
Make It Less difficile: Understanding Genetic Evolution and Global Spread of Clostridioides difficile

 ,
,  ,
,  ,
,  ,
, 
{kind=link}
Abstract
1. Introduction
2. Genetic Evolution of C. difficile Virulence
3. Antimicrobial Resistance Genes
4. Molecular Typing Techniques for C. difficile Strains
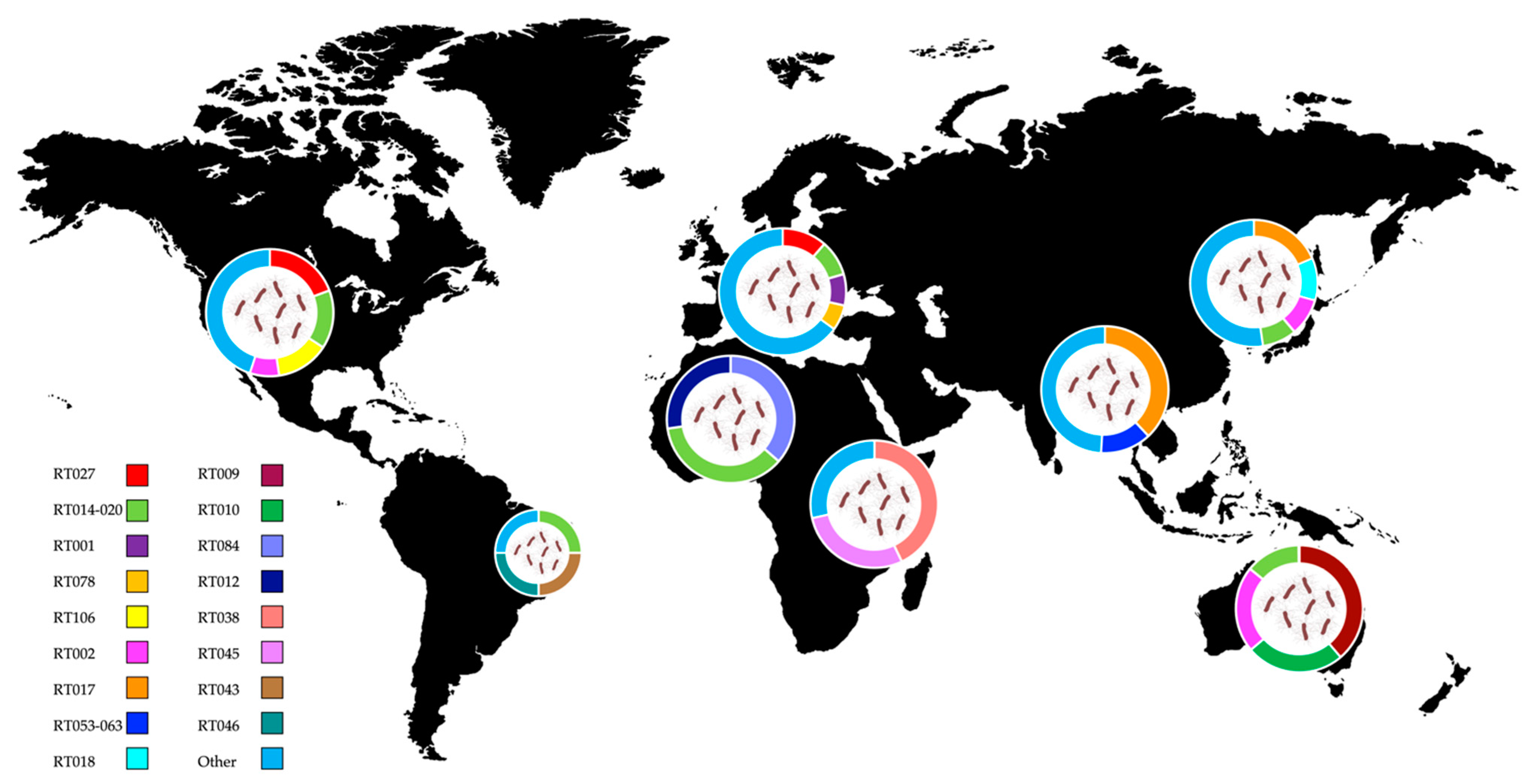
5. Worldwide Distribution of C. difficile Ribotypes
5.1. Europe
5.2. America
5.3. Asia and Middle East
5.4. Oceania
5.5. Africa
6. Future Perspectives to Make It Less difficile
6.1. Genomic Surveillance
6.2. Genetic Engineering
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lawson, P.A.; Citron, D.M.; Tyrrell, K.L.; Finegold, S.M. Reclassification of Clostridium Difficile as Clostridioides Difficile (Hall and O’Toole 1935) Prévot 1938. Anaerobe 2016, 40, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Czepiel, J.; Dróżdż, M.; Pituch, H.; Kuijper, E.J.; Perucki, W.; Mielimonka, A.; Goldman, S.; Wultańska, D.; Garlicki, A.; Biesiada, G. Clostridium Difficile Infection: Review. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1211–1221. [Google Scholar] [CrossRef]
- Hall, I.C.; O’Toole, E. Intestinal Flora in New-Born Infants: With a Description of a New Pathogenic Anaerobe, Bacillus Difficilis. Am. J. Dis. Child. 1935, 49, 390–402. [Google Scholar] [CrossRef]
- Rodriguez-Palacios, A.; LeJeune, J.T. Moist-Heat Resistance, Spore Aging, and Superdormancy in Clostridium Difficile. Appl. Environ. Microbiol. 2011, 77, 3085–3091. [Google Scholar] [CrossRef] [PubMed]
- Dawson, L.F.; Valiente, E.; Donahue, E.H.; Birchenough, G.; Wren, B.W. Hypervirulent Clostridium Difficile PCR-Ribotypes Exhibit Resistance to Widely Used Disinfectants. PLoS ONE 2011, 6, e25754. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P. I Will Survive: DNA Protection in Bacterial Spores. Trends Microbiol. 2007, 15, 172–180. [Google Scholar] [CrossRef]
- Paredes-Sabja, D.; Shen, A.; Sorg, J.A. Clostridium Difficile Spore Biology: Sporulation, Germination, and Spore Structural Proteins. Trends Microbiol. 2014, 22, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Kochan, T.J.; Foley, M.H.; Shoshiev, M.S.; Somers, M.J.; Carlson, P.E.; Hanna, P.C. Updates to Clostridium Difficile Spore Germination. J. Bacteriol. 2018, 200, e00218-18. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.H.; Kennedy, M.J.; Fekety, F.R. Use of Sodium Taurocholate to Enhance Spore Recovery on a Medium Selective for Clostridium Difficile. J. Clin. Microbiol. 1982, 15, 443–446. [Google Scholar] [CrossRef]
- Sorg, J.A.; Sonenshein, A.L. Inhibiting the Initiation of Clostridium Difficile Spore Germination Using Analogs of Chenodeoxycholic Acid, a Bile Acid. J. Bacteriol. 2010, 192, 4983–4990. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.W.; Tsai, P.J.; Lee, C.C.; Ko, W.C.; Hung, Y.P. Inhibition of Spores to Prevent the Recurrence of Clostridioides Difficile Infection—A Possibility or an Improbability? J. Microbiol. Immunol. Infect. 2021, 54, 1011–1017. [Google Scholar] [CrossRef]
- Sandhu, B.K.; McBride, S.M. Clostridioides Difficile. Trends Microbiol. 2018, 26, 1049–1050. [Google Scholar] [CrossRef] [PubMed]
- Baktash, A.; Terveer, E.M.; Zwittink, R.D.; Hornung, B.V.H.; Corver, J.; Kuijper, E.J.; Smits, W.K. Mechanistic Insights in the Success of Fecal Microbiota Transplants for the Treatment of Clostridium Difficile Infections. Front. Microbiol. 2018, 9, 1242. [Google Scholar] [CrossRef] [PubMed]
- Smits, W.K.; Lyras, D.; Lacy, D.B.; Wilcox, M.H.; Kuijper, E.J. Clostridium Difficile Infection. Nat. Rev. Dis. Prim. 2016, 2, 16020. [Google Scholar] [CrossRef]
- McDonald, L.C.; Gerding, D.N.; Johnson, S.; Bakken, J.S.; Carroll, K.C.; Coffin, S.E.; Dubberke, E.R.; Garey, K.W.; Gould, C.V.; Kelly, C.; et al. Clinical Practice Guidelines for Clostridium Difficile Infection in Adults and Children: 2017 Update by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA). Clin. Infect. Dis. 2018, 66, e1–e48. [Google Scholar] [CrossRef] [PubMed]
- Sidler, J.A.; Battegay, M.; Tschudin-Sutter, S.; Widmer, A.F.; Weisser, M. Enterococci, Clostridium Difficile and ESBL-Producing Bacteria: Epidemiology, Clinical Impact and Prevention in ICU Patients. Swiss Med. Wkly. 2014, 144, w14009. [Google Scholar] [CrossRef]
- Vincent, J.L.; Rello, J.; Marshall, J.; Silva, E.; Anzueto, A.; Martin, C.D.; Moreno, R.; Lipman, J.; Gomersall, C.; Sakr, Y.; et al. International Study of the Prevalence and Outcomes of Infection in Intensive Care Units. JAMA 2009, 302, 2323–2329. [Google Scholar] [CrossRef] [PubMed]
- Czepiel, J.; Kędzierska, J.; Biesiada, G.; Birczyńska, M.; Perucki, W.; Nowak, P.; Garlicki, A. Epidemiology of Clostridium Difficile Infection: Results of a Hospital-Based Study in Krakow, Poland. Epidemiol. Infect. 2015, 143, 3235–3243. [Google Scholar] [CrossRef] [PubMed]
- Adelman, M.W.; Woodworth, M.H.; Shaffer, V.O.; Martin, G.S.; Kraft, C.S. Critical Care Management of the Patient with Clostridioides Difficile HHS Public Access. Crit. Care Med. 2021, 49, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Webb, B.J.; Subramanian, A.; Lopansri, B.; Goodman, B.; Jones, P.B.; Ferraro, J.; Stenehjem, E.; Brown, S.M. Antibiotic Exposure and Risk for Hospital-Associated Clostridioides Difficile Infection. Antimicrob. Agents Chemother. 2020, 64, e02169-19. [Google Scholar] [CrossRef]
- Watson, T.; Hickok, J.; Fraker, S.; Korwek, K.; Poland, R.E.; Septimus, E. Evaluating the Risk Factors for Hospital-Onset Clostridium Difficile Infections in a Large Healthcare System. Clin. Infect. Dis. 2018, 66, 1957–1959. [Google Scholar] [CrossRef] [PubMed]
- Anjewierden, S.; Han, Z.; Brown, A.M.; Donskey, C.J.; Deshpande, A. Risk Factors for Clostridioides Difficile Colonization among Hospitalized Adults: A Meta-Analysis and Systematic Review. Infect. Control Hosp. Epidemiol. 2021, 42, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Jukes, C.A.; Ijaz, U.Z.; Buckley, A.; Spencer, J.; Irvine, J.; Candlish, D.; Li, J.V.; Marchesi, J.R.; Douce, G. Bile Salt Metabolism Is Not the Only Factor Contributing to Clostridioides (Clostridium) Difficile Disease Severity in the Murine Model of Disease. Gut Microbes 2020, 11, 481–496. [Google Scholar] [CrossRef]
- Ferreyra, J.A.; Wu, K.J.; Hryckowian, A.J.; Bouley, D.M.; Weimer, B.C.; Sonnenburg, J.L. Gut Microbiota-Produced Succinate Promotes C. Difficile Infection after Antibiotic Treatment or Motility Disturbance. Cell Host Microbe 2014, 16, 770–777. [Google Scholar] [CrossRef]
- Curtis, M.M.; Hu, Z.; Klimko, C.; Narayanan, S.; Deberardinis, R.; Sperandio, V. The Gut Commensal Bacteroides Thetaiotaomicron Exacerbates Enteric Infection through Modification of the Metabolic Landscape. Cell Host Microbe 2014, 16, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.M.; Yalcinkaya, N.; Wu, Q.; Swennes, A.; Tessier, M.E.; Roberts, P.; Miyajima, F.; Savidge, T.; Sorg, J.A. Bile Acid-Independent Protection against Clostridioides Difficile Infection. PLoS Pathog. 2021, 17, e1010015. [Google Scholar] [CrossRef]
- Theriot, C.M.; Bowman, A.A.; Young, V.B. Antibiotic-Induced Alterations of the Gut Microbiota Alter Secondary Bile Acid Production and Allow for Clostridium Difficile Spore Germination and Outgrowth in the Large Intestine. mSphere 2016, 1, e00045-15. [Google Scholar] [CrossRef]
- Spigaglia, P.; Mastrantonio, P.; Barbanti, F. Antibiotic Resistances of Clostridium Difficile. Adv. Exp. Med. Biol. 2018, 1050, 137–159. [Google Scholar]
- Alrahmany, D.; Ereshefsky, B.J.; el Nekidy, W.S.; Harb, G.; Pontiggia, L.; Ghazi, I.M. Risk Factors for Recurrence of Clostridioides Difficile in Hospitalized Patients. J. Infect. Public Health 2021, 14, 1642–1649. [Google Scholar] [CrossRef]
- Van Prehn, J.; Reigadas, E.; Vogelzang, E.H.; Bouza, E.; Hristea, A.; Guery, B.; Krutova, M.; Norén, T.; Allerberger, F.; Coia, J.E.; et al. European Society of Clinical Microbiology and Infectious Diseases: 2021 Update on the Treatment Guidance Document for Clostridioides Difficile Infection in Adults. Clin. Microbiol. Infect. 2021, 27, S1–S21. [Google Scholar] [CrossRef]
- MIC EUCAST. Available online: https://mic.eucast.org/ (accessed on 21 July 2022).
- Freeman, J.; Vernon, J.; Morris, K.; Nicholson, S.; Todhunter, S.; Longshaw, C.; Wilcox, M.H.; Pfeiffer, S.; Delmee, M.; Muytjens, L.; et al. Pan-European Longitudinal Surveillance of Antibiotic Resistance among Prevalent Clostridium Difficile Ribotypes. Clin. Microbiol. Infect. 2015, 21, 248-e9. [Google Scholar] [CrossRef] [PubMed]
- Imwattana, K.; Rodríguez, C.; Riley, T.V.; Knight, D.R. A Species-Wide Genetic Atlas of Antimicrobial Resistance in Clostridioides Difficile. Microb. Genom. 2021, 7, 696. [Google Scholar] [CrossRef]
- Jang, M.-O.; An, J.H.; Jung, S.-I.; Park, K.-H. Refractory Clostridium Difficile Infection Cured With Fecal Microbiota Transplantation in Vancomycin-Resistant Enterococcus Colonized Patient. Intest. Res. 2015, 13, 80–84. [Google Scholar] [CrossRef]
- Fujitani, S.; George, W.L.; Morgan, M.A.; Nichols, S.; Murthy, A.R. Implications for Vancomycin-Resistant Enterococcus Colonization Associated with Clostridium Difficile Infections. Am. J. Infect. Control 2011, 39, 188–193. [Google Scholar] [CrossRef]
- Özsoy, S.; İlki, A. Detection of Vancomycin-Resistant Enterococci (VRE) in Stool Specimens Submitted for Clostridium Difficile Toxin Testing. Braz. J. Microbiol. 2017, 48, 489–492. [Google Scholar] [CrossRef]
- Marra, A.R.; Edmond, M.B.; Wenzel, R.P.; Bearman, G.M.L. Hospital-Acquired Clostridium Difficile-Associated Disease in the Intensive Care Unit Setting: Epidemiology, Clinical Course and Outcome. BMC Infect. Dis. 2007, 7, 42. [Google Scholar] [CrossRef]
- Bauer, M.P.; Notermans, D.W.; van Benthem, B.H.; Brazier, J.S.; Wilcox, M.H.; Rupnik, M.; Monnet, D.L.; van Dissel, J.T.; Kuijper, E.J. Clostridium Difficile Infection in Europe: A Hospital-Based Survey. Lancet 2011, 377, 63–73. [Google Scholar] [CrossRef]
- Rodriguez, C.; Taminiau, B.; van Broeck, J.; Delmée, M.; Daube, G. Clostridium Difficile in Food and Animals: A Comprehensive Review. Adv. Exp. Med. Biol. 2016, 932, 65–92. [Google Scholar]
- Jhung, M.A.; Thompson, A.D.; Killgore, G.E.; Zukowski, W.E.; Songer, G.; Warny, M.; Johnson, S.; Gerding, D.N.; McDonald, L.C.; Limbago, B.M. Toxinotype V Clostridium Difficile in Humans and Food Animals. Emerg. Infect. Dis. 2008, 14, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Goorhuis, A.; Bakker, D.; Corver, J.; Debast, S.B.; Harmanus, C.; Notermans, D.W.; Bergwerff, A.A.; Dekker, F.W.; Kuijper, E.J. Emergence of Clostridium Difficile Infection Due to a New Hypervirulent Strain, Polymerase Chain Reaction Ribotype 078. Clin. Infect. Dis. 2008, 47, 1162–1170. [Google Scholar] [CrossRef]
- Freeman, D.M. Invertible Carnot Groups. Anal. Geom. Metr. Spaces 2014, 2, 248–257. [Google Scholar] [CrossRef]
- Knetsch, C.W.; Connor, T.R.; Mutreja, A.; van Dorp, S.M.; Sanders, I.M.; Browne, H.P.; Harris, D.; Lipman, L.; Keessen, E.C.; Corver, J.; et al. Whole Genome Sequencing Reveals Potential Spread of Clostridium Difficile between Humans and Farm Animals in the Netherlands, 2002 to 2011. Eurosurveillance 2014, 19, 20954. [Google Scholar] [CrossRef] [PubMed]
- Tramuta, C.; Spigaglia, P.; Barbanti, F.; Bianchi, D.M.; Boteva, C.; di Blasio, A.; Zoppi, S.; Zaccaria, T.; Proroga, Y.T.R.; Chiavacci, L.; et al. Comparison of Clostridioides Difficile Strains from Animals and Humans: First Results after Introduction of C. Difficile Molecular Typing and Characterization at the Istituto Zooprofilattico Sperimentale of Piemonte, Liguria e Valle d’Aosta, Italy. Comp. Immunol. Microbiol. Infect. Dis. 2021, 75, 101623. [Google Scholar] [CrossRef] [PubMed]
- Marcos, P.; Whyte, P.; Rogers, T.; McElroy, M.; Fanning, S.; Frias, J.; Bolton, D. The Prevalence of Clostridioides Difficile on Farms, in Abattoirs and in Retail Foods in Ireland. Food Microbiol. 2021, 98, 103781. [Google Scholar] [CrossRef]
- Braun, V.; Hundsberger, T.; Leukel, P.; Sauerborn, M.; von Eichel-Streiber, C. Definition of the Single Integration Site of the Pathogenicity Locus in Clostridium Difficile. Gene 1996, 181, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Martin-Verstraete, I.; Peltier, J.; Dupuy, B. The Regulatory Networks That Control Clostridium Difficile Toxin Synthesis. Toxins 2016, 8, 153. [Google Scholar] [CrossRef]
- Stare, B.G.; Delmée, M.; Rupnik, M. Variant Forms of the Binary Toxin CDT Locus and TcdC Gene in Clostridium Difficile Strains. J. Med. Microbiol. 2007, 56, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Carter, G.P.; Lyras, D.; Allen, D.L.; Mackin, K.E.; Howarth, P.M.; O’Connor, J.R.; Rood, J.I. Binary Toxin Production in Clostridium Difficile Is Regulated by CdtR, a LytTR Family Response Regulator. J. Bacteriol. 2007, 189, 7290. [Google Scholar] [CrossRef]
- Maslanka, J.R.; Gu, C.H.; Zarin, I.; Denny, J.E.; Broadaway, S.; Fett, B.; Mattei, L.M.; Walk, S.T.; Abt, M.C. Detection and Elimination of a Novel Non-Toxigenic Clostridioides Difficile Strain from the Microbiota of a Mouse Colony. Gut Microbes 2020, 12, 1851999. [Google Scholar] [CrossRef]
- Monot, M.; Eckert, C.; Lemire, A.; Hamiot, A.; Dubois, T.; Tessier, C.; Dumoulard, B.; Hamel, B.; Petit, A.; Lalande, V.; et al. Clostridium Difficile: New Insights into the Evolution of the Pathogenicity Locus. Sci. Rep. 2015, 5, 15023. [Google Scholar] [CrossRef]
- Dingle, K.E.; Elliott, B.; Robinson, E.; Griffiths, D.; Eyre, D.W.; Stoesser, N.; Vaughan, A.; Golubchik, T.; Fawley, W.N.; Wilcox, M.H.; et al. Evolutionary History of the Clostridium Difficile Pathogenicity Locus. Genome Biol. Evol. 2014, 6, 36–52. [Google Scholar] [CrossRef]
- Janezic, S.; Dingle, K.; Alvin, J.; Accetto, T.; Didelot, X.; Crook, D.W.; Borden Lacy, D.; Rupnik, M. Comparative Genomics of Clostridioides Difficile Toxinotypes Identifies Module-Based Toxin Gene Evolution. Microb. Genom. 2020, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, M.J.; Tremblay, B.J.M.; Zeng, J.; Wei, X.; Hodgins, H.; Worley, J.; Bry, L.; Dong, M.; Doxey, A.C. Phylogenomics of 8,839 Clostridioides Difficile Genomes Reveals Recombination-Driven Evolution and Diversification of Toxin A and B. PLoS Pathog. 2020, 16, e1009181. [Google Scholar] [CrossRef] [PubMed]
- Steele, J.; Mukherjee, J.; Parry, N.; Tzipori, S. Antibody Against TcdB, but Not TcdA, Prevents Development of Gastrointestinal and Systemic Clostridium Difficile Disease. J. Infect. Dis. 2013, 207, 323–330. [Google Scholar] [CrossRef]
- Gupta, S.B.; Mehta, V.; Dubberke, E.R.; Zhao, X.; Dorr, M.B.; Guris, D.; Molrine, D.; Leney, M.; Miller, M.; Dupin, M.; et al. Antibodies to Toxin B Are Protective Against Clostridium Difficile Infection Recurrence. Clin. Infect. Dis. 2016, 63, 730–734. [Google Scholar] [CrossRef]
- Rounds, J.; Strain, J. Bezlotoxumab for Preventing Recurrent Clostridium Difficile Infections. J. South Dak. State Med. Assoc. 2017, 70, 422–423. [Google Scholar]
- Knight, D.R.; Imwattana, K.; Kullin, B.; Guerrero-Araya, E.; Paredes-Sabja, D.; Didelot, X.; Dingle, K.E.; Eyre, D.W.; Rodríguez, C.; Riley, T.V. Major Genetic Discontinuity and Novel Toxigenic Species in Clostridioides Difficile Taxonomy. Elife 2021, 10, e64325. [Google Scholar] [CrossRef]
- Imwattana, K.; Knight, D.R.; Kullin, B.; Collins, D.A.; Putsathit, P.; Kiratisin, P.; Riley, T.V. Clostridium Difficile Ribotype 017—Characterization, Evolution and Epidemiology of the Dominant Strain in Asia. Emerg. Microbes Infect. 2019, 8, 796–807. [Google Scholar] [CrossRef]
- Knight, D.R.; Kullin, B.; Androga, G.O.; Barbut, F.; Eckert, C.; Johnson, S.; Spigaglia, P.; Tateda, K.; Tsai, P.J.; Riley, T.V. Evolutionary and Genomic Insights into Clostridioides Difficile Sequence Type 11: A Diverse Zoonotic and Antimicrobial-Resistant Lineage of Global One Health Importance. mBio 2019, 10, e00446-19. [Google Scholar] [CrossRef] [PubMed]
- Didelot, X.; Eyre, D.W.; Cule, M.; Ip, C.L.C.; Ansari, M.A.; Griffiths, D.; Vaughan, A.; O’Connor, L.; Golubchik, T.; Batty, E.M.; et al. Microevolutionary Analysis of Clostridium Difficile Genomes to Investigate Transmission. Genome Biol. 2012, 13, R118. [Google Scholar] [CrossRef] [PubMed]
- Janezic, S.; Potocnik, M.; Zidaric, V.; Rupnik, M. Highly Divergent Clostridium Difficile Strains Isolated from the Environment. PLoS ONE 2016, 11, e0167101. [Google Scholar] [CrossRef]
- Ramírez-Vargas, G.; López-Ureña, D.; Badilla, A.; Orozco-Aguilar, J.; Murillo, T.; Rojas, P.; Riedel, T.; Overmann, J.; González, G.; Chaves-Olarte, E.; et al. Novel Clade C-I Clostridium Difficile Strains Escape Diagnostic Tests, Differ in Pathogenicity Potential and Carry Toxins on Extrachromosomal Elements. Sci. Rep. 2018, 8, 13951. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Vargas, G.; Rodriguez, C. Putative Conjugative Plasmids with TcdB and CdtAB Genes in Clostridioides Difficile. Emerg. Infect. Dis. 2020, 26, 2287. [Google Scholar] [CrossRef]
- Dingle, K.E.; Didelot, X.; Quan, T.P.; Eyre, D.W.; Stoesser, N.; Marwick, C.A.; Coia, J.; Brown, D.; Buchanan, S.; Ijaz, U.Z.; et al. A Role for Tetracycline Selection in Recent Evolution of Agriculture-Associated Clostridium Difficile PCR Ribotype 078. MBio 2019, 10, e02790-18. [Google Scholar] [CrossRef]
- Kuijper, E.; Weerdt, J.; Kato, H.; Kato, N.; Dam, A.; Vorm, E.; Weel, J.; Rheenen, C.; Dankert, J. Nosocomial Outbreak of Clostridium Difficile-Associated Diarrhoea Due to a Clindamycin-Resistant Enterotoxin A-Negative Strain. Eur. J. Clin. Microbiol. Infect. Dis. 2001, 20, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Chaar, A.; Feuerstadt, P. Evolution of Clinical Guidelines for Antimicrobial Management of Clostridioides Difficile Infection. Ther. Adv. Gastroenterol. 2021, 14, 17562848211011953. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.J.; Deshpande, A.; Hevener, K.E.; Endres, B.T.; Garey, K.W.; Palmer, K.L.; Hurdle, J.G. Constitutive Expression of the Cryptic VanGCd Operon Promotes Vancomycin Resistance in Clostridioides Difficile Clinical Isolates. J. Antimicrob. Chemother. 2020, 75, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Boekhoud, I.M.; Sidorov, I.; Nooij, S.; Harmanus, C.; Bos-Sanders, I.M.J.G.; Viprey, V.; Spittal, W.; Clark, E.; Davies, K.; Freeman, J.; et al. Haem Is Crucial for Medium-Dependent Metronidazole Resistance in Clinical Isolates of Clostridioides Difficile. J. Antimicrob. Chemother. 2021, 76, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Shen, W.J.; Deshpande, A.; Olaitan, A.O.; Palmer, K.L.; Garey, K.W.; Hurdle, J.G. The Integrity of Heme Is Essential for Reproducible Detection of Metronidazole-Resistant Clostridioides Difficile by Agar Dilution Susceptibility Tests. J. Clin. Microbiol. 2021, 59, e0058521. [Google Scholar] [CrossRef]
- Giardina, G.; Rinaldo, S.; Johnson, K.A.; di Matteo, A.; Brunori, M.; Cutruzzolà, F. NO Sensing in Pseudomonas Aeruginosa: Structure of the Transcriptional Regulator DNR. J. Mol. Biol. 2008, 378, 1002–1015. [Google Scholar] [CrossRef]
- Boekhoud, I.M.; Hornung, B.V.H.; Sevilla, E.; Harmanus, C.; Bos-Sanders, I.M.J.G.; Terveer, E.M.; Bolea, R.; Corver, J.; Kuijper, E.J.; Smits, W.K. Plasmid-Mediated Metronidazole Resistance in Clostridioides Difficile. Nat. Commun. 2020, 11, 598. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, A.; Wu, X.; Huo, W.; Palme, K.L.; Hurdl, J.G. Chromosomal Resistance to Metronidazole in Clostridioides Difficile Can Be Mediated by Epistasis between Iron Homeostasis and Oxidoreductases. Antimicrob. Agents Chemother. 2020, 64, e00415-20. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Das, K.; Degen, D.; Mazumder, A.; Duchi, D.; Wang, D.; Ebright, Y.W.; Ebright, R.Y.; Sineva, E.; Gigliotti, M.; et al. Structural Basis of Transcription Inhibition by Fidaxomicin (Lipiarmycin A3). Mol. Cell 2018, 70, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Babakhani, F.; Seddon, J.; Sears, P. Comparative Microbiological Studies of Transcription Inhibitors Fidaxomicin and the Rifamycins in Clostridium Difficile. Antimicrob. Agents Chemother. 2014, 58, 2934–2937. [Google Scholar] [CrossRef] [PubMed]
- Babakhani, F.; Gomez, A.; Robert, N.; Sears, P. Killing Kinetics of Fidaxomicin and Its Major Metabolite, OP-1118, against Clostridium Difficile. J. Med. Microbiol. 2011, 60, 1213–1217. [Google Scholar] [CrossRef]
- Johnson, S.; Lavergne, V.; Skinner, A.M.; Gonzales-Luna, A.J.; Garey, K.W.; Kelly, C.P.; Wilcox, M.H. Clinical Practice Guideline by the Infectious Diseases Society of America (IDSA) and Society for Healthcare Epidemiology of America (SHEA): 2021 Focused Update Guidelines on Management of Clostridioides Difficile Infection in Adults. Clin. Infect. Dis. 2021, 73, 755–757. [Google Scholar] [CrossRef]
- Peng, Z.; Jin, D.; Kim, H.B.; Stratton, C.W.; Wu, B.; Tang, Y.W.; Suna, X. Update on Antimicrobial Resistance in Clostridium Difficile: Resistance Mechanisms and Antimicrobial Susceptibility Testing. J. Clin. Microbiol. 2017, 55, 1998. [Google Scholar] [CrossRef]
- Baines, S.D.; Wilcox, M.H. Antimicrobial Resistance and Reduced Susceptibility in Clostridium Difficile: Potential Consequences for Induction, Treatment, and Recurrence of C. Difficile Infection. Antibiotics 2015, 4, 267–298. [Google Scholar] [CrossRef] [PubMed]
- Carlson, T.J.; Gonzales-Luna, A.J. Antibiotic Treatment Pipeline for Clostridioides Difficile Infection (CDI): A Wide Array of Narrow-Spectrum Agents. Curr. Infect. Dis. Rep. 2020, 22, 20. [Google Scholar] [CrossRef]
- Sun, J.; Chen, C.; Cui, C.Y.; Zhang, Y.; Liu, X.; Cui, Z.H.; Ma, X.Y.; Feng, Y.; Fang, L.X.; Lian, X.L.; et al. Plasmid-Encoded Tet(X) Genes That Confer High-Level Tigecycline Resistance in Escherichia Coli. Nat. Microbiol. 2019, 4, 1457–1464. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Wang, R.; Liu, D.; Walsh, T.R.; Zhang, R.; Lv, Y.; Ke, Y.; Ji, Q.; Wei, R.; Liu, Z.; et al. Emergence of Plasmid-Mediated High-Level Tigecycline Resistance Genes in Animals and Humans. Nat. Microbiol. 2019, 4, 1450–1456. [Google Scholar] [CrossRef]
- Collins, D.A.; Elliott, B.; Riley, T.V. Molecular Methods for Detecting and Typing of Clostridium Difficile. Pathology 2015, 47, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Krutova, M.; Kinross, P.; Barbut, F.; Hajdu, A.; Wilcox, M.H.; Kuijper, E.J.; Allerberger, F.; Delmée, M.; van Broeck, J.; Vatcheva-Dobrevska, R.; et al. How to: Surveillance of Clostridium Difficile Infections. Clin. Microbiol. Infect. 2018, 24, 469–475. [Google Scholar] [CrossRef]
- Seth-Smith, H.M.B.; Biggel, M.; Roloff, T.; Hinic, V.; Bodmer, T.; Risch, M.; Casanova, C.; Widmer, A.; Sommerstein, R.; Marschall, J.; et al. Transition From PCR-Ribotyping to Whole Genome Sequencing Based Typing of Clostridioides Difficile. Front. Cell Infect. Microbiol. 2021, 11, 681518. [Google Scholar] [CrossRef]
- Neoh, H.M.; Tan, X.E.; Sapri, H.F.; Tan, T.L. Pulsed-Field Gel Electrophoresis (PFGE): A Review of the “Gold Standard” for Bacteria Typing and Current Alternatives. Infect. Genet. Evol. 2019, 74, 103935. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.; Martín, A.; Peláez, T.; Marín, M.; Rodríguez-Creixéms, M.; Bouza, E. An Improved Protocol for Pulsed-Field Gel Electrophoresis Typing of Clostridium Difficile. J. Med. Microbiol. 2005, 54, 155–157. [Google Scholar] [CrossRef]
- Pérez-Losada, M.; Cabezas, P.; Castro-Nallar, E.; Crandall, K.A. Pathogen Typing in the Genomics Era: MLST and the Future of Molecular Epidemiology. Infect. Genet. Evol. 2013, 16, 38–53. [Google Scholar] [CrossRef]
- PubMLST. Clostridioides Difficile. Available online: https://pubmlst.org/organisms/clostridioides-difficile (accessed on 23 September 2022).
- Griffiths, D.; Fawley, W.; Kachrimanidou, M.; Bowden, R.; Crook, D.W.; Fung, R.; Golubchik, T.; Harding, R.M.; Jeffery, K.J.M.; Jolley, K.A.; et al. Multilocus Sequence Typing of Clostridium Difficile. J. Clin. Microbiol. 2010, 48, 770–778. [Google Scholar] [CrossRef]
- Janezic, S.; Rupnik, M. Development and Implementation of Whole Genome Sequencing-Based Typing Schemes for Clostridioides Difficile. Front. Public Health 2019, 7, 309. [Google Scholar] [CrossRef]
- Eyre, D.W.; Golubchik, T.; Gordon, N.C.; Bowden, R.; Piazza, P.; Batty, E.M.; Ip, C.L.C.; Wilson, D.J.; Didelot, X.; O’Connor, L.; et al. A Pilot Study of Rapid Benchtop Sequencing of Staphylococcus Aureus and Clostridium Difficile for Outbreak Detection and Surveillance. BMJ Open 2012, 2, e001124. [Google Scholar] [CrossRef]
- Freeman, J.; Vernon, J.; Pilling, S.; Morris, K.; Nicolson, S.; Shearman, S.; Clark, E.; Palacios-Fabrega, J.A.; Wilcox, M. Five-Year Pan-European, Longitudinal Surveillance of Clostridium Difficile Ribotype Prevalence and Antimicrobial Resistance: The Extended ClosER Study. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 169–177. [Google Scholar] [CrossRef]
- Snydman, D.R.; McDermott, L.A.; Jenkins, S.G.; Goldstein, E.J.C.; Patel, R.; Forbes, B.A.; Johnson, S.; Gerding, D.N.; Thorpe, C.M.; Walk, S.T. Epidemiologic Trends in Clostridioides Difficile Isolate Ribotypes in United States from 2011 to 2016. Anaerobe 2020, 63, 102185. [Google Scholar] [CrossRef]
- Lew, T.; Putsathit, P.; Sohn, K.M.; Wu, Y.; Ouchi, K.; Ishii, Y.; Tated, K.; Riley, T.V.; Collins, D.A. Antimicrobial Susceptibilities of Clostridium Difficile Isolates from 12 Asia-Pacific Countries in 2014 and 2015. Antimicrob. Agents Chemother. 2020, 64, e00296-20. [Google Scholar] [CrossRef] [PubMed]
- Secco, D.A.; Balassiano, I.T.; Boente, R.F.; Miranda, K.R.; Brazier, J.; Hall, V.; dos Santos-Filho, J.; Lobo, L.A.; Nouér, S.A.; Domingues, R.M.C.P. Clostridium Difficile Infection among Immunocompromised Patients in Rio de Janeiro, Brazil and Detection of Moxifloxacin Resistance in a Ribotype 014 Strain. Anaerobe 2014, 28, 85–89. [Google Scholar] [CrossRef]
- Sofjan, A.K.; Islam, M.A.; Halder, K.; Kabir, N.D.; Saleh, A.A.; Miranda, J.; Lancaster, C.; Begum, K.; Alam, M.J.; Garey, K.W. Molecular Epidemiology of Toxigenic Clostridioides Difficile Isolates from Hospitalized Patients and the Hospital Environment in Dhaka, Bangladesh. Anaerobe 2020, 61, 102081. [Google Scholar] [CrossRef] [PubMed]
- Perumalsamy, S.; Lim, S.C.; Riley, T.V. Clostridioides (Clostridium) Difficile Isolated from Paediatric Patients in Western Australia 2019–2020. Pathology 2022, 54, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Djebbar, A.; Sebaihia, M.; Kuijper, E.; Harmanus, C.; Sanders, I.; Benbraham, N.; Hacène, H. First Molecular Characterisation and PCR Ribotyping of Clostridium Difficile Strains Isolated in Two Algerian Hospitals. J. Infect. Dev. Ctries. 2018, 12, 15–21. [Google Scholar] [CrossRef]
- Seugendo, M.; Mshana, S.E.; Hokororo, A.; Okamo, B.; Mirambo, M.M.; von Müller, L.; Gunka, K.; Zimmermann, O.; Groß, U. Clostridium Difficile Infections among Adults and Children in Mwanza/Tanzania: Is It an Underappreciated Pathogen among Immunocompromised Patients in Sub-Saharan Africa? New Microbes New Infect. 2015, 8, 99–102. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhao, H.; Nickle, D.C.; Zeng, Z.; Law, P.Y.T.; Wilcox, M.H.; Chen, L.; Peng, Y.; Meng, J.; Deng, Z.; Albright, A.; et al. Global Landscape of Clostridioides Difficile Phylogeography, Antibiotic Susceptibility, and Toxin Polymorphisms by Post-Hoc Whole-Genome Sequencing from the MODIFY I/II Studies. Infect. Dis. Ther. 2021, 10, 853–870. [Google Scholar] [CrossRef] [PubMed]
- Abdrabou, A.M.M.; Ul Habib Bajwa, Z.; Halfmann, A.; Mellmann, A.; Nimmesgern, A.; Margardt, L.; Bischoff, M.; von Müller, L.; Gärtner, B.; Berger, F.K. Molecular Epidemiology and Antimicrobial Resistance of Clostridioides Difficile in Germany, 2014–2019. Int. J. Med. Microbiol. 2021, 311, 151507. [Google Scholar] [CrossRef]
- Abdrabou, A.M.M.; Bischoff, M.; Mellmann, A.; von Müller, L.; Margardt, L.; Gärtner, B.C.; Berger, F.K.; Haase, G.; Häfner, H.; Hoffmann, R.; et al. Implementation of a Clostridioides Difficile Sentinel Surveillance System in Germany: First Insights for 2019–2021. Anaerobe 2022, 77, 102548. [Google Scholar] [CrossRef]
- Johnson, S.; Citron, D.M.; Gerding, D.N.; Wilcox, M.H.; Goldstein, E.J.C.; Sambol, S.P.; Best, E.L.; Eves, K.; Jensen, E.; Dorr, M.B. Efficacy of Bezlotoxumab in Trial Participants Infected With Clostridioides Difficile Strain BI Associated With Poor Outcomes. Clin. Infect. Dis. 2021, 73, e2616–e2624. [Google Scholar] [CrossRef] [PubMed]
- Romano, V.; Pasquale, V.; Krovacek, K.; Mauri, F.; Demarta, A.; Dumontet, S. Toxigenic Clostridium Difficile PCR Ribotypes from Wastewater Treatment Plants in Southern Switzerland. Appl. Environ. Microbiol. 2012, 78, 6643. [Google Scholar] [CrossRef]
- Skinner, A.M.; Petrella, L.; Siddiqui, F.; Sambol, S.P.; Gulvik, C.A.; Gerding, D.N.; Donskey, C.J.; Johnson, S. Unique Clindamycin-Resistant Clostridioides Difficile Strain Related to Fluoroquinolone-Resistant Epidemic BI/RT027 Strain. Emerg. Infect. Dis. 2020, 26, 247–254. [Google Scholar] [CrossRef]
- FDA. FDA Updates Warnings for Fluoroquinolone Antibiotics. Available online: https://www.fda.gov/news-events/press-announcements/fda-updates-warnings-fluoroquinolone-antibiotics (accessed on 9 November 2022).
- European Medicines Agency. Fluoroquinolone and Quinolone Antibiotics: PRAC Recommends New Restrictions on Use Following Review of Disabling and Potentially Long-Lasting Side Effects. Available online: https://www.ema.europa.eu/en/news/fluoroquinolone-quinolone-antibiotics-prac-recommends-new-restrictions-use-following-review (accessed on 9 November 2022).
- Wieczorkiewicz, J.T.; Lopansri, B.K.; Cheknis, A.; Osmolski, J.R.; Hecht, D.W.; Gerding, D.N.; Johnson, S. Fluoroquinolone and Macrolide Exposure Predict Clostridium Difficile Infection with the Highly Fluoroquinolone- and Macrolide-Resistant Epidemic C. Difficile Strain BI/NAP1/027. Antimicrob. Agents Chemother. 2016, 60, 418–423. [Google Scholar] [CrossRef]
- Tabak, Y.P.; Srinivasan, A.; Yu, K.C.; Kurtz, S.G.; Gupta, V.; Gelone, S.; Scoble, P.J.; Mcdonald, L.C. Hospital-Level High-Risk Antibiotic Use in Relation to Hospital-Associated Clostridioides Difficile Infections: Retrospective Analysis of 2016-2017 Data from US Hospitals HHS Public Access. Infect. Control Hosp. Epidemiol. 2019, 40, 1229–1235. [Google Scholar] [CrossRef]
- Berger, F.K.; Rasheed, S.S.; Araj, G.F.; Mahfouz, R.; Rimmani, H.H.; Karaoui, W.R.; Sharara, A.I.; Dbaibo, G.; Becker, S.L.; von Müller, L.; et al. Molecular Characterization, Toxin Detection and Resistance Testing of Human Clinical Clostridium Difficile Isolates from Lebanon. Int. J. Med. Microbiol. 2018, 308, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.A.; Ashwin, H.; Longshaw, C.M.; Burns, D.A.; Davis, G.L.; Wilcox, M.H. Diversity of Clostridium Difficile PCR Ribotypes in Europe: Results from the European, Multicentre, Prospective, Biannual, Point-Prevalence Study of Clostridium Difficile Infection in Hospitalised Patients with Diarrhoea (EUCLID), 2012 and 2013. Eurosurveillance 2016, 21, 30294. [Google Scholar] [CrossRef] [PubMed]
- Tkalec, V.; Janezic, S.; Skok, B.; Simonic, T.; Mesaric, S.; Vrabic, T.; Rupnik, M. High Clostridium Difficile Contamination Rates of Domestic and Imported Potatoes Compared to Some Other Vegetables in Slovenia. Food Microbiol. 2019, 78, 194–200. [Google Scholar] [CrossRef]
- Lim, S.C.; Foster, N.F.; Elliott, B.; Riley, T.V. High Prevalence of Clostridium Difficile on Retail Root Vegetables, Western Australia. J. Appl. Microbiol. 2018, 124, 585–590. [Google Scholar] [CrossRef]
- Freeman, J.; Vernon, J.; Pilling, S.; Morris, K.; Nicholson, S.; Shearman, S.; Longshaw, C.; Wilcox, M.H. The ClosER Study: Results from a Three-Year Pan-European Longitudinal Surveillance of Antibiotic Resistance among Prevalent Clostridium Difficile Ribotypes, 2011–2014. Clin. Microbiol. Infect. 2018, 24, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Cheknis, A.; Johnson, S.; Chesnel, L.; Petrella, L.; Sambol, S.; Dale, S.E.; Nary, J.; Sears, P.; Citron, D.M.; Goldstein, E.J.C.; et al. Molecular Epidemiology of Clostridioides (Clostridium) Difficile Strains Recovered from Clinical Trials in the US, Canada and Europe from 2006–2009 to 2012–2015. Anaerobe 2018, 53, 38–42. [Google Scholar] [CrossRef]
- Eyre, D.W.; Davies, K.A.; Davis, G.; Fawley, W.N.; Dingle, K.E.; de Maio, N.; Karas, A.; Crook, D.W.; Peto, T.E.A.; Walker, A.S.; et al. Two Distinct Patterns of Clostridium Difficile Diversity Across Europe Indicating Contrasting Routes of Spread. Clin. Infect. Dis. 2018, 67, 1035–1044. [Google Scholar] [CrossRef]
- Piepenbrock, E.; Stelzer, Y.; Berger, F.; Jazmati, N. Changes in Clostridium (Clostridioides) Difficile PCR-Ribotype Distribution and Antimicrobial Resistance in a German Tertiary Care Hospital Over the Last 10 Years. Curr. Microbiol. 2019, 76, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-Luna, A.J.; Carlson, T.J.; Dotson, K.M.; Poblete, K.; Costa, G.; Miranda, J.; Lancaster, C.; Walk, S.T.; Tupy, S.; Begum, K.; et al. PCR Ribotypes of Clostridioides Difficile across Texas from 2011 to 2018 Including Emergence of Ribotype 255. Emerg. Microbes Infect. 2020, 9, 341–347. [Google Scholar] [CrossRef]
- Spinler, J.K.; Gonzales-Luna, A.J.; Raza, S.; Runge, J.K.; Luna, R.A.; Savidge, T.C.; Garey, K.W. Complete Genome Sequence of Clostridioides Difficile Ribotype 255 Strain Mta-79, Assembled Using Oxford Nanopore and Illumina Sequencing. Microbiol. Resour. Announc. 2019, 8, e00935-19. [Google Scholar] [CrossRef]
- O’shaughnessy, R.A.; Habing, G.G.; Gebreyes, W.A.; Bowman, A.S.; Weese, J.S.; Rousseau, J.; Stull, J.W. Clostridioides Difficile on Ohio Swine Farms (2015): A Comparison of Swine and Human Environments and Assessment of on-Farm Risk Factors. Zoonoses Public Health 2019, 66, 861–870. [Google Scholar] [CrossRef]
- Fawley, W.N.; Davies, K.A.; Morris, T.; Parnell, P.; Howe, R.; Wilcox, M.H. Enhanced Surveillance of Clostridium Difficile Infection Occurring Outside Hospital, England, 2011 to 2013. Eurosurveillance 2016, 21, 30295. [Google Scholar] [CrossRef]
- Cheng, A.C.; Collins, D.A.; Elliott, B.; Ferguson, J.K.; Paterson, D.L.; Thean, S.; Riley, T.V. Laboratory-Based Surveillance of Clostridium Difficile Circulating in Australia, September–November 2010. Pathology 2016, 48, 257–260. [Google Scholar] [CrossRef]
- Williamson, C.H.D.; Stone, N.E.; Nunnally, A.E.; Roe, C.C.; Vazquez, A.J.; Lucero, S.A.; Hornstra, H.; Wagner, D.M.; Keim, P.; Rupnik, M.; et al. Identification of Novel, Cryptic Clostridioides Species Isolates from Environmental Samples Collected from Diverse Geographical Locations. Microb. Genom. 2022, 8, 000742. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, G.Z.; Pires, R.N.; Rauber, A.P.; de Lima-Morales, D.; Falci, D.R.; Caierão, J.; Pasqualotto, A.C.; Martins, A.F. Genetic Relatedness, Virulence Factors and Antimicrobial Resistance of C. Difficile Strains from Hospitalized Patients in a Multicentric Study in Brazil. J. Glob. Antimicrob Resist. 2020, 22, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Balassiano, I.T.; Miranda, K.R.; Boente, R.F.; Pauer, H.; Oliveira, I.C.M.; Santos-Filho, J.; Amorim, E.L.T.; Caniné, G.A.; Souza, C.F.; Gomes, M.Z.R.; et al. Characterization of Clostridium Difficile Strains Isolated from Immunosuppressed Inpatients in a Hospital in Rio de Janeiro, Brazil. Anaerobe 2009, 15, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Alcides, A.P.P.; Brazier, J.S.; Pinto, L.J.F.; Balassiano, I.T.; Boente, R.F.; Paula, G.R.; Ferreira, E.O.; Avelar, K.E.S.; Miranda, K.R.; Ferreira, M.C.S.; et al. New PCR Ribotypes of Clostridium Difficile Detected in Children in Brazil. Antonie Van Leeuwenhoek 2007, 92, 53–59. [Google Scholar] [CrossRef]
- Carneiro, L.G.; Pinto, T.C.A.; Moura, H.; Barr, J.; Domingues, R.M.C.P.; Ferreira, E.d.O. MALDI-TOF MS: An Alternative Approach for Ribotyping Clostridioides Difficile Isolates in Brazil. Anaerobe 2021, 69, 102351. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, R.; Sharma, N.; Bahadur, T.; Khullar, S.; Agarwal, S.K.; Gahlowt, A.; Gupta, N.; Kumar, L.; Kabra, S.K.; Dey, A.B. Molecular Characterization of Clostridioides Difficile by Multi-Locus Sequence Typing (MLST): A Study from Tertiary Care Center in India. Anaerobe 2022, 75, 102545. [Google Scholar] [CrossRef]
- Collins, D.A.; Sohn, K.M.; Wu, Y.; Ouchi, K.; Ishii, Y.; Elliott, B.; Riley, T.V.; Tateda, K. Clostridioides Difficile Infection in the Asia-Pacific Region. Emerg. Microbes Infect. 2020, 9, 42–52. [Google Scholar] [CrossRef]
- Collins, D.A.; Hawkey, P.M.; Riley, T.V. Epidemiology of Clostridium Difficile Infection in Asia. Antimicrob. Resist. Infect. Control 2013, 2, 21. [Google Scholar] [CrossRef]
- Collins, D.A.; Riley, T.V. Clostridium Difficile Guidelines. Clin. Infect. Dis. 2018, 67, 1639. [Google Scholar] [CrossRef]
- Kato, H.; Kato, H.; Ito, Y.; Akahane, T.; Izumida, S.; Yokoyama, T.; Kaji, C.; Arakawa, Y. Typing of Clostridium Difficile Isolates Endemic in Japan by Sequencing of SlpA and Its Application to Direct Typing. J. Med. Microbiol. 2010, 59, 556–562. [Google Scholar] [CrossRef]
- Senoh, M.; Kato, H.; Fukuda, T.; Niikawa, A.; Hori, Y.; Hagiya, H.; Ito, Y.; Miki, H.; Abe, Y.; Furuta, K.; et al. Predominance of PCR-Ribotypes, 018 (Smz) and 369 (Trf) of Clostridium Difficile in Japan: A Potential Relationship with Other Global Circulating Strains? J. Med. Microbiol. 2015, 64, 1226–1236. [Google Scholar] [CrossRef]
- Kuwata, Y.; Tanimoto, S.; Sawabe, E.; Shima, M.; Takahashi, Y.; Ushizawa, H.; Fujie, T.; Koike, R.; Tojo, N.; Kubota, T.; et al. Molecular Epidemiology and Antimicrobial Susceptibility of Clostridium Difficile Isolated from a University Teaching Hospital in Japan. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 763–772. [Google Scholar] [CrossRef]
- Aoki, K.; Takeda, S.; Miki, T.; Ishii, Y.; Tateda, K. Antimicrobial Susceptibility and Molecular Characterization Using Whole-Genome Sequencing of Clostridioides Difficile Collected in 82 Hospitals in Japan between 2014 and 2016. Antimicrob. Agents Chemother. 2019, 63, e01259-19. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Luo, Y.; Huang, C.; Cai, J.; Ye, J.; Zheng, Y.; Wang, L.; Zhao, P.; Liu, A.; Fang, W.; et al. Molecular Epidemiology of Clostridium Difficile Infection in Hospitalized Patients in Eastern China. J. Clin. Microbiol. 2017, 55, 801–810. [Google Scholar] [CrossRef]
- Tang, C.; Cui, L.; Xu, Y.; Xie, L.; Sun, P.; Liu, C.; Xia, W.; Liu, G. The Incidence and Drug Resistance of Clostridium Difficile Infection in Mainland China: A Systematic Review and Meta-Analysis. Sci. Rep. 2016, 6, 37865. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, W.; Xiao, T.; Chen, Y.; Lv, T.; Wang, Y.; Zhang, S.; Cai, H.; Chi, X.; Kong, X.; et al. Comparative Genomic and Transmission Analysis of Clostridioides Difficile between Environmental, Animal, and Clinical Sources in China. Emerg. Microbes Infect. 2021, 10, 2244–2255. [Google Scholar] [CrossRef]
- Zhang, W.Z.; Li, W.G.; Liu, Y.Q.; Gu, W.P.; Zhang, Q.; Li, H.; Liu, Z.J.; Zhang, X.; Wu, Y.; Lu, J.X. The Molecular Characters and Antibiotic Resistance of Clostridioides Difficile from Economic Animals in China. BMC Microbiol. 2020, 20, 70. [Google Scholar] [CrossRef] [PubMed]
- Baghani, A.; Mesdaghinia, A.; Kuijper, E.J.; Aliramezani, A.; Talebi, M.; Douraghi, M. High Prevalence of Clostridiodes Diffiicle PCR Ribotypes 001 and 126 in Iran. Sci. Rep. 2020, 10, 4658. [Google Scholar] [CrossRef]
- Sharara, A.; Karaoui, W.R.; Shayto, R.H.; Rimmani, H.; Chalhoub, J.M.; Zahreddine, N.; el Sabbagh, R.; Kanj, S.S.; Mahfouz, R.; Araj, G.; et al. Hypervirulent Clostridium Difficile Strains Are Rarely Associated With Nosocomial CDI in Adults in Lebanon: Results From a Prospective Study on the Incidence, Risk Factors for Relapse, and Outcome of Nosocomial CDI. Am. J. Gastroenterol. 2017, 112, S51. [Google Scholar] [CrossRef]
- Jamal, W.; Pauline, E.; Rotimi, V. A Prospective Study of Community-Associated Clostridium Difficile Infection in Kuwait: Epidemiology and Ribotypes. Anaerobe 2015, 35, 28–32. [Google Scholar] [CrossRef]
- Al-Thani, A.A.; Hamdi, W.S.; Al-Ansari, N.A.; Doiphode, S.H. Polymerase Chain Reaction Ribotyping of Clostridium Difficile Isolates in Qatar: A Hospital-Based Study. BMC Infect. Dis. 2014, 14, 502. [Google Scholar] [CrossRef]
- Hong, S.; Putsathit, P.; George, N.; Hemphill, C.; Huntington, P.G.; Korman, T.M.; Kotsanas, D.; Lahra, M.; McDougall, R.; Moore, C.V.; et al. Laboratory-Based Surveillance of Clostridium Difficile Infection in Australian Health Care and Community Settings, 2013 to 2018. J. Clin. Microbiol. 2020, 58, e01552-20. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.C.; Collins, D.A.; Imwattana, K.; Knight, D.R.; Perumalsamy, S.; Hain-Saunders, N.M.R.; Putsathit, P.; Speers, D.; Riley, T.V. Whole-Genome Sequencing Links Clostridium (Clostridioides) Difficile in a Single Hospital to Diverse Environmental Sources in the Community. J. Appl. Microbiol. 2021, 133, 1156–1168. [Google Scholar] [CrossRef]
- Johnston, M.; Irwin, J.; Roberts, S.; Leung, A.; Andersson, H.S.; Orme, G.; Deroles-Main, J.; Bakker, S. Clostridioides Difficile Infection in a Rural New Zealand Secondary Care Centre: An Incidence Case–Control Study. Intern. Med. J. 2021, 52, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Sathyendran, V.; Mcauliffe, G.N.; Swager, T.; Freeman, J.T.; Taylor, S.L.; Roberts, S.A. Clostridium Difficile as a Cause of Healthcare-Associated Diarrhoea among Children in Auckland, New Zealand: Clinical and Molecular Epidemiology. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1741–1747. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L. The Burden and Etiology of Diarrheal Illness in Developing Countries. Pediatr. Clin. N. Am. 2017, 64, 799–814. [Google Scholar] [CrossRef] [PubMed]
- Janssen, I.; Cooper, P.; Gunka, K.; Rupnik, M.; Wetzel, D.; Zimmermann, O.; Groß, U. High Prevalence of Nontoxigenic Clostridium Difficile Isolated from Hospitalized and Non-Hospitalized Individuals in Rural Ghana. Int. J. Med. Microbiol. 2016, 306, 652–656. [Google Scholar] [CrossRef]
- Seugendo, M.; Janssen, I.; Lang, V.; Hasibuan, I.; Bohne, W.; Cooper, P.; Daniel, R.; Gunka, K.; Kusumawati, R.L.; Mshana, S.E.; et al. Prevalence and Strain Characterization of Clostridioides (Clostridium) Difficile in Representative Regions of Germany, Ghana, Tanzania and Indonesia—A Comparative Multi-Center Cross-Sectional Study. Front. Microbiol. 2018, 9, 1843. [Google Scholar] [CrossRef]
- Berger, F.K.; Mellmann, A.; Bischoff, M.; von Müller, L.; Becker, S.L.; Simango, C.; Gärtner, B. Molecular Epidemiology and Antimicrobial Resistance of Clostridioides Difficile Detected in Chicken, Soil and Human Samples from Zimbabwe. Int. J. Infect. Dis. 2020, 96, 82–87. [Google Scholar] [CrossRef]
- Robishaw, J.D.; Alter, S.M.; Solano, J.J.; Shih, R.D.; DeMets, D.L.; Maki, D.G.; Hennekens, C.H. Genomic Surveillance to Combat COVID-19: Challenges and Opportunities. Lancet Microbe 2021, 2, e481–e484. [Google Scholar] [CrossRef]
- World Health Organization Global Genomic Surveillance Strategy for Pathogens with Pandemic and Epidemic Potential, 2022–2032. Available online: https://www.who.int/publications/i/item/9789240046979 (accessed on 27 July 2022).
- Jahn, K.; Dreifuss, D.; Topolsky, I.; Kull, A.; Ganesanandamoorthy, P.; Fernandez-Cassi, X.; Bamp, C.; Devaux, A.J.; Stachler, E.; Caduff, L.; et al. Early Detection and Surveillance of SARS-CoV-2 Genomic Variants in Wastewater Using COJAC. Nat. Microbiol. 2022, 7, 1151–1160. [Google Scholar] [CrossRef]
- Schumann, V.-F.; Cuadrat, R.R.d.C.; Wyler, E.; Wurmus, R.; Deter, A.; Quedenau, C.; Dohmen, J.; Faxel, M.; Borodina, T.; Blume, A.; et al. SARS-CoV-2 Infection Dynamics Revealed by Wastewater Sequencing Analysis and Deconvolution (Preprint). Sci. Total Environ. 2022, 853, 158931. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Weese, J.S.; Flemming, C.; Odumeru, J.; Warriner, K. Fate of Clostridium Difficile during Wastewater Treatment and Incidence in Southern Ontario Watersheds. J. Appl. Microbiol. 2014, 117, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Moradigaravand, D.; Gouliouris, T.; Ludden, C.; Reuter, S.; Jamrozy, D.; Blane, B.; Naydenova, P.; Judge, K.; Aliyu, S.H.; Hadjirin, N.F.; et al. Genomic Survey of Clostridium Difficile Reservoirs in the East of England Implicates Environmental Contamination of Wastewater Treatment Plants by Clinical Lineages. Microb. Genom. 2018, 4, e000162. [Google Scholar] [CrossRef] [PubMed]
- Gardy, J.L.; Loman, N.J. Towards a Genomics-Informed, Real-Time, Global Pathogen Surveillance System. Nat. Rev. Genet. 2017, 19, 9–20. [Google Scholar] [CrossRef]
- Kuehne, S.A.; Rood, J.I.; Lyras, D. Clostridial Genetics: Genetic Manipulation of the Pathogenic Clostridia. Microbiol. Spectr. 2019, 7, 20. [Google Scholar] [CrossRef]
- Joseph, R.C.; Kim, N.M.; Sandoval, N.R. Recent Developments of the Synthetic Biology Toolkit for Clostridium. Front. Microbiol. 2018, 9, 154. [Google Scholar] [CrossRef]
- Maikova, A.; Kreis, V.; Boutserin, A.; Severinov, K.; Soutourina, O. Using an Endogenous CRISPR-Cas System for Genome Editing in the Human Pathogen Clostridium Difficile. Appl. Environ. Microbiol. 2019, 85, e01416-19. [Google Scholar] [CrossRef]
- Zhu, D.; Wang, S.; Sun, X. FliW and CsrA Govern Flagellin (FliC) Synthesis and Play Pleiotropic Roles in Virulence and Physiology of Clostridioides Difficile R20291. Front. Microbiol. 2021, 12, 735616. [Google Scholar] [CrossRef]
- Hong, W.; Zhang, J.; Cui, G.; Wang, L.; Wang, Y. Multiplexed CRISPR-Cpf1-Mediated Genome Editing in Clostridium Difficile toward the Understanding of Pathogenesis of C. Difficile Infection. ACS Synth. Biol. 2018, 7, 1588–1600. [Google Scholar] [CrossRef]
- McAllister, K.N.; Bouillaut, L.; Kahn, J.N.; Self, W.T.; Sorg, J.A. Using CRISPR-Cas9-Mediated Genome Editing to Generate C. Difficile Mutants Defective in Selenoproteins Synthesis. Sci. Rep. 2017, 7, 14672. [Google Scholar] [CrossRef]
- Wang, S.; Hong, W.; Dong, S.; Zhang, Z.T.; Zhang, J.; Wang, L.; Wang, Y. Genome Engineering of Clostridium Difficile Using the CRISPR-Cas9 System. Clin. Microbiol. Infect. 2018, 24, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Selle, K.; Fletcher, J.R.; Tuson, H.; Schmitt, D.S.; McMillan, L.; Vridhambal, G.S.; Rivera, A.J.; Montgomery, S.A.; Fortier, L.C.; Barrangou, R.; et al. In Vivo Targeting of Clostridioides Difficile Using Phage- Delivered CRISPR-Cas3 Antimicrobials. mBio 2020, 11, e00019-20. [Google Scholar] [CrossRef]
- Roldan, G.A.; Cui, A.X.; Pollock, N.R. Assessing the Burden of Clostridium Difficile Infection in Low- and Middle-Income Countries. J. Clin. Microbiol. 2018, 56, e01747-17. [Google Scholar] [CrossRef] [PubMed]
- Kullin, B.; Abratt, V.R.; Reid, S.J.; Riley, T.V. Clostridioides Difficile Infection in Africa: A Narrative Review. Anaerobe 2022, 74, 102549. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Stewardship. Available online: https://www.ecdc.europa.eu/en/publications-data/directory-guidance-prevention-and-control/prudent-use-antibiotics/antimicrobial (accessed on 10 November 2022).
- CDC. What CDC Is Doing to Reduce C. Diff Infections. Available online: https://www.cdc.gov/cdiff/reducing.html (accessed on 10 November 2022).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mengoli, M.; Barone, M.; Fabbrini, M.; D’Amico, F.; Brigidi, P.; Turroni, S. Make It Less difficile: Understanding Genetic Evolution and Global Spread of Clostridioides difficile. Genes 2022, 13, 2200. https://doi.org/10.3390/genes13122200
Mengoli M, Barone M, Fabbrini M, D’Amico F, Brigidi P, Turroni S. Make It Less difficile: Understanding Genetic Evolution and Global Spread of Clostridioides difficile. Genes. 2022; 13(12):2200. https://doi.org/10.3390/genes13122200
Chicago/Turabian StyleMengoli, Mariachiara, Monica Barone, Marco Fabbrini, Federica D’Amico, Patrizia Brigidi, and Silvia Turroni. 2022. "Make It Less difficile: Understanding Genetic Evolution and Global Spread of Clostridioides difficile" Genes 13, no. 12: 2200. https://doi.org/10.3390/genes13122200
APA StyleMengoli, M., Barone, M., Fabbrini, M., D’Amico, F., Brigidi, P., & Turroni, S. (2022). Make It Less difficile: Understanding Genetic Evolution and Global Spread of Clostridioides difficile. Genes, 13(12), 2200. https://doi.org/10.3390/genes13122200