
Pleiotropic Effects of Common and Rare GCKR Exonic Mutations on Cardiometabolic Traits

 , ,
, ,
Abstract
1. Introduction
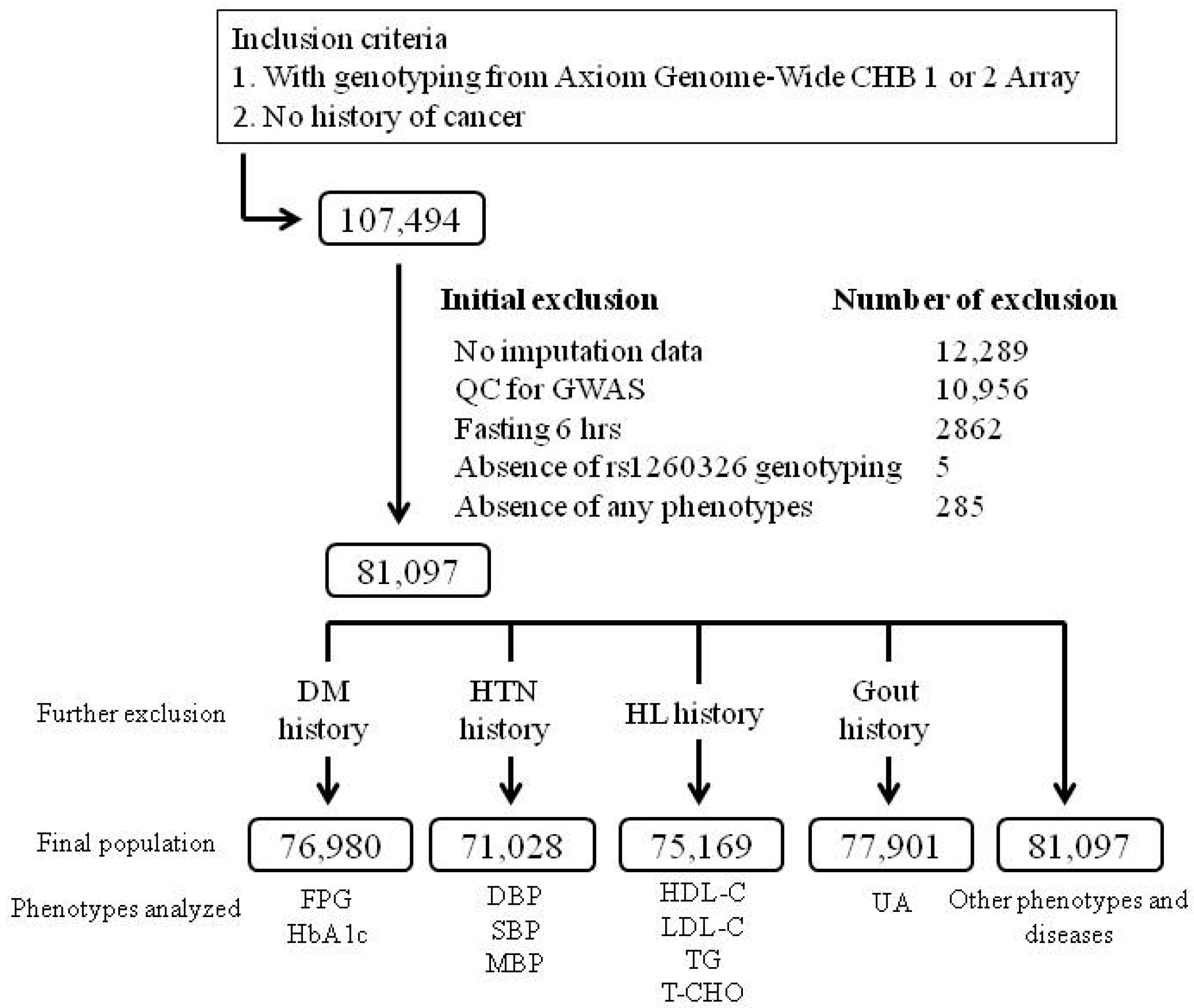
2. Materials and Methods
2.1. TWB Cohort
2.2. Clinical Phenotypes and Laboratory Examinations
2.3. Selection of GCKR Variants and Genotyping
2.4. Regional Plot Association Analysis
2.5. Selection of Rare Exonic GCKR Mutations from the Pre-QC Imputation Data for Analysis
2.6. Statistical Analysis
3. Results
3.1. Association of GCKR rs1260326 Genotypes with Clinical, Metabolic, and Biochemical Phenotypes and Hematological Parameters
3.2. Association of GCKR rs1260326 Genotypes with Risk Factors for Atherosclerosis
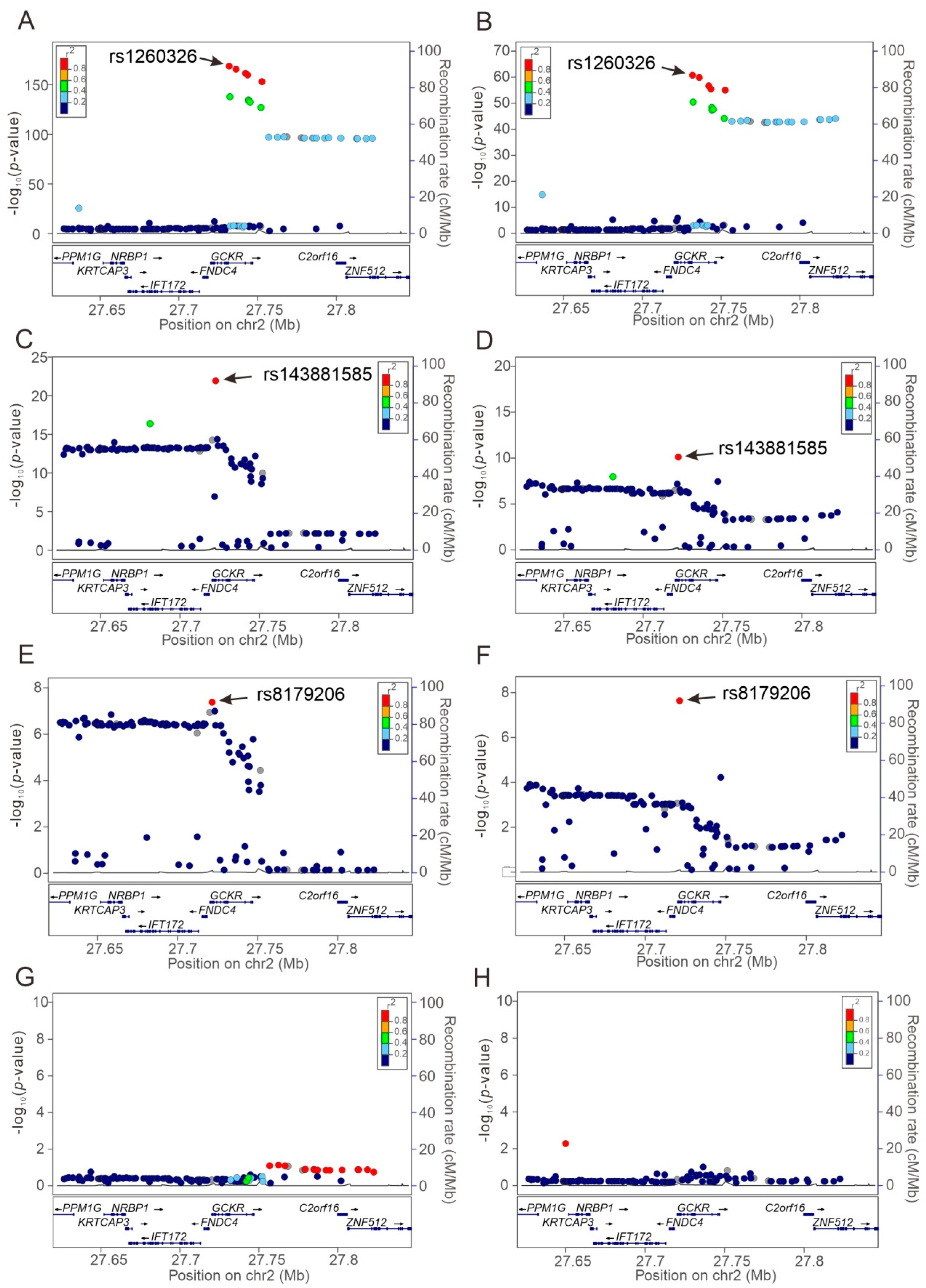
3.3. Regional Plot Association Studies for Determining the Associations of Genetic Variants at Positions 27.62 to 27.85 Mb on Chromosome 2p23.3 with Study Phenotypes
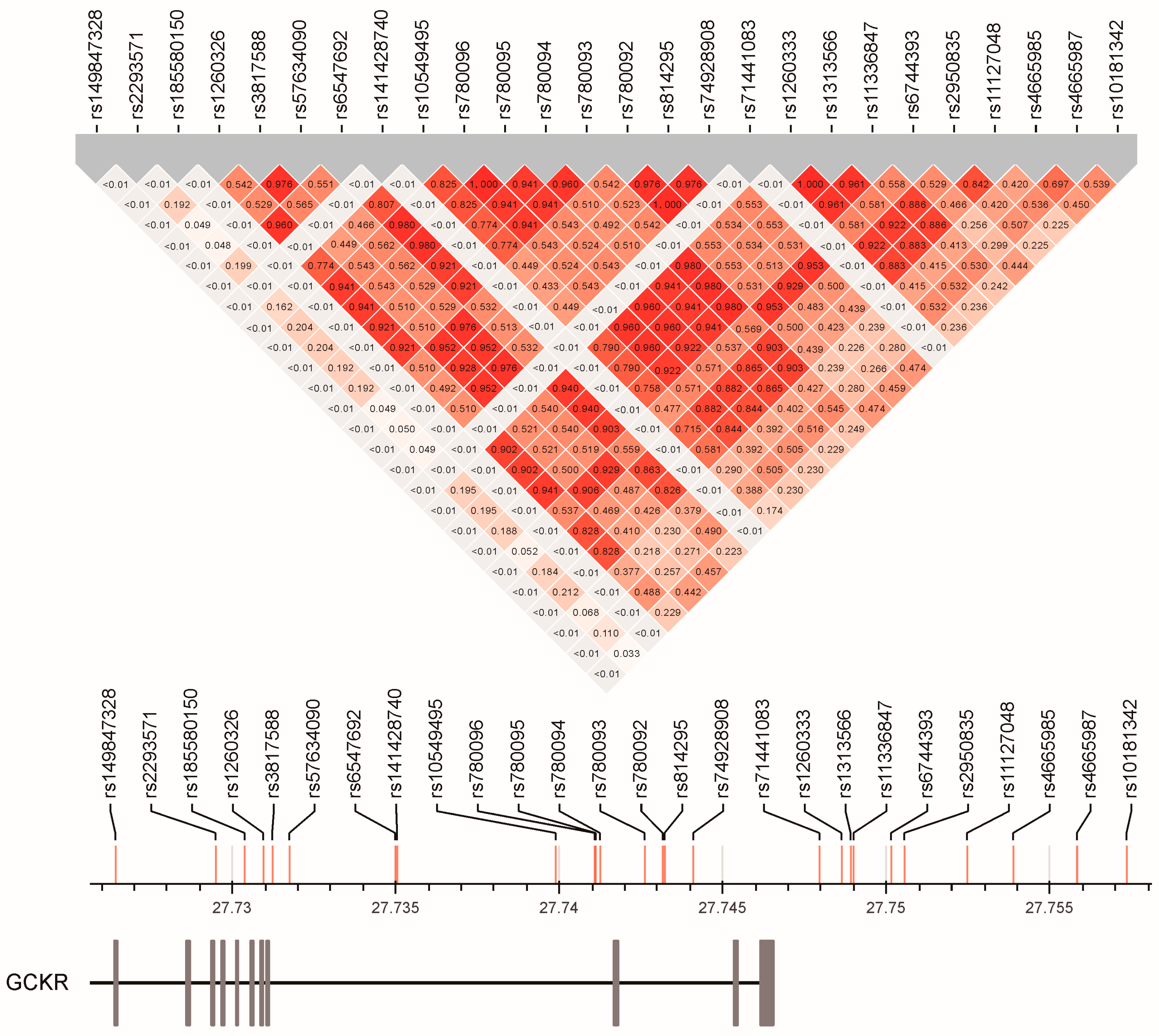
3.4. Linkage Disequilibrium between GCKR Gene Region SNPs
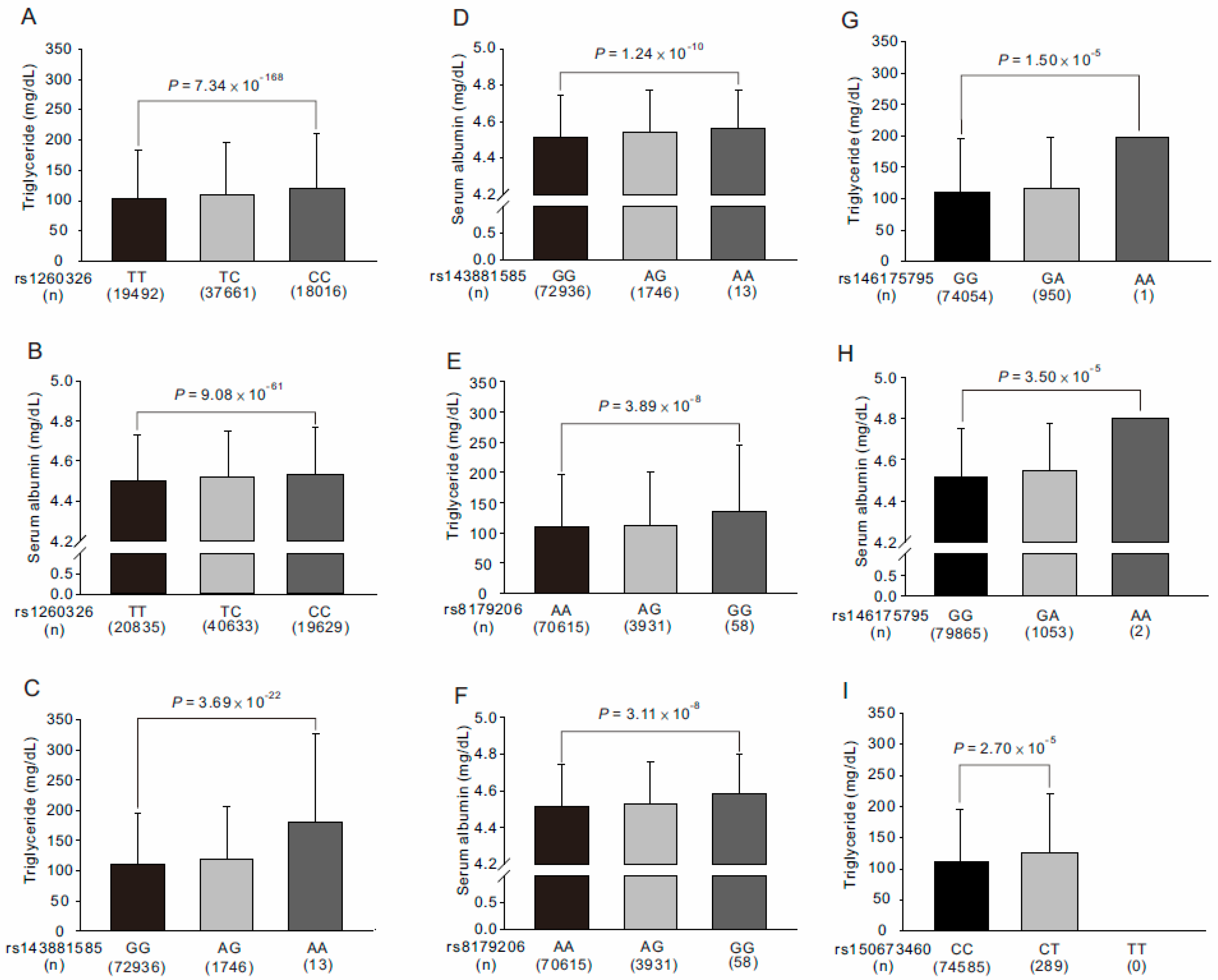
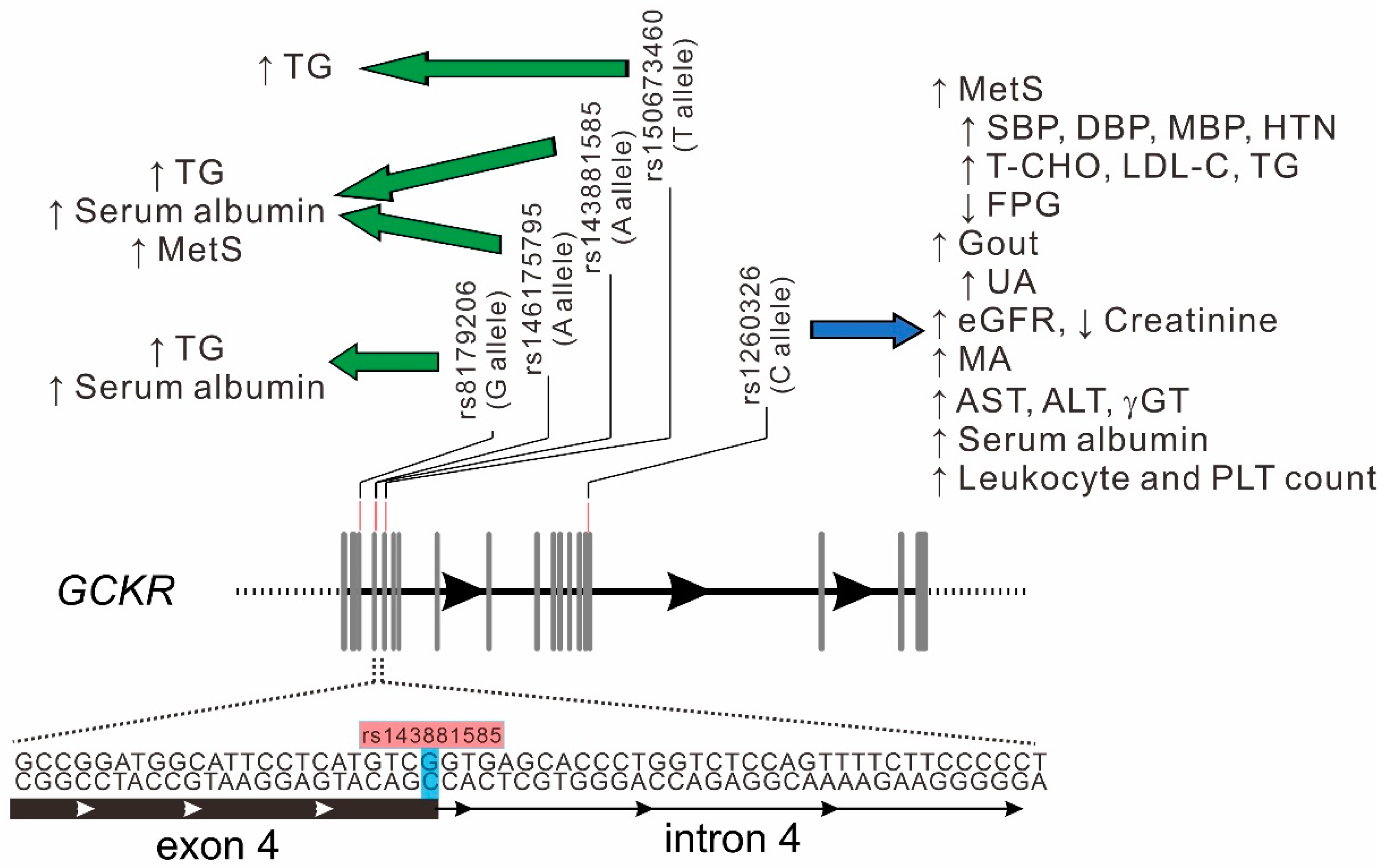
3.5. Association of GCKR rs143881585 and rs8179206 Genotypes with Clinical, Metabolic, and Biochemical Phenotypes, Hematological Parameters, and Risk Factors for Atherosclerosis
3.6. Association between Rare GCKR Exonic Mutations and Clinical Phenotypes and Laboratory Parameters
3.7. Stepwise Linear Regression Analysis for Serum Triglyceride and Albumin Levels
3.8. WGRS from the Combination of GCKR rs143881585 and rs1461755795 Revealed Significant Association with Metabolic Syndrome
4. Discussion
4.1. Pleiotropic Effect of GCKR Gene Locus
4.2. Bidirectional Effects of GCKR rs1260326 Variant on Associated Phenotypes
4.3. Association between the rs143881585 Variant and Serum Triglyceride and Albumin Levels Is Independent of the rs1260326 Variant
4.4. Association of the rs8179206 Variant with Serum Triglyceride and Albumin Levels Is Independent of the rs1260326 Variant
4.5. Role of Rare GCKR Exonic Mutations in Serum Triglyceride Levels
4.6. GCKR Variants and Serum Albumin Levels
4.7. Association between GCKR Variants and Metabolic Syndrome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, M.C.; Coate, K.C.; Winnick, J.J.; An, Z.; Cherrington, A.D. Regulation of hepatic glucose uptake and storage in vivo. Adv. Nutr. 2012, 3, 286–294. [Google Scholar] [CrossRef]
- Iynedjian, P.B. Molecular physiology of mammalian glucokinase. Cell. Mol. Life Sci. 2009, 66, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, A.; Rees, M.G.; Gloyn, A.L. Glucokinase regulatory protein: Complexity at the crossroads of triglyceride and glucose metabolism. Curr. Opin. Lipidol. 2015, 26, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Toyoda, Y.; Miwa, I.; Satake, S.; Anai, M.; Oka, Y. Nuclear location of the regulatory protein of glucokinase in rat liver and translocation of the regulator to the cytoplasm in response to high glucose. Biochem. Biophys. Res. Commun. 1995, 215, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Veiga-da-Cunha, M.; Van Schaftingen, E. Identification of fructose 6-phosphate- and fructose 1-phosphate-binding residues in the regulatory protein of glucokinase. J. Biol. Chem. 2002, 277, 8466–8473. [Google Scholar] [CrossRef] [PubMed]
- Slosberg, E.D.; Desai, U.J.; Fanelli, B.; St Denny, I.; Connelly, S.; Kaleko, M.; Boettcher, B.R.; Caplan, S.L. Treatment of type 2 diabetes by adenoviral-mediated overexpression of the glucokinase regulatory protein. Diabetes 2001, 50, 1813–1820. [Google Scholar] [CrossRef]
- Farrelly, D.; Brown, K.S.; Tieman, A.; Ren, J.; Lira, S.A.; Hagan, D.; Gregg, R.; Mookhtiar, K.A.; Hariharan, N. Mice mutant for glucokinase regulatory protein exhibit decreased liver glucokinase: A sequestration mechanism in metabolic regulation. Proc. Natl. Acad. Sci. USA 1999, 96, 14511–14516. [Google Scholar] [CrossRef]
- Beer, N.L.; Tribble, N.D.; McCulloch, L.J.; Roos, C.; Johnson, P.R.; Orho-Melander, M.; Gloyn, A.L. The P446L variant in GCKR associated with fasting plasma glucose and triglyceride levels exerts its effect through increased glucokinase activity in liver. Hum. Mol. Genet. 2009, 18, 4081–4088. [Google Scholar] [CrossRef] [PubMed]
- Veiga-da-Cunha, M.; Delplanque, J.; Gillain, A.; Bonthron, D.T.; Boutin, P.; Van Schaftingen, E.; Froguel, P. Mutations in the glucokinase regulatory protein gene in 2p23 in obese French caucasians. Diabetologia 2003, 46, 704–711. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Warner, J.P.; Leek, J.P.; Intody, S.; Markham, A.F.; Bonthron, D.T. Human glucokinase regulatory protein (GCKR): cDNA and genomic cloning, complete primary structure, and chromosomal localization. Mamm. Genome Off. J. Int. Mamm. Genome Soc. 1995, 6, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Astle, W.J.; Elding, H.; Jiang, T.; Allen, D.; Ruklisa, D.; Mann, A.L.; Mead, D.; Bouman, H.; Riveros-Mckay, F.; Kostadima, M.A.; et al. The Allelic Landscape of Human Blood Cell Trait Variation and Links to Common Complex Disease. Cell 2016, 167, 1415–1429.e1419. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.H.; Raffield, L.M.; Mousas, A.; Sakaue, S.; Huffman, J.E.; Moscati, A.; Trivedi, B.; Jiang, T.; Akbari, P.; Vuckovic, D.; et al. Trans-ethnic and Ancestry-Specific Blood-Cell Genetics in 746,667 Individuals from 5 Global Populations. Cell 2020, 182, 1198–1213.e1114. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, N.; van Rooij, F.J.; Prins, B.P.; Feitosa, M.F.; Karakas, M.; Eckfeldt, J.H.; Folsom, A.R.; Kopp, J.; Vaez, A.; Andrews, J.S.; et al. Discovery and fine mapping of serum protein loci through transethnic meta-analysis. Am. J. Hum. Genet. 2012, 91, 744–753. [Google Scholar] [CrossRef][Green Version]
- Hoffmann, T.J.; Theusch, E.; Haldar, T.; Ranatunga, D.K.; Jorgenson, E.; Medina, M.W.; Kvale, M.N.; Kwok, P.Y.; Schaefer, C.; Krauss, R.M.; et al. A large electronic-health-record-based genome-wide study of serum lipids. Nat. Genet. 2018, 50, 401–413. [Google Scholar] [CrossRef]
- Hwang, J.Y.; Sim, X.; Wu, Y.; Liang, J.; Tabara, Y.; Hu, C.; Hara, K.; Tam, C.H.; Cai, Q.; Zhao, Q.; et al. Genome-wide association meta-analysis identifies novel variants associated with fasting plasma glucose in East Asians. Diabetes 2015, 64, 291–298. [Google Scholar] [CrossRef]
- Kanai, M.; Akiyama, M.; Takahashi, A.; Matoba, N.; Momozawa, Y.; Ikeda, M.; Iwata, N.; Ikegawa, S.; Hirata, M.; Matsuda, K.; et al. Genetic analysis of quantitative traits in the Japanese population links cell types to complex human diseases. Nat. Genet. 2018, 50, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Klarin, D.; Damrauer, S.M.; Cho, K.; Sun, Y.V.; Teslovich, T.M.; Honerlaw, J.; Gagnon, D.R.; DuVall, S.L.; Li, J.; Peloso, G.M.; et al. Genetics of blood lipids among ~300,000 multi-ethnic participants of the Million Veteran Program. Nat. Genet. 2018, 50, 1514–1523. [Google Scholar] [CrossRef] [PubMed]
- Kolz, M.; Johnson, T.; Sanna, S.; Teumer, A.; Vitart, V.; Perola, M.; Mangino, M.; Albrecht, E.; Wallace, C.; Farrall, M.; et al. Meta-analysis of 28,141 individuals identifies common variants within five new loci that influence uric acid concentrations. PLoS Genet. 2009, 5, e1000504. [Google Scholar] [CrossRef]
- Köttgen, A.; Pattaro, C.; Böger, C.A.; Fuchsberger, C.; Olden, M.; Glazer, N.L.; Parsa, A.; Gao, X.; Yang, Q.; Smith, A.V.; et al. New loci associated with kidney function and chronic kidney disease. Nat. Genet. 2010, 42, 376–384. [Google Scholar] [CrossRef]
- Kraja, A.T.; Vaidya, D.; Pankow, J.S.; Goodarzi, M.O.; Assimes, T.L.; Kullo, I.J.; Sovio, U.; Mathias, R.A.; Sun, Y.V.; Franceschini, N.; et al. A bivariate genome-wide approach to metabolic syndrome: STAMPEED consortium. Diabetes 2011, 60, 1329–1339. [Google Scholar] [CrossRef]
- Ligthart, S.; Vaez, A.; Hsu, Y.H.; Stolk, R.; Uitterlinden, A.G.; Hofman, A.; Alizadeh, B.Z.; Franco, O.H.; Dehghan, A. Bivariate genome-wide association study identifies novel pleiotropic loci for lipids and inflammation. BMC Genom. 2016, 17, 443. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, H.; Yamamoto, K.; Nakaoka, H.; Nakayama, A.; Sakiyama, M.; Chiba, T.; Takahashi, A.; Nakamura, T.; Nakashima, H.; Takada, Y.; et al. Genome-wide association study of clinically defined gout identifies multiple risk loci and its association with clinical subtypes. Ann. Rheum. Dis. 2016, 75, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Orho-Melander, M.; Melander, O.; Guiducci, C.; Perez-Martinez, P.; Corella, D.; Roos, C.; Tewhey, R.; Rieder, M.J.; Hall, J.; Abecasis, G.; et al. Common missense variant in the glucokinase regulatory protein gene is associated with increased plasma triglyceride and C-reactive protein but lower fasting glucose concentrations. Diabetes 2008, 57, 3112–3121. [Google Scholar] [CrossRef] [PubMed]
- Speliotes, E.K.; Yerges-Armstrong, L.M.; Wu, J.; Hernaez, R.; Kim, L.J.; Palmer, C.D.; Gudnason, V.; Eiriksdottir, G.; Garcia, M.E.; Launer, L.J.; et al. Genome-wide association analysis identifies variants associated with nonalcoholic fatty liver disease that have distinct effects on metabolic traits. PLoS Genet. 2011, 7, e1001324. [Google Scholar] [CrossRef]
- Stančáková, A.; Paananen, J.; Soininen, P.; Kangas, A.J.; Bonnycastle, L.L.; Morken, M.A.; Collins, F.S.; Jackson, A.U.; Boehnke, M.L.; Kuusisto, J.; et al. Effects of 34 risk loci for type 2 diabetes or hyperglycemia on lipoprotein subclasses and their composition in 6,580 nondiabetic Finnish men. Diabetes 2011, 60, 1608–1616. [Google Scholar] [CrossRef][Green Version]
- Anstee, Q.M.; Darlay, R.; Cockell, S.; Meroni, M.; Govaere, O.; Tiniakos, D.; Burt, A.D.; Bedossa, P.; Palmer, J.; Liu, Y.L.; et al. Genome-wide association study of non-alcoholic fatty liver and steatohepatitis in a histologically characterised cohort. J. Hepatol. 2020, 73, 505–515. [Google Scholar] [CrossRef]
- Rees, M.G.; Wincovitch, S.; Schultz, J.; Waterstradt, R.; Beer, N.L.; Baltrusch, S.; Collins, F.S.; Gloyn, A.L. Cellular characterisation of the GCKR P446L variant associated with type 2 diabetes risk. Diabetologia 2012, 55, 114–122. [Google Scholar] [CrossRef]
- Rees, M.G.; Ng, D.; Ruppert, S.; Turner, C.; Beer, N.L.; Swift, A.J.; Morken, M.A.; Below, J.E.; Blech, I.; Mullikin, J.C.; et al. Correlation of rare coding variants in the gene encoding human glucokinase regulatory protein with phenotypic, cellular, and kinetic outcomes. J. Clin. Investig. 2012, 122, 205–217. [Google Scholar] [CrossRef]
- Rees, M.G.; Raimondo, A.; Wang, J.; Ban, M.R.; Davis, M.I.; Barrett, A.; Ranft, J.; Jagdhuhn, D.; Waterstradt, R.; Baltrusch, S.; et al. Inheritance of rare functional GCKR variants and their contribution to triglyceride levels in families. Hum. Mol. Genet. 2014, 23, 5570–5578. [Google Scholar] [CrossRef]
- Hsu, L.A.; Chou, H.H.; Teng, M.S.; Wu, S.; Ko, Y.L. Circulating chemerin levels are determined through circulating platelet counts in nondiabetic Taiwanese people: A bidirectional Mendelian randomization study. Atherosclerosis 2021, 320, 61–69. [Google Scholar] [CrossRef]
- Wang, Q.; Dhindsa, R.S.; Carss, K.; Harper, A.R.; Nag, A.; Tachmazidou, I.; Vitsios, D.; Deevi, S.V.V.; Mackay, A.; Muthas, D.; et al. Rare variant contribution to human disease in 281,104 UK Biobank exomes. Nature 2021, 597, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Kocarnik, J.M.; Richard, M.; Graff, M.; Haessler, J.; Bien, S.; Carlson, C.; Carty, C.L.; Reiner, A.P.; Avery, C.L.; Ballantyne, C.M.; et al. Discovery, fine-mapping, and conditional analyses of genetic variants associated with C-reactive protein in multiethnic populations using the Metabochip in the Population Architecture using Genomics and Epidemiology (PAGE) study. Hum. Mol. Genet. 2018, 27, 2940–2953. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ferreira, T.; Morris, A.P.; Medland, S.E.; Genetic Investigation of, A.T.C.; Replication, D.I.G.; Meta-analysis, C.; Madden, P.A.; Heath, A.C.; Martin, N.G.; et al. Conditional and joint multiple-SNP analysis of GWAS summary statistics identifies additional variants influencing complex traits. Nat. Genet. 2012, 44, 369–375, S361–S363. [Google Scholar] [CrossRef]
- Kim, Y.K.; Hwang, M.Y.; Kim, Y.J.; Moon, S.; Han, S.; Kim, B.J. Evaluation of pleiotropic effects among common genetic loci identified for cardio-metabolic traits in a Korean population. Cardiovasc. Diabetol. 2016, 15, 20. [Google Scholar] [CrossRef][Green Version]
- Ling, Y.; Li, X.; Gu, Q.; Chen, H.; Lu, D.; Gao, X. Associations of common polymorphisms in GCKR with type 2 diabetes and related traits in a Han Chinese population: A case-control study. BMC Med. Genet. 2011, 12, 66. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Wang, X.; Lu, M.; Yang, Y.; An, Y.; Zhang, J.; Chen, X.; Li, L.; Li, S.; Jiang, J.; et al. Glucokinase regulatory protein (GCKR) gene rs4425043 polymorphism is associated with overweight and obesity in Chinese women. Lipids 2011, 46, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Hong, S.C.; Yin, R.X.; Yang, Q.; Cao, X.L.; Chen, W.X. Polymorphisms in the GCKR are associated with serum lipid traits, the risk of coronary artery disease and ischemic stroke. Int. J. Clin. Exp. Med. 2015, 8, 10678–10686. [Google Scholar] [PubMed]
- Johansen, C.T.; Wang, J.; Lanktree, M.B.; Cao, H.; McIntyre, A.D.; Ban, M.R.; Martins, R.A.; Kennedy, B.A.; Hassell, R.G.; Visser, M.E.; et al. Excess of rare variants in genes identified by genome-wide association study of hypertriglyceridemia. Nat. Genet. 2010, 42, 684–687. [Google Scholar] [CrossRef]
- Sanghera, D.K.; Hopkins, R.; Malone-Perez, M.W.; Bejar, C.; Tan, C.; Mussa, H.; Whitby, P.; Fowler, B.; Rao, C.V.; Fung, K.A.; et al. Targeted sequencing of candidate genes of dyslipidemia in Punjabi Sikhs: Population-specific rare variants in GCKR promote ectopic fat deposition. PLoS ONE 2019, 14, e0211661. [Google Scholar] [CrossRef]
- Jin, J.L.; Sun, D.; Cao, Y.X.; Zhang, H.W.; Guo, Y.L.; Wu, N.Q.; Zhu, C.G.; Gao, Y.; Dong, Q.T.; Liu, G.; et al. Intensive genetic analysis for Chinese patients with very high triglyceride levels: Relations of mutations to triglyceride levels and acute pancreatitis. EBioMedicine 2018, 38, 171–177. [Google Scholar] [CrossRef]
- Matsunaga, A.; Nagashima, M.; Yamagishi, H.; Saku, K. Variants of Lipid-Related Genes in Adult Japanese Patients with Severe Hypertriglyceridemia. J. Atheroscler. Thromb. 2020, 27, 1264–1277. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.; Xing, C.; Garg, A. Type 1 Hyperlipoproteinemia Due to Compound Heterozygous Rare Variants in GCKR. J. Clin. Endocrinol. Metab. 2016, 101, 3884–3887. [Google Scholar] [CrossRef] [PubMed]
- Kamatani, Y.; Matsuda, K.; Okada, Y.; Kubo, M.; Hosono, N.; Daigo, Y.; Nakamura, Y.; Kamatani, N. Genome-wide association study of hematological and biochemical traits in a Japanese population. Nat. Genet. 2010, 42, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Osman, W.; Okada, Y.; Kamatani, Y.; Kubo, M.; Matsuda, K.; Nakamura, Y. Association of common variants in TNFRSF13B, TNFSF13, and ANXA3 with serum levels of non-albumin protein and immunoglobulin isotypes in Japanese. PLoS ONE 2012, 7, e32683. [Google Scholar] [CrossRef] [PubMed]
- Arques, S. Serum albumin and cardiovascular disease: State-of-the-art review. Ann. De Cardiol. Et D’angeiologie 2020, 69, 192–200. [Google Scholar] [CrossRef]
- Seidu, S.; Kunutsor, S.K.; Khunti, K. Serum albumin, cardiometabolic and other adverse outcomes: Systematic review and meta-analyses of 48 published observational cohort studies involving 1,492,237 participants. Scand. Cardiovasc. J. SCJ 2020, 54, 280–293. [Google Scholar] [CrossRef]
- Loomis, S.J.; Li, M.; Maruthur, N.M.; Baldridge, A.S.; North, K.E.; Mei, H.; Morrison, A.; Carson, A.P.; Pankow, J.S.; Boerwinkle, E.; et al. Genome-Wide Association Study of Serum Fructosamine and Glycated Albumin in Adults Without Diagnosed Diabetes: Results From the Atherosclerosis Risk in Communities Study. Diabetes 2018, 67, 1684–1696. [Google Scholar] [CrossRef]
- Inaba, M.; Okuno, S.; Kumeda, Y.; Yamada, S.; Imanishi, Y.; Tabata, T.; Okamura, M.; Okada, S.; Yamakawa, T.; Ishimura, E.; et al. Glycated albumin is a better glycemic indicator than glycated hemoglobin values in hemodialysis patients with diabetes: Effect of anemia and erythropoietin injection. J. Am. Soc. Nephrol. 2007, 18, 896–903. [Google Scholar] [CrossRef]
- Wu, W.C.; Ma, W.Y.; Wei, J.N.; Yu, T.Y.; Lin, M.S.; Shih, S.R.; Hua, C.H.; Liao, Y.J.; Chuang, L.M.; Li, H.Y. Serum Glycated Albumin to Guide the Diagnosis of Diabetes Mellitus. PLoS ONE 2016, 11, e0146780. [Google Scholar] [CrossRef]
- Freitas, P.A.C.; Ehlert, L.R.; Camargo, J.L. Glycated albumin: A potential biomarker in diabetes. Arch. Endocrinol. Metab. 2017, 61, 296–304. [Google Scholar] [CrossRef]
- Zendjabil, M. Glycated albumin. Clin. Chim. Acta Int. J. Clin. Chem. 2020, 502, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Neelofar, K.; Ahmad, J. An overview of in vitro and in vivo glycation of albumin: A potential disease marker in diabetes mellitus. Glycoconj. J. 2017, 34, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Nathan, D.M.; McGee, P.; Steffes, M.W.; Lachin, J.M. Relationship of glycated albumin to blood glucose and HbA1c values and to retinopathy, nephropathy, and cardiovascular outcomes in the DCCT/EDIC study. Diabetes 2014, 63, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Hovsepian, S.; Javanmard, S.H.; Mansourian, M.; Tajadini, M.; Hashemipour, M.; Kelishadi, R. Interaction of GCKR, MLXIPL and FADS genes polymorphisms with obesity in the occurrence of childhood metabolic syndrome. Middle East J. Fam. Med. 2018, 16, 20–28. [Google Scholar] [CrossRef]
- Rousseaux, J.; Duhamel, A.; Dumont, J.; Dallongeville, J.; Molnar, D.; Widhalm, K.; Manios, Y.; Sjöström, M.; Kafatos, A.; Breidenassel, C.; et al. The n-3 long-chain PUFAs modulate the impact of the GCKR Pro446Leu polymorphism on triglycerides in adolescents. J. Lipid Res. 2015, 56, 1774–1780. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical and Laboratory Parameters | Total | β | SE | p Value * |
|---|---|---|---|---|
| Anthropology | ||||
| Age (years) | 51.0 (41.0–59.0) | 0.0767 | 0.0530 | 0.1481 |
| Waist circumference (cm) | 83.0 (76.0–89.5) | 0.0533 | 0.0253 | 0.0350 |
| Waist–hip ratio | 0.87 ± 0.07 | 0.0002 | 0.0003 | 0.3493 |
| Body mass index (kg/m2) | 23.8 (21.6–26.3) | −0.0318 | 0.0180 | 0.0773 |
| Blood pressure | ||||
| Systolic BP * (mmHg) | 115.0(105.0–127.0) | 0.3931 | 0.0762 | 2.48 × 10−7 |
| Diastolic BP * (mmHg) | 71.0 (65.0–79.0) | 0.2182 | 0.0497 | 1.10 × 10−5 |
| Mean BP * (mmHg) | 86.0 (78.7–94.3) | 0.2765 | 0.0546 | 4.08 × 10−7 |
| Lipid profiles | ||||
| Total cholesterol # (mg/dL) | 171.0 (193.0–216.0) | 0.0052 | 0.0004 | 1.74 × 10−39 |
| HDL cholesterol # (mg/dL) | 53.0 (45.0–63.0) | −0.0003 | 0.0005 | 0.5755 |
| LDL cholesterol # (mg/dL) | 119.0 (99.0–140.0) | 0.0047 | 0.0006 | 1.90 × 10−15 |
| Triglyceride # (mg/dL) | 90.0 (63.0–132.0) | 0.0302 | 0.0011 | 7.34 × 10−168 |
| Glucose metabolism | ||||
| Fasting plasma glucose ** (mg/dL) | 92.0 (87.0–97.0) | −0.6133 | 0.0750 | 2.83 × 10−16 |
| HbA1C ** (%) | 5.6 (5.4–5.9) | −0.0038 | 0.0030 | 0.2031 |
| Uric acid | ||||
| Uric acid *** (mg/dL) | 5.2 (4.4–6.2) | 0.0714 | 0.0055 | 4.72 × 10−38 |
| Renal function | ||||
| Creatinine (mg/dL) | 0.68 (0.57–0.83) | −0.0072 | 0.0011 | 9.45 × 10−12 |
| eGFR (mL/min/1.73 m2) | 100.7 (87.5–116.4) | 1.1035 | 0.1074 | 9.07 × 10−25 |
| Urine albumin (mg/L) | 8.7 (5.4–15.2) | 0.0147 | 0.0023 | 9.64 × 10−11 |
| Liver function | ||||
| AST (U/L) | 23.0 (20.0–27.0) | 0.3812 | 0.0602 | 2.43 × 10−10 |
| ALT (U/L) | 19.0 (14.0–27.0) | 0.3921 | 0.0929 | 2.40 × 10−5 |
| γGT (U/L) | 17.0 (12.0–26.0) | 1.3605 | 0.1550 | 1.68 × 10−18 |
| Serum albumin (g/dL) | 4.5 (4.4–4.6) | 0.0182 | 0.0011 | 9.08 × 10−61 |
| Total bilirubin (mg/dL) | 0.6 (0.5–0.8) | 0.0022 | 0.0013 | 0.0968 |
| Hematological parameters | ||||
| Leukocyte count (103/μL) | 5.7 (4.7–6.8) | 0.0523 | 0.0076 | 8.17 × 10−12 |
| Hematocrit (%) | 41.6 (39.0–44.5) | −0.0387 | 0.0173 | 0.0256 |
| Platelet count (103/μL) | 237.0 (202.0–276.0) | 2.0649 | 0.2833 | 3.17 × 10−13 |
| Red blood cell count (106/μL) | 4.7 (4.4–5.0) | −0.0052 | 0.0022 | 0.0189 |
| Hemoglobin (g/dL) | 13.7 (12.8–14.8) | −0.0118 | 0.0061 | 0.0545 |
| Genotypes | TT | TC | CC | β | SE | p Value * |
|---|---|---|---|---|---|---|
| Diabetes mellitus (%) | 9.8 | 9.4 | 9.4 | −0.0284 | 0.0178 | 0.1105 |
| Hypertension (%) | 21.6 | 22.3 | 23.4 | 0.0639 | 0.0132 | 1.00 × 10−6 |
| Current smoking (%) | 9.2 | 9 | 9.2 | 0.0156 | 0.0182 | 0.3908 |
| Gout (%) | 3.7 | 3.8 | 4.5 | 0.1153 | 0.0266 | 1.40 × 10−5 |
| Microalbuminuria (%) | 10.6 | 11.2 | 12.4 | 0.0953 | 0.0159 | 2.16 × 10−9 |
| Metabolic syndrome (%) | 23.9 | 24.8 | 26.2 | 0.0884 | 0.0133 | 2.51 × 10−11 |
| Phenotypes | Lead SNPs | p Value | Position | Allele # | MAF | LD ## | Function | Amino Acid (Codon) |
|---|---|---|---|---|---|---|---|---|
| Triglyceride (mg/dL) | rs1260326 | 7.34 × 10−168 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| rs143881585 * | 3.69 × 10−22 | 27498323 | G/A | 0.0132 | <0.015 | Synonymous Variant | Ser118Ser | |
| rs8179206 ** | 3.89 × 10−8 | 27497575 | A/G | 0.0271 | 0.029 | Missense variant | Glu77Gly | |
| Serum albumin (mg/L) | rs1260326 | 9.08 × 10−61 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| rs143881585 * | 1.24 × 10−10 | 27498323 | G/A | 0.0132 | <0.015 | Synonymous Variant | Ser118Ser | |
| rs8179206 ** | 3.11 × 10−8 | 27497575 | A/G | 0.0271 | 0.029 | Missense variant | Glu77Gly | |
| Systolic BP (mmHg) | rs1260326 | 2.48 × 10−7 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| Diastolic BP (mmHg) | rs1260326 | 1.10 × 10−5 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| Mean BP (mmHg) | rs1260326 | 4.08 × 10−7 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| Total cholesterol (mg/dL) | rs1260326 | 1.74 × 10−39 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| LDL cholesterol (mg/dL) | rs1260326 | 1.90 × 10−15 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| Fasting plasma glucose (mg/dL) | rs1260326 | 2.83 × 10−16 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| Uric acid (mg/dL) | rs1260326 | 4.72 × 10−38 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| Creatinine (mg/dL) | rs2950835 | 9.45 × 10−12 | 27527678 | A/G | 0.5040 | 0.828 | Downstream gene variant | -- |
| eGFR (mL/min/1.73 m2) | rs2950835 | 9.07 × 10−25 | 27527678 | A/G | 0.5040 | 0.828 | Downstream gene variant | -- |
| Urine albumin (mg/L) | rs1260326 | 9.64 × 10−11 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| AST (U/L) | rs1260326 | 2.43 × 10−10 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| ALT (U/L) | rs12989678 | 2.40 × 10−5 | 27598615 | C/T | 0.4935 | 0.476 | Intron variant | -- |
| γGT (U/L) | rs780093 | 1.68 × 10−18 | 27519736 | T/C | 0.4941 | 0.921 | Intron variant | -- |
| Leukocyte counts (103/μL) | rs6744393 | 8.17 × 10−12 | 27527272 | C/T | 0.3524 | 0.537 | Downstream gene variant | -- |
| Platelet counts (103/μL) | rs6547692 | 3.17 × 10−13 | 27512105 | G/A | 0.4944 | 0.960 | Intron variant | -- |
| Hypertension | rs2950835 | 1.00 × 10−6 | 27527678 | A/G | 0.5040 | 0.828 | Downstream gene variant | -- |
| Gout | rs780094 | 1.40 × 10−5 | 27518370 | T/C | 0.5099 | 0.921 | Intron variant | -- |
| Microalbuminuria | rs6547692 | 2.16 × 10−9 | 27512105 | G/A | 0.4944 | 0.960 | Intron variant | -- |
| Metabolic syndrome | rs1260326 | 2.49 × 10−11 | 27508073 | T/C | 0.4997 | 1 | Missense variant | Pro446Leu |
| Serum Triglyceride Level (75,169 *) | Serum Albumin Level (81,097) | |||||
|---|---|---|---|---|---|---|
| β | r2 | P | β | r2 | P | |
| Age (years) | 0.0031 | 0.0175 | <10−307 | −0.0034 | 0.0243 | <10−307 |
| Sex (male vs. female) | −0.0560 | 0.0189 | 8.11 × 10−226 | −0.1179 | 0.0503 | <10−307 |
| Body mass index (kg/m2) | 0.0223 | 0.1475 | <10−307 | −0.0033 | 0.0028 | 1.23 × 10−52 |
| Current smoking (%) | 0.0819 | 0.0090 | 6.70 × 10−181 | −0.0202 | 0.0005 | 2.47 × 10−12 |
| rs1260326 (TT vs. TC vs. CC) | 0.0328 | 0.0083 | 1.55 × 10−188 | 0.0205 | 0.0032 | 1.58 × 10−73 |
| rs143881585 (GG vs. GA vs. AA) | 0.0499 | 0.0010 | 1.11 × 10−22 | 0.0344 | 0.0005 | 2.14 × 10−11 |
| rs146175795 (GG vs. GA) | 0.0474 | 0.0005 | 8.36 × 10−12 | 0.0401 | 0.0004 | 6.83 × 10−9 |
| rs8179206 (AA vs. AG vs. GG) | 0.0190 | 0.0003 | 2.13 × 10−8 | 0.0194 | 0.0004 | 1.60 × 10−8 |
| rs150673460 (CC vs. CT) | 0.0401 | 0.0001 | 0.0013 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeh, K.-H.; Hsu, L.-A.; Teng, M.-S.; Wu, S.; Chou, H.-H.; Ko, Y.-L. Pleiotropic Effects of Common and Rare GCKR Exonic Mutations on Cardiometabolic Traits. Genes 2022, 13, 491. https://doi.org/10.3390/genes13030491
Yeh K-H, Hsu L-A, Teng M-S, Wu S, Chou H-H, Ko Y-L. Pleiotropic Effects of Common and Rare GCKR Exonic Mutations on Cardiometabolic Traits. Genes. 2022; 13(3):491. https://doi.org/10.3390/genes13030491
Chicago/Turabian StyleYeh, Kuan-Hung, Lung-An Hsu, Ming-Sheng Teng, Semon Wu, Hsin-Hua Chou, and Yu-Lin Ko. 2022. "Pleiotropic Effects of Common and Rare GCKR Exonic Mutations on Cardiometabolic Traits" Genes 13, no. 3: 491. https://doi.org/10.3390/genes13030491
APA StyleYeh, K.-H., Hsu, L.-A., Teng, M.-S., Wu, S., Chou, H.-H., & Ko, Y.-L. (2022). Pleiotropic Effects of Common and Rare GCKR Exonic Mutations on Cardiometabolic Traits. Genes, 13(3), 491. https://doi.org/10.3390/genes13030491