central clock components modulate plant shade avoidance by directly repressing transcriptional activation activity of PIF proteins. Proc. Natl. Acad. Sci. USA 2020, 117, 3261–3269. [Google Scholar] [CrossRef]
- Zhou, Y.; Chebotarov, D.; Kudrna, D.; Llaca, V.; Lee, S.; Rajasekar, S.; Mohammed, N.; Al-Bader, N.; Sobel-Sorenson, C.; Parakkal, P.; et al. A platinum standard pan-genome resource that represents the population structure of Asian rice. Sci. Data 2020, 7, 113. [Google Scholar] [CrossRef]
- Lu, Y.Q. Gene genealogy-based mutation analysis reveals emergence of aus, tropical japonica, and aromatic of Oryza sativa during the later stage of rice domestication. Genes 2023, 14, 1412. [Google Scholar] [CrossRef]
- Yang, Z.H.; Nielsen, R.; Goldman, N.; Pedersen, A.M.K. Codon-substitution models for heterogeneous selection pressure at amino acid sites. Genetics 2000, 155, 431–449. [Google Scholar] [CrossRef]
- Nielsen, R. Molecular signatures of natural selection. Annu. Rev. Genet. 2005, 39, 197–218. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Bramsiepe, J.; Krabberod, A.K.; Bjerkan, K.N.; Alling, R.M.; Johannessen, I.M.; Hornslien, K.S.; Miller, J.R.; Brysting, A.K.; Grini, P.E. Structural evidence for MADS-box type I family expansion seen in new assemblies of Arabidopsis arenosa and A. lyrata. Plant J. 2023, 116, 942–961. [Google Scholar] [CrossRef]
- Lu, Y.Q.; Xie, L.L.; Chen, J.N. A novel procedure for absolute real-time quantification of gene expression patterns. Plant Methods 2012, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.L.; Guan, S.; Zhu, Z.X.; Wang, Y.; Lu, Y.Q. A valid strategy for precise identifications of transcription factor binding sites in combinatorial regulation using bioinformatic and experimental approaches. Plant Methods 2013, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Neff, M.M.; Chory, J. Genetic interactions between phytochrome A, phytochrome B, and cryptochrome 1 during Arabidopsis development. Plant Physiol. 1998, 118, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Lange, H.; Mohr, H. Analysis of phytochrome-mediated anthocyanin synthesis. Plant Physiol. 1971, 47, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Hollander, M.; Wolfe, D. Nonparametric Statistical Methods, 2nd ed.; Wiley & Sons: New York, NY, USA, 1999. [Google Scholar]
- Koo, B.H.; Yoo, S.C.; Park, J.W.; Kwon, C.T.; Lee, B.D.; An, G.; Zhang, Z.Y.; Li, J.J.; Li, Z.C.; Paek, N.C. Natural variation in OsPRR37 regulates heading date and contributes to rice cultivation at a wide range of latitudes. Mol. Plant. 2013, 6, 1877–1888. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.-M.; Feng, L.; Wang, J.; Qiao, W.; Zhang, L.; Cheng, Y.; Yang, Q. Nonfunctional alleles of long-day suppressor genes independently regulate flowering time. J. Integr. Plant Biol. 2016, 58, 540–548. [Google Scholar] [CrossRef]
- Ury, H.K. Comparison of 4 procedures for multiple comparisons among means (pairwise contrasts) for arbitrary sample sizes. Technometrics 1976, 18, 89–97. [Google Scholar] [CrossRef]
- Nagel, D.H.; Doherty, C.J.; Pruneda-Paz, J.L.; Schmitz, R.J.; Ecker, J.R.; Kay, S.A. Genome-wide identification of CCA1 targets uncovers an expanded clock network in Arabidopsis. Proc. Natl. Acad. Sci. USA 2015, 112, E4802–E4810. [Google Scholar] [CrossRef]
- Alabadi, D.; Oyama, T.; Yanovsky, M.J.; Harmon, F.G.; Mas, P.; Kay, S.A. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 2001, 293, 880–883. [Google Scholar] [CrossRef] [PubMed]
- Anwer, M.U.; Davis, S.J. An overview of natural variation studies in the Arabidopsis thaliana circadian clock. Semin. Cell Develop. Biol. 2013, 24, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; Hogenesch, J.B. A circadian gene expression atlas in mammals: Implications for biology and medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [PubMed]
- Covington, M.F.; Maloof, J.N.; Straume, M.; Kay, S.A.; Harmer, S.L. Global transcriptome analysis reveals circadian regulation of key pathways in plant growth and development. Genome Biol. 2008, 9, R130. [Google Scholar] [CrossRef]
- Strutt, S.; Sellinger, T.; Glemin, S.; Tellier, A.; Laurent, S. Joint inference of evolutionary transitions to self-fertilization and demographic history using whole-genome sequences. eLife 2023, 12, e82384. [Google Scholar] [CrossRef]
- Gross, B.L.; Zhao, Z.J. Archaeological and genetic insights into the origins of domesticated rice. Proc. Natl. Acad. Sci. USA 2014, 111, 6190–6197. [Google Scholar] [CrossRef]
- Liu, C.; Qu, X.F.; Zhou, Y.H.; Song, G.Y.; Abiri, N.; Xiao, Y.H.; Liang, F.; Jiang, D.M.; Hu, Z.L.; Yang, D.C. OsPRR37 confers an expanded regulation of the diurnal rhythms of the transcriptome and photoperiodic flowering pathways in rice. Plant Cell Environ. 2018, 41, 630–645. [Google Scholar] [CrossRef]
- Liang, L.W.; Zhang, Z.Y.; Cheng, N.N.; Liu, H.Y.; Song, S.; Hu, Y.; Zhou, X.C.; Zhang, J.; Xing, Y.Z. The transcriptional repressor OsPRR73 links circadian clock and photoperiod pathway to control heading date in rice. Plant Cell Environ. 2021, 44, 842–855. [Google Scholar] [CrossRef]
- Sun, C.H.; Zhang, K.; Zhou, Y.; Xiang, L.; He, C.C.; Zhong, C.; Li, K.; Wang, Q.X.; Yang, C.P.; Wang, Q.; et al. Dual function of clock component OsLHY sets critical day length for photoperiodic flowering in rice. Plant Biotechnol. J. 2021, 19, 1644–1657. [Google Scholar] [CrossRef]
- Morris, W.L.; Ducreux, L.J.M.; Morris, J.; Campbell, R.; Usman, M.; Hedley, P.E.; Prat, S.; Taylor, M.A. Identification of TIMING OF CAB EXPRESSION 1 as a temperature-sensitive negative regulator of tuberization in potato. J. Exp. Bot. 2019, 70, 5703–5714. [Google Scholar] [CrossRef] [PubMed]
- Turck, F.; Fornara, F.; Coupland, G. Regulation and identity of florigen: FLOWERING LOCUS T moves center stage. Annu. Rev. Plant Biol. 2008, 59, 573–594. [Google Scholar] [CrossRef] [PubMed]
- Sawa, M.; Kay, S.A. GIGANTEA directly activates Flowering Locus T in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2011, 108, 11698–11703. [Google Scholar] [CrossRef] [PubMed]
- Hayama, R.; Sarid-Krebs, L.; Richter, R.; Fernandez, V.; Jang, S.; Coupland, G. PSEUDO RESPONSE REGULATORs stabilize CONSTANS protein to promote flowering in response to day length. EMBO J. 2017, 36, 904–918. [Google Scholar] [CrossRef]
- Kojima, S.; Takahashi, Y.; Kobayashi, Y.; Monna, L.; Sasaki, T.; Araki, T.; Yano, M. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 2002, 43, 1096–1105. [Google Scholar] [CrossRef]
- Doi, K.; Izawa, T.; Fuse, T.; Yamanouchi, U.; Kubo, T.; Shimatani, Z.; Yano, M.; Yoshimura, A. Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-Iike gene expression independently of Hd1. Genes Dev. 2004, 18, 926–936. [Google Scholar] [CrossRef]
- Kim, S.L.; Choi, M.; Jung, K.H.; An, G. Analysis of the early-flowering mechanisms and generation of T-DNA tagging lines in Kitaake, a model rice cultivar. J. Exp. Bot. 2013, 64, 4169–4182. [Google Scholar] [CrossRef]
 ) were estimated from neighboring polymorphic sites, with colors showing origins of parental sequences. Regions of uncertain origin are shown in horizontally dashed lines. Os-specific mutations are shown by a circle for indel (colors for different ones) or letter for substitution (red for the nonsynonymous and black for the synon-ymous). The arrow ends at the stop codon of each allele, with the allele designated in lower letter following the locus. Drawings are not proportional to the genic regions but show relative positions of features. (b) Eight alleles of OsPRR95. Two tentative allelets (OsPRR95_b′ and _d′) are included here. Different indels are shown in colors. (c) Nine alleles of OsPRR37. (d) Six alleles of OsPRR59. An early stop codon caused by mutation is shown by x. (e) Seven alleles of OsPRR1. (f) Relative con-tributions of parental genomes to the oscillator loci in Asian rice. The components of the molecular clock are in square with parental origins of their sequences in colors. Arrows indicate flow of time, with dark ones for the night. (g) Allelic mutation rates across genic regions of OsPRR1. Seven alleles shown in e are included. The baseline (3′-875) is from 875-bp genomic region downstream of the 3′ region. Letters a and b indicate a significant t-test after correction for multiple comparisons at ex-perimental error rate of 0.05. See Table 2 for detail. (h) Allelic mutation rates across genic regions of OsPRR37. Nine alleles (shown in (c)) are included. The baseline (3′-1035) is from 1035-bp genomic region downstream of the 3′ region. Format of tests follows that of (g).
) were estimated from neighboring polymorphic sites, with colors showing origins of parental sequences. Regions of uncertain origin are shown in horizontally dashed lines. Os-specific mutations are shown by a circle for indel (colors for different ones) or letter for substitution (red for the nonsynonymous and black for the synon-ymous). The arrow ends at the stop codon of each allele, with the allele designated in lower letter following the locus. Drawings are not proportional to the genic regions but show relative positions of features. (b) Eight alleles of OsPRR95. Two tentative allelets (OsPRR95_b′ and _d′) are included here. Different indels are shown in colors. (c) Nine alleles of OsPRR37. (d) Six alleles of OsPRR59. An early stop codon caused by mutation is shown by x. (e) Seven alleles of OsPRR1. (f) Relative con-tributions of parental genomes to the oscillator loci in Asian rice. The components of the molecular clock are in square with parental origins of their sequences in colors. Arrows indicate flow of time, with dark ones for the night. (g) Allelic mutation rates across genic regions of OsPRR1. Seven alleles shown in e are included. The baseline (3′-875) is from 875-bp genomic region downstream of the 3′ region. Letters a and b indicate a significant t-test after correction for multiple comparisons at ex-perimental error rate of 0.05. See Table 2 for detail. (h) Allelic mutation rates across genic regions of OsPRR37. Nine alleles (shown in (c)) are included. The baseline (3′-1035) is from 1035-bp genomic region downstream of the 3′ region. Format of tests follows that of (g).
) were estimated from neighboring polymorphic sites, with colors showing origins of parental sequences. Regions of uncertain origin are shown in horizontally dashed lines. Os-specific mutations are shown by a circle for indel (colors for different ones) or letter for substitution (red for the nonsynonymous and black for the synon-ymous). The arrow ends at the stop codon of each allele, with the allele designated in lower letter following the locus. Drawings are not proportional to the genic regions but show relative positions of features. (b) Eight alleles of OsPRR95. Two tentative allelets (OsPRR95_b′ and _d′) are included here. Different indels are shown in colors. (c) Nine alleles of OsPRR37. (d) Six alleles of OsPRR59. An early stop codon caused by mutation is shown by x. (e) Seven alleles of OsPRR1. (f) Relative con-tributions of parental genomes to the oscillator loci in Asian rice. The components of the molecular clock are in square with parental origins of their sequences in colors. Arrows indicate flow of time, with dark ones for the night. (g) Allelic mutation rates across genic regions of OsPRR1. Seven alleles shown in e are included. The baseline (3′-875) is from 875-bp genomic region downstream of the 3′ region. Letters a and b indicate a significant t-test after correction for multiple comparisons at ex-perimental error rate of 0.05. See Table 2 for detail. (h) Allelic mutation rates across genic regions of OsPRR37. Nine alleles (shown in (c)) are included. The baseline (3′-1035) is from 1035-bp genomic region downstream of the 3′ region. Format of tests follows that of (g).
) were estimated from neighboring polymorphic sites, with colors showing origins of parental sequences. Regions of uncertain origin are shown in horizontally dashed lines. Os-specific mutations are shown by a circle for indel (colors for different ones) or letter for substitution (red for the nonsynonymous and black for the synon-ymous). The arrow ends at the stop codon of each allele, with the allele designated in lower letter following the locus. Drawings are not proportional to the genic regions but show relative positions of features. (b) Eight alleles of OsPRR95. Two tentative allelets (OsPRR95_b′ and _d′) are included here. Different indels are shown in colors. (c) Nine alleles of OsPRR37. (d) Six alleles of OsPRR59. An early stop codon caused by mutation is shown by x. (e) Seven alleles of OsPRR1. (f) Relative con-tributions of parental genomes to the oscillator loci in Asian rice. The components of the molecular clock are in square with parental origins of their sequences in colors. Arrows indicate flow of time, with dark ones for the night. (g) Allelic mutation rates across genic regions of OsPRR1. Seven alleles shown in e are included. The baseline (3′-875) is from 875-bp genomic region downstream of the 3′ region. Letters a and b indicate a significant t-test after correction for multiple comparisons at ex-perimental error rate of 0.05. See Table 2 for detail. (h) Allelic mutation rates across genic regions of OsPRR37. Nine alleles (shown in (c)) are included. The baseline (3′-1035) is from 1035-bp genomic region downstream of the 3′ region. Format of tests follows that of (g).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Number of Alleles | 5′ Region | Coding Region(s) | ||||
|---|---|---|---|---|---|---|---|
| Length * (nt) | Mutations | Mutation Density | Length * (nt) | Mutations | Mutation Density | ||
| OsCCA1 | 6 | 1014 | 1 | 0.0010 | 2160 | 7 | 0.0032 |
| OsPRR95 | 8 | 1031 | 16 | 0.0155 | 1872 | 4 | 0.0021 |
| OsPRR37 | 9 | 1036 | 4 | 0.0039 | 2230 | 15 | 0.0067 |
| OsPRR59 | 6 | 1015 | 15 | 0.0148 | 2088 | 9 | 0.0043 |
| OsPRR1 | 7 | 1305 | 28 | 0.0215 | 1557 | 8 | 0.0051 |
| Mutation Rate Per Allele | Probability for One Tailed t-Test | |||||
|---|---|---|---|---|---|---|
| Locus | 5′ | Coding | Intergenic a | 5′ vs. Int. b | Coding vs. Int. | 5′ vs. Coding |
| CCA1/LHY | 0.00026 (0.00016) | 0.00046 (0.00023) | 0.00109 (0.00045) | 0.036 | 0.059 | 0.086 |
| PRR95 | 0.00194 (0.00066) | 0.00027 (0.00013) | 0.00108 (0.00035) | 0.141 | 0.087 | 0.034 |
| PRR37 | 0.00043 (0.00013) | 0.00075 (0.00022) | 0.00216 (0.00027) | <0.001 c | 0.163 | 0.018 |
| PRR59 | 0.00246 (0.00058) | 0.00074 (0.00030) | 0.00211 (0.00054) | 0.332 | 0.224 | 0.131 |
| PRR1 | 0.00307 (0.00029) | 0.00077 (0.00020) | 0.00230 (0.00067) | 0.009 c | 0.115 | <0.0001 c |
| Locus | Number of Alleles | 5′ Region | Coding Region(s) | ||||
|---|---|---|---|---|---|---|---|
| Length a (nt) | Mutations b | Mutation Density c | Length a (nt) | Mutations b | Mutation Density | ||
| CCA1 | 8 | 1002 | 11 | 0.0130 | 1827 | 4 | 0.0022 |
| PRR9 | 7 | 1012 | 5 | 0.0049 | 1407 | 8 | 0.0057 |
| PRR7 | 9 | 1018 | 10 | 0.0098 | 2184 | 13 | 0.0060 |
| PRR5 | 8 | 1011 | 10 | 0.0099 | 2010 | 13 | 0.0065 d |
| PRR1 | 10 | 1020 | 12 | 0.0118 | 1857 | 4 | 0.0022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, G.; Chen, M.; Mo, R.; Li, N.; Xu, Y.; Lu, Y. Linking New Alleles at the Oscillator Loci to Flowering and Expansion of Asian Rice. Genes 2023, 14, 2027. https://doi.org/10.3390/genes14112027
Gao G, Chen M, Mo R, Li N, Xu Y, Lu Y. Linking New Alleles at the Oscillator Loci to Flowering and Expansion of Asian Rice. Genes. 2023; 14(11):2027. https://doi.org/10.3390/genes14112027
Chicago/Turabian StyleGao, Guangtong, Maoxian Chen, Rong Mo, Nan Li, Yunzhang Xu, and Yingqing Lu. 2023. "Linking New Alleles at the Oscillator Loci to Flowering and Expansion of Asian Rice" Genes 14, no. 11: 2027. https://doi.org/10.3390/genes14112027
APA StyleGao, G., Chen, M., Mo, R., Li, N., Xu, Y., & Lu, Y. (2023). Linking New Alleles at the Oscillator Loci to Flowering and Expansion of Asian Rice. Genes, 14(11), 2027. https://doi.org/10.3390/genes14112027