
Chromosome Changes in Soma and Germ Line: Heritability and Evolutionary Outcome


{kind=link}
Abstract
1. Introduction
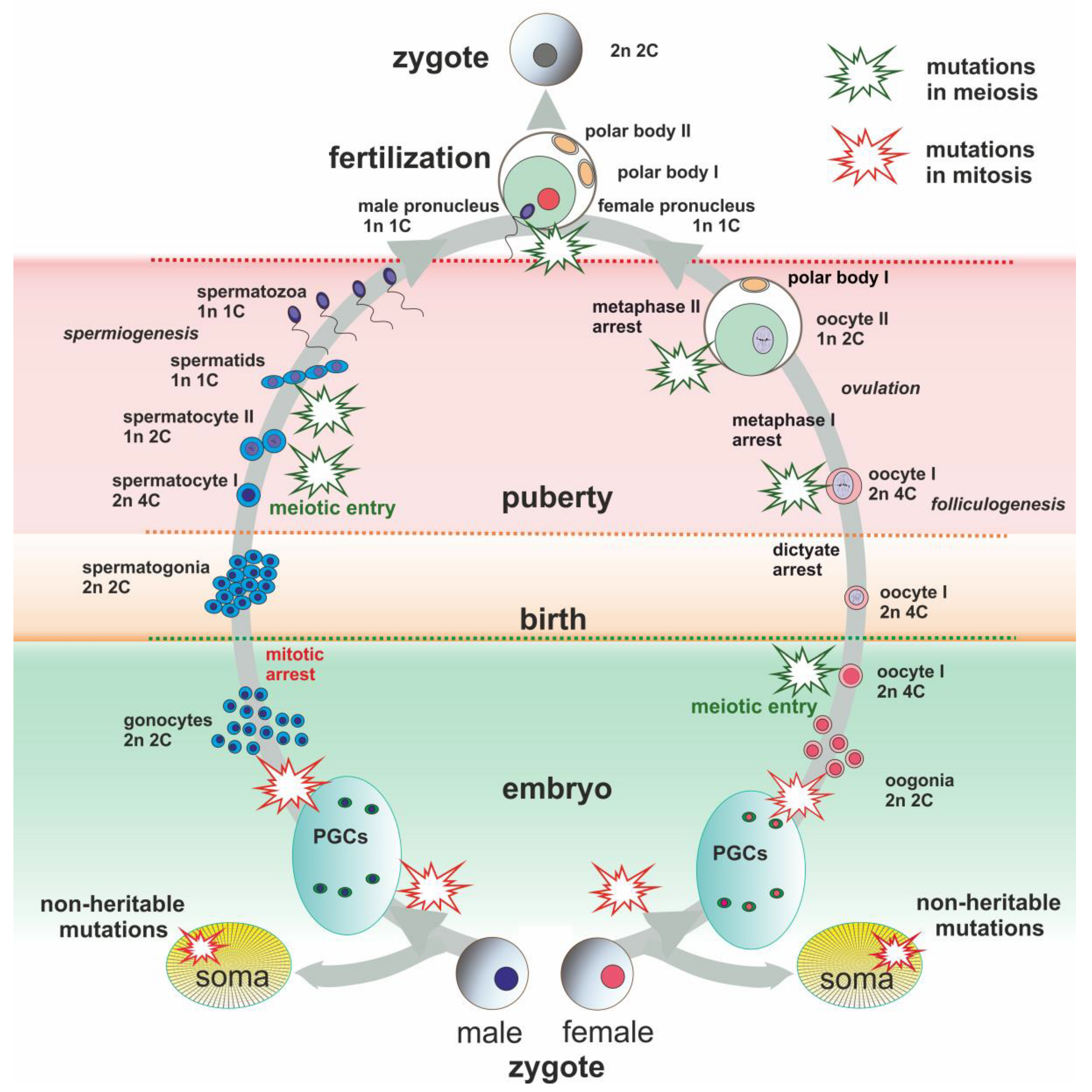
2. Chromosome Rearrangements in Early Development
2.1. Start of a New Life
2.2. Early Development, Alterations, and Mosaicism
3. Gametogenesis
3.1. Mitotic and Meiotic Checkpoints
3.2. Chromothripsis in Germ Line
3.3. Female Meiotic Drive as an Evolutionary Force
4. Chromosomal Speciation
4.1. Robertsonian Translocations and WARTs
4.2. Looking for a Model of Robertsonian Translocations Origin
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nägeli, C. Zur Entwickelungsgeschichte Des Pollens Bei Den Phanerogamen; Orell, Füssli und Comp: Zürich, Switzerland, 1842. [Google Scholar]
- Sutton, W.S. On the morphology of the chromosome group in Brachystola magna. Biol. Bull. 1902, 4, 24–39. [Google Scholar] [CrossRef]
- Boveri, T. Ergebnisse über Die Konstitution Der Chromatischen Substanz Des Zellkerns; Fischer G.: Jena, Germany, 1904. [Google Scholar]
- Ju, Y.S.; Martincorena, I.; Gerstung, M.; Petljak, M.; Alexandrov, L.B.; Rahbari, R.; Wedge, D.C.; Davies, H.R.; Ramakrishna, M.; Fullam, A.; et al. Somatic mutations reveal asymmetric cellular dynamics in the early human embryo. Nature 2017, 543, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Munisha, M.; Schimenti, J.C. Genome maintenance during embryogenesis. DNA Repair 2021, 106, 103195. [Google Scholar] [CrossRef] [PubMed]
- Stephens, P.J.; Greenman, C.D.; Fu, B.; Yang, F.; Bignell, G.R.; Mudie, L.J.; Pleasance, E.D.; Lau, K.W.; Beare, D.; Stebbings, L.A.; et al. Massive genomic rearrangement acquired in a single catastrophic event during cancer development. Cell 2011, 144, 27–40. [Google Scholar] [CrossRef]
- Liu, P.; Erez, A.; Nagamani, S.C.S.; Dhar, S.U.; Kołodziejska, K.E.; Dharmadhikari, A.V.; Cooper, M.L.; Wiszniewska, J.; Zhang, F.; Withers, M.A.; et al. Chromosome catastrophes involve replication mechanisms generating complex genomic rearrangements. Cell 2011, 146, 889–903. [Google Scholar] [CrossRef]
- Baca, S.C.; Prandi, D.; Lawrence, M.S.; Mosquera, J.M.; Romanel, A.; Drier, Y.; Park, K.; Kitabayashi, N.; MacDonald, T.Y.; Ghandi, M.; et al. Punctuated evolution of prostate cancer genomes. Cell 2013, 153, 666–677. [Google Scholar] [CrossRef]
- Pellestor, F.; Gaillard, J.B.; Schneider, A.; Puechberty, J.; Gatinois, V. Chromoanagenesis, the mechanisms of a genomic chaos. Semin. Cell Dev. Biol. 2022, 123, 90–99. [Google Scholar] [CrossRef]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef]
- Zhao, B.; Rothenberg, E.; Ramsden, D.A.; Lieber, M.R. The molecular basis and disease relevance of non-homologous DNA end joining. Nat. Rev. Mol. Cell Biol. 2020, 21, 765–781. [Google Scholar] [CrossRef]
- Chen, S.; Lee, L.; Naila, T.; Fishbain, S.; Wang, A.; Tomkinson, A.E.; Lees-Miller, S.P.; He, Y. Structural basis of long-range to short-range synaptic transition in NHEJ. Nature 2021, 593, 294–298. [Google Scholar] [CrossRef]
- Kim, S.; Peterson, S.E.; Jasin, M.; Keeney, S. Mechanisms of germ line genome instability. Semin. Cell Dev. Biol. 2016, 54, 177–187. [Google Scholar] [CrossRef]
- Hattori, A.; Fukami, M. Established and novel mechanisms leading to de novo genomic rearrangements in the human germline. Cytogenet. Genome Res. 2020, 160, 167–176. [Google Scholar] [CrossRef]
- Kot, P.; Yasuhara, T.; Shibata, A.; Hirakawa, M.; Abe, Y.; Yamauchi, M.; Matsuda, N. Mechanism of chromosome rearrangement arising from single-strand breaks. Biochem. Biophys. Res. Commun. 2021, 572, 191–196. [Google Scholar] [CrossRef]
- Vyatkin, A.D.; Otnyukov, D.V.; Leonov, S.V.; Belikov, A.V. Comprehensive patient-level classification and quantification of driver events in TCGA PanCanAtlas cohorts. PLoS Genet. 2022, 18, e1009996. [Google Scholar] [CrossRef]
- Zhang, C.Z.; Pellman, D. Cancer genomic rearrangements and copy number alterations from errors in cell division. Annu. Rev. Cancer Biol. 2022, 6. [Google Scholar] [CrossRef]
- Takei, Y.; Yun, J.; Zheng, S.; Ollikainen, N.; Pierson, N.; White, J.; Shah, S.; Thomassie, J.; Suo, S.; Eng, C.H.; et al. Integrated spatial genomics reveals global architecture of single nuclei. Nature 2021, 590, 344–350. [Google Scholar] [CrossRef]
- Tortora, M.M.; Salari, H.; Jost, D. Chromosome dynamics during interphase: A biophysical perspective. Curr. Opin. Genet. Dev. 2020, 61, 37–43. [Google Scholar] [CrossRef]
- Xie, L.; Liu, Z. Single-cell imaging of genome organization and dynamics. Mol. Syst. Biol. 2021, 17, e9653. [Google Scholar] [CrossRef]
- Cavalli, G. Understanding 3D genome organization by multidisciplinary methods. Nat. Rev. Mol. Cell Biol. 2021, 22, 511–528. [Google Scholar] [CrossRef]
- Tanabe, H.; Müller, S.; Neusser, M.; von Hase, J.; Calcagno, E.; Cremer, M.; Solovei, I.; Cremer, C.; Cremer, T. Evolutionary conservation of chromosome territory arrangements in cell nuclei from higher primates. Proc. Natl. Acad. Sci. USA 2002, 99, 4424–4429. [Google Scholar] [CrossRef]
- Guerrero, R.F.; Kirkpatrick, M. Local adaptation and the evolution of chromosome fusions. Evolution 2014, 68, 2747–2756. [Google Scholar] [CrossRef] [PubMed]
- Matveevsky, S.; Tretiakov, A.; Kashintsova, A.; Bakloushinskaya, I.; Kolomiets, O. Meiotic nuclear architecture in distinct mole vole hybrids with Robertsonian translocations: Chromosome chains, stretched centromeres, and distorted recombination. Int. J. Mol. Sci. 2020, 21, 7630. [Google Scholar] [CrossRef] [PubMed]
- Winick-Ng, W.; Kukalev, A.; Harabula, I.; Zea-Redondo, L.; Szabó, D.; Meijer, M.; Serebreni, L.; Zhang, Y.; Bianco, S.; Chiariello, A.M.; et al. Cell-type specialization is encoded by specific chromatin topologies. Nature 2021, 599, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Salari, H.; Di Stefano, M.; Jost, D. Spatial organization of chromosomes leads to heterogeneous chromatin motion and drives the liquid-or gel-like dynamical behavior of chromatin. Genome Res. 2022, 32, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Kragesteen, B.K.; Spielmann, M.; Paliou, C.; Heinrich, V.; Schöpflin, R.; Esposito, A.; Annunziatella, C.; Bianco, S.; Chiariello, A.M.; Jerković, I.; et al. Dynamic 3D chromatin architecture contributes to enhancer specificity and limb morphogenesis. Nat. Genet. 2018, 50, 1463–1473. [Google Scholar] [CrossRef]
- Bhat, P.; Honson, D.; Guttman, M. Nuclear compartmentalization as a mechanism of quantitative control of gene expression. Nat. Rev. Mol. Cell Biol. 2021, 22, 653–670. [Google Scholar] [CrossRef]
- Glaser, J.; Mundlos, S. 3D or Not 3D: Shaping the Genome during Development. Cold Spring Harb. Perspect. Biol. 2021, a040188. [Google Scholar] [CrossRef]
- Lupiáñez, D.G.; Kraft, K.; Heinrich, V.; Krawitz, P.; Brancati, F.; Klopocki, E.; Horn, D.; Kayserili, H.; Opitz, J.M.; Laxova, R.; et al. Disruptions of topological chromatin domains cause pathogenic rewiring of gene-enhancer interactions. Cell 2015, 161, 1012–1025. [Google Scholar] [CrossRef]
- Schneider, I.; Ellenberg, J. Mysteries in embryonic development: How can errors arise so frequently at the beginning of mammalian life? PLoS Biol. 2019, 17, e3000173. [Google Scholar] [CrossRef]
- Shukla, V.; Høffding, M.K.; Hoffmann, E.R. Genome diversity and instability in human germ cells and preimplantation embryos. Semin. Cell Dev. Biol. 2021, 113, 132–147. [Google Scholar] [CrossRef]
- Reichmann, J.; Nijmeijer, B.; Hossain, M.J.; Eguren, M.; Schneider, I.; Politi, A.Z.; Roberti, M.J.; Hufnagel, L.; Hiiragi, T.; Ellenberg, J. Dual-spindle formation in zygotes keeps parental genomes apart in early mammalian embryos. Science 2018, 361, 189–193. [Google Scholar] [CrossRef]
- Cavazza, T.; Takeda, Y.; Politi, A.Z.; Aushev, M.; Aldag, P.; Baker, C.; Choudhary, M.; Bucevičius, J.; Lukinavičius, G.; Elder, K.; et al. Parental genome unification is highly error-prone in mammalian embryos. Cell 2021, 184, 2860–2877. [Google Scholar] [CrossRef]
- So, C.; Menelaou, K.; Uraji, J.; Harasimov, K.; Steyer, A.M.; Seres, K.B.; Bucevičius, J.; Lukinavičius, G.; Möbius, W.; Sibold, C.; et al. Mechanism of spindle pole organization and instability in human oocytes. Science 2022, 375, 6581. [Google Scholar] [CrossRef]
- Ing-Simmons, E.; Rigau, M.; Vaquerizas, J.M. Emerging mechanisms and dynamics of three-dimensional genome organisation at zygotic genome activation. Curr. Opin. Cell Biol. 2022, 74, 37–46. [Google Scholar] [CrossRef]
- Ghosh, R.P.; Meyer, B.J. Spatial organization of chromatin: Emergence of chromatin structure during development. Annu. Rev. Cell Dev. Biol. 2021, 37, 199–232. [Google Scholar] [CrossRef]
- Dai, X.; Guo, X. Decoding and rejuvenating human ageing genomes: Lessons from mosaic chromosomal alterations. Ageing Res. Rev. 2021, 68, 101342. [Google Scholar] [CrossRef]
- Campbell, I.M.; Shaw, C.A.; Stankiewicz, P.; Lupski, J.R. Somatic mosaicism: Implications for disease and transmission genetics. Trends Genet. 2015, 31, 382–392. [Google Scholar] [CrossRef]
- Guttenbach, M.; Koschorz, B.; Bernthaler, U.; Grimm, T.; Schmid, M. Sex chromosome loss and aging: In situ hybridization studies on human interphase nuclei. Am. J. Hum. Genet. 1995, 57, 1143. [Google Scholar]
- Russell, L.M.; Strike, P.; Browne, C.E.; Jacobs, P.A. X chromosome loss and ageing. Cytogenet. Genome Res. 2007, 116, 181–185. [Google Scholar] [CrossRef]
- Machiela, M.J.; Zhou, W.; Karlins, E.; Sampson, J.N.; Freedman, N.D.; Yang, Q.; Hicks, B.; Dagnall, C.; Hautman, C.; Jacobs, K.B.; et al. Female chromosome X mosaicism is age-related and preferentially affects the inactivated X chromosome. Nat. Commun. 2016, 7, 11843. [Google Scholar] [CrossRef]
- Ouseph, M.M.; Hasserjian, R.P.; Dal Cin, P.; Lovitch, S.B.; Steensma, D.P.; Nardi, V.; Weinberg, O.K. Genomic alterations in patients with somatic loss of the Y chromosome as the sole cytogenetic finding in bone marrow cells. Haematologica 2021, 106, 555. [Google Scholar] [CrossRef]
- Tuck-Muller, C.M.; Chen, H.; Martínez, J.E.; Shen, C.C.; Li, S.; Kusyk, C.; Batista, D.A.; Bhatnagar, Y.M.; Dowling, E.; Wertelecki, W. Isodicentric Y chromosome: Cytogenetic, molecular and clinical studies and review of the literature. Hum. Genet. 1995, 96, 119–129. [Google Scholar] [CrossRef]
- Chandley, A.C.; Ambros, P.; McBeath, S.; Hargreave, T.B.; Kilanowski, F.; Spowart, G. Short arm dicentric Y chromosome with associated statural defects in a sterile man. Hum. Genet. 1986, 73, 350–353. [Google Scholar] [CrossRef]
- Iannuzzi, A.; Parma, P.; Iannuzzi, L. Chromosome abnormalities and fertility in domestic bovids: A review. Animals 2021, 11, 802. [Google Scholar] [CrossRef]
- Biesecker, L.G.; Spinner, N.B. A genomic view of mosaicism and human disease. Nat. Rev. Genet. 2013, 14, 307–320. [Google Scholar] [CrossRef]
- McLaren, A. Primordial germ cells in the mouse. Dev. Biol. 2003, 262, 1–15. [Google Scholar] [CrossRef]
- Kobayashi, T.; Surani, M.A. On the origin of the human germline. Development 2018, 145, dev150433. [Google Scholar] [CrossRef]
- Zhao, J.; Lu, P.; Wan, C.; Huang, Y.; Cui, M.; Yang, X.; Hu, Y.; Zheng, Y.; Dong, J.; Wang, M.; et al. Cell-fate transition and determination analysis of mouse male germ cells throughout development. Nat. Commun. 2021, 12, 6839. [Google Scholar] [CrossRef]
- Nicholls, P.K.; Schorle, H.; Naqvi, S.; Hu, Y.C.; Fan, Y.; Carmell, M.A.; Dobrinski, I.; Watson, A.L.; Carlson, D.F.; Fahrenkrug, S.C.; et al. Mammalian germ cells are determined after PGC colonization of the nascent gonad. Proc. Natl. Acad. Sci. USA 2019, 116, 25677–25687. [Google Scholar] [CrossRef]
- Hancock, G.V.; Wamaitha, S.E.; Peretz, L.; Clark, A.T. Mammalian primordial germ cell specification. Development 2021, 148, dev189217. [Google Scholar] [CrossRef]
- Yang, X.; Breuss, M.W.; Xu, X.; Antaki, D.; James, K.N.; Stanley, V.; Ball, L.L.; George, R.D.; Wirth, S.A.; Cao, B.; et al. Developmental and temporal characteristics of clonal sperm mosaicism. Cell 2021, 184, 4772–4783. [Google Scholar] [CrossRef] [PubMed]
- Kruger, A.N.; Mueller, J.L. Mechanisms of meiotic drive in symmetric and asymmetric meiosis. Cell. Mol. Life Sci. 2021, 78, 3205–3218. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.N.; Johnson, R.T. Mammalian cell fusion: Studies on the regulation of DNA synthesis and mitosis. Nature 1970, 225, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Elledge, S.J. Cell cycle checkpoints: Preventing an identity crisis. Science 1996, 274, 1664–1672. [Google Scholar] [CrossRef] [PubMed]
- Elkouby, Y.M.; Mullins, M.C. Coordination of cellular differentiation, polarity, mitosis and meiosis—New findings from early vertebrate oogenesis. Dev. Biol. 2017, 430, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Musacchio, A.; Salmon, E.D. The spindle-assembly checkpoint in space and time. Nat. Rev. Mol. Cell Biol. 2007, 8, 379–393. [Google Scholar] [CrossRef]
- Vleugel, M.; Hoogendoorn, E.; Snel, B.; Kops, G.J. Evolution and function of the mitotic checkpoint. Dev. Cell 2012, 23, 239–250. [Google Scholar] [CrossRef]
- Musacchio, A. The molecular biology of spindle assembly checkpoint signaling dynamics. Curr. Biol. 2015, 2, R1002–R1018. [Google Scholar] [CrossRef]
- Santaguida, S.; Amon, A. Short- and long-term effects of chromosome mis-segregation and aneuploidy. Nat. Rev. Mol. Cell Biol. 2015, 16, 473–485. [Google Scholar] [CrossRef]
- Cohen-Sharir, Y.; McFarland, J.M.; Abdusamad, M.; Marquis, C.; Bernhard, S.V.; Kazachkova, M.; Tang, H.; Ippolito, M.R.; Laue, K.; Zerbib, J.; et al. Aneuploidy renders cancer cells vulnerable to mitotic checkpoint inhibition. Nature 2021, 590, 486–491. [Google Scholar] [CrossRef]
- Silk, A.D.; Zasadil, L.M.; Holland, A.J.; Vitre, B.; Cleveland, D.W.; Weaver, B.A. Chromosome missegregation rate predicts whether aneuploidy will promote or suppress tumors. Proc. Natl. Acad. Sci. USA 2013, 110, E4134–E4141. [Google Scholar] [CrossRef]
- Lukow, D.A.; Sheltzer, J.M. Chromosomal instability and aneuploidy as causes of cancer drug resistance. Trends Cancer 2022, 8, 43–53. [Google Scholar] [CrossRef]
- Abraham, R.T. Cell cycle checkpoint signaling through the ATM and ATR kinases. Genes Dev. 2001, 15, 2177–2196. [Google Scholar] [CrossRef]
- Panagopoulos, A.; Altmeyer, M. The hammer and the dance of cell cycle control. Trends Biochem. Sci. 2021, 46, 301–314. [Google Scholar] [CrossRef]
- Burgoyne, P.S.; Mahadevaiah, S.K.; Turner, J.M. The consequences of asynapsis for mammalian meiosis. Nat. Rev. Genet. 2009, 10, 207–216. [Google Scholar] [CrossRef]
- Rodriguez-Rodriguez, J.A.; Lewis, C.; McKinley, K.L.; Sikirzhytski, V.; Corona, J.; Maciejowski, J.; Khodjakov, A.; Cheeseman, I.M.; Jallepalli, P.V. Distinct roles of RZZ and Bub1–KNL1 in mitotic checkpoint signaling and kinetochore expansion. Curr. Biol. 2018, 28, 3422–3429. [Google Scholar] [CrossRef]
- Ichijima, Y.; Ichijima, M.; Lou, Z.; Nussenzweig, A.; Camerini-Otero, R.D.; Chen, J.; Andreassen, P.R.; Namekawa, S.H. MDC1 directs chromosome-wide silencing of the sex chromosomes in male germ cells. Genes Dev. 2011, 25, 959–971. [Google Scholar] [CrossRef]
- Aylon, Y.; Liefshitz, B.; Kupiec, M. The CDK regulates repair of double-strand breaks by homologous recombination during the cell cycle. EMBO J. 2004, 23, 4868–4875. [Google Scholar] [CrossRef]
- Matveevsky, S.; Chassovnikarova, T.; Grishaeva, T.; Atsaeva, M.; Malygin, V.; Bakloushinskaya, I.; Kolomiets, O. Kinase CDK2 in mammalian meiotic prophase I: Screening for hetero-and homomorphic sex chromosomes. Int. J. Mol. Sci. 2021, 22, 1969. [Google Scholar] [CrossRef]
- MacQueen, A.J.; Hochwagen, A. Checkpoint mechanisms: The puppet masters of meiotic prophase. Trends Cell Biol. 2011, 21, 393–400. [Google Scholar] [CrossRef]
- Subramanian, V.V.; Hochwagen, A. The meiotic checkpoint network: Step-by-step through meiotic prophase. Cold Spring Harb. Perspect. Biol. 2014, 6, a016675. [Google Scholar] [CrossRef]
- Hua, R.; Liu, M. Sexual dimorphism in mouse meiosis. Front. Cell Dev. Biol. 2021, 9, 670599. [Google Scholar] [CrossRef]
- Ishiguro, K.I. Sexually dimorphic properties in meiotic chromosome. Sex. Dev. 2022, 1–11. [Google Scholar] [CrossRef]
- Hu, X.; Shen, B.; Liao, S.; Ning, Y.; Ma, L.; Chen, J.; Lin, X.; Zhang, D.; Li, Z.; Zheng, C.; et al. Gene knockout of Zmym3 in mice arrests spermatogenesis at meiotic metaphase with defects in spindle assembly checkpoint. Cell Death Dis. 2017, 8, e2910. [Google Scholar] [CrossRef]
- Lane, S.; Kauppi, L. Meiotic spindle assembly checkpoint and aneuploidy in males versus females. Cell. Mol. Life Sci. 2019, 76, 1135–1150. [Google Scholar] [CrossRef]
- Rinaldi, V.D.; Bolcun-Filas, E.; Kogo, H.; Kurahashi, H.; Schimenti, J.C. The DNA damage checkpoint eliminates mouse oocytes with chromosome synapsis failure. Mol. Cell 2017, 67, 1026–1036. [Google Scholar] [CrossRef]
- Kouznetsova, A.; Wang, H.; Bellani, M.; Camerini-Otero, R.D.; Jessberger, R.; Hoog, C. BRCA1-mediated chromatin silencing is limited to oocytes with a small number of asynapsed chromosomes. J. Cell Sci. 2009, 122, 2446–2452. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pepling, M.E. Regulation of meiotic prophase one in mammalian oocytes. Front. Cell Dev. Biol. 2021, 9, 667306. [Google Scholar] [CrossRef] [PubMed]
- Stringer, J.M.; Winship, A.; Zerafa, N.; Wakefield, M.; Hutt, K. Oocytes can efficiently repair DNA double-strand breaks to restore genetic integrity and protect offspring health. Proc. Natl. Acad. Sci. USA 2020, 117, 11513–11522. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Y.; Feng, X.; Xie, F.Y.; Li, S.; Chen, L.N.; Luo, S.M.; Yin, S.; Ou, X.H. Double-strand breaks induce short-scale DNA replication and damage amplification in the fully grown mouse oocytes. Genetics 2021, 218, iyab054. [Google Scholar] [CrossRef]
- Kloosterman, W.P.; Guryev, V.; van Roosmalen, M.; Duran, K.J.; de Bruijn, E.; Bakker, S.C.; Letteboer, T.; van Nesselrooij, B.; Hochstenbach, R.; Poot, M.; et al. Chromothripsis as a mechanism driving complex de novo structural rearrangements in the germline. Hum. Mol. Genet. 2011, 20, 1916–1924. [Google Scholar] [CrossRef]
- Pellestor, F.; Gatinois, V.; Puechberty, J.; Geneviève, D.; Lefort, G. Chromothripsis: Potential origin in gametogenesis and preimplantation cell divisions. A review. Fertil Steril. 2014, 102, 1785–1796. [Google Scholar] [CrossRef]
- Hurst, L.D.; Ellegren, H. Sex biases in the mutation rate. Trends Genet. 1998, 14, 446–452. [Google Scholar] [CrossRef]
- Eisfeldt, J.; Pettersson, M.; Petri, A.; Nilsson, D.; Feuk, L.; Lindstrand, A. Hybrid sequencing resolves two germline ultra-complex chromosomal rearrangements consisting of 137 breakpoint junctions in a single carrier. Hum. Genet. 2021, 140, 775–790. [Google Scholar] [CrossRef]
- Capilla, L.; Sánchez-Guillén, R.A.; Farre, M.; Paytuví-Gallart, A.; Malinverni, R.; Ventura, J.; Larkin, D.M.; Ruiz-Herrera, A. Mammalian comparative genomics reveals genetic and epigenetic features associated with genome reshuffling in Rodentia. Genome Biol. Evol. 2016, 8, 3703–3717. [Google Scholar] [CrossRef][Green Version]
- Romanenko, S.A.; Perelman, P.L.; Trifonov, V.A.; Graphodatsky, A.S. Chromosomal evolution in Rodentia. Heredity 2012, 108, 4–16. [Google Scholar] [CrossRef]
- Sandler, L.; Novitski, E. Meiotic drive as an evolutionary force. Am. Nat. 1957, 91, 105–110. [Google Scholar] [CrossRef]
- Lindholm, A.K.; Dyer, K.A.; Firman, R.C.; Fishman, L.; Forstmeier, W.; Holman, L.; Johannesson, H.; Knief, U.; Kokko, H.; Larracuente, A.M.; et al. The ecology and evolutionary dynamics of meiotic drive. Trends Ecol. Evol. 2016, 31, 315–326. [Google Scholar] [CrossRef]
- de Villena, F.; Sapienza, C. Transmission ratio distortion in offspring of heterozygous female carriers of Robertsonian translocations. Hum. Genet. 2001, 108, 31–36. [Google Scholar] [CrossRef]
- de Villena, F.P.M.; Sapienza, C. Female meiosis drives karyotypic evolution in mammals. Genetics 2001, 159, 1179–1189. [Google Scholar] [CrossRef]
- Blackmon, H.; Justison, J.; Mayrose, I.; Goldberg, E.E. Meiotic drive shapes rates of karyotype evolution in mammals. Evolution 2019, 73, 511–523. [Google Scholar] [CrossRef]
- Tambovtseva, V.G.; Matveevsky, S.N.; Kashintsova, A.A.; Tretiakov, A.V.; Kolomiets, O.L.; Bakloushinskaya, I.Y. A meiotic mystery in experimental hybrids of the eastern mole vole (Ellobius tancrei, Mammalia, Rodentia). Vavilov J. Genet. Breed. 2019, 23, 239–243. [Google Scholar] [CrossRef]
- Wurster, D.H.; Benirschke, K. Indian muntjac, Muntiacus muntjak: A deer with a low diploid chromosome number. Science 1970, 168, 1364–1366. [Google Scholar] [CrossRef]
- Gallardo, M.H.; Bickham, J.W.; Honeycutt, R.L.; Ojeda, R.A.; Köhler, N. Discovery of tetraploidy in a mammal. Nature 1999, 401, 341. [Google Scholar] [CrossRef]
- White, M.J.D. Modes of Speciation; Freeman: San Francisco, CA, USA, 1978. [Google Scholar]
- King, M. Species Evolution: The Role of Chromosome Change; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Capanna, E. Robertsonian numerical variation in animal speciation: Mus musculus an emblematic model. In Mechanism of Speciation; Barigozzi, C., Ed.; Alan Liss: New York, NY, USA, 1982; pp. 155–177. [Google Scholar]
- Darwin, C. On the Origin of Species by Natural Selection; Murray: London, UK, 1859. [Google Scholar]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1937. [Google Scholar]
- Mayr, E.; Provine, W.B. The Evolutionary Synthesis; Harvard University Press: Cambridge, MA, USA, 1980. [Google Scholar]
- Gould, S.J.; Eldrige, N. Punctuated equilibria: The tempo and mode of evolution reconsidered. Paleobiology 1977, 3, 115–151. [Google Scholar] [CrossRef]
- Heasley, L.R.; Sampaio, N.; Argueso, J.L. Systemic and rapid restructuring of the genome: A new perspective on punctuated equilibrium. Curr. Genet. 2021, 67, 57–63. [Google Scholar] [CrossRef]
- Heng, J.; Heng, H.H. Genome chaos: Creating new genomic information essential for cancer macroevolution. Semin. Cancer Biol. 2020. [Google Scholar] [CrossRef]
- Pellestor, F.; Gatinois, V. Chromoanagenesis: A piece of the macroevolution scenario. Mol. Cytogenet. 2020, 13, 3. [Google Scholar] [CrossRef]
- Mayr, E. The Growth of Biological Thought: Diversity, Evolution, and Inheritance; Harvard University Press: Cambridge, MA, USA, 1982. [Google Scholar]
- Goldschmidt, R. The Material Basis of Evolution; Yale University Press: New Haven, CT, USA, 1940. [Google Scholar]
- Shapiro, J.A. What can evolutionary biology learn from cancer biology? Prog. Biophys. Mol. Biol. 2021, 165, 19–28. [Google Scholar] [CrossRef]
- Pellestor, F. Chromothripsis and the Macroevolution Theory. Methods Mol. Biol. 2018, 1769, 43–49. [Google Scholar] [CrossRef]
- Okasha, S. Cancer and the levels of selection. Br. J. Philos. Sci. 2022, in press. [Google Scholar] [CrossRef]
- Pearse, A.M.; Swift, K. Transmission of devil facial-tumour disease. Nature 2006, 439, 549. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A. Revisiting the central dogma in the 21st century. Ann. N. Y. Acad. Sci. 2009, 1178, 6–28. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A. Rethinking the (im)possible in evolution. Progr. Biophys. Mol. Biol. 2013, 111, 92–96. [Google Scholar] [CrossRef]
- Darlington, C.D. The Evolution of Genetic Systems; Cambridge University Press: Cambridge, UK, 1939. [Google Scholar]
- Deakin, J.E.; Potter, S.; O’Neill, R.; Ruiz-Herrera, A.; Cioffi, M.B.; Eldridge, M.D.; Fukui, K.; Marshall Graves, J.A.; Griffin, D.; Grutzner, F.; et al. Chromosomics: Bridging the gap between genomes and chromosomes. Genes 2019, 10, 627. [Google Scholar] [CrossRef]
- Franchini, P.; Kautt, A.F.; Nater, A.; Antonini, G.; Castiglia, R.; Meyer, A.; Solano, E. Reconstructing the evolutionary history of chromosomal races on islands: A genome-wide analysis of natural house mouse populations. Mol. Biol. Evol. 2020, 37, 2825–2837. [Google Scholar] [CrossRef]
- Jackson, C.E.; Xu, S.; Ye, Z.; Pfrender, M.E.; Lynch, M.; Colbourne, J.K.; Shaw, J.R. Chromosomal rearrangements preserve adaptive divergence in ecological speciation. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wallace, B. On coadaptation in Drosophila. Am. Nat. 1953, 87, 343–358. [Google Scholar] [CrossRef]
- Oliver-Bonet, M.; Benet, J.; Sun, F.; Navarro, J.; Abad, C.; Liehr, T.; Starke, H.; Greene, C.; Ko, E.; Martin, R.H. Meiotic studies in two human reciprocal translocations and their association with spermatogenic failure. Hum. Reprod. 2005, 20, 683–688. [Google Scholar] [CrossRef]
- Liu, P.; Carvalho, C.M.; Hastings, P.J.; Lupski, J.R. Mechanisms for recurrent and complex human genomic rearrangements. Curr. Opin. Genet. Dev. 2012, 22, 211–220. [Google Scholar] [CrossRef]
- Turner, D.J.; Miretti, M.; Rajan, D.; Fiegler, H.; Carter, N.P.; Blayney, M.L.; Beck, S.; Hurles, M.E. Germline rates of de novo meiotic deletions and duplications causing several genomic disorders. Nat. Genet. 2008, 40, 90–95. [Google Scholar] [CrossRef]
- Zhang, S.; Lei, C.; Wu, J.; Sun, H.; Zhou, J.; Zhu, S.; Wu, J.; Fu, J.; Sun, Y.; Lu, D.; et al. Analysis of segregation patterns of quadrivalent structures and the effect on genome stability during meiosis in reciprocal translocation carriers. Hum. Reprod. 2018, 33, 757–767. [Google Scholar] [CrossRef]
- Chandley, A.C.; McBeath, S.; Speed, R.M.; Yorston, L.; Hargreave, T.B. Pericentric inversion in human chromosome 1 and the risk for male sterility. J. Med. Genet. 1987, 24, 325–334. [Google Scholar] [CrossRef]
- León-Ortiz, A.M.; Panier, S.; Sarek, G.; Vannier, J.B.; Patel, H.; Campbell, P.J.; Boulton, S.J. A distinct class of genome rearrangements driven by heterologous recombination. Mol. Cell 2018, 69, 292–305. [Google Scholar] [CrossRef]
- Vara, C.; Paytuví-Gallart, A.; Cuartero, Y.; Álvarez-González, L.; Marín-Gual, L.; Garcia, F.; Florit-Sabater, B.; Capilla, L.; Sanchéz-Guillén, R.A.; Sarrate, Z.; et al. The impact of chromosomal fusions on 3D genome folding and recombination in the germ line. Nat. Commun. 2021, 12, 2981. [Google Scholar] [CrossRef]
- Monroe, J.; Srikant, T.; Carbonell-Bejerano, P.; Becker, C.; Lensink, M.; Exposito-Alonso, M.; Klein, M.; Hildebrandt, J.; Neumann, M.; Kliebenstein, D.; et al. Mutation bias reflects natural selection in Arabidopsis thaliana. Nature 2022, 602, 101–105. [Google Scholar] [CrossRef]
- Simakov, O.; Bredeson, J.; Berkoff, K.; Marletaz, F.; Mitros, T.; Schultz, D.T.; O’Connell, B.L.; Dear, P.; Martinez, D.E.; Steele, R.E.; et al. Deeply conserved synteny and the evolution of metazoan chromosomes. Sci. Adv. 2022, 8, eabi5884. [Google Scholar] [CrossRef]
- Hochstenbach, R.; van Binsbergen, E.; Engelen, J.; Nieuwint, A.; Polstra, A.; Poddighe, P.; Ruivenkamp, C.; Sikkema-Raddatz, B.; Smeets, D.; Poot, M. Array analysis and karyotyping: Workflow consequences based on a retrospective study of 36,325 patients with idiopathic developmental delay in the Netherlands. Eur. J. Med. Genet. 2009, 52, 161–169. [Google Scholar] [CrossRef]
- Gustavsson, I. Distribution and effects of the 1/29 Robertsonian translocation in cattle. J. Dairy Sci. 1964, 62, 825–835. [Google Scholar] [CrossRef]
- Iannuzzi, L.; King, W.A.; Di Berardino, D. Chromosome evolution in domestic bovids as revealed by chromosome banding and FISH-mapping techniques. Cytogenet. Genome Res. 2009, 126, 49–62. [Google Scholar] [CrossRef]
- Jiménez, J.M.; Morales, R.; Molina, A.; Moreno-Millán, M.; Demyda Peyrás, S. The effect of the Robertsonian translocation 1/29 on the fertility of beef cattle reared under extensive conditions: A 30 years retrospective study. Reprod. Domest. Anim. 2021. [Google Scholar] [CrossRef]
- Gropp, A.; Winking, H. Robertsonian translocations: Cytology, meiosis, segregation patterns and biological consequences of heterozygosity. Symp. Zool. Soc. Lond. 1981, 47, 141–181. [Google Scholar]
- Bogdanov, Y.F.; Kolomiets, O.L.; Lyapunova, E.A.; Yanina, I.Y.; Mazurova, T.F. Synaptonemal complexes and chromosome chains in the rodent Ellobius talpinus heterozygous for ten Robertsonian translocations. Chromosoma 1986, 94, 94–102. [Google Scholar] [CrossRef]
- Ratomponirina, C.; Andrianivo, J.; Rumpler, Y. Spermatogenesis in several intra- andinterspecific hybrids of the lemur (Lemur). J. Reprod. Fertil. 1982, 66, 717–721. [Google Scholar] [CrossRef] [PubMed]
- Volleth, M.; Heller, K.G.; Yong, H.S.; Müller, S. Karyotype evolution in the horseshoe bat Rhinolophus sedulus by whole-arm reciprocal translocation (WART). Cytogenet. Genome Res. 2014, 143, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Nunes, A.C.; Catalan, J.; Lopez, J.; Ramalhinho, M.G.; Mathias, M.L.; Britton-Davidian, J. Fertility assessment in hybrids between monobrachially homologous Rb races of the house mouse from the island of Madeira: Implications for modes of chromosomal evolution. Heredity 2011, 106, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Britton-Davidian, J.; Robinson, T.J.; Veyrunes, F. Systematics and evolution of the African pygmy mice, subgenus Nannomys: A review. Acta Oecologica 2012, 42, 41–49. [Google Scholar] [CrossRef]
- Tapisso, J.T.; Gabriel, S.I.; Cerveira, A.M.; Britton-Davidian, J.; Ganem, G.; Searle, J.B.; Ramalhinho, M.D.G.; Mathias, M.D.L. Spatial and temporal dynamics of contact zones between chromosomal races of house mice, Mus musculus domesticus, on Madeira Island. Genes 2020, 11, 748. [Google Scholar] [CrossRef]
- Potter, S.; Bragg, J.G.; Blom, M.P.; Deakin, J.E.; Kirkpatrick, M.; Eldridge, M.D.; Moritz, C. Chromosomal speciation in the genomics era: Disentangling phylogenetic evolution of rock-wallabies. Front. Genet. 2017, 8, 10. [Google Scholar] [CrossRef]
- Potter, S.; Bragg, J.G.; Turakulov, R.; Eldridge, M.D.; Deakin, J.; Kirkpatrick, M.; Edwards, R.J.; Moritz, C. Limited introgression between rock-wallabies with extensive chromosomal rearrangements. Mol. Biol. Evol. 2022, 39, msab333. [Google Scholar] [CrossRef]
- Pavlova, S.V.; Kolomiets, O.L.; Bulatova, N.S.; Searle, J.B. Demonstration of a WART in a hybrid zone of the common shrew (Sorex araneus Linnaeus, 1758). Comp. Cytogenet. 2008, 2, 115–120. [Google Scholar]
- Romanenko, S.A.; Lyapunova, E.A.; Saidov, A.S.; O’Brien, P.; Serdyukova, N.A.; Ferguson-Smith, M.A.; Graphodatsky, A.S.; Bakloushinskaya, I. Chromosome translocations as a driver of diversification in mole voles Ellobius (Rodentia, Mammalia). Int. J. Mol. Sci. 2019, 20, 4466. [Google Scholar] [CrossRef]
- Giménez, M.D.; Förster, D.W.; Jones, E.P.; Jóhannesdóttir, F.; Gabriel, S.I.; Panithanarak, T.; Scascitelli, M.; Merico, V.; Garagna, S.; Searle, J.B.; et al. A half-century of studies on a chromosomal hybrid zone of the house mouse. J. Hered. 2017, 108, 25–35. [Google Scholar] [CrossRef]
- Kartavtseva, I.V.; Sheremetyeva, I.N.; Pavlenko, M.V. Intraspecies multiple chromosomal variations including rare tandem fusion in the Russian Far Eastern endemic evoron vole Alexandromys evoronensis (Rodentia, Arvicolinae). Comp. Cytogenet. 2021, 15, 393. [Google Scholar] [CrossRef]
- Bazykin, A.D. Hypothetical mechanism of speciation. Evolution 1969, 23, 685–687. [Google Scholar] [CrossRef]
- Branco, M.R.; Pombo, A. Intermingling of chromosome territories in interphase suggests role in translocations and transcription-dependent associations. PLoS Biol. 2006, 4, e138. [Google Scholar] [CrossRef]
- Miura, H.; Hiratani, I. Cell cycle dynamics and developmental dynamics of the 3D genome: Toward linking the two timescales. Curr. Opin. Genet. Dev. 2022, 73, 101898. [Google Scholar] [CrossRef]
- Slijepcevic, P. Telomeres and mechanisms of Robertsonian fusion. Chromosoma 1998, 107, 136–140. [Google Scholar] [CrossRef]
- Garagna, S.; Broccoli, D.; Redi, C.A.; Searle, J.B.; Cooke, H.J.; Capanna, E. Robertsonian metacentrics of the mouse lose telomeric sequences but retain some minor satellite DNA in the pericentromeric area. Chromosoma 1995, 103, 685–692. [Google Scholar] [CrossRef]
- Nanda, I.; Schneider-Rasp, S.; Winking, H.; Schmid, M. Loss of telomeric sites in the chromosomes of Mus musculus domesticus (Rodentia: Muridae) during Robertsonian rearrangements. Chromosom. Res. 1995, 3, 399–409. [Google Scholar] [CrossRef]
- Clark, F.E.; Akera, T. Unravelling the mystery of female meiotic drive: Where we are. Open Biol. 2021, 11, 210074. [Google Scholar] [CrossRef] [PubMed]
- Henikoff, S.; Ahmad, K.; Malik, H.S. The centromere paradox: Stable inheritance with rapidly evolving DNA. Science 2001, 293, 1098–1102. [Google Scholar] [CrossRef] [PubMed]
- Kursel, L.E.; Malik, H.S. The cellular mechanisms and consequences of centromere drive. Curr. Opin. Cell Biol. 2018, 52, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Kumon, T.; Ma, J.; Akins, R.B.; Stefanik, D.; Nordgren, C.E.; Kim, J.; Levine, M.T.; Lampson, M.A. Parallel pathways for recruiting effector proteins determine centromere drive and suppression. Cell 2021, 184, 4904–4918. [Google Scholar] [CrossRef] [PubMed]
- Akera, T.; Chmatal, L.; Trimm, E.; Yang, K.; Aonbangkhen, C.; Chenoweth, D.M.; Janke, C.; Schultz, R.M.; Lampson, M.A. Spindle asymmetry drives non-Mendelian chromosome segregation. Science 2017, 358, 668–672. [Google Scholar] [CrossRef] [PubMed]
- Farré, M.; Robinson, T.J.; Ruiz-Herrera, A. An Integrative Breakage Model of genome architecture, reshuffling and evolution: The Integrative Breakage Model of genome evolution, a novel multidisciplinary hypothesis for the study of genome plasticity. BioEssays 2015, 37, 479–488. [Google Scholar] [CrossRef]
- Matveevsky, S.; Kolomiets, O.; Bogdanov, A.; Alpeeva, E.; Bakloushinskaya, I. Meiotic chromosome contacts as a plausible prelude for Robertsonian translocations. Genes 2020, 11, 386. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.D.; de Borja, R.; Young, M.D.; Fuligni, F.; Rosic, A.; Roberts, N.D.; Hajjar, S.; Layeghifard, M.; Novokmet, A.; Kowalski, P.E.; et al. Rearrangement bursts generate canonical gene fusions in bone and soft tissue tumors. Science 2018, 361, eaam8419. [Google Scholar] [CrossRef]
- Hattori, A.; Okamura, K.; Terada, Y.; Tanaka, R.; Katoh-Fukui, Y.; Matsubara, Y.; Matsubara, K.; Kagami, M.; Horikawa, R.; Fukami, M. Transient multifocal genomic crisis creating chromothriptic and non-chromothriptic rearrangements in prezygotic testicular germ cells. BMC Med. Genom. 2019, 12, 77. [Google Scholar] [CrossRef]
- Ghevaria, H.; SenGupta, S.; Naja, R.; Odia, R.; Exeter, H.; Serhal, P.; Gonzalez, X.V.; Sun, X.; Delhanty, J. Next generation sequencing detects premeiotic errors in human oocytes. Int. J. Mol. Sci. 2022, 23, 665. [Google Scholar] [CrossRef]
- Bergero, R.; Ellis, P.; Haerty, W.; Larcombe, L.; Macaulay, I.; Mehta, T.; Mogensen, M.; Murray, D.; Nash, W.; Neale, M.J.; et al. Meiosis and beyond-understanding the mechanistic and evolutionary processes shaping the germline genome. Biol. Rev. 2021, 96, 822–841. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakloushinskaya, I. Chromosome Changes in Soma and Germ Line: Heritability and Evolutionary Outcome. Genes 2022, 13, 602. https://doi.org/10.3390/genes13040602
Bakloushinskaya I. Chromosome Changes in Soma and Germ Line: Heritability and Evolutionary Outcome. Genes. 2022; 13(4):602. https://doi.org/10.3390/genes13040602
Chicago/Turabian StyleBakloushinskaya, Irina. 2022. "Chromosome Changes in Soma and Germ Line: Heritability and Evolutionary Outcome" Genes 13, no. 4: 602. https://doi.org/10.3390/genes13040602
APA StyleBakloushinskaya, I. (2022). Chromosome Changes in Soma and Germ Line: Heritability and Evolutionary Outcome. Genes, 13(4), 602. https://doi.org/10.3390/genes13040602