
Validation of a Chromosome 14 Risk Haplotype for Idiopathic Epilepsy in the Belgian Shepherd Dog Found to Be Associated with an Insertion in the RAPGEF5 Gene

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Haplotype Risk and Frequency
3.2. Candidate Gene Investigation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belanger, J.; Famula, T.; Gershony, L.; Palij, M.; Oberbauer, A. Genome-wide association analysis of idiopathic epilepsy in the Belgian shepherd. Canine Med. Genet. 2020, 7, e33549. [Google Scholar] [CrossRef]
- Famula, T.; Oberbauer, A.; Brown, K. Heritability of epileptic seizures in the Belgian tervueren. J. Small Anim. Pract. 1997, 38, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Gulløv, C.; Toft, N.; Berendt, M. A longitudinal study of survival in Belgian Shepherds with genetic epilepsy. J. Vet. Intern. Med. 2012, 26, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Koskinen, L.L.; Seppälä, E.H.; Weissl, J.; Jokinen, T.S.; Viitmaa, R.; Hänninen, R.L.; Quignon, P.; Fischer, A.; André, C.; Lohi, H. ADAM23 is a common risk gene for canine idiopathic epilepsy. BMC Genet. 2017, 18, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberbauer, A.; Grossman, D.; Irion, D.; Schaffer, A.; Eggleston, M.; Famula, T. The genetics of epilepsy in the Belgian tervuren and sheepdog. J. Hered. 2003, 94, 57–63. [Google Scholar] [CrossRef] [Green Version]
- De Risio, L.; Bhatti, S.; Muñana, K.; Penderis, J.; Stein, V.; Tipold, A.; Berendt, M.; Farqhuar, R.; Fischer, A.; Long, S. International veterinary epilepsy task force consensus proposal: Diagnostic approach to epilepsy in dogs. BMC Vet. Res. 2015, 11, 148. [Google Scholar]
- Rincon, G.; Tengvall, K.; Belanger, J.M.; Lagoutte, L.; Medrano, J.F.; André, C.; Thomas, A.; Lawley, C.T.; Hansen, M.S.; Lindblad-Toh, K. Comparison of buccal and blood-derived canine DNA, either native or whole genome amplified, for array-based genome-wide association studies. BMC Res. Notes 2011, 4, 226. [Google Scholar] [CrossRef]
- Koskinen, L.L.; Seppälä, E.H.; Belanger, J.M.; Arumilli, M.; Hakosalo, O.; Jokinen, P.; Nevalainen, E.M.; Viitmaa, R.; Jokinen, T.S.; Oberbauer, A.M. Identification of a common risk haplotype for canine idiopathic epilepsy in the ADAM23 gene. BMC Genom. 2015, 16, 465. [Google Scholar] [CrossRef] [Green Version]
- van Hagen, M.A.; Schipper, L.; Oosterveer-van der Doelen, M.A.; Vos-Loohuis, M.; Gehring, R.; Leegwater, P.A. Analysis of polymorphisms of canine Cytochrome P 450-CYP2D15. J. Vet. Pharmacol. Ther. 2020, 43, 602–607. [Google Scholar] [CrossRef]
- Dahlgren, S.; Ziener, M.L.; Lingaas, F. A genome-wide association study identifies a region strongly associated with symmetrical onychomadesis on chromosome 12 in dogs. Anim. Genet. 2016, 47, 708–716. [Google Scholar] [CrossRef]
- Forsberg, S.K.; Kierczak, M.; Ljungvall, I.; Merveille, A.-C.; Gouni, V.; Wiberg, M.; Lundgren Willesen, J.; Hanås, S.; Lequarre, A.-S.; Mejer Sørensen, L. The shepherds’ tale: A genome-wide study across 9 dog breeds implicates two loci in the regulation of fructosamine serum concentration in Belgian shepherds. PLoS ONE 2015, 10, e0123173. [Google Scholar]
- Jagannathan, V.; Drögemüller, C.; Leeb, T.; Consortium, D.B.V.D.; Aguirre, G.; André, C.; Bannasch, D.; Becker, D.; Davis, B.; Ekenstedt, K. A comprehensive biomedical variant catalogue based on whole genome sequences of 582 dogs and eight wolves. Anim. Genet. 2019, 50, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsson, M.; Tengvall, K.; Frankowiack, M.; Kierczak, M.; Bergvall, K.; Axelsson, E.; Tintle, L.; Marti, E.; Roosje, P.; Leeb, T. Genome-wide analyses suggest mechanisms involving early B-cell development in canine IgA deficiency. PLoS ONE 2015, 10, e0133844. [Google Scholar]
- Stern, J.A.; Hsue, W.; Song, K.-H.; Ontiveros, E.S.; Luis Fuentes, V.; Stepien, R.L. Severity of mitral valve degeneration is associated with chromosome 15 loci in whippet dogs. PLoS ONE 2015, 10, e0141234. [Google Scholar] [CrossRef] [PubMed]
- Vaysse, A.; Ratnakumar, A.; Derrien, T.; Axelsson, E.; Rosengren Pielberg, G.; Sigurdsson, S.; Fall, T.; Seppälä, E.H.; Hansen, M.S.; Lawley, C.T. Identification of genomic regions associated with phenotypic variation between dog breeds using selection mapping. PLoS Genet. 2011, 7, e1002316. [Google Scholar] [CrossRef] [Green Version]
- Wolf, Z.T.; Brand, H.A.; Shaffer, J.R.; Leslie, E.J.; Arzi, B.; Willet, C.E.; Cox, T.C.; McHenry, T.; Narayan, N.; Feingold, E. Genome-wide association studies in dogs and humans identify ADAMTS20 as a risk variant for cleft lip and palate. PLoS Genet. 2015, 11, e1005059. [Google Scholar] [CrossRef] [Green Version]
- Gershony, L.C.; Belanger, J.M.; Hytönen, M.K.; Lohi, H.; Oberbauer, A.M. Whole Genome Sequencing Reveals Multiple Linked Genetic Variants on Canine Chromosome 12 Associated with Risk for Symmetrical Lupoid Onychodystrophy (SLO) in the Bearded Collie. Genes 2021, 12, 1265. [Google Scholar] [CrossRef]
- Garrison, E.; Marth, G. Haplotype-based variant detection from short-read sequencing. arXiv 2012, arXiv:1207.3907. [Google Scholar]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef]
- Bithell, A.; Alberta, J.; Hornby, F.; Stiles, C.D.; Williams, B.P. Expression of the guanine nucleotide exchange factor, mr-gef, is regulated during the differentiation of specific subsets of telencephalic neurons. Dev. Brain Res. 2003, 146, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Knott, S.; Forty, L.; Craddock, N.; Thomas, R.H. Epilepsy and bipolar disorder. Epilepsy Behav. 2015, 52, 267–274. [Google Scholar] [PubMed]
- Winden, K.D.; Bragin, A.; Engel, J.; Geschwind, D.H. Molecular alterations in areas generating fast ripples in an animal model of temporal lobe epilepsy. Neurobiol. Dis. 2015, 78, 35–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seppälä, E.H.; Koskinen, L.L.; Gulløv, C.H.; Jokinen, P.; Karlskov-Mortensen, P.; Bergamasco, L.; Baranowska Körberg, I.; Cizinauskas, S.; Oberbauer, A.M.; Berendt, M. Identification of a novel idiopathic epilepsy locus in Belgian Shepherd dogs. PLoS ONE 2012, 7, e33549. [Google Scholar] [CrossRef] [PubMed]
- Bithell, A.; Hsu, T.; Kandanearatchi, A.; Landau, S.; Everall, I.P.; Tsuang, M.T.; Chana, G.; Williams, B.P. Expression of the Rap1 guanine nucleotide exchange factor, MR-GEF, is altered in individuals with bipolar disorder. PLoS ONE 2010, 5, e10392. [Google Scholar] [CrossRef]
- Barrett, K.T.; Choudhary, A.; Charkhand, B.; Scantlebury, M.H. Animal Models in Epileptic Spasms and the Development of Novel Treatment Options. J. Clin. Neurophysiol. 2022; ahead of print. [Google Scholar] [CrossRef]
- Yang, C.; Shi, Y.; Li, X.; Guan, L.; Li, H.; Lin, J. Cadherins and the pathogenesis of epilepsy. Cell Biochem. Funct. 2022, 40, 336–348. [Google Scholar] [CrossRef]
- Griffin, J.N.; Del Viso, F.; Duncan, A.R.; Robson, A.; Hwang, W.; Kulkarni, S.; Liu, K.J.; Khokha, M.K. RAPGEF5 regulates nuclear translocation of β-catenin. Dev. Cell 2018, 44, 248–260.e244. [Google Scholar] [CrossRef] [Green Version]
- Alharatani, R.; Griffin, J.N.; Liu, K.J. Expression of the guanine nucleotide exchange factor, RAPGEF5, during mouse and human embryogenesis. Gene Expr. Patterns 2019, 34, 119057. [Google Scholar] [CrossRef]
- Ishiura, H.; Mitsui, J.; Yoshimura, J.; Matsukawa, M.K.; Fujiyama, A.; Toyoshima, Y.; Kakita, A.; Takahashi, H.; Suzuki, Y.; Sugano, S. Expansions of intronic TTTCA and TTTTA repeats in benign adult familial myoclonic epilepsy. Nat. Genet. 2018, 50, 581–590. [Google Scholar] [CrossRef]
- Maeta, K.; Hattori, S.; Ikutomo, J.; Edamatsu, H.; Bilasy, S.E.; Miyakawa, T.; Kataoka, T. Comprehensive behavioral analysis of mice deficient in Rapgef2 and Rapgef6, a subfamily of guanine nucleotide exchange factors for Rap small GTPases possessing the Ras/Rap-associating domain. Mol. Brain 2018, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Consonni, S.V.; Maurice, M.M.; Bos, J.L. DEP domains: Structurally similar but functionally different. Nat. Rev. Mol. Cell Biol. 2014, 15, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.H.; Lee, H.; Song, D.H.; Eom, J.-H.; Kim, S.C.; Lee, H.-S.; Lee, H.; Lee, J.-O. Connecting two proteins using a fusion alpha helix stabilized by a chemical cross linker. Nat. Commun. 2016, 7, 11031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
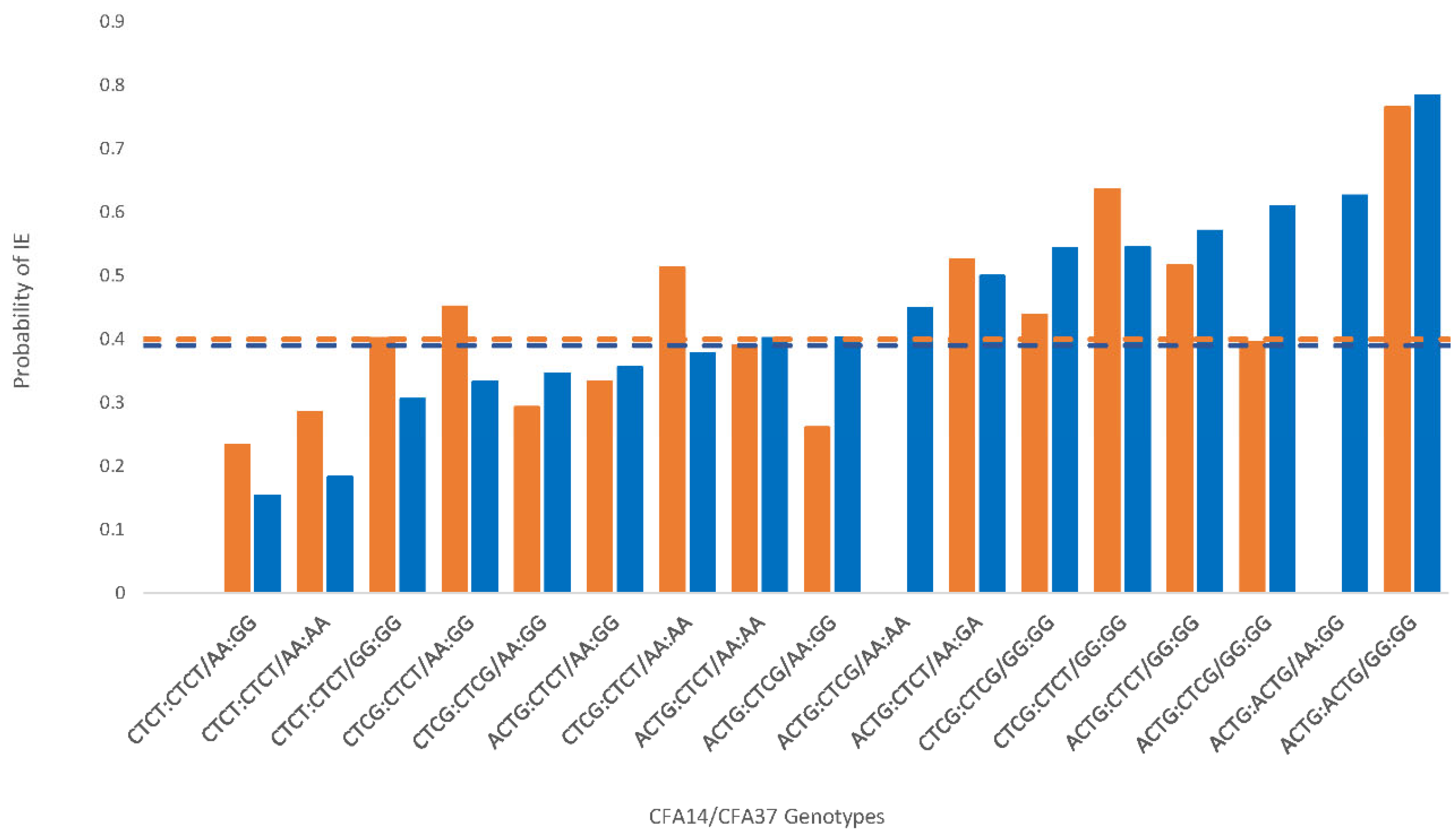

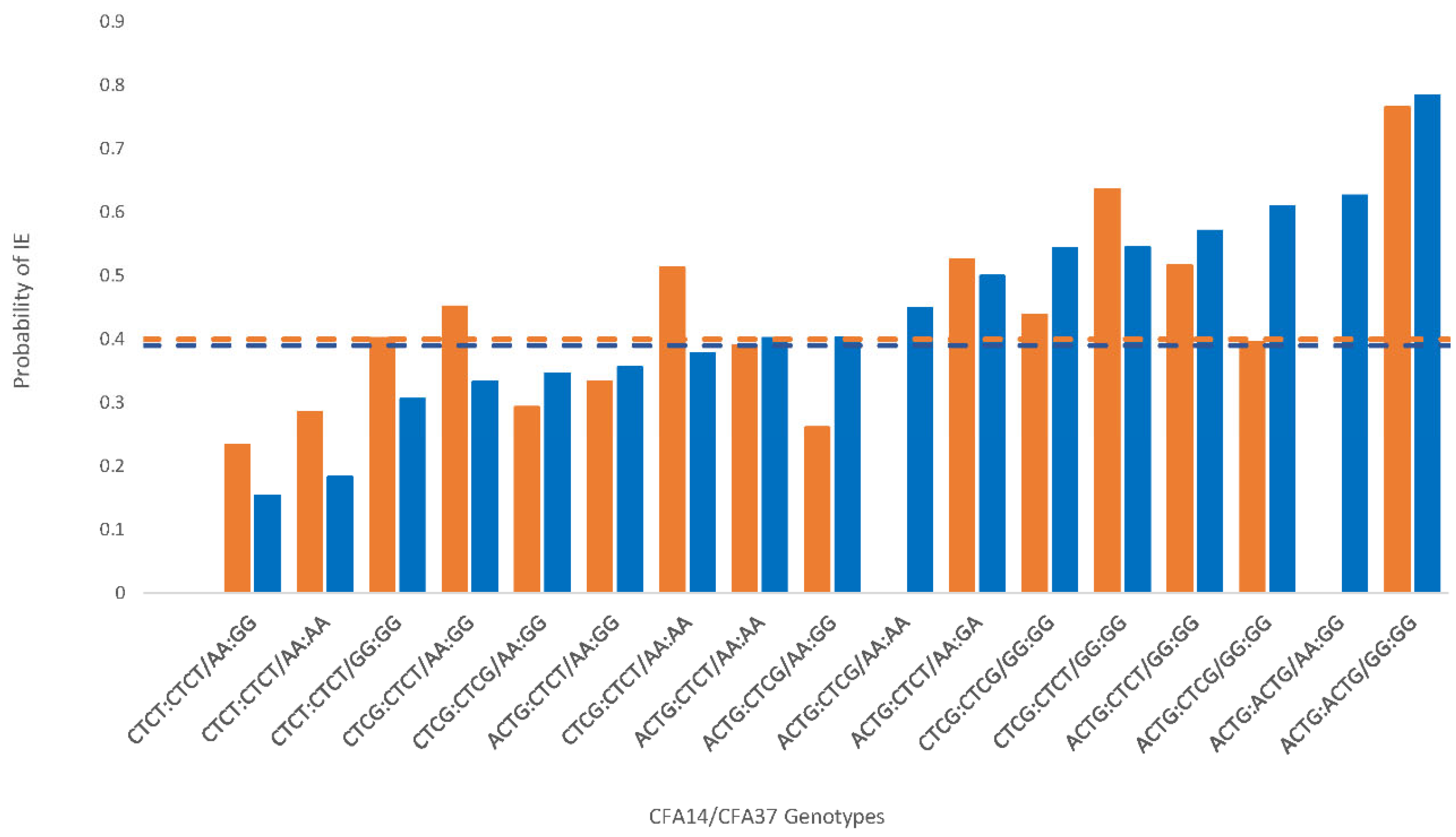{kind=link}
| New Cohort | Combined Cohort | |||||||
|---|---|---|---|---|---|---|---|---|
| CFA14 Four SNP Risk Haplotype | Control (n = 74) | IE Case (n = 47) | OR (95% CI) | p-Value | Control (n = 117) | IE Case (n = 66) | OR (95% CI) | p-Value |
| ACTG (htz or hmz *) vs. | 25 | 20 | 1.45 (0.68–3.08) | 3.42 × 10−1 | 31 | 34 | 2.95 (1.56–5.56) | 8.01 × 10−4 |
| non-ACTG | 49 | 27 | 86 | 32 | ||||
| CTCG (htz or hmz) vs. | 13 | 11 | 1.43 (0.58–3.54) | 4.87 × 10−1 | 24 | 19 | 1.41 (0.71–2.82) | 3.72 × 10−1 |
| non-CTCG | 61 | 36 | 81 | 47 | ||||
| CTCT (htz or hmz) vs. | 69 | 42 | 0.61 (0.17–2.23) | 5.08 × 10−1 | 99 | 55 | 0.32 (0.12–0.87) | 2.33 × 10−2 |
| non-CTCT | 5 | 5 | 6 | 11 | ||||
| CFA37 two SNP risk haplotype | Control (n = 74) | IE Case (n = 47) | OR (95% CI) | p-value | Control (n = 117) | IE Case (n = 66) | OR (95% CI) | p-value |
| GG (htz or hmz) vs. | 60 | 41 | 1.59 (0.57–4.49) | 4.57 × 10−1 | 85 | 60 | 2.32 (0.89–6.04) | 9.03 × 10−2 |
| non-GG | 14 | 6 | 20 | 6 | ||||
| AA (htz or hmz) vs. | 51 | 19 | 0.31 (0.14–0.66) | 2.54 × 10−3 | 66 | 21 | 0.27 (0.14–0.51) | 5.57 × 10−5 |
| non-AA | 23 | 28 | 39 | 49 | ||||
| Belgian Shepherds 2n = 1192 | Other Breeds 2n = 4484 | |||
|---|---|---|---|---|
| CFA14 Alleles | Total | % Freq | Total | % Freq |
| ACCG | 0 | 0 | 6 | 0.13 |
| ACCT | 0 | 0 | 2 | 0.04 |
| ACTG | 233 | 19.55 | 211 | 4.71 |
| ACTT | 3 | 0.25 | 89 | 1.98 |
| CCCG | 3 | 0.25 | 1000 | 22.30 |
| CCCT | 8 | 0.67 | 776 | 17.31 |
| CCTG | 0 | 0 | 25 | 0.56 |
| CCTT | 0 | 0 | 77 | 1.72 |
| CTCG | 159 | 13.34 | 704 | 15.70 |
| CTCT | 786 | 65.94 | 1585 | 35.35 |
| CTTT | 0 | 0 | 9 | 0.20 |
| Total | 1192 | 100 | 4484 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belanger, J.M.; Heinonen, T.; Famula, T.R.; Mandigers, P.J.J.; Leegwater, P.A.; Hytönen, M.K.; Lohi, H.; Oberbauer, A.M. Validation of a Chromosome 14 Risk Haplotype for Idiopathic Epilepsy in the Belgian Shepherd Dog Found to Be Associated with an Insertion in the RAPGEF5 Gene. Genes 2022, 13, 1124. https://doi.org/10.3390/genes13071124
Belanger JM, Heinonen T, Famula TR, Mandigers PJJ, Leegwater PA, Hytönen MK, Lohi H, Oberbauer AM. Validation of a Chromosome 14 Risk Haplotype for Idiopathic Epilepsy in the Belgian Shepherd Dog Found to Be Associated with an Insertion in the RAPGEF5 Gene. Genes. 2022; 13(7):1124. https://doi.org/10.3390/genes13071124
Chicago/Turabian StyleBelanger, Janelle M., Tiina Heinonen, Thomas R. Famula, Paul J. J. Mandigers, Peter A. Leegwater, Marjo K. Hytönen, Hannes Lohi, and Anita M. Oberbauer. 2022. "Validation of a Chromosome 14 Risk Haplotype for Idiopathic Epilepsy in the Belgian Shepherd Dog Found to Be Associated with an Insertion in the RAPGEF5 Gene" Genes 13, no. 7: 1124. https://doi.org/10.3390/genes13071124
APA StyleBelanger, J. M., Heinonen, T., Famula, T. R., Mandigers, P. J. J., Leegwater, P. A., Hytönen, M. K., Lohi, H., & Oberbauer, A. M. (2022). Validation of a Chromosome 14 Risk Haplotype for Idiopathic Epilepsy in the Belgian Shepherd Dog Found to Be Associated with an Insertion in the RAPGEF5 Gene. Genes, 13(7), 1124. https://doi.org/10.3390/genes13071124