
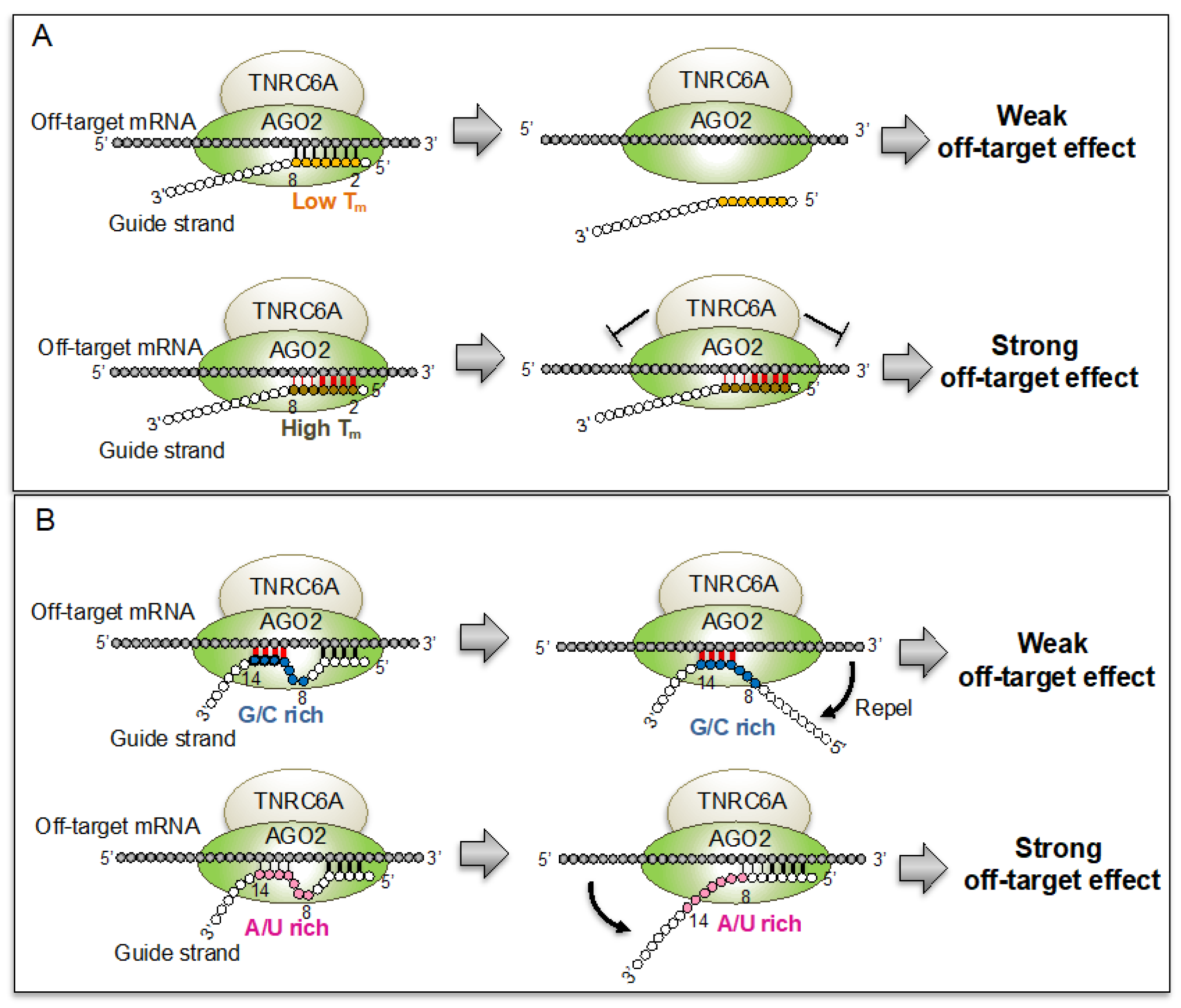
The siRNA Off-Target Effect Is Determined by Base-Pairing Stabilities of Two Different Regions with Opposite Effects



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Kobayashi, Y.; Tian, S.; Ui-Tei, K. The siRNA Off-Target Effect Is Determined by Base-Pairing Stabilities of Two Different Regions with Opposite Effects. Genes 2022, 13, 319. https://doi.org/10.3390/genes13020319
Kobayashi Y, Tian S, Ui-Tei K. The siRNA Off-Target Effect Is Determined by Base-Pairing Stabilities of Two Different Regions with Opposite Effects. Genes. 2022; 13(2):319. https://doi.org/10.3390/genes13020319
Chicago/Turabian StyleKobayashi, Yoshiaki, Shen Tian, and Kumiko Ui-Tei. 2022. "The siRNA Off-Target Effect Is Determined by Base-Pairing Stabilities of Two Different Regions with Opposite Effects" Genes 13, no. 2: 319. https://doi.org/10.3390/genes13020319
APA StyleKobayashi, Y., Tian, S., & Ui-Tei, K. (2022). The siRNA Off-Target Effect Is Determined by Base-Pairing Stabilities of Two Different Regions with Opposite Effects. Genes, 13(2), 319. https://doi.org/10.3390/genes13020319