
KIBRA Gene Variant Is Associated with Ability in Chess and Science

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Participants
2.3. Genetic Analysis
2.3.1. Russian and Tatar Samples
2.3.2. Kazakh Samples
2.4. Statistical Analyses
3. Results
3.1. Association of the KIBRA Gene Variant with Chess Player Status
3.2. Association of the KIBRA Gene Variant with PhD Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schneider, A.; Huentelman, M.J.; Kremerskothen, J.; Duning, K.; Spoelgen, R.; Nikolich, K. KIBRA: A new gateway to learning and memory? Front. Aging Neurosci. 2010, 2, 4. [Google Scholar] [CrossRef]
- Johannsen, S.; Duning, K.; Pavenstädt, H.; Kremerskothen, J.; Boeckers, T. Temporal-spatial expression and novel biochemical properties of the memory-related protein KIBRA. Neuroscience 2008, 155, 1165–1173. [Google Scholar] [CrossRef]
- Papassotiropoulos, A.; Stephan, D.A.; Huentelman, M.J.; Hoerndli, F.J.; Craig, D.W.; Pearson, J.V.; Huynh, K.-D.; Brunner, F.; Corneveaux, J.; Osborne, D.; et al. Common Kibra Alleles Are Associated with Human Memory Performance. Science 2006, 314, 475–478. [Google Scholar] [CrossRef]
- Milnik, A.; Heck, A.; Vogler, C.; Heinze, H.-J.; de Quervain, D.J.-F.; Papassotiropoulos, A. Association of KIBRA with episodic and working memory: A meta-analysis. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2012, 159B, 958–969. [Google Scholar] [CrossRef]
- Muse, J.; Emery, M.; Sambataro, F.; Lemaitre, H.; Tan, H.-Y.; Chen, Q.; Kolachana, B.S.; Das, S.; Callicott, J.; Weinberger, D.R.; et al. WWC1 Genotype Modulates Age-Related Decline in Episodic Memory Function Across the Adult Life Span. Biol. Psychiatry 2013, 75, 693–700. [Google Scholar] [CrossRef]
- Zlomuzica, A.; Preusser, F.; Roberts, S.; Woud, M.L.; Lester, K.J.; Dere, E.; Eley, T.C.; Margraf, J. The role of KIBRA in reconstructive episodic memory. Mol. Med. 2018, 24, 7. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Wedow, R.; Okbay, A.; Kong, E.; Maghzian, O.; Zacher, M.; Nguyen-Viet, T.A.; Bowers, P.; Sidorenko, J.; Karlsson Linnér, R.; et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat. Genet. 2018, 50, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Kazantseva, A.V.; Enikeeva, R.F.; Davydova, Y.D.; Mustafin, R.N.; Takhirova, Z.R.; Malykh, S.B.; Lobaskova, M.M.; Tikhomirova, T.N.; Khusnutdinova, E.K. The role of the KIBRA and APOE genes in developing spatial abilities in humans. Vavilov J. Genet. Breed. 2021, 25, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Schuck, N.W.; Doeller, C.F.; Schjeide, B.M.; Schröder, J.; Frensch, P.A.; Bertram, L.; Li, S.C. Aging and KIBRA/WWC1 genotype affect spatial memory processes in a virtual navigation task. Hippocampus 2013, 23, 919–930. [Google Scholar] [CrossRef]
- Rovira, E.; Mackie, R.S.; Clark, N.; Squire, P.N.; Hendricks, M.D.; Pulido, A.M.; Greenwood, P.M. A role for attention during wilderness navigation: Comparing effects of BDNF, KIBRA, and CHRNA4. Neuropsychology 2016, 30, 709–719. [Google Scholar] [CrossRef]
- Malanchini, M.; Rimfeld, K.; Shakeshaft, N.G.; McMillan, A.; Schofield, K.L.; Rodic, M.; Rossi, V.; Kovas, Y.; Dale, P.S.; Tucker-Drob, E.M.; et al. Evidence for a unitary structure of spatial cognition beyond general intelligence. NPJ Sci. Learn. 2020, 5, 9. [Google Scholar] [CrossRef]
- Kell, H.J.; Lubinski, D.; Benbow, C.P.; Steiger, J.H. Creativity and technical innovation: Spatial ability’s unique role. Psychol. Sci. 2013, 24, 1831–1836. [Google Scholar] [CrossRef]
- Rodic, M.; Tikhomirova, T.; Kolienko, T.; Malykh, S.; Bogdanova, O.; Zueva, D.Y.; Gynku, E.I.; Wan, S.; Zhou, X.; Kovas, Y. Spatial complexity of character-based writing systems and arithmetic in primary school: A longitudinal study. Front. Psychol. 2015, 6, 333. [Google Scholar] [CrossRef]
- Frydman, M.; Lynn, R. The general intelligence and spatial abilities of gifted young Belgian chess players. Br. J. Psychol. 1992, 83, 233–235. [Google Scholar] [CrossRef]
- Burgoyne, A.P.; Sala, G.; Gobet, F.; Macnamara, B.N.; Campitelli, G.; Hambrick, D.Z. The relationship between cognitive ability and chess skill: A comprehensive meta-analysis. Intelligence 2016, 59, 72–83. [Google Scholar] [CrossRef]
- Smith, E.T.; Bartlett, J.C.; Krawczyk, D.C.; Basak, C. Are the advantages of chess expertise on visuo-spatial working-memory capacity domain specific or domain general? Mem. Cognit. 2021, 49, 1600–1616. [Google Scholar] [CrossRef]
- Wilson, R.S.; Barral, S.; Lee, J.H.; Leurgans, S.E.; Foroud, T.M.; Sweet, R.A.; Graff-Radford, N.; Bird, T.D.; Mayeux, R.; Bennett, D.A. Heritability of different forms of memory in the Late Onset Alzheimer’s Disease Family Study. J. Alzheimers Dis. 2011, 23, 249–255. [Google Scholar] [CrossRef]
- Vogler, C.; Gschwind, L.; Coynel, D.; Freytag, V.; Milnik, A.; Egli, T.; Heck, A.; de Quervain, D.J.; Papassotiropoulos, A. Substantial SNP-based heritability estimates for working memory performance. Transl. Psychiatry 2014, 4, e438. [Google Scholar] [CrossRef]
- Ellis, D.M.; Ball, B.H.; Kimpton, N.; Brewer, G.A. The role of working memory capacity in analytic and multiply-constrained problem-solving in demanding situations. Q. J. Exp. Psychol. 2020, 73, 920–928. [Google Scholar] [CrossRef]
- Shelton, J.T.; Elliott, E.M.; Matthews, R.A.; Hill, B.D.; Gouvier, W.D. The relationships of working memory, secondary memory, and general fluid intelligence: Working memory is special. J. Exp. Psychol. Learn. Mem. Cogn. 2010, 36, 813–820. [Google Scholar] [CrossRef]
- Little, J.; Higgins, J.P.T.; Ioannidis, J.P.A.; Moher, D.; Gagnon, F.; von Elm, E.; Khoury, M.J.; Cohen, B.; Davey-Smith, G.; Grimshaw, J.; et al. Strengthening the reporting of genetic association studies (STREGA): An extension of the STROBE Statement. Hum. Genet. 2009, 125, 131–151. [Google Scholar] [CrossRef]
- Akhmetov, I.I.; Astranenkova, I.V.; Rogozkin, V.A. Association of PPARD gene polymorphism with human physical performance. Mol. Biol. 2007, 41, 852–857. [Google Scholar]
- Al-Khelaifi, F.; Yousri, N.A.; Diboun, I.; Semenova, E.A.; Kostryukova, E.S.; Kulemin, N.A.; Borisov, O.V.; Andryushchenko, L.B.; Larin, A.K.; Generozov, E.V.; et al. Genome-Wide Association Study Reveals a Novel Association Between MYBPC3 Gene Polymorphism, Endurance Athlete Status, Aerobic Capacity and Steroid Metabolism. Front. Genet. 2020, 11, 595. [Google Scholar] [CrossRef]
- Kikuchi, N.; Moreland, E.; Homma, H.; Semenova, E.A.; Saito, M.; Larin, A.K.; Kobatake, N.; Yusupov, R.A.; Okamoto, T.; Nakazato, K.; et al. Genes and Weightlifting Performance. Genes 2021, 13, 25. [Google Scholar] [CrossRef]
- Fesenko, D.O.; Ivanovsky, I.D.; Ivanov, P.L.; Zemskova, E.Y.; Agapitova, A.S.; Polyakov, S.A.; Fesenko, O.E.; Filippova, M.A.; Zasedatelev, A.S. A Biochip for Genotyping Polymorphisms Associated with Eye, Hair, Skin Color, AB0 Blood Group, Sex, Y Chromosome Core Haplogroup, and Its Application to Study the Slavic Population. Mol. Biol. 2022, 56, 860–880. [Google Scholar] [CrossRef]
- Eynon, N.; Nasibulina, E.S.; Banting, L.K.; Cieszczyk, P.; Maciejewska-Karlowska, A.; Sawczuk, M.; Bondareva, E.A.; Shagimardanova, R.R.; Raz, M.; Sharon, Y.; et al. The FTO A/T polymorphism and elite athletic performance: A study involving three groups of European athletes. PLoS One 2013, 8, e60570. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Hakimullina, A.M.; Popov, D.V.; Lyubaeva, E.V.; Missina, S.S.; Vinogradova, O.L.; Williams, A.G.; Rogozkin, V.A. Association of the VEGFR2 gene His472Gln polymorphism with endurance-related phenotypes. Eur. J. Appl. Physiol. 2009, 107, 95–103. [Google Scholar] [CrossRef]
- Semenova, E.A.; Zempo, H.; Miyamoto-Mikami, E.; Kumagai, H.; Larin, A.K.; Sultanov, R.I.; Babalyan, K.A.; Zhelankin, A.V.; Tobina, T.; Shiose, K.; et al. Genome-Wide Association Study Identifies CDKN1A as a Novel Locus Associated with Muscle Fiber Composition. Cells 2022, 11, 3910. [Google Scholar] [CrossRef]
- GTEx Portal. Available online: https://gtexportal.org (accessed on 25 November 2022).
- Piras, I.S.; Krate, J.; Schrauwen, I.; Corneveaux, J.J.; Serrano, G.E.; Sue, L.; Beach, T.G.; Huentelman, M.J. Whole transcriptome profiling of the human hippocampus suggests an involvement of the KIBRA rs17070145 polymorphism in differential activation of the MAPK signaling pathway. Hippocampus 2017, 27, 784–793. [Google Scholar] [CrossRef]
- Peng, S.; Zhang, Y.; Zhang, J.; Wang, H.; Ren, B. ERK in learning and memory: A review of recent research. Int. J. Mol. Sci. 2010, 11, 222–232. [Google Scholar] [CrossRef]
- Ciccarelli, A.; Giustetto, M. Role of ERK signaling in activity-dependent modifications of histone proteins. Neuropharmacology 2014, 80, 34–44. [Google Scholar] [CrossRef]
- Palombo, D.J.; Amaral, R.S.; Olsen, R.K.; Müller, D.J.; Todd, R.M.; Anderson, A.K.; Levine, B. KIBRA polymorphism is associated with individual differences in hippocampal subregions: Evidence from anatomical segmentation using high-resolution MRI. J. Neurosci. 2013, 33, 13088–13093. [Google Scholar] [CrossRef]
- Witte, A.V.; Köbe, T.; Kerti, L.; Rujescu, D.; Flöel, A. Impact of KIBRA Polymorphism on Memory Function and the Hippocampus in Older Adults. Neuropsychopharmacology 2016, 41, 781–790. [Google Scholar] [CrossRef]
- Li, R.; Wan, W.; Li, J. KIBRA polymorphism modulates gray matter volume to influence cognitive ability in the elderly. Brain Imaging Behav. 2020, 14, 1388–1394. [Google Scholar] [CrossRef]
- Corneveaux, J.J.; Liang, W.S.; Reiman, E.M.; Webster, J.A.; Myers, A.J.; Zismann, V.L.; Joshipura, K.D.; Pearson, J.V.; Hu-Lince, D.; Craig, D.W.; et al. Evidence for an association between KIBRA and late-onset Alzheimer’s disease. Neurobiol. Aging 2010, 31, 901–909. [Google Scholar] [CrossRef]
- Reiman, E.M.; Chen, K.; Alexander, G.E.; Caselli, R.J.; Bandy, D.; Osborne, D.; Saunders, A.M.; Hardy, J. Correlations between apolipoprotein E epsilon4 gene dose and brain-imaging measurements of regional hypometabolism. Proc. Natl. Acad. Sci. USA 2005, 102, 8299–8302. [Google Scholar] [CrossRef]
- Porter, T.; Burnham, S.C.; Doré, V.; Savage, G.; Bourgeat, P.; Begemann, K.; Milicic, L.; Ames, D.; Bush, A.I.; Maruff, P.; et al. KIBRA is associated with accelerated cognitive decline and hippocampal atrophy in APOE ε4-positive cognitively normal adults with high Aβ-amyloid burden. Sci. Rep. 2018, 8, 2034. [Google Scholar] [CrossRef]
- Stickel, A.; Kawa, K.; Walther, K.; Glisky, E.; Richholt, R.; Huentelman, M.; Ryan, L. Age-Modulated Associations between KIBRA, Brain Volume, and Verbal Memory among Healthy Older Adults. Front. Aging Neurosci. 2018, 9, 431. [Google Scholar] [CrossRef]
- Ling, J.; Huang, Y.; Zhang, L.; Wei, D.; Cheng, W. Association of KIBRA polymorphism with risk of Alzheimer’s disease: Evidence based on 20 case-control studies. Neurosci. Lett. 2018, 662, 77–83. [Google Scholar] [CrossRef]
- Wang, Y.; Zuo, C.; Wang, D.; Tao, S.; Hao, L. Reduced Thalamus Volume and Enhanced Thalamus and Fronto-Parietal Network Integration in the Chess Experts. Cereb. Cortex 2020, 30, 5560–5569. [Google Scholar] [CrossRef]
- Vinkhuyzen, A.A.; van der Sluis, S.; Posthuma, D.; Boomsma, D.I. The heritability of aptitude and exceptional talent across different domains in adolescents and young adults. Behav. Genet. 2009, 39, 380–392. [Google Scholar] [CrossRef]
- Yasuda, Y.; Hashimoto, R.; Ohi, K.; Fukumoto, M.; Takamura, H.; Iike, N.; Yoshida, T.; Hayashi, N.; Takahashi, H.; Yamamori, H.; et al. Association study of KIBRA gene with memory performance in a Japanese population. World J. Biol. Psychiatry 2010, 11, 852–857. [Google Scholar] [CrossRef]
- Valeeva, E.V.; Ahmetov, I.I.; Rees, T. Psychogenetics and sport. In Sports, Exercise, and Nutritional Genomics; Academic Press: Cambridge, MA, USA, 2019; pp. 147–165. [Google Scholar] [CrossRef]
- Boulygina, E.A.; Borisov, O.V.; Valeeva, E.V.; Semenova, E.A.; Kostryukova, E.S.; Kulemin, N.A.; Larin, A.K.; Nabiullina, R.M.; Mavliev, F.A.; Akhatov, A.M.; et al. Whole genome sequencing of elite athletes. Biol. Sport. 2020, 37, 295–304. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Hall, E.C.R.; Semenova, E.A.; Pranckevičienė, E.; Ginevičienė, V. Advances in sports genomics. Adv. Clin. Chem. 2022, 107, 215–263. [Google Scholar] [CrossRef]
- Lounsbury, J.W.; Foster, N.; Patel, H.; Carmody, P.; Gibson, L.W.; Stairs, D.R. An investigation of the personality traits of scientists versus nonscientists and their relationship with career satisfaction. R&D Manag. 2012, 42, 47–59. [Google Scholar] [CrossRef]
- Webb, R.M.; Lubinski, D.; Benbow, C.P. Spatial ability: A neglected dimension in talent searches for intellectually precocious youth. J. Educ. Psychol. 2007, 99, 397–420. [Google Scholar] [CrossRef]
- Harris, J.A. Measured intelligence, achievement, openness to experience, and creativity. Personal. Individ. Differ. 2004, 36, 913–929. [Google Scholar] [CrossRef]
- Anglim, J.; Dunlop, P.D.; Wee, S.; Horwood, S.; Wood, J.K.; Marty, A. Personality and intelligence: A meta-analysis. Psychol. Bull. 2022, 148, 301–336. [Google Scholar] [CrossRef]
- Remoli, T.C.; Santos, F.H. Interactions between working memory and creativity: A systematic review. Psicol. Estud. 2017, 22, 53–65. [Google Scholar] [CrossRef]
- Mitolo, M.; Gardini, S.; Caffarra, P.; Ronconi, L.; Venneri, A.; Pazzaglia, F. Relationship between spatial ability, visuospatial working memory and self-assessed spatial orientation ability: A study in older adults. Cogn. Process. 2015, 16, 165–176. [Google Scholar] [CrossRef]
- UK Biobank GWAS round 2 results [released 1st August 2018]. Available online: http://www.nealelab.is/uk-biobank/ (accessed on 25 November 2022).
- Okbay, A.; Wu, Y.; Wang, N.; Jayashankar, H.; Bennett, M.; Nehzati, S.M.; Sidorenko, J.; Kweon, H.; Goldman, G.; Gjorgjieva, T.; et al. Polygenic prediction of educational attainment within and between families from genome-wide association analyses in 3 million individuals. Nat. Genet. 2022, 54, 437–449. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Groups | Males | Females | ||
|---|---|---|---|---|
| n | Age, Years | n | Age, Years | |
| Kazakh chess players | 46 | 17.0 (3.9) | 13 | 16.3 (4.4) |
| Kazakh controls | 111 | 27.5 (4.6) | 66 | 25.0 (3.6) |
| Russian chess players | 44 | 22.2 (3.2) | 23 | 22.8 (3.1) |
| Russian PhD holders | 34 | 27.6 (2.8) | 43 | 30.4 (4.9) |
| Russian controls | 831 | 27.1 (4.3) | 608 | 27.2 (4.4) |
| Tatar chess players | 48 | 21.9 (3.8) | 20 | 21.4 (3.2) |
| Tatar PhD holders | 25 | 30.2 (4.7) | 17 | 32.2 (6.5) |
| Tatar controls | 278 | 25.7 (4.8) | 272 | 25.1 (4.3) |
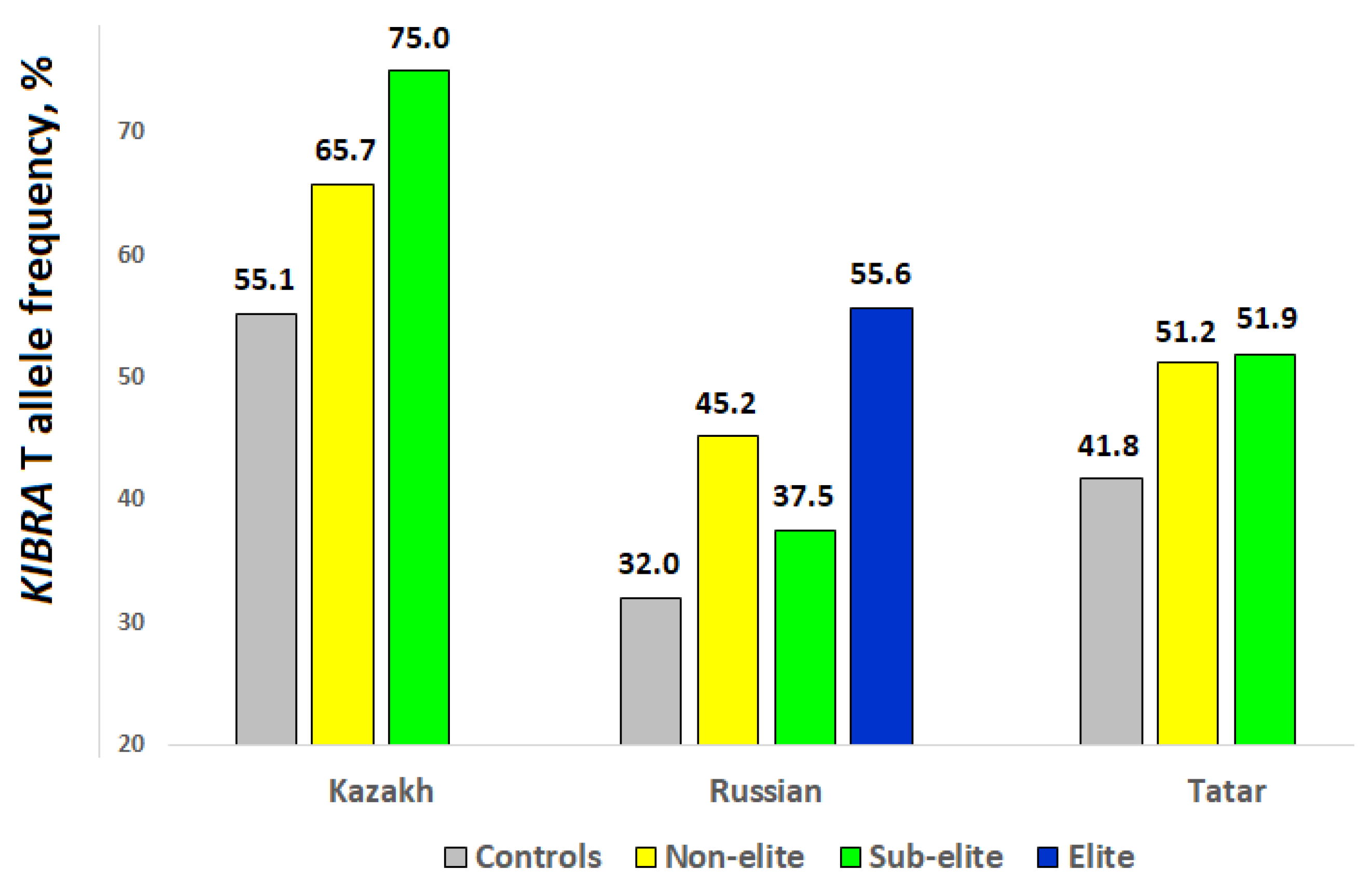
| Cohort | n | KIBRA Genotypes | T Allele, % | p | ||
|---|---|---|---|---|---|---|
| CC | CT | TT | ||||
| Kazakh chess players | 59 | 4 | 31 | 24 | 66.9 | 0.024 * |
| Kazakh controls | 177 | 34 | 91 | 52 | 55.1 | - |
| Russian chess players | 67 | 20 | 34 | 13 | 44.8 | 0.0027 * |
| Russian PhD holders | 77 | 27 | 37 | 13 | 40.9 | 0.026 * |
| Russian controls | 1439 | 666 | 626 | 147 | 32.0 | - |
| Tatar chess players | 68 | 16 | 34 | 18 | 51.5 | 0.035 * |
| Tatar PhD holders | 42 | 9 | 25 | 8 | 48.8 | 0.252 |
| Tatar controls | 550 | 197 | 246 | 107 | 41.8 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmetov, I.I.; Valeeva, E.V.; Yerdenova, M.B.; Datkhabayeva, G.K.; Bouzid, A.; Bhamidimarri, P.M.; Sharafetdinova, L.M.; Egorova, E.S.; Semenova, E.A.; Gabdrakhmanova, L.J.; et al. KIBRA Gene Variant Is Associated with Ability in Chess and Science. Genes 2023, 14, 204. https://doi.org/10.3390/genes14010204
Ahmetov II, Valeeva EV, Yerdenova MB, Datkhabayeva GK, Bouzid A, Bhamidimarri PM, Sharafetdinova LM, Egorova ES, Semenova EA, Gabdrakhmanova LJ, et al. KIBRA Gene Variant Is Associated with Ability in Chess and Science. Genes. 2023; 14(1):204. https://doi.org/10.3390/genes14010204
Chicago/Turabian StyleAhmetov, Ildus I., Elena V. Valeeva, Meruert B. Yerdenova, Gaukhar K. Datkhabayeva, Amal Bouzid, Poorna Manasa Bhamidimarri, Liliya M. Sharafetdinova, Emiliya S. Egorova, Ekaterina A. Semenova, Leysan J. Gabdrakhmanova, and et al. 2023. "KIBRA Gene Variant Is Associated with Ability in Chess and Science" Genes 14, no. 1: 204. https://doi.org/10.3390/genes14010204
APA StyleAhmetov, I. I., Valeeva, E. V., Yerdenova, M. B., Datkhabayeva, G. K., Bouzid, A., Bhamidimarri, P. M., Sharafetdinova, L. M., Egorova, E. S., Semenova, E. A., Gabdrakhmanova, L. J., Yusupov, R. A., Larin, A. K., Kulemin, N. A., Generozov, E. V., Hamoudi, R., Kustubayeva, A. M., & Rees, T. (2023). KIBRA Gene Variant Is Associated with Ability in Chess and Science. Genes, 14(1), 204. https://doi.org/10.3390/genes14010204