
The BDNF Val66Met Polymorphism (rs6265) Modulates Inflammation and Neurodegeneration in the Early Phases of Multiple Sclerosis

 , , , , , , ,
, , , , , , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. MS Patients
2.2. SNP Val66Met Analysis
2.3. CSF Collection and Analysis
2.4. MRI
2.5. Statistical Analysis
3. Results
3.1. Role of BDNF Gene in Disease Characteristics of MS Patients
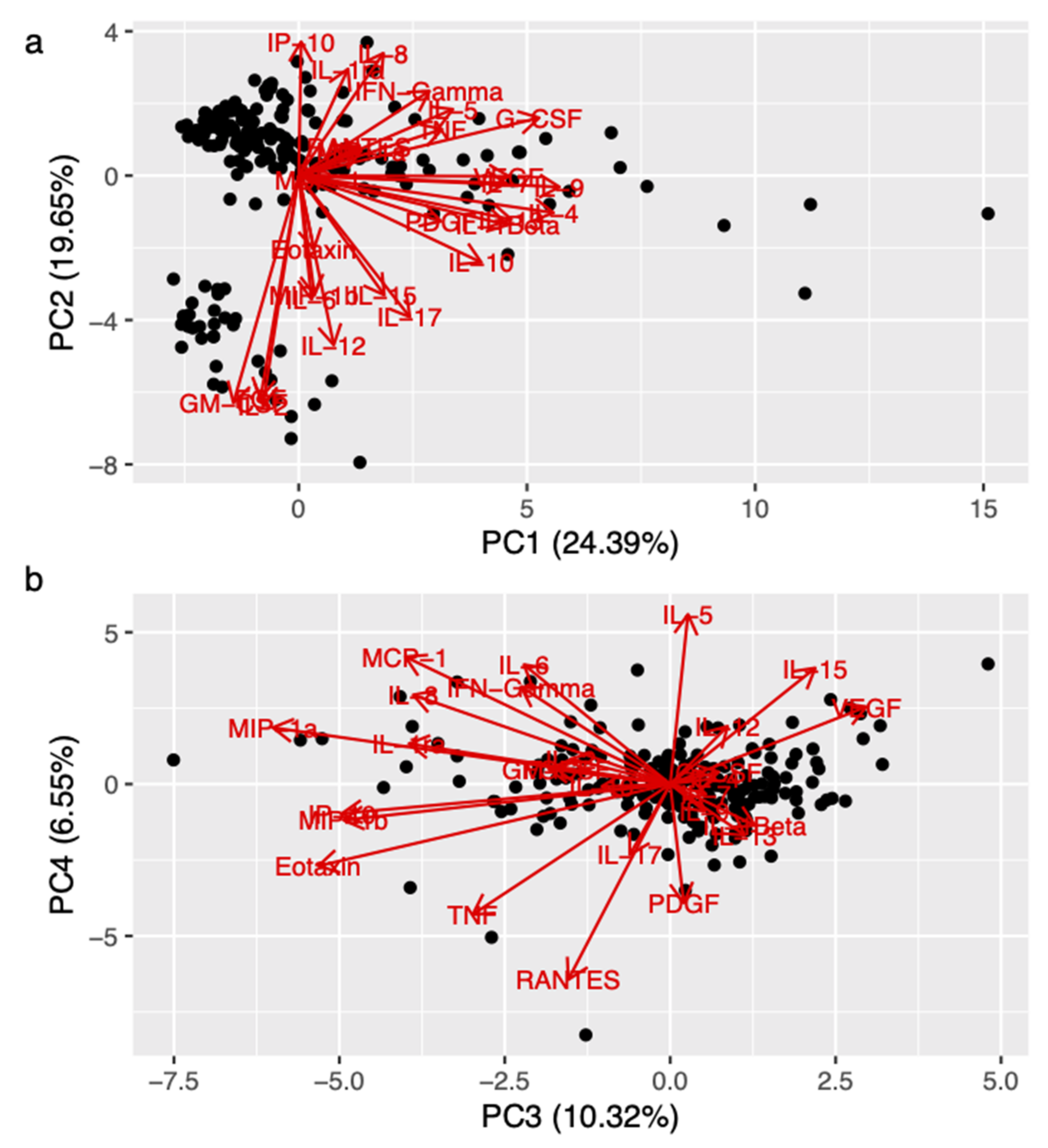
3.2. Role of BDNF Gene in the Regulation of Pro-Inflammatory Cytokine Levels in MS Patients
3.3. Role of BDNF Gene on MRI Structural Measurements in MS Patients
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thompson, A.J.; Baranzini, S.E.; Geurts, J.; Hemmer, B.; Ciccarelli, O. Multiple sclerosis. Lancet 2018, 391, 1622–1636. [Google Scholar] [CrossRef]
- Mandolesi, G.; Gentile, A.; Musella, A.; Fresegna, D.; De Vito, F.; Bullitta, S.; Sepman, H.; Marfia, G.A.; Centonze, D. Synaptopathy connects inflammation and neurodegeneration in multiple sclerosis. Nat. Rev. Neurol. 2015, 11, 711–724. [Google Scholar] [CrossRef]
- Centonze, D.; Muzio, L.; Rossi, S.; Furlan, R.; Bernardi, G.; Martino, G. The link between inflammation, synaptic transmission and neurodegeneration in multiple sclerosis. Cell Death Differ. 2009, 17, 1083–1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, F.R.; Musella, A.; De Vito, F.; Fresegna, D.; Bullitta, S.; Vanni, V.; Guadalupi, L.; Stampanoni Bassi, M.; Buttari, F.; Mandolesi, G.; et al. Tumor Necrosis Factor and Interleukin-1β Modulate Synaptic Plasticity during Neuroinflammation. Neural Plast. 2018, 2018, 8430123. [Google Scholar] [CrossRef] [Green Version]
- Rossi, S.; Furlan, R.; De Chiara, V.; Motta, C.; Studer, V.; Mori, F.; Musella, A.; Bergami, A.; Muzio, L.; Bernardi, G.; et al. Interleukin-1β causes synaptic hyperexcitability in multiple sclerosis. Ann. Neurol. 2011, 71, 76–83. [Google Scholar] [CrossRef]
- Rossi, S.; Motta, C.; Studer, V.; Barbieri, F.; Buttari, F.; Bergami, A.; Sancesario, G.; Bernardini, S.; De Angelis, G.; Martino, G.; et al. Tumor necrosis factor is elevated in progressive multiple sclerosis and causes excitotoxic neurodegeneration. Mult. Scler. J. 2013, 20, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–423. [Google Scholar] [CrossRef] [Green Version]
- Egan, M.F.; Kojima, M.; Callicott, J.; Goldberg, T.E.; Kolachana, B.S.; Bertolino, A.; Zaitsev, E.; Gold, B.; Goldman, D.; Dean, M.; et al. The BDNF val66met Polymorphism Affects Activity-Dependent Secretion of BDNF and Human Memory and Hippocampal Function. Cell 2003, 112, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Bath, K.G.; Lee, F.S. Variant BDNF (Val66Met) impact on brain structure and function. Cogn. Affect. Behav. Neurosci. 2006, 6, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.D.; Züchner, S.; McQuoid, D.R.; Payne, M.E.; MacFall, J.R.; Steffens, D.C.; Speer, M.C.; Krishnan, K.R.R. The Brain-Derived Neurotrophic Factor VAL66MET Polymorphism and Cerebral White Matter Hyperintensities in Late-Life Depression. Am. J. Geriatr. Psychiatry 2008, 16, 263–271. [Google Scholar] [CrossRef]
- Zivadinov, R.; Weinstock-Guttman, B.; Benedict, R.; Tamaño-Blanco, M.; Hussein, S.; Abdelrahman, N.; Durfee, J.; Ramanathan, M. Preservation of gray matter volume in multiple sclerosis patients with the Met allele of the rs6265 (Val66Met) SNP of brain-derived neurotrophic factor. Hum. Mol. Genet. 2007, 16, 2659–2668. [Google Scholar] [CrossRef] [Green Version]
- Shen, T.; You, Y.; Joseph, C.; Mirzaei, M.; Klistorner, A.; Graham, S.L.; Gupta, V. BDNF Polymorphism: A Review of Its Diagnostic and Clinical Relevance in Neurodegenerative Disorders. Aging Dis. 2018, 9, 523–536. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Lian, D.; Wu, J.; Liu, Y.; Zhu, M.; Sun, J.; He, D.; Li, L. Brain-derived neurotrophic factor reduces inflammation and hippocampal apoptosis in experimental Streptococcus pneumoniae meningitis. J. Neuroinflammation 2017, 14, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 Revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coupé, P.; Tourdias, T.; Linck, P.; Romero, J.E.; Manjón, J.V. LesionBrain: An Online Tool for White Matter Lesion Segmentation. In Patch-Based Techniques in Medical Imaging; Bai, W., Sanroma, G., Wu, G., Munsell, B., Zhan, Y., Coupé, P., Eds.; Springer: Cham, Switzerland, 2018; pp. 95–103. [Google Scholar] [CrossRef] [Green Version]
- Anastasia, A.; Deinhardt, K.; Chao, M.; Will, N.E.; Irmady, K.; Lee, F.S.; Hempstead, B.L.; Bracken, C. Val66Met polymorphism of BDNF alters prodomain structure to induce neuronal growth cone retraction. Nat. Commun. 2013, 4, 2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.-Y.; Patel, P.D.; Sant, G.; Meng, C.-X.; Teng, K.; Hempstead, B.L.; Lee, F.S. Variant Brain-Derived Neurotrophic Factor (BDNF) (Met66) Alters the Intracellular Trafficking and Activity-Dependent Secretion of Wild-Type BDNF in Neurosecretory Cells and Cortical Neurons. J. Neurosci. 2004, 24, 4401–4411. [Google Scholar] [CrossRef]
- Hartmann, M.; Heumann, R.; Lessmann, V. Synaptic secretion of BDNF after high-frequency stimulation of glutamatergic synapses. EMBO J. 2001, 20, 5887–5897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Sarchielli, P.; Greco, L.; Stipa, A.; Floridi, A.; Gallai, V. Brain-derived neurotrophic factor in patients with multiple sclerosis. J. Neuroimmunol. 2002, 132, 180–188. [Google Scholar] [CrossRef]
- Nociti, V. What is the role of Brain derived neurotrophic factor in Multiple Sclerosis neuroinflammation? Neuroimmunol. Neuroinflammation 2020, 7, 291–299. [Google Scholar] [CrossRef]
- Ksiazek-Winiarek, D.J.; Szpakowski, P.; Glabinski, A. Neural Plasticity in Multiple Sclerosis: The Functional and Molecular Background. Neural Plast. 2015, 2015, 307175. [Google Scholar] [CrossRef] [Green Version]
- Bassi, M.S.; Iezzi, E.; Landi, D.; Monteleone, F.; Gilio, L.; Simonelli, I.; Musella, A.; Mandolesi, G.; De Vito, F.; Furlan, R.; et al. Delayed treatment of MS is associated with high CSF levels of IL-6 and IL-8 and worse future disease course. J. Neurol. 2018, 265, 2540–2547. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.; Newcombe, J.; Cuzner, M.; Woodroofe, N. Expression of monocyte chemoattractant protein-1 and other β-chemokines by resident glia and inflammatory cells in multiple sclerosis lesions. J. Neuroimmunol. 1998, 84, 238–249. [Google Scholar] [CrossRef]
- Chen, X.-L.; Zhang, Q.; Zhao, R.; Medford, R.M. Superoxide, H2O2, and iron are required for TNF-α-induced MCP-1 gene expression in endothelial cells: Role of Rac1 and NADPH oxidase. Am. J. Physiol. Circ. Physiol. 2004, 286, H1001–H1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanuma, N.; Sakuma, H.; Sasaki, A.; Matsumoto, Y. Chemokine expression by astrocytes plays a role in microglia/macrophage activation and subsequent neurodegeneration in secondary progressive multiple sclerosis. Acta Neuropathol. 2006, 112, 195–204. [Google Scholar] [CrossRef]
- Bassi, M.S.; Gilio, L.; Iezzi, E.; Moscatelli, A.; Pekmezovic, T.; Drulovic, J.; Furlan, R.; Finardi, A.; Mandolesi, G.; Musella, A.; et al. Age at Disease Onset Associates With Oxidative Stress, Neuroinflammation, and Impaired Synaptic Plasticity in Relapsing-Remitting Multiple Sclerosis. Front. Aging Neurosci. 2021, 13, 694651. [Google Scholar] [CrossRef] [PubMed]
- Makar, T.K.; Trisler, D.; Sura, K.; Sultana, S.; Patel, N.; Bever, C.T. Brain derived neurotrophic factor treatment reduces inflammation and apoptosis in experimental allergic encephalomyelitis. J. Neurol. Sci. 2008, 270, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wei, N.; Zhu, J.; Lu, T.; Chen, Z.; Xu, G.; Liu, X. Effects of Brain-Derived Neurotrophic Factor on Local Inflammation in Experimental Stroke of Rat. Mediat. Inflamm. 2010, 2010, 372423. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Z.; Xu, D.; Wang, Y.; Li, L. Selective Loss of Brain-Derived Neurotrophic Factor Exacerbates Brain Injury by Enhancing Neuroinflammation in Experimental Streptococcus pneumoniae Meningitis. Front. Immunol. 2020, 11, 1357. [Google Scholar] [CrossRef]
- Sandrini, L.; Castiglioni, L.; Amadio, P.; Werba, J.P.; Eligini, S.; Fiorelli, S.; Zarà, M.; Castiglioni, S.; Bellosta, S.; Lee, F.S.; et al. Impact of BDNF Val66Met Polymorphism on Myocardial Infarction: Exploring the Macrophage Phenotype. Cells 2020, 9, 1084. [Google Scholar] [CrossRef] [PubMed]
- Mero, I.-L.; Smestad, C.; Lie, B.A.; Lorentzen, R.; Sandvik, L.; Landrø, N.I.; Aarseth, J.H.; Myhr, K.-M.; Celius, E.G.; Harbo, H.F. Polymorphisms of the BDNF gene show neither association with multiple sclerosis susceptibility nor clinical course. J. Neuroimmunol. 2012, 244, 107–110. [Google Scholar] [CrossRef]
- Blanco, Y.; Gómez-Choco, M.; Arostegui, J.; Casanova, B.; Martínez-Rodríguez, J.; Boscá, I.; Munteis, E.; Yagüe, J.; Graus, F.; Saiz, A. No association of the Val66Met polymorphism of brain-derived neurotrophic factor (BDNF) to multiple sclerosis. Neurosci. Lett. 2006, 396, 217–219. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, S.; Schott, B.; Ban, M.; Compston, D.; Sawcer, S.; Sailer, M. The BDNF-Val66Met polymorphism: Implications for susceptibility to multiple sclerosis and severity of disease. J. Neuroimmunol. 2005, 167, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Mirowska-Guzel, D.; Mach, A.; Gromadzka, G.; Czlonkowski, A.; Czlonkowska, A. BDNF A196G and C270T gene polymorphisms and susceptibility to multiple sclerosis in the polish population. Gender differences. J. Neuroimmunol. 2008, 193, 170–172. [Google Scholar] [CrossRef] [PubMed]
- Portaccio, E.; Bellinvia, A.; Prestipino, E.; Nacmias, B.; Bagnoli, S.; Razzolini, L.; Pastò, L.; Niccolai, C.; Goretti, B.; Fonderico, M.; et al. The Brain-Derived Neurotrophic Factor Val66Met Polymorphism Can Protect Against Cognitive Impairment in Multiple Sclerosis. Front. Neurol. 2021, 12, 645220. [Google Scholar] [CrossRef] [PubMed]
- Fera, F.; Passamonti, L.; Cerasa, A.; Gioia, M.C.; Liguori, M.; Manna, I.; Valentino, P.; Quattrone, A. The BDNF Val66Met Polymorphism Has Opposite Effects on Memory Circuits of Multiple Sclerosis Patients and Controls. PLoS ONE 2013, 8, e61063. [Google Scholar] [CrossRef] [PubMed]
- Liguori, M.; Fera, F.; Gioia, M.C.; Valentino, P.; Manna, I.; Condino, F.; Cerasa, A.; La Russa, A.; Clodomiro, A.; Paolillo, A.; et al. Investigating the role of brain-derived neurotrophic factor in relapsing-remitting multiple sclerosis. Genes Brain Behav. 2006, 6, 177–183. [Google Scholar] [CrossRef]
- Ramasamy, D.P.; Ramanathan, M.; Cox, J.L.; Antulov, R.; Weinstock-Guttman, B.; Bergsland, N.; Benedict, R.H.; Dwyer, M.G.; Minagar, A.; Zivadinov, R. Effect of Met66 allele of the BDNF rs6265 SNP on regional gray matter volumes in patients with multiple sclerosis: A voxel-based morphometry study. Pathophysiology 2011, 18, 53–60. [Google Scholar] [CrossRef] [PubMed]
- De Meo, E.; Portaccio, E.; Prestipino, E.; Nacmias, B.; Bagnoli, S.; Razzolini, L.; Pastò, L.; Niccolai, C.; Goretti, B.; Bellinvia, A.; et al. Effect of BDNF Val66Met polymorphism on hippocampal subfields in multiple sclerosis patients. Mol. Psychiatry 2021, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dinacci, D.; Tessitore, A.; Russo, A.; De Bonis, M.L.; Lavorgna, L.; Picconi, O.; Sacco, R.; Bonavita, S.; Gallo, A.; Servillo, G.; et al. BDNF Val66Met polymorphism and brain volumes in multiple sclerosis. Neurol. Sci. 2010, 32, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Bueller, J.A.; Aftab, M.; Sen, S.; Gomez-Hassan, D.; Burmeister, M.; Zubieta, J.-K. BDNF Val66Met Allele Is Associated with Reduced Hippocampal Volume in Healthy Subjects. Biol. Psychiatry 2006, 59, 812–815. [Google Scholar] [CrossRef] [PubMed]
- Frodl, T.; Schüle, C.; Schmitt, G.; Born, C.; Baghai, T.; Zill, P.; Bottlender, R.; Rupprecht, R.; Bondy, B.; Reiser, M.; et al. Association of the Brain-Derived Neurotrophic Factor Val66Met Polymorphism With Reduced Hippocampal Volumes in Major Depression. Arch. Gen. Psychiatry 2007, 64, 410–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, K.; Walss-Bass, C.; Nery, F.G.; A Nicoletti, M.; Hatch, J.P.; Frey, B.N.; Monkul, E.S.; Zunta-Soares, G.B.; Bowden, C.L.; A Escamilla, M.; et al. Neuronal Correlates of Brain-derived Neurotrophic Factor Val66Met Polymorphism and Morphometric Abnormalities in Bipolar Disorder. Neuropsychopharmacology 2009, 34, 1904–1913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezawas, L.; Verchinski, B.A.; Mattay, V.S.; Callicott, J.; Kolachana, B.S.; Straub, R.E.; Egan, M.F.; Meyer-Lindenberg, A.; Weinberger, D.R. The Brain-Derived Neurotrophic Factor val66met Polymorphism and Variation in Human Cortical Morphology. J. Neurosci. 2004, 24, 10099–10102. [Google Scholar] [CrossRef] [PubMed]
- Black, I.B. Trophic regulation of synaptic plasticity. J. Neurobiol. 1999, 41, 108–118. [Google Scholar] [CrossRef]
- Narisawa-Saito, M.; Iwakura, Y.; Kawamura, M.; Araki, K.; Kozaki, S.; Takei, N.; Nawa, H. Brain-derived Neurotrophic Factor Regulates Surface Expression of α-Amino-3-hydroxy-5-methyl-4-isoxazoleproprionic Acid Receptors by Enhancing the N-Ethylmaleimide-sensitive Factor/GluR2 Interaction in Developing Neocortical Neurons. J. Biol. Chem. 2002, 277, 40901–40910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363. [Google Scholar] [CrossRef]
- Nociti, V.; Santoro, M.; Quaranta, D.; Losavio, F.A.; De Fino, C.; Giordano, R.; Palomba, N.; Rossini, P.M.; Guerini, F.R.; Clerici, M.; et al. BDNF rs6265 polymorphism methylation in Multiple Sclerosis: A possible marker of disease progression. PLoS ONE 2018, 13, e0206140. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| RR-MS | N | 218 |
|---|---|---|
| Age at diagnosis, years | Median (IQR) | 34 (25.9–44.8) |
| Sex, F/M | N (%) | 155/63 (71.1/28.9) |
| Disease duration, years | Median (IQR) | 0.25 (0.06–1.4) |
| EDSS at diagnosis | Median (IQR) | 1.5 (1–2) |
| Radiological activity, yes | N/tot (%) | 87/206 (42.2) |
| OCB, yes | N/tot (%) | 165/214 (77.1) |
| RR-MS | N | Met Carriers (82) | Val/Val (136) | p |
|---|---|---|---|---|
| Age at diagnosis, years | Median (IQR) | 36 (26.8–45.3) | 33.3 (25.2–44.1) | 0.269 |
| Sex, F/M | N (%) | 52/30 (63/37) | 103/33 (78/24) | 0.052 |
| Disease duration, years | Median (IQR) | 0.4 (0.1–2) | 0.2 (0.1–1.1) | 0.039 * |
| EDSS at diagnosis | Median (IQR) | 1.5 (1–2) | 1.5 (1–2) | 0.390 |
| Radiological activity, yes | N/tot (%) | 39/78 (50) | 48/128 (37.5) | 0.078 |
| OCB, yes | N/tot (%) | 64/82 (78) | 101/132 (76.5) | 0.795 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dolcetti, E.; Bruno, A.; Azzolini, F.; Gilio, L.; Moscatelli, A.; De Vito, F.; Pavone, L.; Iezzi, E.; Gambardella, S.; Giardina, E.; et al. The BDNF Val66Met Polymorphism (rs6265) Modulates Inflammation and Neurodegeneration in the Early Phases of Multiple Sclerosis. Genes 2022, 13, 332. https://doi.org/10.3390/genes13020332
Dolcetti E, Bruno A, Azzolini F, Gilio L, Moscatelli A, De Vito F, Pavone L, Iezzi E, Gambardella S, Giardina E, et al. The BDNF Val66Met Polymorphism (rs6265) Modulates Inflammation and Neurodegeneration in the Early Phases of Multiple Sclerosis. Genes. 2022; 13(2):332. https://doi.org/10.3390/genes13020332
Chicago/Turabian StyleDolcetti, Ettore, Antonio Bruno, Federica Azzolini, Luana Gilio, Alessandro Moscatelli, Francesca De Vito, Luigi Pavone, Ennio Iezzi, Stefano Gambardella, Emiliano Giardina, and et al. 2022. "The BDNF Val66Met Polymorphism (rs6265) Modulates Inflammation and Neurodegeneration in the Early Phases of Multiple Sclerosis" Genes 13, no. 2: 332. https://doi.org/10.3390/genes13020332
APA StyleDolcetti, E., Bruno, A., Azzolini, F., Gilio, L., Moscatelli, A., De Vito, F., Pavone, L., Iezzi, E., Gambardella, S., Giardina, E., Ferese, R., Buttari, F., Rizzo, F. R., Furlan, R., Finardi, A., Musella, A., Mandolesi, G., Guadalupi, L., Centonze, D., & Stampanoni Bassi, M. (2022). The BDNF Val66Met Polymorphism (rs6265) Modulates Inflammation and Neurodegeneration in the Early Phases of Multiple Sclerosis. Genes, 13(2), 332. https://doi.org/10.3390/genes13020332