
Correlation between Genomic Variants and Worldwide Epidemiology of Prostate Cancer

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
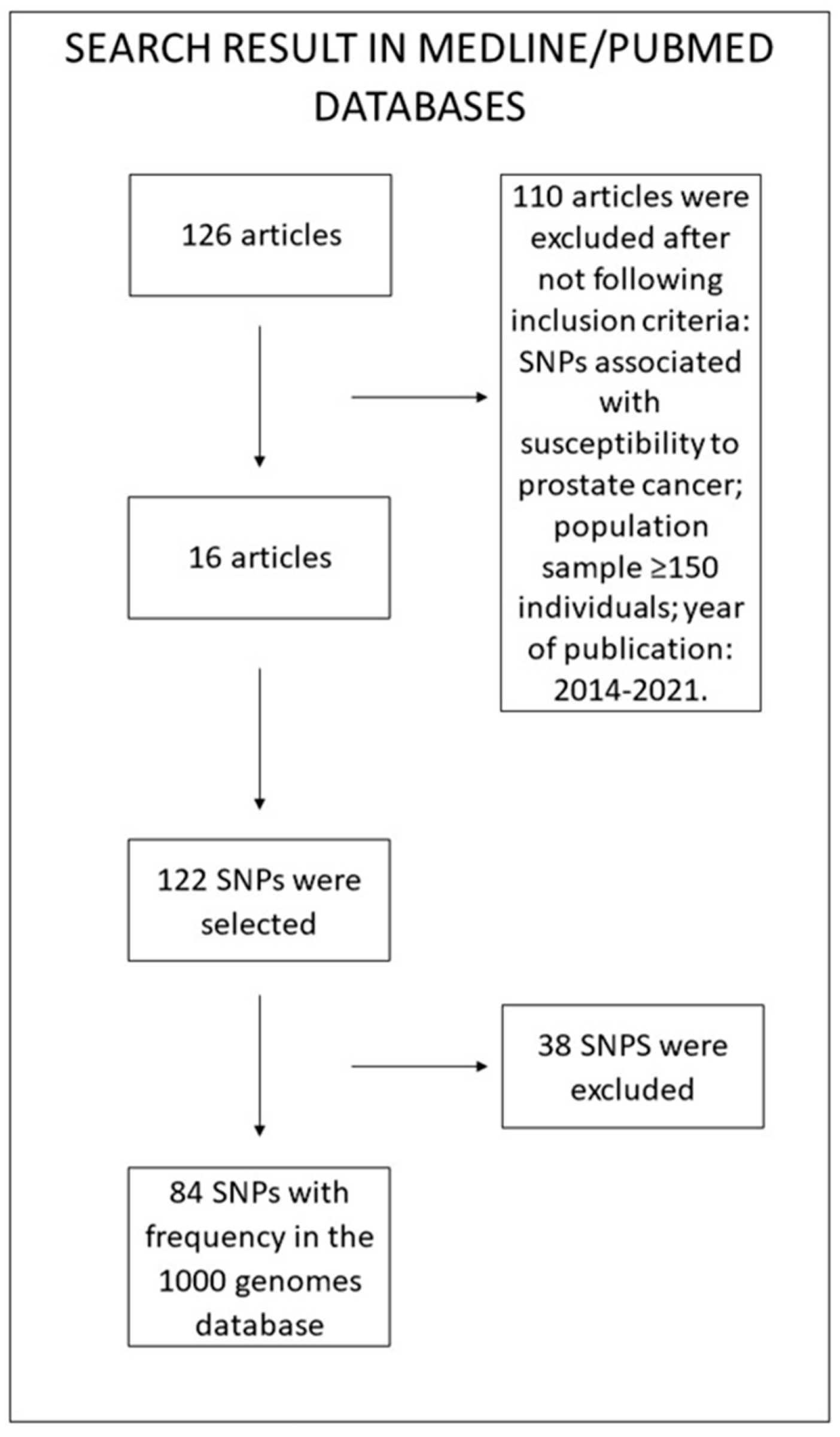
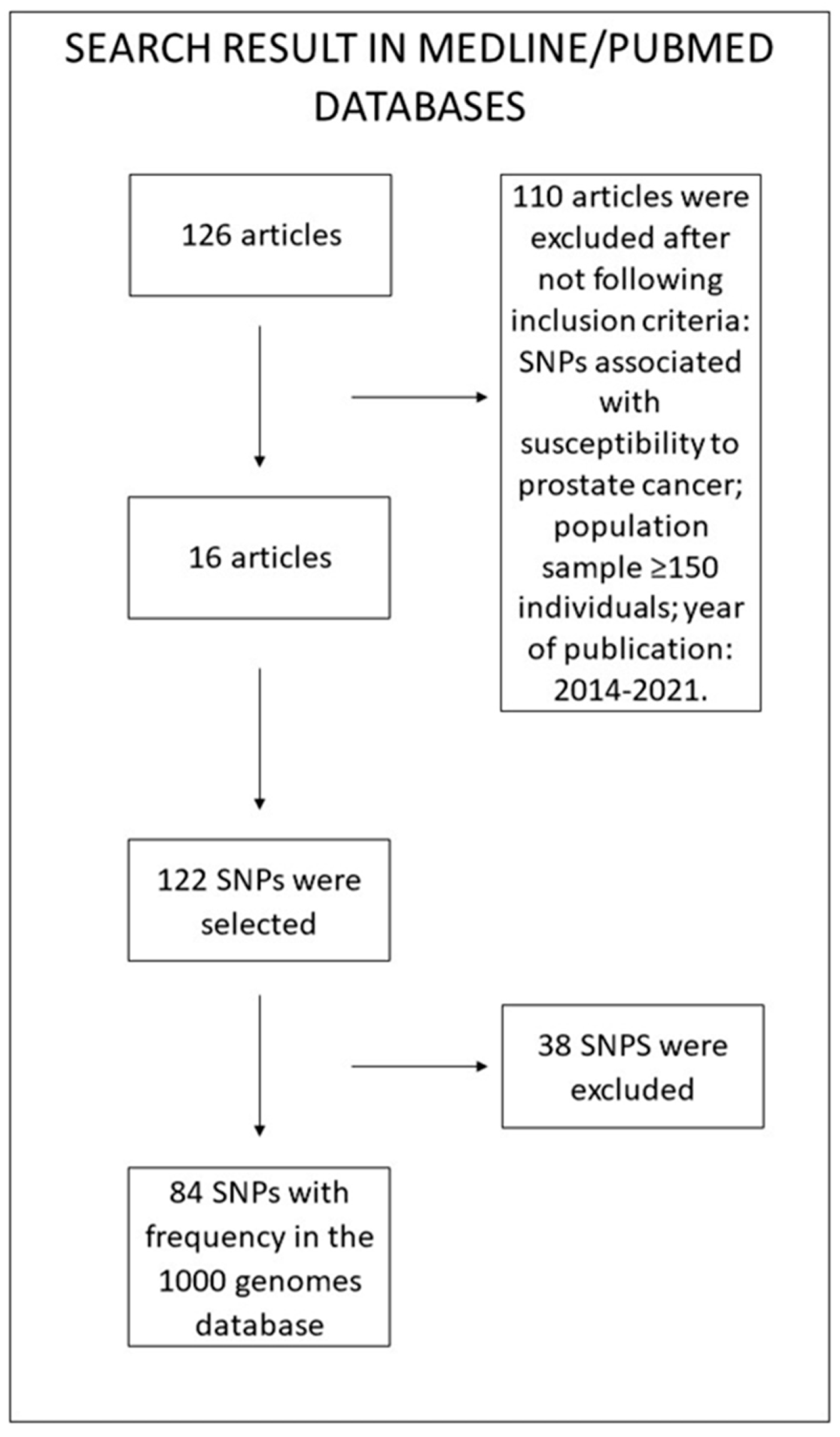
2. Materials and Methods
2.1. SNP Determination
2.2. Epidemiological and Genetic Data
2.3. Statistical Analysis
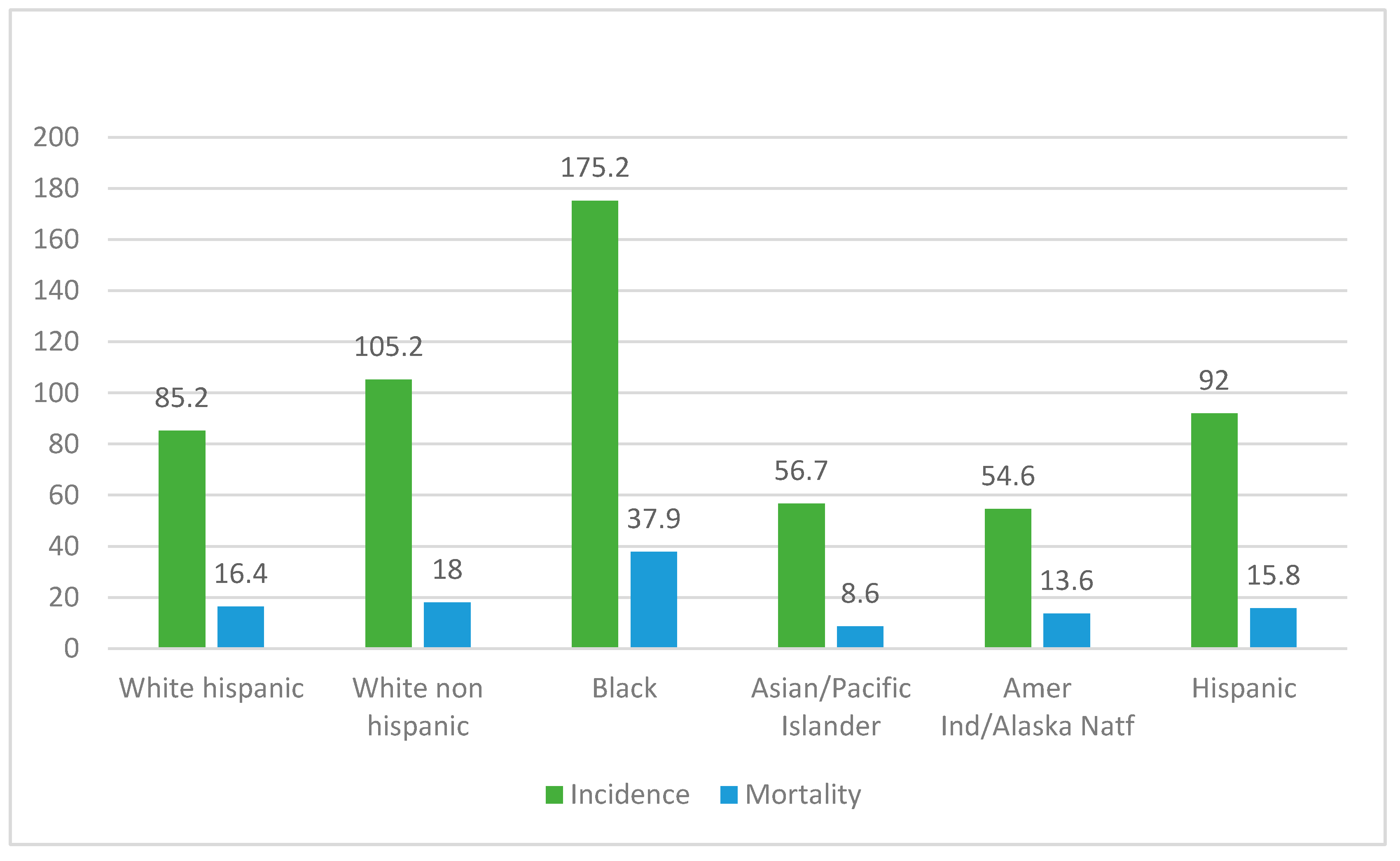
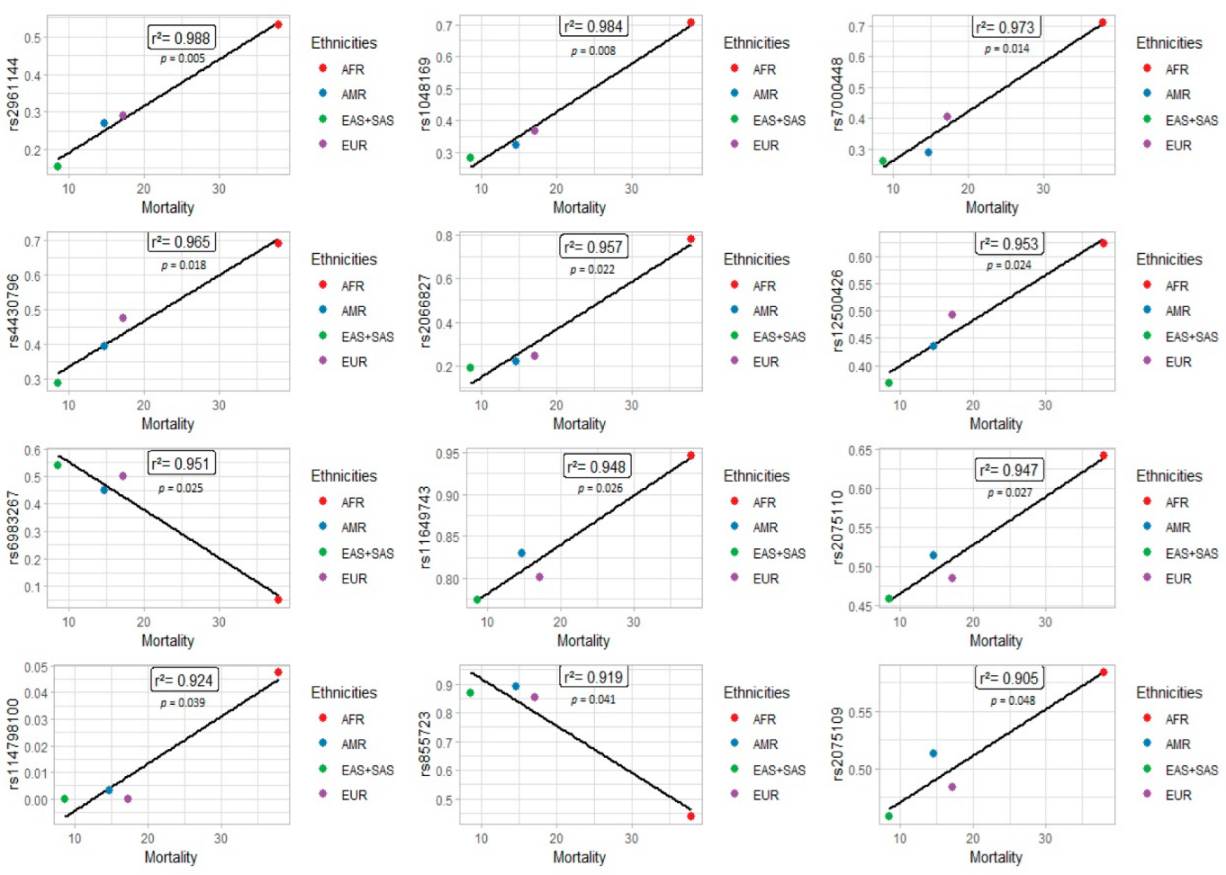
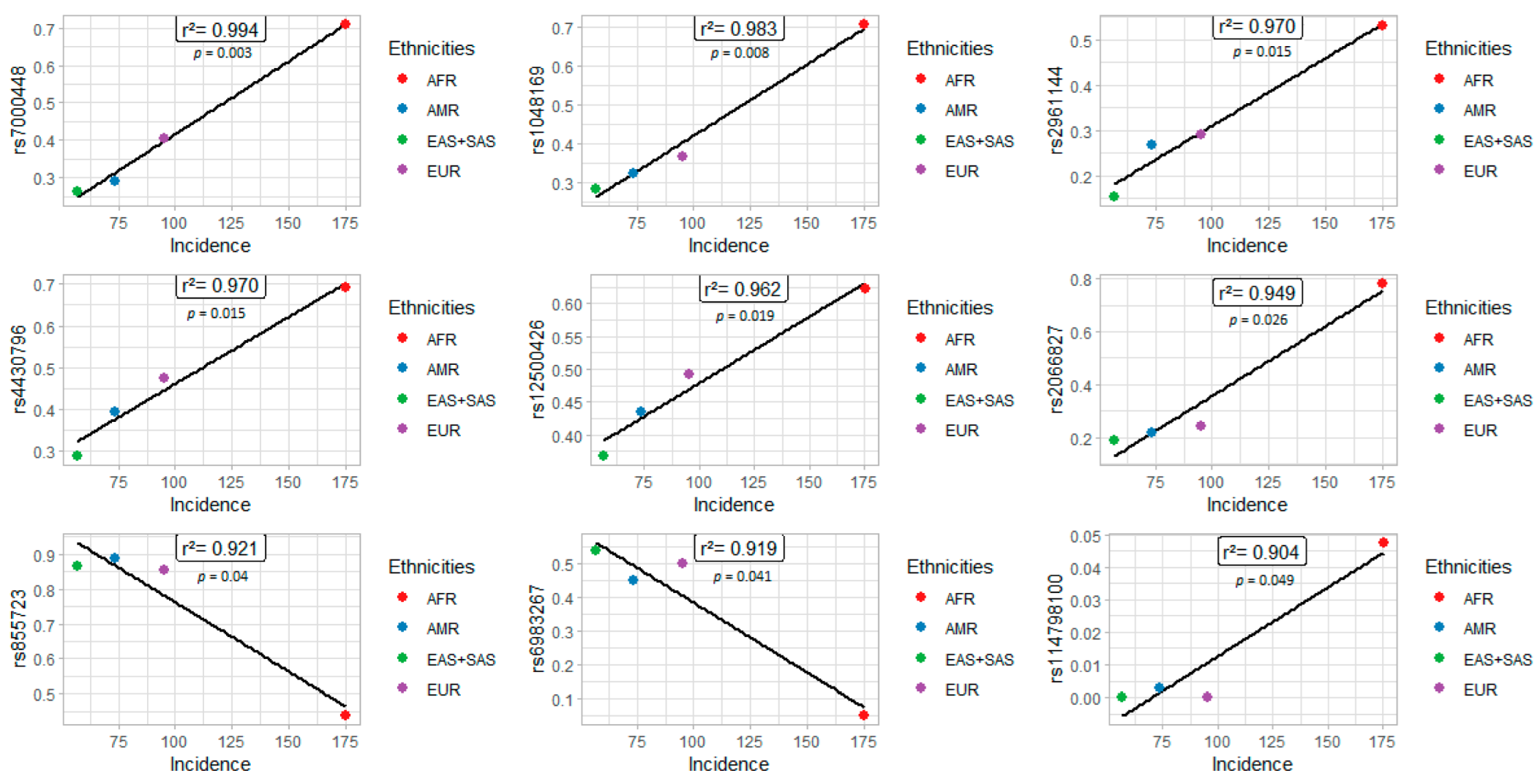
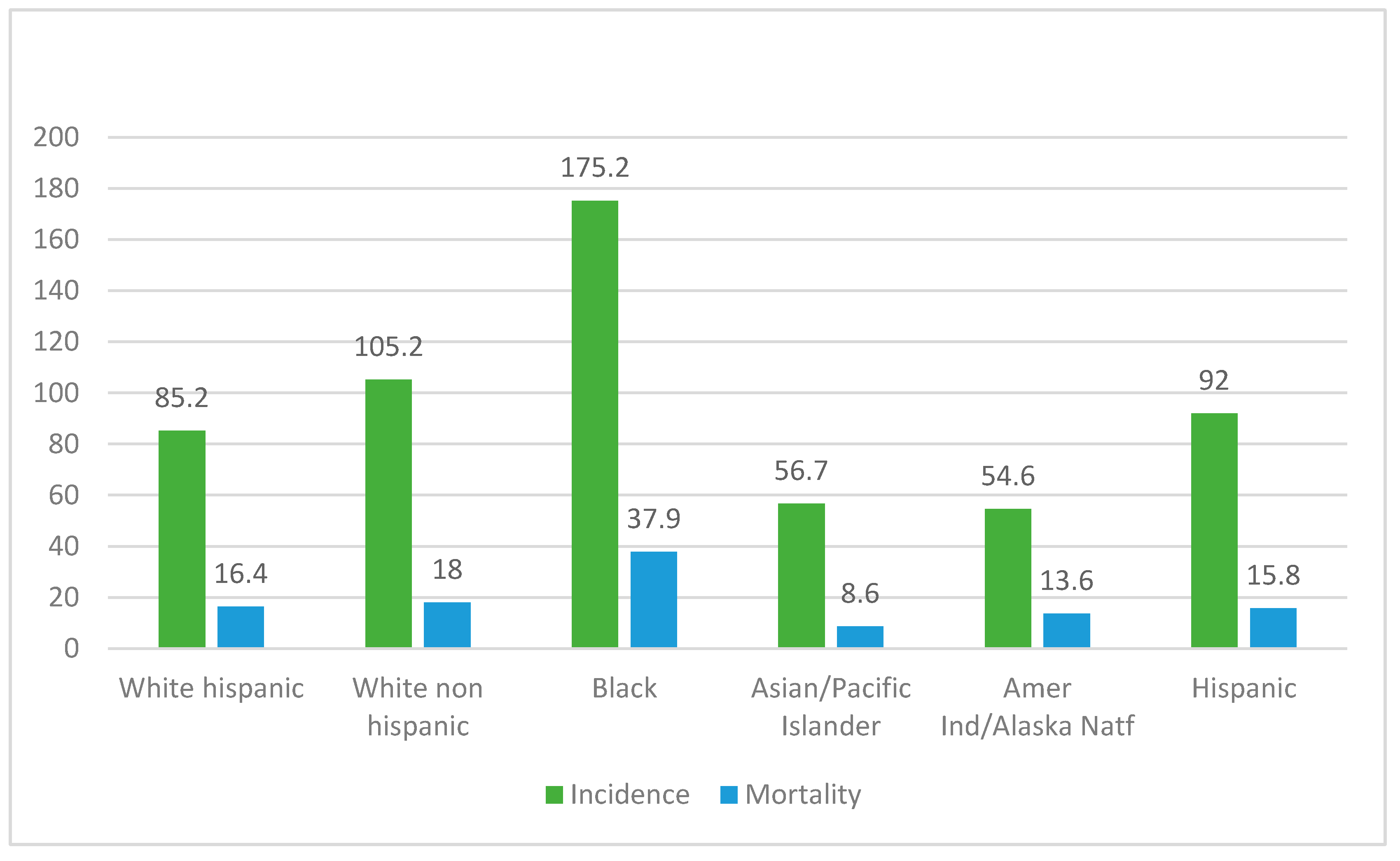
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Conti, D.V.; Darst, B.F.; Moss, L.C.; Saunders, E.J.; Sheng, X.; Chou, A.; Schumacher, F.R.; Al Olama, A.A.; Benlloch, S.; Dadaev, T.; et al. Trans-ancestry genome-wide association meta-analysis of prostate cancer identifies new susceptibility loci and informs genetic risk prediction. Nat. Genet. 2021, 53, 65–75. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute, Surveillance, Epidemiology, and End Results Program (SEER). Cancer Stat Facts: Prostate Cancer. Available online: https://seer.cancer.gov/statfacts/html/prost.html (accessed on 30 March 2021).
- Saad, M.; Mokrab, Y.; Halabi, N.; Shan, J.; Razali, R.; Kunji, K.; Syed, N.; Temanni, R.; Subramanian, M.; Ceccarelli, M.; et al. Genetic predisposition to cancer across people of different ancestries in Qatar: A population-based, cohort study. Lancet Oncol. 2022, 23, 341–352. [Google Scholar] [CrossRef]
- Mucci, L.A.; Hjelmborg, J.B.; Harris, J.R.; Czene, K.; Havelick, D.J.; Scheike, T.; Graff, R.E.; Holst, K.; Möller, S.; Unger, R.H.; et al. Familial risk and heritability of cancer among twins in nordic countries. JAMA J. Am. Med. Assoc. 2016, 315, 68–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allemailem, K.S.; Almatroudi, A.; Alrumaihi, F.; Almansour, N.M.; Aldakheel, F.M.; Rather, R.A.; Afroze, D.; Rah, B. Single nucleotide polymorphisms (SNPs) in prostate cancer: Its implications in diagnostics and therapeutics. Am. J. Transl. Res. 2021, 13, 3868–3889. [Google Scholar]
- Johnson, J.R.; Woods-Burnham, L.; Hooker, S.E.J.; Batai, K.; Kittles, R.A. Genetic contributions to prostate cancer disparities in men of West African descent. Front. Oncol. 2021, 11, 4327. [Google Scholar] [CrossRef]
- Rebbeck, T.R. Prostate cancer genetics: Variation by race, ethnicity, and geography. Semin. Radiat. Oncol. 2016, 27, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Sun, J.; Zhou, H.; Wang, F.; Zhao, C.; Li, K.; Fan, C.; Ding, G.; Wang, J. HNF1B inhibits cell proliferation via repression of SMAD6 expression in prostate cancer. J. Cell. Mol. Med. 2020, 24, 14539–14548. [Google Scholar] [CrossRef]
- Dundr, P.; Bártů, M.; Hojný, J.; Michálková, R.; Hájková, N.; Stružinská, I.; Krkavcová, E.; Hadravský, L.; Kleissnerová, L.; Kopejsková, J.; et al. HNF1B, EZH2 and ECI2 in prostate carcinoma. Molecular, immunohistochemical and clinico-pathological study. Sci. Rep. 2020, 10, 14365. [Google Scholar] [CrossRef]
- Zheng, S.L.; Sun, J.; Wiklund, F.; Smith, S.; Stattin, P.; Li, G.; Adami, H.-O.; Hsu, F.-C.; Zhu, Y.; Bälter, K.; et al. Cumulative association of five genetic variants with prostate cancer. N. Engl. J. Med. 2008, 358, 910–919. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Zheng, S.L.; Wiklund, F.; Isaacs, S.D.; Purcell, L.D.; Gao, Z.; Hsu, F.-C.; Kim, S.-T.; Liu, W.; Zhu, Y.; et al. Evidence for two independent prostate cancer risk–associated loci in the HNF1B gene at 17q12. Nat. Genet. 2008, 40, 1153–1155. [Google Scholar] [CrossRef] [PubMed]
- Mitsudomi, T.; Yatabe, Y. Epidermal growth factor receptor in relation to tumor development: EGFR gene and cancer. FEBS J. 2009, 277, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Li, Z.; Zheng, H.; Chan, M.T.; Wu, W.K.K. CCAT2: A novel oncogenic long non-coding RNA in human cancers. Cell Prolif. 2017, 50, e12342. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Xiong, G.; Guo, W.; Jiang, G.; Li, Y.; Li, H. Long non-coding RNA CCAT2 promotes prostate cancer cell proliferation and invasion by regulating the Wnt/β-catenin signaling pathway. Oncol. Lett. 2020, 20, 97. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.L.; Keller, E.T. The role of Wnts in bone metastases. Cancer Metastasis Rev. 2006, 25, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Thiele, S.; Rauner, M.; Goettsch, C.; Rachner, T.D.; Benad, P.; Fuessel, S.; Erdmann, K.; Hamann, C.; Baretton, G.B.; Wirth, M.P.; et al. Expression profile of WNT molecules in prostate cancer and its regulation by aminobisphosphonates. J. Cell. Biochem. 2011, 112, 1593–1600. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, X.; Xu, Z.; He, Y.; Guo, C.; He, L.; Huan, C.; Cai, C.; Huang, J.; Zhang, J.; et al. PDLIM5 inhibits STUB1-mediated degradation of SMAD3 and promotes the migration and invasion of lung cancer cells. J. Biol. Chem. 2020, 295, 13798–13811. [Google Scholar] [CrossRef]
- Schumacher, F.R.; Al Olama, A.A.; Berndt, S.I.; Benlloch, S.; Ahmed, M.; Saunders, E.J.; Dadaev, T.; Leongamornlert, D.; Anokian, E.; Cieza-Borrella, C.; et al. Association analyses of more than 140,000 men identify 63 new prostate cancer susceptibility loci. Nat. Genet. 2018, 50, 928–936. [Google Scholar] [CrossRef] [Green Version]
- Farashi, S.; Kryza, T.; Clements, J.; Batra, J. Post-GWAS in prostate cancer: From genetic association to biological contribution. Nat. Cancer 2018, 19, 46–59. [Google Scholar] [CrossRef]
- Beuten, J.; Gelfond, J.A.L.; Martinez-Fierro, M.L.; Weldon, K.S.; Crandall, A.C.; Rojas-Martinez, A.; Thompson, I.M.; Leach, R. Association of chromosome 8q variants with prostate cancer risk in Caucasian and Hispanic men. Carcinogenesis 2009, 30, 1372–1379. [Google Scholar] [CrossRef]
- Tan, C.; Hu, W.; Huang, Y.; Zhou, J.; Zheng, S. Risk of eighteen genome-wide association study-identified genetic variants for colorectal cancer and colorectal adenoma in Han Chinese. Oncotarget 2016, 7, 77651–77663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Gao, M.; Yin, Z.; Yan, L.; Cui, L. Association between lncRNA CASC8 polymorphisms and the risk of cancer: A meta-analysis. Cancer Manag. Res. 2018, 10, 3141–3148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, A.B.; Ukoli, F.; Freeman, V.; Bennett, F.; Aiken, W.; Tulloch, T.; Coard, K.; Angwafo, F.; Kittles, R.A. 8q24 risk alleles in West African and Caribbean men. Prostate 2012, 72, 1366–1373. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Tang, Y.; Li, S.; Zhao, F.; Ying, J.; Qu, Y.; Niu, X.; Mu, D. Cumulative evidence of relationships between multiple variants in 8q24 region and cancer incidence. Medicine 2020, 99, e20716. [Google Scholar] [CrossRef] [PubMed]
- Shen, A.; Liu, L.; Huang, Y.; Shen, Z.; Wu, M.; Chen, X.; Wu, X.; Lin, X.; Chen, Y.; Li, L.; et al. Targeting Spindle-Associated Protein HAUS6 Prevents Cell Growth of Colorectal Cancer Cells by Activating the P53/P21 Pathway. Res. Square 2020. preprint. [Google Scholar]
- Lin, X.; Chen, Z.; Gao, P.; Gao, Z.; Chen, H.; Qi, J.; Liu, F.; Ye, D.; Jiang, H.; Na, R.; et al. TEX15: A DNA repair gene associated with prostate cancer risk in Han Chinese. Prostate 2017, 77, 1271–1278. [Google Scholar] [CrossRef]
- Han, Y.; Rand, K.A.; Hazelett, D.; Ingles, S.A.; Kittles, R.A.; Strom, S.S.; Rybicki, B.A.; Nemesure, B.; Isaacs, W.B.; Stanford, J.L.; et al. Prostate cancer susceptibility in men of African ancestry at 8q24. JNCI J. Natl. Cancer Inst. 2016, 108, djv431. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | SNP ID | Gene Consequence | Alleles | Ancestral | Placement | Clinical Impact | MAF | Frequency from the 1000 Genomes Project | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AFR | EAS | SAS | EUR | AMR | ||||||||
| OR2A5 | rs2961144 | Missense Variant | A/G | A | GRCh38.p13 chr 7 | Not Reported | 0.30 | 0.5318 | 0.0079 | 0.298 | 0.2903 | 0.269 |
| HAUS6 | rs1048169 | 3’ UTR Variant | T/C/G | C | GRCh38.p13 chr 9 | Not Reported | 0.42 | 0.7073 | 0.2073 | 0.36 | 0.3668 | 0.323 |
| CASC8 | rs7000448 | Intron Variant | C/T | T | GRCh38.p13 chr 8 | Not Reported | 0.41 | 0.711 | 0.2659 | 0.258 | 0.4046 | 0.29 |
| HNF1B | rs4430796 | Intron Variant | A/C/G/T | G | GRCh38.p13 chr 17 | Not Reported | 0.49 | 0.6906 | 0.2768 | 0.297 | 0.4751 | 0.395 |
| CDKN1B | rs2066827 | Missense Variant | T/A/C/G | T | GRCh38.p13 chr 12 | Benign | 0.36 | 0.7837 | 0.0556 | 0.319 | 0.2425 | 0.218 |
| PDL1M5 | rs12500426 | Intron Variant | A/C | C | GRCh38.p13 chr 4 | Not Reported | 0.48 | 0.6225 | 0.4931 | 0.241 | 0.4931 | 0.435 |
| CCAT2 | rs6983267 | Non-Coding Transcript Variant | G/T | G | GRCh38.p13 chr 8 | Not Reported | 0.39 | 0.041 | 0.0477 | 0.6121 | 0.468 | 0.5010 |
| HNF1B | rs11649743 | Intron Variant | A/G | G | GRCh38.p13 chr 17 | Not Reported | 0.37 | 0.9470 | 0.6617 | 0.885 | 0.8012 | 0.829 |
| EGFR | rs2075110 | Intron Variant | C/G/T | T | GRCh38.p13 chr 7 | Not Reported | 0.48 | 0.643 | 0.3522 | 0.565 | 0.565 | 0.514 |
| PCAT2 | rs114798100 | Intron Variant | A/G | A | GRCh38.p13 chr 8 | Not Reported | 0.01 | 0.0477 | 0 | 0 | 0 | 0.003 |
| WNT1 | rs855723 | Regulatory Region Variant | G/A/C/T | G | GRCh38.p13 chr 12 | Not Reported | 0.25 | 0.4365 | 0.996 | 0.737 | 0.8549 | 0.889 |
| EGFR | rs2075109 | Intron Variant | T/C | C | GRCh38.p13 chr 7 | Not Reported | 0.50 | 0.584 | 0.3522 | 0.564 | 0.4841 | 0.513 |
| Gene | SNP ID | Gene consequence | Alleles | Ancestral | Placement | Clinical Impact | MAF | Frequency from the 1000 Genomes Project | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AFR | EAS | SAS | EUR | AMR | ||||||||
| CASC8 | rs7000448 | Intron Variant | C/T | T | GRCh38.p13 chr 8 | Not Reported | 0.41 | 0.711 | 0.2659 | 0.258 | 0.4046 | 0.29 |
| HAUS6 | rs1048169 | 3 Prime UTR Variant | T/C/G | C | GRCh38.p13 chr 9 | Not Reported | 0.42 | 0.7073 | 0.2073 | 0.36 | 0.3668 | 0.323 |
| NF1B | rs4430796 | Intron Variant | A/C/G/T | G | GRCh38.p13 chr 17 | Not Reported | 0.49 | 0.6906 | 0.2768 | 0.297 | 0.4751 | 0.395 |
| OR2A5 | rs2961144 | Missense Variant | A/G | A | GRCh38.p13 chr 7 | Not Reported | 0.30 | 0.5318 | 0.0079 | 0.298 | 0.903 | 0.269 |
| PDL1M5 | rs12500426 | Intron Variant | A/C | C | GRCh38.p13 chr 4 | Not Reported | 0.48 | 0.6225 | 0.4931 | 0.241 | 0.4931 | 0.435 |
| CDKN1B | rs2066827 | Missense Variant | T/A/C/G | T | GRCh38.p13 chr 12 | Benign | 0.36 | 0.7837 | 0.0556 | 0.319 | 0.2425 | 0.218 |
| WNT1 | rs855723 | Regulatory Region Variant | G/A/C/T | G | GRCh38.p13 chr 12 | Not Reported | 0.25 | 0.4365 | 0.996 | 0.737 | 0.8549 | 0.889 |
| CCAT2 | rs6983267 | Non-Coding Transcript Exon Variant | G/T | G | GRCh38.p13 chr 8 | Not Reported | 0.39 | 0.0477 | 0.6121 | 0.468 | 0.5010 | 0.450 |
| PCAT2 | rs114798100 | Intron Variant | A/G | A | GRCh38.p13 chr 8 | Not Reported | 0.01 | 0.0477 | 0 | 0 | 0 | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, G.M.; Gellen, L.P.A.; da Veiga Borges Leal, D.F.; Pastana, L.F.; Vinagre, L.W.M.S.; Aquino, V.T.; Fernandes, M.R.; de Assumpção, P.P.; Burbano, R.M.R.; dos Santos, S.E.B.; et al. Correlation between Genomic Variants and Worldwide Epidemiology of Prostate Cancer. Genes 2022, 13, 1039. https://doi.org/10.3390/genes13061039
Vieira GM, Gellen LPA, da Veiga Borges Leal DF, Pastana LF, Vinagre LWMS, Aquino VT, Fernandes MR, de Assumpção PP, Burbano RMR, dos Santos SEB, et al. Correlation between Genomic Variants and Worldwide Epidemiology of Prostate Cancer. Genes. 2022; 13(6):1039. https://doi.org/10.3390/genes13061039
Chicago/Turabian StyleVieira, Giovana Miranda, Laura Patrícia Albarello Gellen, Diana Feio da Veiga Borges Leal, Lucas Favacho Pastana, Lui Wallacy Morikawa Souza Vinagre, Vitória Teixeira Aquino, Marianne Rodrigues Fernandes, Paulo Pimentel de Assumpção, Rommel Mario Rodríguez Burbano, Sidney Emanuel Batista dos Santos, and et al. 2022. "Correlation between Genomic Variants and Worldwide Epidemiology of Prostate Cancer" Genes 13, no. 6: 1039. https://doi.org/10.3390/genes13061039
APA StyleVieira, G. M., Gellen, L. P. A., da Veiga Borges Leal, D. F., Pastana, L. F., Vinagre, L. W. M. S., Aquino, V. T., Fernandes, M. R., de Assumpção, P. P., Burbano, R. M. R., dos Santos, S. E. B., & dos Santos, N. P. C. (2022). Correlation between Genomic Variants and Worldwide Epidemiology of Prostate Cancer. Genes, 13(6), 1039. https://doi.org/10.3390/genes13061039