
Repeatome Analyses and Satellite DNA Chromosome Patterns in Deschampsia sukatschewii, D. cespitosa, and D. antarctica (Poaceae)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Genomic DNA Extraction and Sequencing
2.3. Sequence Analysis and Identification of DNA Repeats
2.4. Chromosome Spread Preparation
2.5. Fluorescence In Situ Hybridization
2.6. Chromosome Analysis
3. Results
3.1. Comparative Analyses of the Repetitive DNA Sequences
3.2. BLAST Analysis
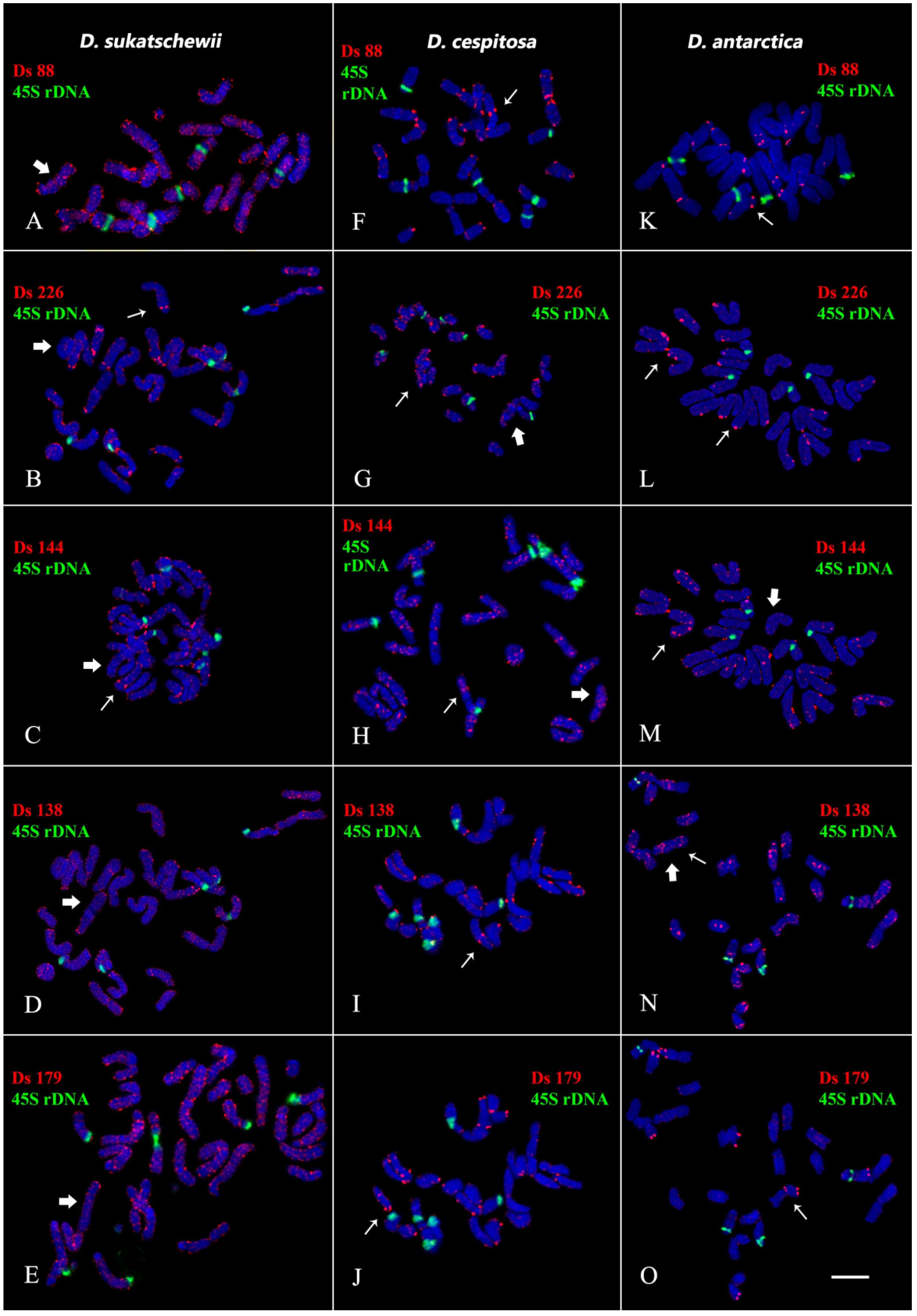
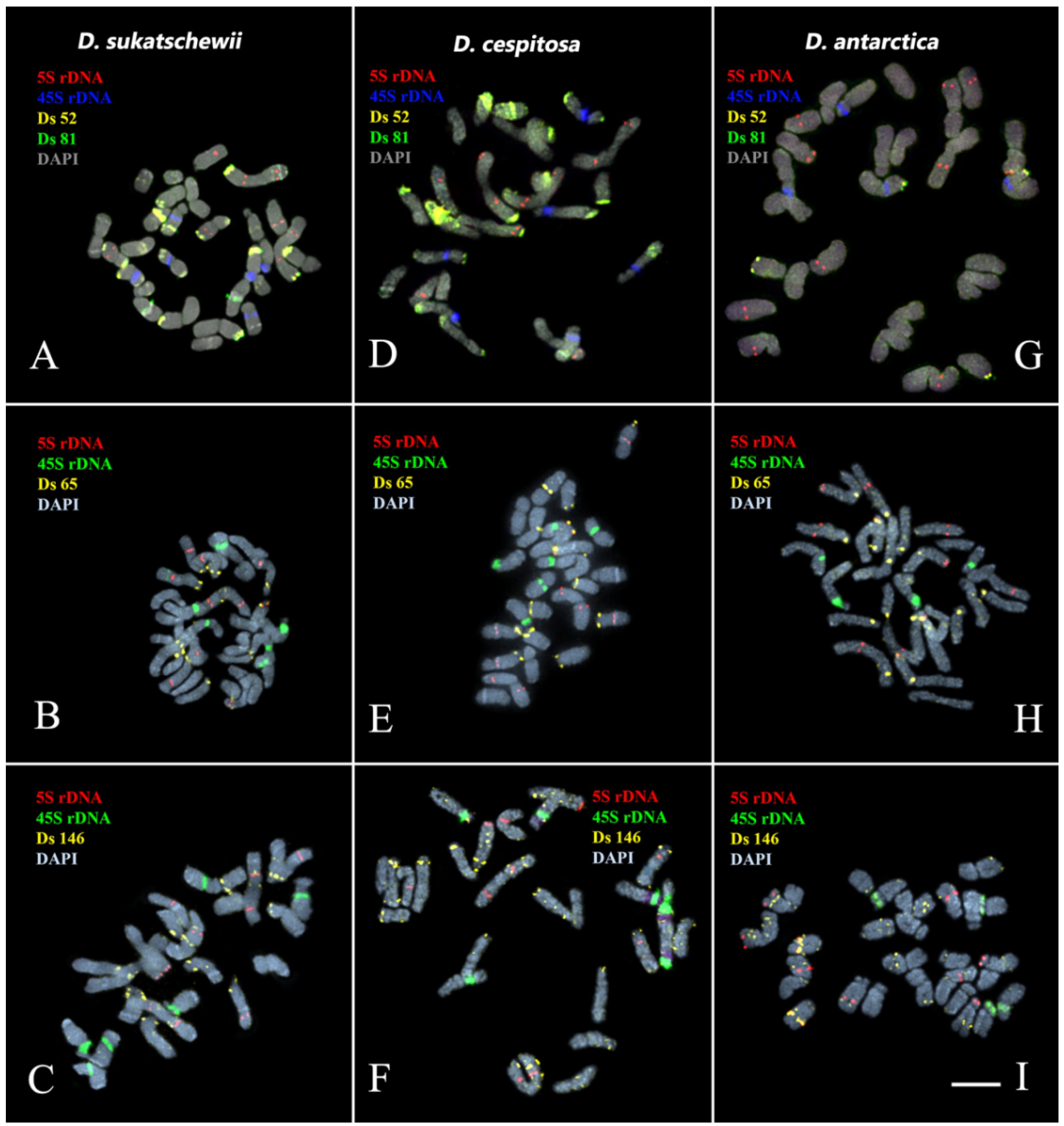
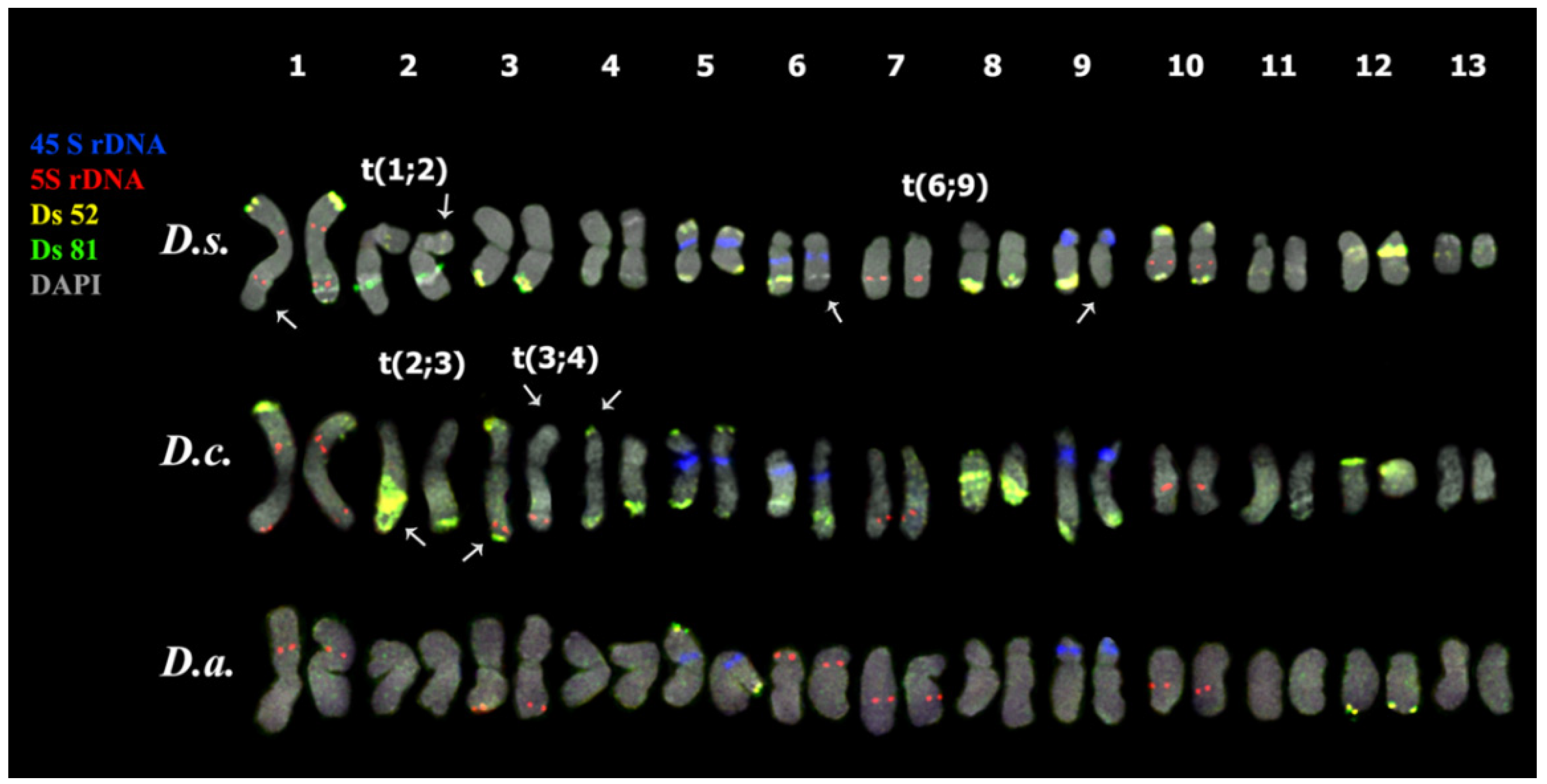
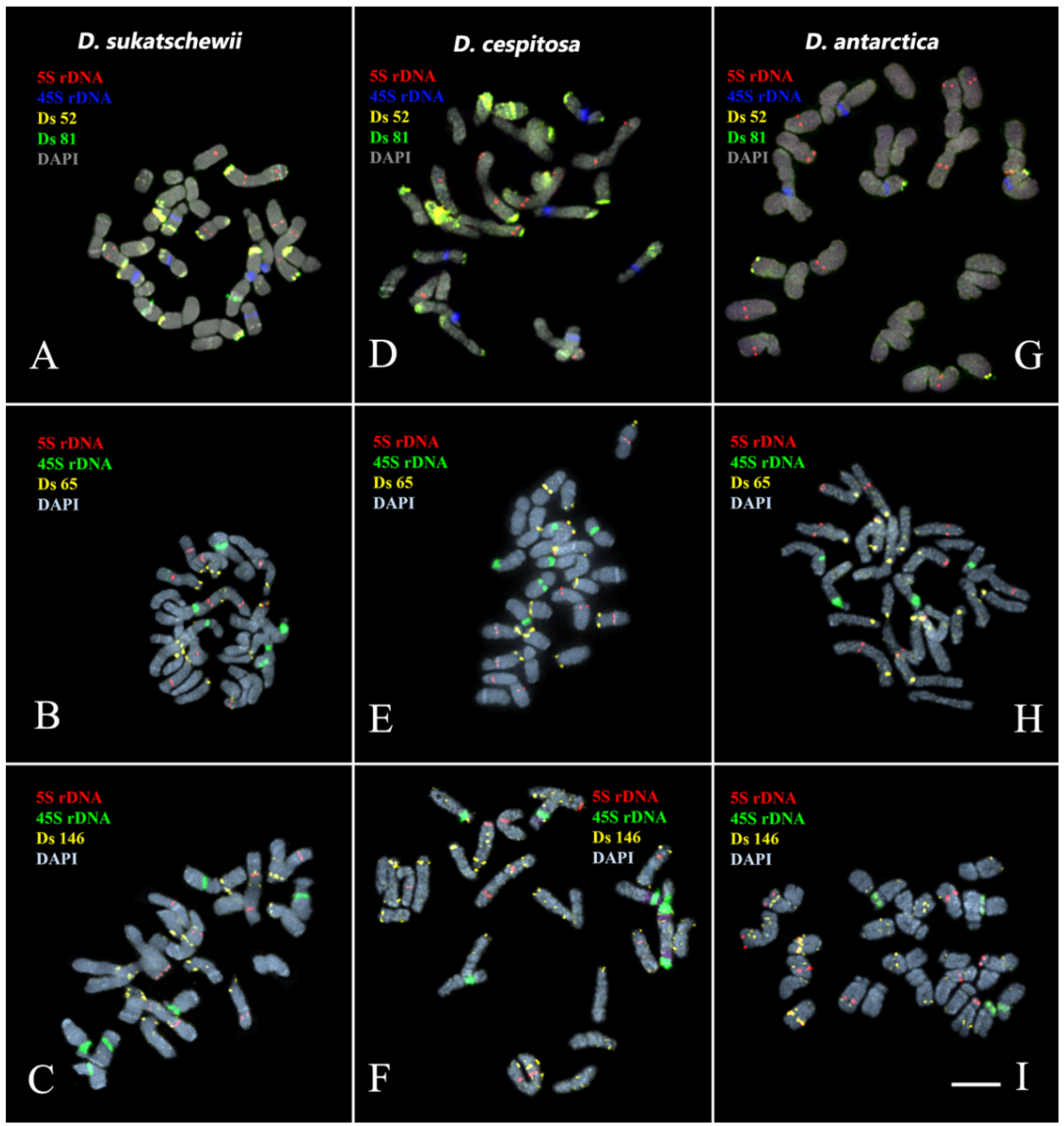
3.3. Chromosomal Structural Variations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tzvelev, N.N. Arctic Flora of the USSR; Nauka: Leningrad, USSR, 1964; Volume 2. [Google Scholar]
- Hulten, E. Flora of Alaska and Neighboring Territories; Stanford University Press: Stanford, UK, 1968. [Google Scholar]
- Alberdi, M.; Bravo, L.A.; Gutierrez, A.; Gidekel, M.; Corcuera, L.J. Ecophysiology of Antarctic vascular plants. Physiol. Plant 2002, 115, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Löve, A.; Löve, D. Chromosome numbers of central and northwest European plant species. In “Opera Botanica” a Societate Botanica Lundensi, 5; Almqvist & Wiksell: Stockholm, Sweden, 1961. [Google Scholar]
- Hulten, E.; Fries, M. Atlas of North European Vascular Plants. North of the Tropic of Cancer; Koeltz Scientific Books: Konigstein, Germany, 1986. [Google Scholar]
- Amosova, A.V.; Zoshchuk, S.A.; Rodionov, A.V.; Ghukasyan, L.; Samatadze, T.E.; Punina, E.O.; Loskutov, I.G.; Yurkevich, O.Y.; Muravenko, O.V. Molecular cytogenetics of valuable Arctic and sub-Arctic pasture grass species from the Aveneae/Poeae tribe complex (Poaceae). BMC Genet. 2019, 20, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, D.M. Chromosome numbers of Falkland Islands angiosperms. BAS Bull. 1967, 14, 69–82. [Google Scholar]
- Moore, D.M. Studies in Colobanthus quitensis (Kunth) Bartl. and Deschampsia antarctica Desv. II. Taxonomy, distribution and relationships. BAS Bull. 1970, 23, 63–80. [Google Scholar]
- Chiapella, J.; Zuloaga, F.O. A Revision of Deschampsia, Avenella, and Vahlodea (Poaceae, Poeae, Airinae) in South America. Ann. Mo. Bot. Gard. 2010, 97, 141–162. [Google Scholar] [CrossRef]
- Lee, J.; Noh, E.K.; Choi, H.S.; Shin, S.C.; Park, H.; Lee, H. Transcriptome sequencing of the Antarctic vascular plant Deschampsia antarctica Desv. under abiotic stress. Planta 2013, 237, 823–836. [Google Scholar] [CrossRef]
- Byun, M.Y.; Lee, J.; Cui, L.H.; Kang, Y.; Oh, T.K.; Park, H.; Lee, H.; Kim, W.T. Constitutive expression of DaCBF7, an Antarctic vascular plant Deschampsia antarctica CBF homolog, resulted in improved cold tolerance in transgenic rice plants. Plant Sci. 2015, 236, 61–74. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Akram, N.A.; Ashraf, M.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P. Plant responses to environmental stresses—From gene to biotechnology. AoB PLANTS 2017, 9, plx025. [Google Scholar] [CrossRef] [Green Version]
- Androsiuk, P.; Chwedorzewska, K.J.; Dulska, J.; Milarska, S.; Giełwanowska, I. Retrotransposon based genetic diversity of Deschampsia antarctica Desv. from King George Island (Maritime Antarctic). Ecol. Evol. 2021, 11, 648–663. [Google Scholar] [CrossRef]
- Cardone, S.; Sawatani, P.; Rush, P.; García, A.M.; Poggio, L.; Schrauf, G. Karyological studies in Deschampsia antarctica Desv. (Poaceae). Polar Biol. 2009, 32, 427–433. [Google Scholar] [CrossRef]
- Amosova, A.V.; Bolsheva, N.L.; Samatadze, T.E.; Twardovska, M.O.; Zoshchuk, S.A.; Andreev, I.O.; Badaeva, E.D.; Kunakh, V.A.; Muravenko, O.V. Molecular cytogenetic analysis of Deschampsia antarctica Desv. (Poaceae), Maritime Antarctic. PLoS ONE 2015, 10, e0138878. [Google Scholar] [CrossRef]
- Amosova, A.V.; Bolsheva, N.L.; Zoshchuk, S.A.; Twardovska, M.O.; Yurkevich, O.Y.; Andreev, I.O.; Samatadze, T.E.; Badaeva, E.D.; Kunakh, V.A.; Muravenko, O.V. Comparative molecular cytogenetic characterization of seven Deschampsia (Poaceae) species. PLoS ONE 2017, 12, e0175760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flavell, R.B.; O’Dell, M.; Hutchinson, J. Nucleotide sequence organization in plant chromosomes and evidence for sequence translocation during evolution. Cold Spring Harb. Symp. Quant. Biol. 1981, 45, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Flavell, R.B. Repetitive DNA and chromosome evolution in plants. Philos. Trans. R. Soc. B Biol. Sci. 1986, 312, 227–242. [Google Scholar]
- Schmidt, T.; Heslop-Harrison, J.S. Genomes, genes and junk: The large-scale organization of plant chromosomes. Trends Plant Sci. 1998, 3, 195–199. [Google Scholar] [CrossRef]
- Mehrotra, S.; Goyal, V. Repetitive sequences in plant nuclear DNA: Types, distribution, evolution and function. Genom. Proteom. Bioinform. 2014, 12, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Ramos, M.A. Satellite DNA: An Evolving Topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef]
- Grebenstein, B.; Grebenstein, O.; Sauer, W.; Hemleben, V. Characterization of a highly repeated DNA component of perennial oats (Helictotrichon, Poaceae) with sequence similarity to a A-genome-specific satellite DNA of rice (Oryza). Theor. Appl. Genet. 1995, 90, 1101–1105. [Google Scholar] [CrossRef] [PubMed]
- Winterfeld, G.; Röser, M. Chromosomal localization and evolution of satellite DNAs and heterochromatin in grasses (Poaceae), especially tribe Aveneae. Plant Syst. Evol. 2007, 264, 75–100. [Google Scholar] [CrossRef]
- Röser, M.; Winterfeld, G.; Doring, E.; Schneider, J. Chromosome evolution in grass tribes Aveneae/Poeae (Poaceae): In-sights from karyotype structure and molecular phylogeny. Schlechtendalia 2014, 28, 1–21. [Google Scholar]
- Amosova, A.; Ghukasyan, L.; Yurkevich, O.; Bolsheva, N.; Samatadze, T.; Zoshchuk, S.; Muravenko, O. Cytogenomics of Deschampsia P. Beauv (Poaceae) species based on sequence analyses and FISH mapping of CON/COM satellite DNA families. Plants 2021, 10, 1105. [Google Scholar] [CrossRef] [PubMed]
- Novák, P.; Neumann, P.; Macas, J. Graph-based clustering and characterization of repetitive sequences in next-generation sequencing data. BMC Bioinform. 2010, 11, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novák, P.; Neumann, P.; Pech, J.; Steinhaisl, J.; Macas, J. RepeatExplorer: A galaxybased web server for genome-wide characterization of eukaryotic repetitive elements from next-generation sequence reads. Bioinformatics 2013, 29, 792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novák, P.; Neumann, P.; Macas, J. Global analysis of repetitive DNA from unassembled sequence reads using RepeatExplorer2. Nat. Protoc. 2020, 15, 3745–3776. [Google Scholar] [CrossRef]
- Macas, J.; Kejnovský, E.; Neumann, P.; Novák, P.; Koblížková, A.; Vyskot, B. Next generation sequencing-based analysis of repetitive DNA in the model dioecious plant Silene latifolia. PLoS ONE 2011, 6, e27335. [Google Scholar] [CrossRef]
- Renny-Byfield, S.; Kovařík, A.; Chester, M.; Nichols, R.A.; Macas, J.; Novák, P.; Leitch, A.R. Independent, rapid and targeted loss of highly repetitive DNA in natural and synthetic allopolyploids of Nicotiana tabacum. PLoS ONE 2012, 7, e36963. [Google Scholar] [CrossRef] [Green Version]
- Křivánková, A.; Kopecký, D.; Stočes, Š.; Doležel, J.; Hřibová, E. Repetitive DNA: A versatile tool for karyotyping in Festuca pratensis Huds. Cytogenet. Genome Res. 2017, 151, 96–105. [Google Scholar] [CrossRef]
- Said, M.; Hřibová, E.; Danilova, T.V.; Karafiátová, M.; Čížková, J.; Friebe, B.; Doležel, J.; Gill, B.S.; Vrána, J. The Agropyron cristatum karyotype, chromosome structure and cross-genome homoeology as revealed by fluorescence in situ hybridization with tandem repeats and wheat single-gene probes. Theor. Appl. Genet. 2018, 131, 2213–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwyrtková, J.; Němečková, A.; Čížková, J.; Holušová, K.; Kapustová, V.; Svačina, R.; Kopecký, D.; Till, B.J.; Doležel, J.; Hřibová, E. Comparative analyses of DNA repeats and identification of a novel Fesreba centromeric element in fescues and ryegrasses. BMC Plant Biol. 2020, 20, 280. [Google Scholar] [CrossRef] [PubMed]
- González, M.L.; Chiapella, J.O.; Urdampilleta, J.D. Genomic differentiation of Deschampsia antarctica and D. cespitosa (Poaceae) based on satellite DNA. Bot. J. Linn. Soc. 2020, 194, 326–341. [Google Scholar] [CrossRef]
- González, M.L.; Chiapella, J.O.; Urdampilleta, J.D. Chromosomal differentiation of Deschampsia (Poaceae) based on four satellite DNA families. Front. Genet. 2021, 12, 728664. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.G.; De Lange, P.J.; Ferguson, A.R. Nuclear DNA variation, chromosome numbers and polyploidy in the endemic and indigenous grass flora of New Zealand. Ann. Bot. 2005, 96, 1293–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greimler, J.; Temsch, E.M.; Xue, Z.; Weiss-Schneeweiss, H.; Volkova, P.; Peintinger, M.; Wasowicz, P.; Shang, H.; Schanzer, I.; Chiapella, J.O. Genome size variation in Deschampsia cespitosa sensu lato (Poaceae) in Eurasia. Plant Syst. Evol. 2022, 308, 9. [Google Scholar] [CrossRef]
- Novak, P.; Robledillo, L.A.; Koblizkova, A.; Vrbova, I.; Neumann, P.; Macas, J. TAREAN: A computational tool for identification and characterization of satellite DNA from unassembled short reads. Nucleic Acid Res. 2017, 45, e111. [Google Scholar] [CrossRef]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, W.L.; Bedbrook, J.R. Cloning and characterization of ribosomal RNA genes from wheat and barley. Nucleic Acids Res. 1979, 7, 1869–1885. [Google Scholar] [CrossRef]
- Gerlach, W.L.; Dyer, T.A. Sequence organization of the repeating units in the nucleus of wheat which contain 5S rRNA genes. Nucleic Acids Res. 1980, 8, 4851–4855. [Google Scholar] [CrossRef] [Green Version]
- Muravenko, O.V.; Yurkevich, O.Y.; Bolsheva, N.L.; Samatadze, T.E.; Nosova, I.V.; Zelenina, D.A.; Volkov, A.; Popov, K.V.; Zelenin, A.V. Comparison of genomes of eight species of sections Linum and Adenolinum from the genus Linum based on chromosome banding, molecular markers and RAPD analysis. Genetica 2009, 135, 245–255. [Google Scholar] [CrossRef]
- Waring, M.; Britten, R.J. Nucleotide sequence repetition—A rapidly reassociating fraction of mouse DNA. Science 1966, 154, 791–794. [Google Scholar] [CrossRef]
- Britten, R.J.; Kohne, D.E. Repeated sequences in DNA. Hundreds of thousands of copies of DNA sequences have been incorporated into the genomes of higher organisms. Science 1968, 161, 529–540. [Google Scholar] [CrossRef]
- Bennetzen, J.L.; Wang, H. The contributions of transposable elements to the structure, function, and evolution of plant genomes. Annu. Rev. Plant Biol. 2014, 65, 505–530. [Google Scholar] [CrossRef] [PubMed]
- Meštrović, N.; Mravinac, B.; Pavlek, M.; Vojvoda-Zeljko, T.; Šatović, E.; Plohl, M. Structural and functional liaisons between transposable elements and satellite DNAs. Chromosome Res. 2015, 23, 583–596. [Google Scholar] [CrossRef] [PubMed]
- SanMiguel, P.; Tikhonov, A.; Jin, Y.-K.; Motchoulskaia, N.; Zakharov, D.; Melake-Berhan, A.; Springer, P.S.; Edwards, K.J.; Lee, M.; Avramova, Z.; et al. Nested retrotransposons in the inter-genic regions of the maize genome. Science 1996, 274, 765–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SanMiguel, P.; Bennetzen, J.L. Evidence that a recent increase in maize genome size was caused by the massive amplification of intergene retrotranposons. Ann. Bot. 1998, 82, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Makalowski, W. The human genome structure and organization. Acta Biochim. Pol. 2001, 48, 587–598. [Google Scholar] [CrossRef]
- Finnegan, D.J. Eukaryotic transposable elements and genome evolution. Trends Genet. 1989, 5, 103–107. [Google Scholar] [CrossRef]
- Makałowski, W.; Gotea, V.; Pande, A.; Makałowska, I. Transposable elements: Classification, identification, and their use as a tool for comparative genomics. In Evolutionary Genomics. Methods in Molecular Biology; Anisimova, M., Ed.; Humana: New York, NY, USA, 2019; Volume 1910, pp. 170–270. [Google Scholar] [CrossRef] [Green Version]
- Wicker, T.; Sabot, F.; Hua-Van, A.; Bennetzen, J.L.; Capy, P.; Chalhoub, B.; Flavell, A.; Leroy, P.; Morgante, M.; Panaud, O.; et al. A unified classification system for eukaryotic transposable elements. Nat. Rev. Genet. 2007, 8, 973–982. [Google Scholar] [CrossRef]
- Neumann, P.; Novák, P.; Hoštáková, N.; Macas, J. Systematic survey of plant LTR-retrotransposons elucidates phylogenetic relationships of their polyprotein domains and provides a reference for element classification. Mob. DNA 2019, 10, 1. [Google Scholar] [CrossRef]
- Vitte, C.; Panaud, O. LTR retrotransposons and flowering plant genome size: Emergence of the increase/decrease model. Cytogenet. Genome Res. 2005, 110, 91–107. [Google Scholar] [CrossRef]
- Baucom, R.; Estill, J.; Chaparro, C.; Upshaw, N.; Jogi, A.; Deragon, J.-M.; Westerman, R.P.; SanMiguel, P.J.; Bennetzen, J.L. Exceptional diversity, non-random distribution, and rapid evolution of retroelements in the B73 maize genome. PLoS Genet. 2009, 5, e1000732. [Google Scholar] [CrossRef]
- Macas, J.; Novák, P.; Pellicer, J.; Čížková, J.; Koblížková, A.; Neumann, P.; Fuková, I.; Doležel, J.; Kelly, L.J.; Leitch, I.J. In depth characterization of repetitive DNA in 23 plant genomes reveals sources of genome size variation in the vegume vribe Fabeae. PLoS ONE 2015, 10, e0143424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-J.; Gao, L.-I. Rapid and recent evolution of LTR retrotransposons drives rice genome evolution during the speciation of AA-genome Oryza species. G3 2017, 7, 1875–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCann, J.; Macas, J.; Novák, P.; Stuessy, T.F.; Villasenor, J.L.; Weiss-Schneweiss, H. Differential genome size and repetitive DNA evolution in diploid species of Melampodium sect Melampodium (Asteraceae). Front. Plant Sci. 2020, 11, 362. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zheng, Z.; Li, Y.; Hu, H.; Wang, Z.; Du, X. Which factors contribute most to genome size variation within angiosperms? Ecol. Evol. 2021, 11, 2660–2668. [Google Scholar] [CrossRef]
- Becher, H.; Powell, R.F.; Brown, M.R.; Metherell, C.; Pellicer, J.; Leitch, I.J.; Twyford, A.D. The nature of intraspecific and interspecific genome size variation in taxonomically complex eyebrights. Ann. Bot. 2021, 128, 639–651. [Google Scholar] [CrossRef]
- Kawano, S. Cytogeography and evolution of the Deschampsia caespitosa complex. Can. J. Bot. 1963, 41, 719–742. [Google Scholar] [CrossRef]
- Bennett, M.D.; Smith, J.B.; Lewis Smith, R.I. DNA amounts of angiosperms from the Antarctic and South Georgia. Environ. Exp. Bot. 1982, 22, 307–318. [Google Scholar] [CrossRef]
- Pascual-Díaz, J.P.; Serçe, S.; Hradecká, I.; Vanek, M.; Özdemir, B.S.; Sultana, N.; Vural, M.; Vitales, D.; Garcia, S. Genome size constancy in Antarctic populations of Colobanthus quitensis and Deschampsia antarctica. Polar Biol. 2020, 43, 1407–1413. [Google Scholar] [CrossRef]
- Liu, Q.; Li, X.; Zhou, X.; Li, M.; Zhang, F.; Schwarzacher, T.; Heslop-Harrison, J.S. The repetitive DNA landscape in Avena (Poaceae): Chromosome and genome evolution defined by major repeat classes in whole-genome sequence reads. BMC Plant Biol. 2019, 19, 226. [Google Scholar] [CrossRef] [Green Version]
- Stapley, J.; Santure, A.W.; Dennis, S.R. Transposable elements as agents of rapid adaptation may explain the genetic paradox of invasive species. Mol. Ecol. 2015, 24, 2241–2252. [Google Scholar] [CrossRef]
- Holderegger, R.; Stehlic, I.; Lewis, R.I.; Smith Abbott, R.J. Population of Antarctic hairgrass (Deschampsia antarctica) show low genetic diversity. Arct. Antarct. Alp. Res. 2003, 35, 214–217. [Google Scholar] [CrossRef]
- Chwedorzewska, K.J.; Bednarek, P.T. Genetic variability in the Antarctic hairgrass Deschampsia antarctica Desv from Maritime Antarctic and subantarctic sites. Pol. J. Ecol. 2008, 56, 209–216. [Google Scholar]
- Chwedorzewska, K.J.; Bednarek, P.T. Genetic and epigenetic studies on populations of Deschampsia antarctica Desv. from contrasting environments at King George Island (Antarctic). Pol. Polar Res. 2011, 32, 15–26. [Google Scholar] [CrossRef]
- Moreau-Mhiri, C.; Morel, J.B.; Audeon, C.; Ferault, M.; Grandbastien, M.A.; Lucas, H. Regulation of expression of the tobacco Tnt1 retrotransposon in heterologous species following pathogen-related stresses. Plant J. 1996, 9, 409–419. [Google Scholar] [CrossRef]
- Takeda, S.; Sugimoto, K.; Otsuki, H.; Hirochika, H. Transcriptional activation of the tobacco retrotransposon Tto1 by wounding and methyl jasmonate. Plant Mol. Biol. 1998, 36, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Voronova, A.; Jansons, Ā.; Ruņģis, D. Expression of retrotransposon-like sequences in Scots pine (Pinus sylvestris) in response to heat stress. Environ. Exp. Biol. 2011, 9, 121–127. [Google Scholar]
- Schrader, L.; Kim, J.W.; Ence, D.; Zimin, A.; Klein, A.; Wyschetzki, K.; Weichselgartner, T.; Kemena, C.; Stökl, J.; Schultner, E.; et al. Transposable element islands facilitate adaptation to novel environments in an invasive species. Nat. Commun. 2014, 5, 5495. [Google Scholar] [CrossRef] [Green Version]
- Cuadrado, A.; Cardoso, M.; Jouve, N. Physical organization of simple sequence repeats (SSRs) in Triticeae: Structural, functional and evolutionary implications. Cytogenet. Genome Res. 2008, 120, 210–219. [Google Scholar] [CrossRef]
- Dou, Q.; Liu, R.; Yu, F. Chromosomal organization of repetitive DNAs in Hordeum bogdanii and H. brevisubulatum (Poaceae). Comp. Cytogenet. 2016, 10, 465–481. [Google Scholar] [CrossRef] [Green Version]
- McStay, B. Nucleolar organizer regions: Genomic ‘dark matter’ requiring illumination. Genes Dev. 2016, 30, 1598–1610. [Google Scholar] [CrossRef] [Green Version]
- Kubis, S.; Schmidt, T.; Heslop-Harrison, J.S. Repetitive DNA elements as a major component of plant genomes. Ann. Bot. 1998, 82, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Hemleben, V.; Kovařík, A.; Torres-Ruiz, R.A.; Volkov, R.A.; Beridze, T. Plant highly repeated satellite DNA: Molecular evolution, distribution and use for identification of hybrids. Syst. Biodivers. 2007, 5, 277–289. [Google Scholar] [CrossRef]
- Macas, J.; Mészáros, T.; Nouzová, M. PlantSat: A Specialized Database for Plant Satellite Repeats. Bioinformatics 2002, 18, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, S.; Raina, S.N. Organization and evolution of highly repeated satellite DNA sequences in plant chromosomes. Cytogenet. Genome Res. 2005, 109, 15–26. [Google Scholar] [CrossRef]
- Plohl, M.; Meštrovic, N.; Mravinac, B. Satellite DNA evolution. In Repetitive DNA.; Garrido-Ramos, M.A., Ed.; Karger: Granada, Spain, 2012; pp. 126–152. [Google Scholar] [CrossRef]
- Plohl, M.; Petrović, V.; Luchetti, A.; Ricci, A.; Šatović, E.; Passamonti, M.; Mantovani, B. Long-term conservation vs high sequence divergence: The case of an extraordinarily old satellite DNA in bivalve mollusks. Heredity 2010, 104, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Ramos, M.A. Satellite DNA in Plants: More than Just Rubbish. Cytogenet. Genome Res. 2015, 146, 153–170. [Google Scholar] [CrossRef]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef] [Green Version]
- Lower, S.S.; McGurk, M.P.; Clark, A.G.; Barbash, D.A. Satellite DNA Evolution: Old Ideas, New Approaches. Curr. Opin. Genet. Dev. 2018, 49, 70–78. [Google Scholar] [CrossRef]
- Wei, L.; Liu, B.; Zhang, C.; Yu, Y.; Yang, X.; Dou, Q.; Dong, Q. Identification and characterization of satellite DNAs in Poa, L. Mol. Cytogenet. 2020, 13, 1–8. [Google Scholar] [CrossRef]
- Ugarkovic, D. Functional elements residing within satellite DNAs. EMBO Rep. 2005, 6, 1035–1039. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.S. Comparative Genome Organization in Plants: From Sequence and Markers to Chromatin and Chromosomes. Plant Cell 2000, 12, 617–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heslop-Harrison, J.S. Planning for remodelling: Nuclear architecture, chromatin and chromosomes. Trends Plant Sci. 2003, 8, 195–197. [Google Scholar] [CrossRef]
- Grebenstein, B.; Grebenstein, O.; Sauer, W.; Hemleben, V. Distribution and complex organization of satellite DNA sequences in Aveneae species. Genome 1996, 39, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- González, M.L.; Chiapella, J.O.; Urdampilleta, J.D. Characterization of some satellite DNA families in Deschampsia antarctica (Poaceae). Polar Biol. 2018, 41, 457–468. [Google Scholar] [CrossRef]
- Miklos, G.L.G.; Gill, A.C. Nucleotide sequences of highly repeated DNAs; compilation and comments. Genet. Res. 1982, 39, 1–30. [Google Scholar] [CrossRef]
- Badaeva, E.D.; Chikida, N.N.; Belousova, M.K.; Ruban, A.S.; Surzhikov, S.A.; Zoshchuk, S.A. A new insight on the evolution of polyploid Aegilops species from the complex Crassa: Molecular-cytogenetic analysis. Plant Syst. Evol. 2021, 307, 3. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tandem Repeat | Oligo FISH Probe Name/Length, bp | Oligo FISH Probe Sequence |
|---|---|---|
| Ds 52 | Ds 52_1/20 | AATTTGAACCCCTGGACCTC |
| Ds 52_2/20 | ACCCCTTTTATCCAAATGCC | |
| Ds 56 | Ds 56_1/20 | ACCAGCTCATTTCGGAACAC |
| Ds 56_2/20 | AATTCAGGTTCTACGTGCGG | |
| Ds 65 | Ds 65_1/21 | CTCCAAAACAAAGCTTTGGTG |
| Ds 65_1/20 | AAGGCTTGTCCATGGAATTG | |
| Ds 81 | Ds 81_1/20 | GCCTGACACCCTGACTTAGC |
| Ds 81_2/20 | GAAAAGATGCACTGATCGCA | |
| Ds 83 | Ds 83_1/20 | GCCAGAAGTATCCCAAACGA |
| Ds 83_2/20 | TAGTGTGTTATGGCCCACGA | |
| Ds 88 | Ds 88 _1/23 | CGGTTTCGAAGGCGTTAGAAAGA |
| Ds 88 _2/20 | ACTCGAAATTCGATGGAACG | |
| Ds 124 | Ds 124_1/20 | TGCAAAATTTCTTGACACCG |
| Ds 124_2/20 | GCGTGAAATTCCCACAGATT | |
| Ds 138 | Ds 138_1/20 | GTCTACCCCTTTGACCGGAT |
| Ds 138_2/20 | CCAATGAACGTTTTCCTTCC | |
| Ds 144 | Ds 144_1/20 | GGGGGTAGCTCAATGGAACT |
| Ds 144_2/20 | TTATGTTCATTTGTGTTTGT | |
| Ds 146 | Ds 146_1/20 | ATACCACCTTGTGAAAAGTA |
| Ds 146_2/20 | TCCCTTTCCTCATTGGATCA | |
| Ds 179 | Ds 179_1/23 | ATGGCACATGATGAAACGCGTTT |
| Ds 179_2/20 | TTTAATACGGGACTGGGCTG | |
| Ds 226 | Ds 226_1/20 | AGCATGGAAAACCAAGTTGG |
| Repeat Name | Genome Proportion (%) | ||
|---|---|---|---|
| D. antarctica | D. cespitosa | D. sukatschewii | |
| Retrotransposons (Class I) | 43.41 | 42.13 | 41.21 |
| Ty1-Copia | 19.21 | 15.27 | 15.77 |
| Unclassified Ty1-Copia elements | - | 0.17 | 0.01 |
| Ale | - | 0.02 | 0.02 |
| Angela | 10.28 | 6.84 | 7.49 |
| Ikeros | 0.51 | 0.34 | 0.47 |
| Ivana | - | 0.18 | 0.18 |
| SIRE | 8.25 | 7.54 | 7.38 |
| TAR | 0.17 | 0.18 | 0.22 |
| Ty3-Gypsy | 10.13 | 25.53 | 21.16 |
| non-chromovirus Athila | 0.27 | 2.85 | 3.83 |
| non-chromovirus Tat- Ogre | - | 0.22 | 0.13 |
| non-chromovirus Tat-Retand | 1.94 | 7.02 | 4.09 |
| chromovirus CRM | 0.56 | 0.23 | 0.22 |
| chromovirus Tekay | 7.36 | 15.21 | 12.89 |
| LINE | 0.03 | 0.02 | 0.01 |
| Unclassified LTR elements | 14.03 | 1.31 | 4.27 |
| Transposons (Class II) | 2.58 | 2.89 | 2.48 |
| Cacta | 2.44 | 2.84 | 2.34 |
| MuDR_Mutator | 0.13 | 0.01 | 0.12 |
| PIF_Harbinger | 0.01 | 0.04 | 0.02 |
| Ribosomal DNA | 0.06 | 0.6 | 0.29 |
| Unclassified repeats | 7.45 | 5.86 | 9.18 |
| Satellite DNA | 2.07 | 2.85 | 1.61 |
| Organelle | 4.81 | 1.04 | 2.73 |
| Repetitive DNA | 60.37 | 55.37 | 57.5 |
| Putative satellites | 13 high confident 7 low confident | 18 high confident 9 low confident | 12 high confident 9 low confident |
| SatDNA/Genome Proportion, %/Repeat Length, bp (Our Data) * | BLAST Homology (Available NCBI Data) | ||
|---|---|---|---|
| D. sukatschewii | D. cespitosa | D. antarctica | |
| Ds 52/0.33/184 | Dc 87/0.23/184 | not found | Poa pratensis clone PpTR-3 microsatellite sequence KY618841.1, Poa pratensis clone PpTR-2 microsatellite sequence KY618840.1, 78% of identity with Ds 52. |
| Ds 56/0.29/366 | Dc 38/0.48/366 | Da 272/0.025/366 | D. cespitosa satellite D12 sequence MT548102.1, D. antarctica clone 1 satellite D12 sequence MT548072.1, 99% of identity with Ds 56, H. convolutum satellite DNA (ID: pCON1_3). |
| Ds 65/0.22/314 | Dc 89/0.23/314 | not found | D. cespitosa satellite D31 sequence MT548119.1, D. antarctica satellite D31 sequence MT548089.1, 97–98% of identity with Ds 65. |
| Ds 81/0.14/369 | Dc 135/0.078/369 | not found | Leymus triticoides clone Lt1-4 satellite sequence EU629350.1, 83% of identity with Ds 81. |
| Ds 83/0.13/355 | Dc 125/0.1/355 | Da 197/0.076/355 | D. antarctica clone 2 satellite D17 sequence MT548144.1, D. cespitosa satellite D17 sequence MT548106.1, 70–98% of identity with Ds 83, H. compressum satellite DNA (ID pCOM2_4) Z68786.1. |
| Ds 88/0.11/379 | Dc 17/0.69 380 (75% of identity with Ds 88)/; Dc 77/0.31/379 | DA 238/0.042/379 | D. cespitosa satellite D10 sequence MT548100.1, D. antarctica satellite D4 sequence MT548064.1, 96–99% of identity with Ds 88, Secale cereale clone BAC 114I10 satellite pSc200 sequence KT724946.1, 83% of identity with Ds 88. |
| Ds 124/0.046/569 | Dc 234/0.016/563 | Da 351/0.013/563 | D. antarctica satellite D13 sequence MT548073.1, D. cespitosa satellite D13 sequence MT548103.1, 99–100% of identity with Ds 124, H. convolutum satellite DNA (ID pCON2_2). |
| Ds 138/0.036/158 | Dc 177/0.031/158 | Da 154/0.11/158 | D. cespitosa satellite D5 sequence MT548095.1, D. antarctica satellite D6 sequence MT548066.1, 100% of identity with Ds 138, D. antarctica clone 1 satellite D5 sequence MT548133.1, 94% identity with Ds 138, D. cespitosa satellite D29 sequence MT548117.1, D. antarctica satellite D29 sequence MT548087.1, D. antarctica satellite D20 sequence MT548081.1, D. cespitosa satellite D20 sequence MT548110.1, 73–77% of identity with Ds 138, Festuca pratensis satellite TR7 sequence. |
| Ds 142/0.035/658 | Dc 163/0.041/658 | Da 107/0.18/658 | D. antarctica satellite D5 sequence MT548065.1, D. antarctica clone 2 satellite D4 sequence MT548131.1, 99% of identity with Ds 142, Triticum aestivum cultivar Chinese Spring clone BAC 36I14, complete sequence. |
| Ds 144/0.034/352 | Dc 146/0.063/352 | Da 322/0.013 /342 (70% of identity with Ds 144) | D. antarctica clone 3 satellite D1 sequence MT548124.1, D. cespitosa satellite D7 sequence MT548097.1, both 72–73% of identity with Ds 144. |
| Ds 146/0.033/344 | not found | not found | not detected. |
| Ds 158/0.03/350 | Dc 238/0.016/358 | Da 116/0.17/351 | D. antarctica satellite D16 sequence MT548076.1, D. cespitosa satellite D16 sequence MT548105.1, 98–94% of identity with Ds 158, D. cespitosa satellite D22 sequence MT548111.1, D. antarctica satellite D22 sequence MT548083.1, 67% of identity with Ds 158. |
| Ds 166/0.027/174 | Dc 302/0.01/174 | not found | D. antarctica satellite D25 sequence MT548086.1, D. cespitosa satellite D25 sequence MT548113.1, 98–99% of identity with Ds 166. |
| Ds 179/0.023/318 | Dc 261/0.013/318 | not found | D. antarctica satellite D3 sequence MT548063.1, D. cespitosa satellite D3 sequence MT548094.1, 91–89% of identity with Ds 179, Setaria viridis cultivar ME034v chromosome 1. |
| Ds 182/0.022/343 | Dc 211/0.02/343 | Da 225/0.051/341 | D. cespitosa satellite D33 sequence MT548121.1, D. antarctica satellite D33 sequence MT548091.1, 70% of identity with Ds 182, D. cespitosa satellite D20 sequence MT548110.1, D. antarctica satellite D20 sequence MT548081.1, 74% of identity with Ds 182, D. cespitosa satellite D5 sequence MT548095.1, D. antarctica satellite D6 sequence MT548066.1, 70% of identity with Ds 182, H. pratense satellite DNA (ID pPRA1_2). |
| Ds 211/0.015/171 | Dc 174/0.034/171 | Da 204/0.067/171 | D. cespitosa satellite D6 sequence MT548096.1, D. antarctica satellite D7 sequence MT548067.1, D. antarctica clone 1 satellite D6 sequence MT548134.1, 98% of identity with Ds 211. |
| Ds 226/0.014/345 | Dc 106/0.16/ 352 (67% of identity with Ds 226) | Da 97/0.21 /342; Da 129/0.14/343 | D. cespitosa satellite D1 sequence MT548092.1, D. antarctica satellite D1 sequence MT548061.1, 75% of identity with Ds 226, D. cespitosa satellite D20 sequence MT548110.1, D. cespitosa satellite D5 sequence MT548095.1, D. antarctica satellite D6 sequence MT548066.1, 70% of identity with Ds 226, Festuca pratensis satellite TR4 sequence. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amosova, A.V.; Yurkevich, O.Y.; Bolsheva, N.L.; Samatadze, T.E.; Zoshchuk, S.A.; Muravenko, O.V. Repeatome Analyses and Satellite DNA Chromosome Patterns in Deschampsia sukatschewii, D. cespitosa, and D. antarctica (Poaceae). Genes 2022, 13, 762. https://doi.org/10.3390/genes13050762
Amosova AV, Yurkevich OY, Bolsheva NL, Samatadze TE, Zoshchuk SA, Muravenko OV. Repeatome Analyses and Satellite DNA Chromosome Patterns in Deschampsia sukatschewii, D. cespitosa, and D. antarctica (Poaceae). Genes. 2022; 13(5):762. https://doi.org/10.3390/genes13050762
Chicago/Turabian StyleAmosova, Alexandra V., Olga Yu. Yurkevich, Nadezhda L. Bolsheva, Tatiana E. Samatadze, Svyatoslav A. Zoshchuk, and Olga V. Muravenko. 2022. "Repeatome Analyses and Satellite DNA Chromosome Patterns in Deschampsia sukatschewii, D. cespitosa, and D. antarctica (Poaceae)" Genes 13, no. 5: 762. https://doi.org/10.3390/genes13050762
APA StyleAmosova, A. V., Yurkevich, O. Y., Bolsheva, N. L., Samatadze, T. E., Zoshchuk, S. A., & Muravenko, O. V. (2022). Repeatome Analyses and Satellite DNA Chromosome Patterns in Deschampsia sukatschewii, D. cespitosa, and D. antarctica (Poaceae). Genes, 13(5), 762. https://doi.org/10.3390/genes13050762