Abstract
The societal burden of non-communicable disease is closely linked with environmental exposures and lifestyle behaviours, including the adherence to a poor maternal diet from the earliest preimplantation period of the life course onwards. Epigenetic variations caused by a compromised maternal nutritional status can affect embryonic development. This review summarises the main epigenetic modifications in mammals, especially DNA methylation, histone modifications, and ncRNA. These epigenetic changes can compromise the health of the offspring later in life. We discuss different types of nutritional stressors in human and animal models, such as maternal undernutrition, seasonal diets, low-protein diet, high-fat diet, and synthetic folic acid supplement use, and how these nutritional exposures epigenetically affect target genes and their outcomes. In addition, we review the concept of thrifty genes during the preimplantation period, and some examples that relate to epigenetic change and diet. Finally, we discuss different examples of maternal diets, their effect on outcomes, and their relationship with assisted reproductive technology (ART), including their implications on epigenetic modifications.
1. Introduction
Non-communicable diseases (NCDs), such as diabetes, cardiovascular diseases (CVDs), neurological disorders, obesity, and some cancers, are on the rise, leading to around 34 million deaths worldwide per annum [1]. The significant increase in NCDs is closely linked to environmental exposures and lifestyle behaviours. It is also clear that exposure to these different largely modifiable conditions (i.e., poor diet, stress, and chemicals) from the preimplantation period onwards is important in the origin of these diseases [2]. The relationship between exposure to the maternal environment and the development and health of the foetus is captured in the developmental origins of health and disease (DOHaD) hypothesis [3].
The research in DOHaD, which has increased substantially from the 1990s, has shown that the maternal environment during pregnancy can affect programming during development and increase the risk of offspring developing long-term diseases [3,4]. Professor David Barker was one of the pioneer researchers to demonstrate the DOHaD phenomenon in his epidemiological studies in 1989, linking perinatal weight and the subsequent growth trajectory with health and disease risk in later life [5,6]. Since the development of the DOHaD hypothesis, many authors have linked birth weight with the development of NCDs. Initially, only low birth weight was related; however, some studies found that this relationship was U-shaped, indicating that high birth weight can also relate to the risk of developing pathologies associated with metabolic syndrome [7]. The maternal environment is one of the most studied conditions that can influence offspring phenotype. The maternal environment comprises both extrinsic and intrinsic factors. Extrinsic factors act from the outside and are related to the environmental conditions (pollution, exposure to chemicals, etc.), but intrinsic factors act from within an individual and are directly dependent on the mother’s condition (eating behaviours, metabolism, lifestyle, etc.). Despite the importance of human studies, animal studies provide an advantage in that environmental exposures can be controlled (extrinsic factors) and maternal and subject confounders can be reduced (intrinsic factors) [8]. These studies have demonstrated that the origin of NCDs can be attributable to the maternal and gestational environment, laying the foundation for DOHaD. These studies have further progressed to reveal that an array of environmental factors experienced during pregnancy, including toxins, stress, diet, and lifestyle, contribute to physiological and metabolic development, impacting foetal growth and offspring phenotype and health.
DOHaD research in nutritional studies has now found links between the quality of maternal diet during pregnancy and its possible contribution to changes in the genome expression of the offspring in utero. These changes act to coordinate the physiological, metabolic and growth characteristics, and are mediated through epigenetic mechanisms [9,10]. Epigenetic changes refer to different molecular states that affect the regulation of gene expression by changing the structural organisation of the DNA, but without changing the sequence [11]. Epigenetic changes during foetal development can depend on microenvironmental variations. Thus, environmental factors, such as maternal diet, during early pregnancy can affect maternal metabolism, which can also impact offspring development [12,13]. The main epigenetic changes include DNA methylation, histone modifications, and the expression of non-coding RNAs (ncRNAs). Whilst the association between the in utero environment and offspring health has been widely described over the years, the underlying molecular processes influencing the epigenome are generally unknown. The study of epigenetic changes that occur during pregnancy is not limited to natural conditions, but may also operate in clinical settings, such as ART. The next goal in DOHaD research will be to understand the molecular details linking the maternal and clinical environment to the epigenome, and the subsequent biological steps that lead to the onset of disease risk in later life [14].
This review will discuss the evidence on how maternal diet in mammalian models can impose epigenetic modifications in offspring, and can lead to the development of disease in adulthood. We will also review the thrifty gene concept, where dietary-mediated epigenetic changes cause downstream gene expression changes that directly control metabolism and homeostasis [15]. Finally, we will consider clinical ART conditions and their link with maternal diet in light of the preimplantation epigenetics underlying DOHaD.
2. DOHaD and Epigenetic Modifications during Development
In the framework of the DOHaD hypothesis, epigenetic mechanisms have been considered as a possible precursor of changes in developmental programming [16]. In the early pregnancy periods, maternal malnutrition (overnutrition or undernutrition) can alter development throughout gestation [17], and one of the possible causes of these alterations could be linked to epigenetic changes. The intrauterine environment is susceptible to any small changes, which can induce responses in the offspring that could lead to a higher risk of developing NCD later in life [17,18].
Different studies pointed towards the possible idea that, across generations, a compromised diet can affect the phenotype and epigenome of the next generation, making them prone to developing metabolic [19] and even mental illnesses [20], by epigenetic variations that are subject to variations in diet [20]. These premises represent a starting point to understand how lifestyle and dietary behaviours during pregnancy can alter an offspring’s health by triggering epigenetic modifications during development.
These epigenetic modifications will contribute to the expression, activity level, or silencing of a specific gene (examples below). These changes may be induced by means of adaptation to the environment and survival, as envisaged in the DOHaD hypothesis. Firstly, we will summarise the main types of epigenetic regulation of gene expression that occur during development.
2.1. DNA Methylation
One of the most common epigenetic modifications is DNA methylation. DNA methylation is of great importance in the regulation of genome functions and chromatin stability, among others [11].
DNA methylation is caused by the attachment of a methyl group to cytosine, which can generally occur when cytosine is located next to guanine at a cytosine–phosphate–guanine (CpG) site [21]. When there are high levels of DNA methylation at the CpG sites in the promoter regions of a gene, transcription factors are prevented from binding to the DNA, thus inhibiting the expression of that gene [21,22]. Conversely, when the CpG sites on CpG islands are unmethylated, gene expression is promoted [23,24].
In mammals, after fertilization, a general de-methylation process occurs, followed by de novo methylation as the parental genome becomes superseded by the new embryonic genome. The methylation of CpG sites during the first days after fertilization is catalysed by DNA methyltransferases (DNMTs),and is essential for tissue differentiation and maintenance during development [25,26].
DNA methylation has a crucial role in the development of the embryo and cell lineage specification, thereby regulating differential gene expression [22]. The pattern of DNA methylation in the periconceptional period may respond to environmental conditions to permit adaptations in development. This modification is well studied, and its understanding can elucidate how the environment and epigenetics in the early stages of development affect health in later life [22,27] (reviewed previously: [21,22,27,28]) (Table 1).

Table 1.
Maternal diets models and epigenetic modifications in offspring.
Some studies have focused on maternal exposure to, or ingestion of, toxic substances (tobacco, mercury, or arsenic), suggesting that, from conception, the embryo is susceptible to undergoing epigenetic changes (DNA methylation) when the maternal environment is compromised [56]. For example, exposure to arsenic during early pregnancy may be associated with changes in DNA methylation in the foetus, leading to reduced birth weight [56,57], and such exposures may increase the rate of miscarriage [58].
DNA methylation is crucial in the control of gene expression, as aberrant changes in DNA methylation patterns are associated with many diseases and cancers [59]. Alterations in DNA methylation patterns can be caused by genetic factors (polymorphism and mutations), or by factors such as diet, and the imbalance of essential nutrients in the metabolism of methyl groups [60].
2.2. Histone Modification
Histones are essential for DNA folding in eukaryotic cells, and also play a critical role in the regulation of gene expression. Histones are susceptible to different post-transcriptional modifications in the amino-terminal tail domain of histones, such as ubiquitination, phosphorylation, methylation, and acetylation, and these markers are often called histone codes [14].
Acetylation and methylation are the most common and most studied modifications in development [61]. Histone acetylation, through histone acetyltransferase activity, is related to gene expression and an open chromatin state, while deacetylation is related to gene silencing [62]. The methylation of histones by methyltransferases can promote or decrease gene transcription. Histone methylation has a crucial role in the assembly and compaction of heterochromatin, so when methylation increases the chemical attractions between DNA and histone tails, transcription decreases because the DNA compacts to nucleosomes and transcription-polymerase factors cannot access the DNA [63,64] (reviewed previously: [23,61,62,64,65]).
Histone modification is an important means for regulating gene expression, as these modifications will affect the function and expression of proteins, which can affect the development of the embryos [23,66] (Table 1). Previous studies have found that histone modification can affect embryonic development, even to the point of embryonic lethality [67].
2.3. Non-Coding RNAs (miRNA, lncRNA)
Another epigenetic modification is non-coding RNA (ncRNA) [68,69]. ncRNAs can be classified according to their size, into small (sncRNAs) and long ncRNAs (lncRNAs). In mammals, sncRNAs [70] include microRNA (miRNA) [68] and Piwi RNA [69]. lncRNAs are involved in the emergence and development of germ cells and embryos [71]. lncRNAs are associated with different molecular processes, such as splicing, regulation of gene expression (transcriptional and post-transcriptional processes), and chromatin modifications [72] ]. Their mechanism of action is complex, and can involve the creation of repressive domains (of many kilobases) [73], and can be classified into the following three models: (i) competitor, competing with other molecules by binding to the DNA; (ii) recruiter/activator, related to epigenetic modifications by activating epigenetic modifiers (i.e., promoting DNA methylation); (iii) precursor, where lncRNA can be processed by RNase to produce shorter active RNAs (involved in post-transcriptional regulation) [74,75,76]. In mammals, lncRNAs also have an essential role in X chromosome inactivation [77] (reviewed previously: [77,78,79]).
Although ncRNAs are very important in embryological development and are known to be a key regulator in chromosome formation, their mechanism and alteration in relation to maternal diet is still unclear.
3. Diet Models and Epigenetic Modifications: Examples and Biological Meaning
Nutrition is an important player in gene regulation, as alterations in nutrition can influence gene expression, and, more importantly, it has been described that early alterations in maternal nutrition during development may be responsible for the development of chronic diseases through epigenetic mechanisms [80,81].
Human and animal studies of maternal nutrition during pregnancy, or just the preimplantation period, have shown that it affects the epigenome and offspring phenotype, and can be categorised into different models and studies, as outlined in the following sections.
3.1. Dutch Famine
The effects of the Dutch famine of 1944–1945, during World War II, when Amsterdam was occupied for a five-month period by the Nazis, has been the subject of numerous epidemiological studies. During the famine, adult women received a daily food ration of some 400–800 calories, regardless of their social class. Immediately after the occupation, food became plentiful again, rations increased to 2000 calories, and, since medical records were detailed, the effects of malnutrition at key periods of pregnancy could be investigated [32,82,83]. Studies showed that men and women conceived during the famine, and who experienced famine during early pregnancy, were at an increased risk of developing metabolic diseases, such as obesity or diabetes II, in adulthood. Moreover, limited nutrient intake in early pregnancy decreased the development of pancreatic β-cells, resulting in glucose intolerance later in life [84,85].
Thus, among other modifications, a reduction was found in DNA methylation in famine-exposed offspring compared to same-sex controls, on the promoter of the insulin-like growth factor 2 (IGF2) gene that modulates foetal development and growth [27,29], and is implicated in metabolic syndrome [29]. Apart from low birth weight and CVD, early exposure to famine also caused low lipoprotein levels in offspring [30,31]. A recent study has shown an association between epigenetic changes in genes involved in the regulation of lipid metabolism and adipogenesis in offspring exposed to famine in early gestation [32]. Thus, the hypermethylated regions of the genes serine/threonine kinase Pim-3 (PIM3 influences cell growth and energy metabolism [86]), 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase-3 promoter (PFKFB3 is involved in glycolysis), and methyltransferase 8 (METTL8 is involved in adipogenesis) affected the expression of these genes by an increase in DNA methylation, and contributed to the development of adverse metabolic profiles. Additionally, men born after exposure to famine in early gestation had a two-fold increase in the risk of developing schizophrenia, and decreased intracortical volumes and total brain volumes, suggesting that early brain development may be a risk factor for developing schizophrenia [87,88].
3.2. Seasonal Diets: The Gambian Example
One example where seasonal diet affects offspring outcomes is in the rural economy in the African country the Gambia. This study distinguished a hungry/wet season (June–October) and a harvest/dry season (November–May). Epidemiological and longitudinal studies have suggested that there is a relationship between the timing of conception during the seasons and increased susceptibility to uterine growth restriction [89], an increase in premature death, and the development of NCD and CVD in adulthood [90].
One main objective of these studies was to find the underlying mechanisms, such as epigenetic regulation during the periconceptional period, that made these seasons change the health risk in adulthood. The modification of DNA methylation patterns has been associated with maternal diet during seasonal periods in the Gambia, since the foetal development period is sensitive to prenatal exposures, and drastic changes in DNA methylation can affect the reprogramming that occurs during embryogenesis [91,92]. Studies on Gambian populations have shown that different seasons (affecting food availability) influence the DNA methylation of metastable epialleles (MEs), since DNA methylation relies on nutritional inputs of methyl donors (folate, choline, and betaine) and essential cofactors (B2, B6, and B12 vitamins), which could be influenced by rainy and dry seasons [93]. MEs are genomic loci; DNA methylation patterns are originated from all three germ layers (before gastrulation) of MEs in an individual, and are maintained in differentiated tissues [33,93]. Therefore, the study of MEs provides a useful tool to identify the role of the periconceptional environment in the establishment of epigenetic changes, such as DNA methylation, in the offspring. Waterland et al. [33] analysed BolA family member 3 (BOLA3), paired box 8 (PAX8), SLIT and NTRK-like family member 1 (SLITRK1), and zinc finger FYVE-type containing 28 (ZFYVE28) as MEs in peripheral blood, and found that in individuals conceived during the nutritionally challenged rainy season, DNA methylation in MEs was higher. These data suggest that periconceptual nutritional status may influence DNA methylation changes at MEs in the early stages of development (preimplantation period), and impact the offspring’s health later in life [33]. Additionally, James et al. analysed one-carbon biomarkers in maternal plasma at the time of conception and 50 ME loci in their offspring’s blood in a Gambian population. They observed increased ME methylation, higher plasma folate, vitamin B2, betaine, and cysteine, and lower plasma homocysteine in rainy season conceptions. These preliminary results suggest that rainy and dry seasons could influence infant DNA methylation [34,94]. Some of these regions with altered methylations during the different seasonal periods, and their implication in the health of the offspring, have been closely related to one-carbon metabolism [93,95]. However, studies of the Gambian seasonal diet have not yet found a direct association between specific epigenetic changes and a given disease.
3.3. Low-Protein Diets (LPD)
Maternal malnutrition has been associated with the development of metabolic diseases in adult offspring, such as obesity, insulin resistance, and diabetes [13,17,35,96]. However, the mechanisms underlying the development of metabolic diseases in offspring have not been fully elucidated. Epigenetic modifications have been implicated as contributory since the levels of proteins and some essential amino acids and methyl group donors may influence epigenetic regulation [97]. Maternal LPD has recognisable effects from the preimplantation period through to late lactation, affecting the phenotype of the offspring, and increasing the risk of developing CVD, neurological disease, and diabetes [13,17,24,35,36,96,98,99,100,101,102].
In a mouse model, male offspring from mothers exposed to an LPD (10% protein) during gestation and lactation had reduced body weight compared with controls, and exhibited a loss of methyl groups from CpGs in the promoter of the leptin gene in adipose cells, coincident with reduced levels of leptin mRNA [103]. Leptin is an appetite-regulating hormone, and controls body weight through food intake and energy expenditure. Therefore, the imbalance in leptin levels, caused by maternal LPD, suggests that these alterations could not prepare the individual for an environment with an excess of food, leading to an increased risk of obesity [103,104].
Another study exploring the epigenetic changes associated with maternal LPD was conducted in a pig model [36]. The mothers were fed LPD (6%) one month before fertilisation and during pregnancy. Here, the epigenetic regulation of glucose-6-phosphatase (G6PC), which has a crucial role in glucose regulation and the risk of hyperglycaemia, was studied. The study showed that the G6PC promoter was hypomethylated in male offspring, together with increased methylation of H3K4 in the G6PC promoter in the liver. Therefore, LPD during pregnancy caused activation of the G6PC gene in males, linked with the development of hyperglycaemia and diabetes in adulthood [36]. Similar offspring outcomes were shown in another study using LPD (9% protein) in a pregnant rat model. Here, the effect of LPD on glucose transporter 4 (GLUT4), involved in insulin tolerance in the skeletal muscle in the offspring, was investigated [37]. Histone H3 and H4 were acetylated and histone H3K4 was dimethylated in the GLUT4 promoter region in female offspring, providing evidence that glucose metabolism may be adapted by epigenetic processes in LPD offspring in a sex-dependent manner. The resulting low expression of GLUT4 is associated with metabolic syndrome and the development of type II diabetes.
In a further maternal mouse LPD model, the modification of histone acetylation and increased H3K9 methylation in the promoter domain of the cholesterol 7α-hydroxylase gene (Cyp7a1), which catabolizes the conversion of cholesterol to bile acids, was associated with decreased expression in the liver of male offspring [105]. Moreover, similar to previous studies in mice, decreased acetylation and increased methylation of histone H3K9 were reported, which regulates the Cyp7a1 promoter, reducing expression and increasing the risk of metabolic diseases, including insulin resistance and raised cholesterol levels, in offspring [38].
In other studies using the pig LPD (6.5% protein) model during gestation, hepatic cytochrome C gene (CYCS) expression was increased in the liver, and was associated with increased methylation of specific CpG sites in the CYCS promoter in new-born LPD offspring [40]. CYCS expression is associated with mitochondrial energy metabolism and production, and with increased oxidative stress. A study of ewes born on a restricted maternal diet (LPD) obtained similar results, showing increased cytochrome C protein in the liver of LPD offspring, indicating increased mitochondrial activity [41].
These varied examples of maternal LPD during gestation, across mammalian species, show how epigenetic changes, induced by certain dietary exposures, may be associated with the development of CVD, insulin resistance, eating disorders, obesity, and other metabolic diseases in offspring.
3.4. High-Fat Diets (HFD)
Several studies have focused on the effect of excess maternal fat and caloric consumption on offspring epigenetic changes and disease risk, such as obesity. Obesity is a complex disease caused by different factors, including high-fat diets, stress, eating disorders, a sedentary lifestyle, and genetic factors. The increase in obesity is a major threat to health worldwide, regardless of gender and age. In both developed and developing countries, a change to a more sedentary lifestyle and the excess consumption of fatty foods have had a negative effect on population health [106]. Currently, approximately 36.5% of men and 38% of women worldwide are classified as obese [107], and in the last twenty years, the prevalence of maternal obesity has increased alarmingly [108,109]. Maternal HFD (and obesity) affects the embryo and foetus during gestation, causing changes at the epigenetic level during reprogramming, promoting the propensity of the embryo to develop diseases (such as insulin resistance), due to exposure to this environment in utero [110]. Some of the epigenetic modifications due to maternal HFD will be discussed. In vivo studies are valuable in identifying the link between maternal HFD and disease outcomes in offspring, as well as in determining the most vulnerable time point in development [111].
During pregnancy, maternal obesity profoundly influences the risk of obesity and metabolic diseases in the offspring; for example, the offspring of mothers fed an HFD (45% fat) for 12 weeks showed a non-alcoholic steatohepatitis phenotype (most prominent in males) and altered methylation patterns (hypermethylation) in genes implicated in liver fibrosis and lipid accumulation, such as Ephb2 (ephrin type-B receptor 2) and Fgf21 (fibroblast growth factor 21), suggesting that epigenetic changes could favour the development of fatty liver disease in the offspring [42]. Similarly, and as mentioned above, leptin regulation is essential to prevent metabolic diseases. In a maternal HFD (60% fat) mouse model, hypermethylation at the promotor region of adiponectin (visceral fat) and leptin receptor genes (visceral fat and liver have a role in adipogenesis), and decreased DNA methylation at the promotor of the leptin gene, were reported in the adult offspring [112]. Indeed, in a mouse model, the offspring of mothers exposed to an HFD during pregnancy showed modifications in the methylation status of the adiponectin promoter region in the visceral fat, and changes in methylation in the leptin promoter region, suggesting that these epigenetic modifications may modulate adiponectin and leptin expression in the adipose tissue, potentially leading to a phenomenon similar to the metabolic syndrome [10,43].
Another study presented an association of epigenetic modifications in the Pomc (proopiomelanocortin—biomarker for obesity detection) promoters in female rats. The adult female offspring of HFD dams (60% fat) had increased body weight and blood leptin levels, and their weight gain was related to Pomc expression. The hypothalamic Pomc promoter region was hypermethylated in the offspring of HFD-fed mothers, showing a long-term effect on Pomc methylation programming [44]. Consistent with this notion, the offspring of female rats fed an HFD (34% fat) before and during gestation and lactation developed a metabolic syndrome-like phenotype and an altered Pomc promoter methylation pattern in the hypothalamus. However, the male offspring were more predisposed to alterations in the methylation patterns, and were the only offspring that showed hypermethylation in different CpG islands and the promoter region of the Agrp (agouti-related peptide) gene in the hypothalamus [45]. Additionally, in humans, a study demonstrated that epigenetic changes, especially leukocyte methylation levels in the promoter region of the Pomc gene, may be associated with weight regain in obese men [46]. The biological significance of this hypermethylation is related to the dysregulation of leptin, leptin receptors, and the Agrp-Pomc system in the brain, and weight gain in offspring, which may lead to obesity later in life. Interestingly, another maternal over-feeding model, with Wistar rats fed an HFD (34% fat) during gestation and lactation, described changes in the gene expression and hypermethylation pattern of the insulin receptor (InsR) DNA promoter in the hypothalamus in a sex-specific manner, leading to a metabolic syndrome-like phenotype in the male offspring; however, lower methylation levels of Pomc were associated with weight loss [46].
In a macaque maternal HFD (35% fat) model during gestation, the offspring exhibited hyperacetylation of the foetal hepatic tissue at H3K14, and increased H3K9 and H3K18 acetylation (not significant) [47]. These histone changes were related to an increase in the expression of offspring genes, including glutamic pyruvic transaminase 2 (GPT2, which is essential in the maintenance of obesity in the postnatal period) and retinol dehydrogenase 12 (Rdh12 regulates feeding behaviour and adaptation to circadian rhythms). Rdh12 is also related to the family of short-chain alcohol dehydrogenases [48], so its deregulation could cause non-alcoholic fatty liver disease [49]. Additional findings in the foetal heart of non-human primate offspring, following maternal HFD and obesity during pregnancy, showed dysregulation in 80 miRNAs [51] involved in the migration of the cardiac neural crest during development [113].
A recent study using HFD (60%) during pregnancy and lactation in rats found that maternal HFD affects cannabinoid receptor 1 (Cnr1) expression and its underlying epigenetic status in the brain of offspring at early ages [20]. Gawlinski et al. found that Crn1 mRNA decreased and miR-212-5p and CpG island methylation increased in the CnIr1 promoter in the cortex of male offspring before adulthood. Interestingly, these authors found the opposite effect in the hippocampus of adolescent male offspring; the mRNA of the Cnr1 increased, thus increasing its expression and, at the same time, decreasing the expression of miR-212-5p and the methylation of CpG islands in the gene promoter. The authors related these expression changes in the Cnr1 to the coding of the CB1 receptor (related to depressive behaviour), and to the phenotype observed in the offspring, who presented a depressive-like phenotype [20].
These examples show that maternal HFD during gestation produces changes in the offspring epigenome across different mechanisms and species, including primates, which increase the risk of developing diseases such as non-alcoholic fatty liver disease, diabetes, obesity, CVD, and other NCDs.
3.5. One-Carbon Metabolism: Folic Acid Supplement Use
One-carbon metabolism is related to pathways involved in the methylation of proteins, nucleic acids, and phospholipids, such as the pathway of folate and methionine, among others. The nutrients associated with one-carbon metabolism (riboflavin, folates, B12, among others) must be obtained from food because the body does not synthesize them. However, these nutrients are very important for the development of the gametes, embryos, and the health of the offspring later in life [114,115,116]. Alterations in the uterine environment, such as changes in the maternal diet, can compromise one-carbon metabolism regulation, which can trigger endocrine and metabolic changes during development [117,118]. One-carbon metabolism covers a large number of metabolic pathways that supply methyl groups required for DNA methylation [119]. Methylation reactions are imparted during embryonic and foetal development; therefore, dietary deficiencies of certain nutrients associated with one-carbon metabolism (i.e., vitamin B6, vitamin C, folate, choline, etc.) can affect epigenetic processes in utero, leading to changes in methylation that may promote the development of diseases in the offspring later in life [120,121]. In this review, we focus our one-carbon metabolism discussion on the importance of folate in the maternal diet during the preimplantation period and gestation. One of the best-known examples is the absence of folates in the maternal diet during the first months of pregnancy, which can cause diseases during neural tube development in the foetus (“spina bifida”) [122,123]. Other studies in human cohorts found an association between folic acid supplementation during the periconception period and increased methylation in IGF2 in the offspring [124]. DNA methylation during development is related to methyl donor groups obtained from one-carbon metabolism, so there are several associated studies [115,119,125,126,127,128,129,130].
In vivo studies of a rat diet comprising a maternal folic acid supplement used during pregnancy showed that nutrient imbalance produced global DNA hypermethylation in the cortex of the adult offspring [52]. These animals were analysed in behavioural tests, and a longer escape latency was observed compared to the controls. The cortex is associated with cognitive behaviour, decision making, and memory, so changes in the DNA methylation of this brain region can trigger brain disorders, such as Alzheimer’s disease and bipolar disorder [53]. Folic acid supplement use (5 mg/day) during pregnancy in rats induced a significant reduction in DNA methylation in the mammary glands of adult offspring [54]. Surprisingly, these findings have been associated with an increased risk of breast cancer, so high folate levels during pregnancy and lactation are associated with an increased risk of breast cancer in offspring.
A mouse embryonic stem cell culture model has been used to determine the effects of restricted folate supplementation (0.5 mg/L folate) in early development, revealing hypomethylation of the long interspersed nuclear element-1 (LINE-1) promoter domain and increased expression of LINE-1, which is related to neural tube defects [55]. This change was associated with decreased transformation of stem cells into embryonic neural tissue, indicating neural tube defects.
The above examples of manipulating folic acid supplement use during pregnancy elucidate some of the epigenetic mechanisms that are regulated. An unbalanced folate diet can induce the development of neurological diseases, and even cancer, in offspring. However, more studies on the underlying mechanisms are needed to better understand folic acid supplementation.
A summary of all the examples revised in the different diet models mentioned above is shown in Table 1.
4. From Preimplantation Period to Development: Epigenetics and Thrifty Genes
Nutrient accessibility during pregnancy affects crucial events, such as cell growth and division, that are necessary for optimal gametogenesis and embryo development [131,132]. One of the periods on which we would like to place special emphasis, and in accordance with the premises of the DOHaD hypothesis, is the preimplantation period. The preimplantation period encompasses all the morphological, structural, and epigenetic (mainly DNA methylation and histone modifications) enhancements and remodelling that occur in the embryo until just before implantation in the uterus [133]. This is a very short period, but is highly vulnerable to changes in the environment, in which epigenetic modifications can affect gene expression in the developing embryo, which can lead to health defects in adult life; for example, it has been shown that dietary changes during the preimplantation period alone (especially in murine models) can lead to neurological pathologies, NCD, and CVD in the offspring during adulthood [13,131,134,135]. One possible explanation for these events is the thrifty gene hypothesis. This hypothesis was proposed by the geneticist James Neel in 1962, its premise being the study of diabetes and the genes that promote its expression [136]. This hypothesis refers to hunter-gatherer populations, in which the predisposition of these genes allowed them to accumulate fat more efficiently during periods of greater nutrient abundance. Thus, the hypothesis proposes that the expression of certain genes, whilst increasing disease susceptibility in later life, may have, historically, been advantageous in periods of famine or lack of nutrients, but today, the expression of these gene may be detrimental [136,137,138]. Over the years, some authors have proposed some candidate thrifty genes. However, we have found that few have linked these changes to maternal diet [139].
One of the basic units of cellular protein translation and cellular growth is the ribosome, and it has been shown that rDNA can be activated by epigenetic mechanisms [140,141]; for instance, the transcription of rDNA can be silenced by hypermethylation of the rDNA promoter, induced by environmental conditions [140]. Through hypermethylation of the rDNA promoter, rDNA transcription can be silenced, so if the environmental conditions to which the mother is exposed can affect epigenetic regulation, they may also affect rDNA expression for development [142]. Ribosomal RNA expression is sensitive to maternal diet, and, using the mouse LPD model, it was shown that poor diet during the preimplantation period was sufficient to activate a lifetime pattern of enhanced rRNA expression, only when the dietary restriction was lifted during either the foetal or adult stages, reflecting a thrifty phenotype response [142]. A similar effect in response to gestational HFD on epigenetic changes in rRNA expression has been reported [143]. A more recent study has proposed the phosphatase and tensin Homolog (PTEN) as a possible thrifty gene that undergoes epigenetic modifications related to maternal diet. However, the epigenetic regulations involved that affect PTEN expression/silencing have not yet been found [139].
Therefore, we consider that the study and identification of affected genes by maternal diet models could be of great interest Thus, epigenetic changes that can occur in these genes during the preimplantation period can lead to the phenotype of the offspring being altered.
5. ART and Epigenetic Modifications
ART includes a range of techniques, such as superovulation, sperm capacitation, intrauterine insemination (IUI), intracytoplasmic sperm injection (ICSI), in vitro fertilization (IVF) and culture, cryopreservation, and embryo transfer for infertility treatment in humans and the production of transgenic farm animals [144,145]. ART treatment can interfere with epigenetic changes, particularly DNA methylation [146], starting from the preimplantation period in humans and animals [147,148], affecting early embryogenesis and offspring health [149,150,151]. The procedures implemented during ART treatment, such as ovarian stimulation using hormones, the in vitro maturation of oocytes and preimplantation embryos, and the use of ICSI and embryo cryopreservation, expose the embryo to environmental conditions that differ from those in spontaneous fertilization and embryogenesis, and which may alter the normal epigenetic mechanisms [149,152].
5.1. Human Studies
DNA methylation, following ART, has been investigated at different stages of development. Genome-wide DNA methylation profiling in cord blood and placenta revealed differences in the methylation status between ART and spontaneous conceiving populations at certain CpG sites [153,154,155,156]. In ICSI-manipulated oocytes, this can include an abnormal methylation status at imprinted genes at differentially methylated region 1 (DMR1), H19 DMR, and PEG1 DMR [157]. Following ICSI treatment, day 3 embryos also showed aberrant methylation patterns at the same imprinted loci [158]. The aberrant methylation pattern of these DMRs is linked to imprinting disorders, such as Silver–Russell syndrome (SRS) and Beckwith–Weidemann syndrome (BWS) [159,160,161,162,163]. All the BWS patients conceived via ART had DNA methylation errors at H19/Igf2 IG DMR and/or KCNQ10T1:TSS DMR, and the DNA methylation error rates were significantly higher when compared to the spontaneously conceived BWS patients (100% versus 43.6% DNA methylation error rates, respectively) [151]. Interestingly, the DNA methylation levels of H19/Igf2 DMR and KCNQ10T1 DMR were significantly lower in the placentas of IVF/ICSI patients compared to the spontaneously conceived patients [164]. However, no differences in DNA methylation levels were reported between IVF and the spontaneous controls at individual CpG sites and entire DMRs of KvDMR1 in cord blood and placental villi, as well as PEG10 in placental villi [165]. These findings suggest that certain epigenetic variations in ART-conceived embryos may resolve throughout development, whereas others do not, and might associate with offspring outcomes. To further support this hypothesis, a positive correlation was observed between the DNA methylation of ERVFRD-1 and placental weight, as well as ERVFRD-1 and SNURF with birth weight, whereas the epigenetic variations at birth, as a consequence of ART, were largely resolved by adulthood [164,166]. Moreover, the epigenetic changes at certain genes and the transmission to offspring might be tissue-specific [164,165]; for example, the differences in methylation levels between ART and spontaneous conceptions for the H19/Igf2 and KCNQ1OT1 DMRs observed in the placenta were absent in the cord blood [164].
5.2. Animal Studies
Animal studies provide more insight into the prenatal period to help understand the origin and flow of epigenetic changes. The profiling of DNA methylation levels in mouse embryonic cells, blastocysts, the placenta, and the foetus displayed differences between ART and the controls [152,167,168,169]. Similarly to humans, some epigenetic alterations at certain genes among the ART population arise during the periconception and preimplantation period, and can be maintained throughout pregnancy. Aberrant methylation patterns at certain CpG sites of the Igf2/H19 imprinting control region (ICR) are observed in the IVF mouse embryonic stem cells (ESCs), placenta, foetal brain, and liver tissues of certain mouse strains [170,171]. Differences in methylation status at the H19 ICR were also reported in ART blastocysts, embryos, aorta, and placental tissues [149,172]. Nevertheless, the presence of alterations might also differ among the crossed strains [150,171,173]. A reduction in methylation for KvDMR1 and KCNQ1OT1 was observed in the ART placentas compared to the controls, whereas no difference was observed in the foetal brain and liver tissues [150,152,171,172]. Other examples are the small nuclear ribonucleoprotein polypeptide N (Snrpn), PEG1 and PEG3 ICRs that showed an altered methylation status in certain tissues, whereas no difference was observed in others [150,171,172,174]. Altogether, these alterations can be correlated to the aberrant expression levels of the relative genes, blastocyst maturity, and foetal development, and the imprinting disorders observed among ART offspring [149,152]. Moreover, the alterations in the methylation levels of SCAP/SCREPF1-2 in the lungs and the promoter of the endothelial nitric oxide synthase (eNOS) gene in the aortic and vascular tissue implicate the role of epigenetic alterations as a basis for the cardiovascular and respiratory dysfunction observed among ART-conceived offspring [167,174,175,176].
Whether these epigenetic changes are due to the underlying subfertility or to the ART treatment in humans warrants further investigation, yet similar datasets from animal models suggest that the effect of ART cannot be ignored; for example, ART was associated with alterations in DNA methylation in embryos at H19 and PEG 1 DMRs in both human and animal models [158,172]. In placentas, an alteration in the KCNQ1OT1 methylation status was associated with ART in human and animal models [164,171], whereas the outcomes on KvDMR1 and PEG10 ICR were inconsistent between humans and animals [150,165]. Besides, differences in methylation patterns have also been reported between different ART procedures [149,152,164,168], which emphasizes the effect of ART manipulation on the epigenetic profile and subsequent disease risk
6. Diet and ART Outcomes
In the sub-fertile population, diet has been related to fertility [177,178] and implicated in the intermediate ART outcomes, such as fertilization rate [179] and embryo yield [180], as well as clinical outcomes, including, importantly, pregnancy [181] and live birth rates [179]. Adherence to a healthy dietary pattern increased the number of mature oocytes [181,182]. Higher fertility and implantation rates were associated with the Iranian traditional medicine (ITM)-based diet and “pro-fertility” diet, respectively [181,183]. Increased embryo yield was observed with higher adherence towards the Mediterranean and ITM-based diets [180,181]. The chance of pregnancy was also improved with increased adherence to the “pro-fertility”, Mediterranean and ITM-based diets [181,183,184,185]. Furthermore, Twigt et al. reported an association between the chance of ongoing pregnancy and higher preconception dietary risk scores [186]. Besides, the rate of live birth was also improved by healthy dietary behaviours in women [179,183,187]. In contrast, unhealthy dietary behaviour was linked to a lower mature oocyte count, embryo quality, pregnancy chance, and live birth rate in human and mouse models [182,188,189]. Moreover, Li et al. investigated the effect of antioxidant intake, and showed improvements in the oocyte yield and live birth rate, which were dependent on the source and type of antioxidant [190].
A diet–epigenetic–ART connection might, in part, explain the ART outcomes observed, yet studies investigating this connection are limited [172,191]; for example, a healthy and balanced maternal diet could, through correcting the epigenetic alterations caused by ART, modify the corresponding outcomes [172,192]. On the other hand, an unbalanced diet (i.e., HFD diets) may pose epigenetic alterations that might not be observed otherwise [193]. A suggested mechanism behind this connection can be related to the effect of diet on the molecular interactions and biomarker levels within the body, which can induce epigenetic alterations; for example, improved pregnancy and live birth rates were associated with folate intake, which was also observed with adherence to the Mediterranean diet, which showed an association with the folate and vitamin B6 levels [179,184,185]. Subsequently, Rahimi et al. showed that DNA methylation defects induced by ART were partially restored with moderate folic acid supplementation in a mouse embryo and placenta [172]. In terms of phenotypic changes, moderate folic acid supplementation reduced the number of embryos with developmental delays, associated with ART treatment [172], which can further affect the disease risk in offspring [194,195].
7. Conclusions
Maternal diet could induce metabolic and physiological changes in offspring through epigenetic modifications. The altered epigenetic regulation of genes is associated with increased predisposition to disease later in life. Despite the advances in sequencing, there are still many mechanisms that need to be studied to understand the exact contributions of developmentally induced epigenetic markers and their effect on the risk of disease development. Understanding the relationship between maternal diet during pregnancy, epigenetic markers, and disease development may allow the discovery of therapeutic targets for the prevention and treatment of NCDs.
There is now great interest in understanding the environmental basis affecting epigenetic modifications that may be the origin of diseases in offspring. In humans, we have mentioned examples of epidemiological and ART-associated studies that could be used to understand the origins of some diseases or syndromes. However, animal studies are more flexible and controlled, as defined conditions can provide us with more information on the environmental effect during embryonic development and the outcome of the offspring in a shorter time, and, interestingly, we found that in both the natural condition and ART, maternal diet influences the offspring outcomes (Figure 1).
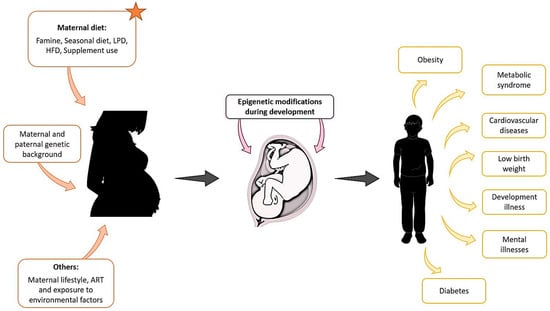
Figure 1.
Maternal exposure to environmental factors, diet changes, genetic background, and other parameters such as lifestyle, can affect the development of the foetus from the first stages of pregnancy and compromise the health of the offspring later in life.
Moreover, the theory of thrifty genes provides an interesting area to understand the epigenetic regulation during the preimplantation period. For this reason, we consider that the study of these genes during early development can be crucial to understand the effect of maternal diet on epigenetic modifications.
Author Contributions
Conceptualisation and writing—original draft preparation, I.P.-S.; writing—review and editing, B.H., D.A.O., R.P.M.S.-T. and S.W.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This review has received funding from the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant agreement No. 812660 (DohART-NET) and under Colciencias doctoral scholarship (convocatoria 646-Doctorados en el exterior) and Newton-Caldas Fund scholarship (convenio 334 of 2015).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Tom P Fleming for his comments and for improving the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Agrp | Agouti-related peptide |
| ART | Assisted reproductive technology |
| BOLA3 | BolA family member 3 |
| BWS | Beckwith–Weidemann syndrome |
| CpG | Cytosine nucleotide phosphate bonded to a guanine nucleotide |
| Cnr1 | Cannabinoid receptor 1 |
| CVD | Cardiovascular diseases |
| CYCS | Cytochrome C gene |
| Cyp7a1 | Hepatic cholesterol 7α-hydroxylase |
| DMR | Differentially methylated region |
| DNMTs | DNA methyltransferases |
| DOHaD | Developmental origins of health and disease |
| ESC | Embryonic stem cell |
| Ephb2 | Ephrin type-B receptor 2 |
| eNOS | Endothelial nitric oxide synthase |
| Fgf21 | Fibroblast growth factor 21 |
| G6PC | Glucose-6-phosphatase |
| GLUT4 | Glucose transporter 4 |
| GPT2 | Glutamic pyruvic transaminase 2 |
| H3K4 | Histone H3 and H4 |
| HFD | High-fat diet |
| ICR | Imprinting control region |
| ICSI | Intracytoplasmic sperm injection |
| IGF2 | Insulin-like growth factor 2 |
| InsR | Insulin receptor |
| ITM | Iranian traditional medicine |
| IUI | Intrauterine insemination |
| IVF | In vitro fertilization |
| LINE-1 | Long interspersed nucleotide element-1 |
| lncRNAs | Long ncRNAs |
| LPD | Low-protein diet |
| ME | Metastable epialleles |
| METTL8 | Methyltransferase 8 |
| NCD | Non-communicable diseases |
| ncRNAs | Non-coding RNAs |
| PAX8 | Paired box 8 |
| PDR | Preconception dietary risk |
| PFKFB3 | 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 |
| PIM3 | serine/threonine kinase Pim-3 |
| Pomc | Proopiomelanocortin |
| PTEN | Phosphatase and tensin homolog |
| RDH12 | Retinol dehydrogenase 12 |
References
- Stanaway, J.D.; Afshin, A.; Gakidou, E.; Lim, S.S.; Abate, D.; Abate, K.H.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; et al. Global, Regional, and National Comparative Risk Assessment of 84 Behavioural, Environmental and Occupational, and Metabolic Risks or Clusters of Risks for 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1923–1994. [Google Scholar] [CrossRef]
- Bateson, P. Fetal experience and good adult design. Int. J. Epidemiol. 2001, 30, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.P. The origins of the developmental origins theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef]
- Lin, Y.-J. Metabolic syndrome in children and adolescents born premature and small-for-gestational age: A scenario of Developmental Origins of Health and Disease (DOHaD). Pediatr. Neonatol. 2018, 59, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.; Osmond, C.; Winter, P.; Margetts, B.; Simmonds, S. Weight in Infancy and Death from Ischaemic Heart Disease. Lancet 1989, 334, 577–580. [Google Scholar] [CrossRef]
- Limesand, S.W.; Thornburg, K.L.; Harding, J.E. 30th anniversary for the developmental origins of endocrinology. J. Endocrinol. 2019, 242, E1–E4. [Google Scholar] [CrossRef]
- Blackmore, H.L.; Ozanne, S.E. Maternal diet-induced obesity and offspring cardiovascular health. J. Dev. Orig. Health Dis. 2013, 4, 338–347. [Google Scholar] [CrossRef]
- Bianco-Miotto, T.; Craig, J.M.; Gasser, Y.P.; Van Dijk, S.J.; Ozanne, S.E. Epigenetics and DOHaD: From basics to birth and beyond. J. Dev. Orig. Health Dis. 2017, 8, 513–519. [Google Scholar] [CrossRef]
- Ramírez-Alarcón, K.; Sánchez-Agurto, A.; Lamperti, L.; Martorell, M. Epigenetics, Maternal Diet and Metabolic Programming. Open Biol. J. 2019, 7, 45–51. [Google Scholar] [CrossRef]
- Masuyama, H.; Hiramatsu, Y. Effects of a High-Fat Diet Exposure in Utero on the Metabolic Syndrome-Like Phenomenon in Mouse Offspring through Epigenetic Changes in Adipocytokine Gene Expression. Endocrinology 2012, 153, 2823–2830. [Google Scholar] [CrossRef]
- Yamada, L.; Chong, S. Epigenetic studies in Developmental Origins of Health and Disease: Pitfalls and key considerations for study design and interpretation. J. Dev. Orig. Health Dis. 2017, 8, 30–43. [Google Scholar] [CrossRef]
- Slater-Jefferies, J.L.; Lillycrop, K.A.; Townsend, P.A.; Torrens, C.; Hoile, S.P.; Hanson, M.A.; Burdge, G.C. Feeding a protein-restricted diet during pregnancy induces altered epigenetic regulation of peroxisomal proliferator-activated receptor-α in the heart of the offspring. J. Dev. Orig. Health Dis. 2011, 2, 250–255. [Google Scholar] [CrossRef]
- Watkins, A.J.; Lucas, E.S.; Wilkins, A.; Cagampang, F.R.A.; Fleming, T.P. Maternal Periconceptional and Gestational Low Protein Diet Affects Mouse Offspring Growth, Cardiovascular and Adipose Phenotype at 1 Year of Age. PLoS ONE 2011, 6, e28745. [Google Scholar] [CrossRef] [PubMed]
- Lillycrop, K.; Burdge, G.C. Maternal diet as a modifier of offspring epigenetics. J. Dev. Orig. Health Dis. 2015, 6, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Speakman, J.R. Thrifty genes for obesity and the metabolic syndrome—Time to call off the search? Diabetes Vasc. Dis. Res. 2006, 3, 7–11. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.; Cooper, C.; Thornburg, K. Effect of in utero and early-life conditions on adult health and disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef]
- Fleming, T.; Velazquez, M.; Eckert, J.; Lucas, E.; Watkins, A. Nutrition of females during the peri-conceptional period and effects on foetal programming and health of offspring. Anim. Reprod. Sci. 2012, 130, 193–197. [Google Scholar] [CrossRef]
- Franczak, A.; Zglejc, K.; Waszkiewicz, E.; Wojciechowicz, B.; Martyniak, M.; Sobotka, W.; Okrasa, S.; Kotwica, G. Periconceptional undernutrition affects in utero methyltransferase expression and steroid hormone concentrations in uterine flushings and blood plasma during the peri-implantation period in domestic pigs. Reprod. Fertil. Dev. 2017, 29, 1499–1508. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, C.M.; Figueiredo, V.P.; Barbosa, M.A.; Cardoso, L.M.; Alzamora, A.C. Maternal high-fat diet triggers metabolic syndrome disorders that are transferred to first and second offspring generations. Br. J. Nutr. 2020, 123, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Gawliński, D.; Gawlińska, K.; Smaga, I. Maternal High-Fat Diet Modulates Cnr1 Gene Expression in Male Rat Offspring. Nutrients 2021, 13, 2885. [Google Scholar] [CrossRef]
- Jang, H.S.; Shin, W.J.; Lee, J.E.; Do, J.T. CpG and Non-CpG Methylation in Epigenetic Gene Regulation and Brain Function. Genes 2017, 8, 148. [Google Scholar] [CrossRef] [PubMed]
- Bestor, T.H.; Edwards, J.R.; Boulard, M. Notes on the role of dynamic DNA methylation in mammalian development. Proc. Natl. Acad. Sci. USA 2015, 112, 6796–6799. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Quan, S.; Yang, G.; Chen, M.; Ye, Q.; Wang, G.; Yu, H.; Wang, Y.; Qiao, S.; Zeng, X. Nutritional Status Impacts Epigenetic Regulation in Early Embryo Development: A Scoping Review. Adv. Nutr. 2021, 12, 1877–1892. [Google Scholar] [CrossRef]
- Cao, J.; Zhu, Q.; Liu, L.; Glazier, B.J.; Hinkel, B.C.; Liang, C.; Shi, H. Global Transcriptome Analysis of Brown Adipose Tissue of Diet-Induced Obese Mice. Int. J. Mol. Sci. 2018, 19, 1095. [Google Scholar] [CrossRef] [PubMed]
- Bestor, T. Activation of mammalian DNA methyltransferase by cleavage of a Zn binding regulatory domain. EMBO J. 1992, 11, 2611–2617. [Google Scholar] [CrossRef]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA Methyltransferases Dnmt3a and Dnmt3b Are Essential for De Novo Methylation and Mammalian Development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef]
- Felix, J.F.; Cecil, C.A.M. Population DNA methylation studies in the Developmental Origins of Health and Disease (DOHaD) framework. J. Dev. Orig. Health Dis. 2018, 10, 306–313. [Google Scholar] [CrossRef]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef]
- Heijmans, B.T.; Tobi, E.; Stein, A.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef]
- Tobi, E.; Lumey, L.H.; Talens, R.P.; Kremer, D.; Putter, H.; Stein, A.; Slagboom, P.; Heijmans, B.T. DNA methylation differences after exposure to prenatal famine are common and timing- and sex-specific. Hum. Mol. Genet. 2009, 18, 4046–4053. [Google Scholar] [CrossRef]
- Tobi, E.W.; Slieker, R.C.; Stein, A.D.; Suchiman, H.E.D.; Slagboom, P.E.; Van Zwet, E.W.; Heijmans, B.T.; Lumey, L.H. Early gestation as the critical time-window for changes in the prenatal environment to affect the adult human blood methylome. Int. J. Epidemiol. 2015, 44, 1211–1223. [Google Scholar] [CrossRef] [PubMed]
- Tobi, E.W.; Slieker, R.C.; Luijk, R.; Dekkers, K.F.; Stein, A.D.; Xu, K.M.; Biobank-based Integrative Omics Studies Consortium; Slagboom, P.E.; van Zwet, E.W.; Lumey, L.H.; et al. DNA methylation as a mediator of the association between prenatal adversity and risk factors for metabolic disease in adulthood. Sci. Adv. 2018, 4, eaao4364. [Google Scholar] [CrossRef]
- Waterland, R.A.; Kellermayer, R.; Laritsky, E.; Rayco-Solon, P.; Harris, R.A.; Travisano, M.; Zhang, W.; Torskaya, M.S.; Zhang, J.; Shen, L.; et al. Season of Conception in Rural Gambia Affects DNA Methylation at Putative Human Metastable Epialleles. PLoS Genet. 2010, 6, e1001252. [Google Scholar] [CrossRef]
- James, P.T.; Dominguez-Salas, P.; Hennig, B.J.; Moore, S.E.; Prentice, A.M.; Silver, M.J. Maternal One-Carbon Metabolism and Infant DNA Methylation between Contrasting Seasonal Environments: A Case Study from The Gambia. Curr. Dev. Nutr. 2018, 3, nzy082. [Google Scholar] [CrossRef]
- Watkins, A.; Ursell, E.; Panton, R.; Papenbrock, T.; Hollis, L.; Cunningham, C.; Wilkins, A.; Perry, V.H.; Sheth, B.; Kwong, W.Y.; et al. Adaptive Responses by Mouse Early Embryos to Maternal Diet Protect Fetal Growth but Predispose to Adult Onset Disease. Biol. Reprod. 2008, 78, 299–306. [Google Scholar] [CrossRef]
- Jia, Y.; Cong, R.; Li, R.; Yang, X.; Sun, Q.; Parvizi, N.; Zhao, R. Maternal Low-Protein Diet Induces Gender-Dependent Changes in Epigenetic Regulation of the Glucose-6-Phosphatase Gene in Newborn Piglet Liver. J. Nutr. 2012, 142, 1659–1665. [Google Scholar] [CrossRef]
- Zheng, S.; Rollet, M.; Pan, Y.-X. Protein restriction during gestation alters histone modifications at the glucose transporter 4 (GLUT4) promoter region and induces GLUT4 expression in skeletal muscle of female rat offspring. J. Nutr. Biochem. 2012, 23, 1064–1071. [Google Scholar] [CrossRef]
- Ferrell, J.; Boehme, S.; Li, F.; Chiang, J.Y.L. Cholesterol 7α-hydroxylase-deficient mice are protected from high-fat/high-cholesterol diet-induced metabolic disorders. J. Lipid Res. 2016, 57, 1144–1154. [Google Scholar] [CrossRef] [PubMed]
- Sohi, G.; Marchand, K.; Revesz, A.; Arany, E.; Hardy, D.B. Maternal Protein Restriction Elevates Cholesterol in Adult Rat Offspring Due to Repressive Changes in Histone Modifications at the Cholesterol 7α-Hydroxylase Promoter. Mol. Endocrinol. 2011, 25, 785–798. [Google Scholar] [CrossRef]
- Altmann, S.; Muràni, E.; Schwerin, M.; Metges, C.C.; Wimmers, K.; Ponsuksili, S. Somatic cytochrome c (CYCS) gene expression and promoter-specific DNA methylation in a porcine model of prenatal exposure to maternal dietary protein excess and restriction. Br. J. Nutr. 2012, 107, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Yakubu, D.P.; Mostyn, A.; Hyatt, M.A.; Kurlak, L.O.; Budge, H.; Stephenson, T.; Symonds, M.E. Ontogeny and nutritional programming of mitochondrial proteins in the ovine kidney, liver and lung. Reproduction 2007, 134, 823–830. [Google Scholar] [CrossRef][Green Version]
- Wankhade, U.; Zhong, Y.; Kang, P.; Alfaro, M.; Chintapalli, S.V.; Thakali, K.M.; Shankar, K. Enhanced offspring predisposition to steatohepatitis with maternal high-fat diet is associated with epigenetic and microbiome alterations. PLoS ONE 2017, 12, e0175675. [Google Scholar] [CrossRef]
- Lihn, A.S.; Pedersen, S.B.; Richelsen, B. Adiponectin: Action, regulation and association to insulin sensitivity. Obes. Rev. 2005, 6, 13–21. [Google Scholar] [CrossRef]
- Marco, A.; Kisliouk, T.; Tabachnik, T.; Meiri, N.; Weller, A. Overweight and CpG methylation of the Pomc promoter in offspring of high-fat-diet-fed dams are not “reprogrammed” by regular chow diet in rats. FASEB J. 2014, 28, 4148–4157. [Google Scholar] [CrossRef] [PubMed]
- Fitoussi, Y.; Corbe, C.; Hamard, H.; Perdriel, G. A glaucoma follow-up of 1500 persons over 20 years. J. Fr. Ophtalmol. 1988, 11, 151–154. [Google Scholar]
- Crujeiras, A.B.; Campion, J.; Díaz-Lagares, A.; Milagro, F.; Goyenechea, E.; Abete, I.; Casanueva, F.F.; Martínez, J.A. Association of weight regain with specific methylation levels in the NPY and POMC promoters in leukocytes of obese men: A translational study. Regul. Pept. 2013, 186, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Aagaard-Tillery, K.M.; Grove, K.; Bishop, J.; Ke, X.; Fu, Q.; McKnight, R.; Lane, R.H. Developmental origins of disease and determinants of chromatin structure: Maternal diet modifies the primate fetal epigenome. J. Mol. Endocrinol. 2008, 41, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Hellman, U.; Wernstedt, C.; Eriksson, U. The Retinal Pigment Epithelial-specific 11-cis Retinol Dehydrogenase Belongs to the Family of Short Chain Alcohol Dehydrogenases. J. Biol. Chem. 1995, 270, 1107–1112. [Google Scholar] [CrossRef]
- Luo, M.; Mengos, A.E.; Stubblefield, T.M.; Mandarino, L.J. High Fat Diet-Induced Changes in Hepatic Protein Abundance in Mice. J. Proteom. Bioinform. 2012, 5, 60–66. [Google Scholar] [CrossRef]
- Masuyama, H.; Mitsui, T.; Nobumoto, E.; Hiramatsu, Y. The Effects of High-Fat Diet Exposure in Utero on the Obesogenic and Diabetogenic Traits Through Epigenetic Changes in Adiponectin and Leptin Gene Expression for Multiple Generations in Female Mice. Endocrinology 2015, 156, 2482–2491. [Google Scholar] [CrossRef] [PubMed]
- Maloyan, A.; Muralimanoharan, S.; Huffman, S.; Cox, L.A.; Nathanielsz, P.W.; Myatt, L.; Nijland, M.J. Identification and comparative analyses of myocardial miRNAs involved in the fetal response to maternal obesity. Physiol. Genom. 2013, 45, 889–900. [Google Scholar] [CrossRef]
- Sable, P.; Randhir, K.; Kale, A.; Chavan-Gautam, P.; Joshi, S. Maternal micronutrients and brain global methylation patterns in the offspring. Nutr. Neurosci. 2015, 18, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Rao, J.S.; Keleshian, V.L.; Klein, S.; Rapoport, S.I. Epigenetic modifications in frontal cortex from Alzheimer’s disease and bipolar disorder patients. Transl. Psychiatry 2012, 2, e132. [Google Scholar] [CrossRef] [PubMed]
- Ly, A.; Lee, H.; Chen, J.; Sie, K.K.Y.; Renlund, R.; Medline, A.; Sohn, K.-J.; Croxford, R.; Thompson, L.U.; Kim, Y.-I. Effect of Maternal and Postweaning Folic Acid Supplementation on Mammary Tumor Risk in the Offspring. Cancer Res. 2011, 71, 988–997. [Google Scholar] [CrossRef]
- Chang, S.; Wang, L.; Guan, Y.; Shangguan, S.; Du, Q.; Wang, Y.; Zhang, T.; Zhang, Y. Long interspersed nucleotide element-1 hypomethylation in folate-deficient mouse embryonic stem cells. J. Cell. Biochem. 2013, 114, 1549–1558. [Google Scholar] [CrossRef]
- Marsit, C.J. Influence of environmental exposure on human epigenetic regulation. J. Exp. Biol. 2015, 218, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Hopenhayn, C.; Ferreccio, C.; Browning, S.R.; Huang, B.; Peralta, C.; Gibb, H.; Hertz-Picciotto, I. Arsenic Exposure from Drinking Water and Birth Weight. Epidemiology 2013, 14, 593–602. [Google Scholar] [CrossRef]
- Sohel, N.; Vahter, M.; Ali, M.; Rahman, M.; Rahman, A.; Streatfield, P.K.; Kanaroglou, P.S.; Persson, L. Spatial patterns of fetal loss and infant death in an arsenic-affected area in Bangladesh. Int. J. Health Geogr. 2010, 9, 53. [Google Scholar] [CrossRef]
- Jin, Z.; Liu, Y. DNA methylation in human diseases. Genes Dis. 2018, 5, 1–8. [Google Scholar] [CrossRef]
- Greco, E.A.; Lenzi, A.; Migliaccio, S.; Gessani, S. Epigenetic Modifications Induced by Nutrients in Early Life Phases: Gender Differences in Metabolic Alteration in Adulthood. Front. Genet. 2019, 10, 795. [Google Scholar] [CrossRef]
- Shi, L.; Wu, J. Epigenetic regulation in mammalian preimplantation embryo development. Reprod. Biol. Endocrinol. 2009, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, Z.; Jia, J.; Du, T.; Zhang, N.; Tang, Y.; Fang, Y.; Fang, D. Overview of Histone Modification. Adv. Exp. Med. Biol. 2021, 1283, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Vakoc, C.; Mandat, S.A.; Olenchock, B.A.; Blobel, G.A. Histone H3 Lysine 9 Methylation and HP1γ Are Associated with Transcription Elongation through Mammalian Chromatin. Mol. Cell 2005, 19, 381–391. [Google Scholar] [CrossRef]
- Morgan, H.D.; Santos, F.; Green, K.; Dean, W.; Reik, W. Epigenetic reprogramming in mammals. Hum. Mol. Genet. 2005, 14, R47–R58. [Google Scholar] [CrossRef]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv. Exp. Med. Biol. 2020, 1253, 3–55. [Google Scholar] [CrossRef]
- Kurup, J.T.; Han, Z.; Jin, W.; Kidder, B.L. H4K20me3 methyltransferase SUV420H2 shapes the chromatin landscape of pluripotent embryonic stem cells. Development 2020, 147, dev188516. [Google Scholar] [CrossRef] [PubMed]
- Dovey, O.M.; Foster, C.T.; Cowley, S.M. Histone deacetylase 1 (HDAC1), but not HDAC2, controls embryonic stem cell differentiation. Proc. Natl. Acad. Sci. USA 2010, 107, 8242–8247. [Google Scholar] [CrossRef]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA Translation and Stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef]
- Todeschini, A.-L.; Teysset, L.; Delmarre, V.; Ronsseray, S. The Epigenetic Trans-Silencing Effect in Drosophila Involves Maternally-Transmitted Small RNAs Whose Production Depends on the piRNA Pathway and HP1. PLoS ONE 2010, 5, e11032. [Google Scholar] [CrossRef]
- Kawasaki, H.; Taira, K.; Morris, K. siRNA Induced Transcriptional Gene Silencing in Mammalian Cells. Cell Cycle 2005, 4, 442–448. [Google Scholar] [CrossRef]
- Bouckenheimer, J.; Assou, S.; Riquier, S.; Hou, C.; Philippe, N.; Sansac, C.; Lavabre-Bertrand, T.; Commes, T.; Lemaître, J.-M.; Boureux, A.; et al. Long non-coding RNAs in human early embryonic development and their potential in ART. Hum. Reprod. Update 2016, 23, 19–40. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Kwon, J.T.; Kim, J.; Jeong, J.; Kim, J.; Lee, S.; Cho, C. Profiling of testis-specific long noncoding RNAs in mice. BMC Genom. 2018, 19, 539. [Google Scholar] [CrossRef]
- Liang, M.; Li, W.; Tian, H.; Hu, T.; Wang, L.; Lin, Y.; Li, Y.; Huang, H.; Sun, F. Sequential expression of long noncoding RNA as mRNA gene expression in specific stages of mouse spermatogenesis. Sci. Rep. 2014, 4, srep05966. [Google Scholar] [CrossRef]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol. 2012, 14, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Kraus, P.; Sivakamasundari, V.; Lim, S.L.; Xing, X.; Lipovich, L.; Lufkin, T. Making sense of Dlx1 antisense RNA. Dev. Biol. 2013, 376, 224–235. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Luk, A.C.-S.; Chan, W.Y.; Rennert, O.M.; Lee, T.-L. Long noncoding RNAs in spermatogenesis: Insights from recent high-throughput transcriptome studies. Reproduction 2014, 147, R131–R141. [Google Scholar] [CrossRef]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.; Ganai, N. Long non-coding RNAs: Mechanism of action and functional utility. Non-Coding RNA Res. 2016, 1, 43–50. [Google Scholar] [CrossRef]
- Wang, J.; Wu, Z.; Li, D.; Li, N.; Dindot, S.V.; Satterfield, M.C.; Bazer, F.W.; Wu, G. Nutrition, Epigenetics, and Metabolic Syndrome. Antioxid. Redox Signal. 2012, 17, 282–301. [Google Scholar] [CrossRef]
- Heindel, J.J.; Vandenberg, L.N. Developmental origins of health and disease: A paradigm for understanding disease etiology and prevention. Curr. Opin. Pediatr. 2015, 27, 248–253. [Google Scholar] [CrossRef]
- Hoffman, D.J.; Reynolds, R.; Hardy, D.B. Developmental origins of health and disease: Current knowledge and potential mechanisms. Nutr. Rev. 2017, 75, 951–970. [Google Scholar] [CrossRef] [PubMed]
- Elias, S.G.; Peeters, P.H.M.; Grobbee, D.E.; van Noord, P.A.H. Transient caloric restriction and cancer risk (The Netherlands). Cancer Causes Control 2007, 18, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Roseboom, T.J. Epidemiological evidence for the developmental origins of health and disease: Effects of prenatal undernutrition in humans. J. Endocrinol. 2019, 242, T135–T144. [Google Scholar] [CrossRef]
- Roseboom, T.; De Rooij, S.; Painter, R. The Dutch famine and its long-term consequences for adult health. Early Hum. Dev. 2006, 82, 485–491. [Google Scholar] [CrossRef]
- Van Abeelen, A.; de Rooij, S.; Osmond, C.; Painter, R.; Veenendaal, M.; Bossuyt, P.; Elias, S.; Grobbee, D.; van der Schouw, Y.; Barker, D.; et al. The sex-specific effects of famine on the association between placental size and later hypertension. Placenta 2011, 32, 694–698. [Google Scholar] [CrossRef]
- Beharry, Z.; Mahajan, S.; Zemskova, M.; Lin, Y.-W.; Tholanikunnel, B.G.; Xia, Z.; Smith, C.D.; Kraft, A.S. The Pim protein kinases regulate energy metabolism and cell growth. Proc. Natl. Acad. Sci. USA 2011, 108, 528–533. [Google Scholar] [CrossRef]
- Hoek, H.W.; Susser, E.; Buck, K.A.; Lumey, L.H.; Lin, S.P.; Gorman, J.M. Schizoid personality disorder after prenatal exposure to famine. Am. J. Psychiatry 1996, 153, 1637–1639. [Google Scholar] [CrossRef]
- Kyle, U.G.; Pichard, C. The Dutch Famine of 1944–1945: A pathophysiological model of long-term consequences of wasting disease. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 388–394. [Google Scholar] [CrossRef]
- Quilter, C.R.; Harvey, K.M.; Bauer, J.; Skinner, B.M.; Gomez, M.; Shrivastava, M.; Doel, A.M.; Drammeh, S.; Dunger, D.B.; Moore, S.E.; et al. Identification of methylation changes associated with positive and negative growth deviance in Gambian infants using a targeted methyl sequencing approach of genomic DNA. FASEB BioAdv. 2021, 3, 205–230. [Google Scholar] [CrossRef]
- Moore, S.E.; Cole, T.J.; Poskitt, E.M.E.; Sonko, B.J.; Whitehead, R.G.; McGregor, I.A.; Prentice, A.M. Season of birth predicts mortality in rural Gambia. Nature 1997, 388, 434. [Google Scholar] [CrossRef] [PubMed]
- Bernal, A.J.; Jirtle, R.L. Epigenomic disruption: The effects of early developmental exposures. Birth Defects Res. Part A Clin. Mol. Teratol. 2010, 88, 938–944. [Google Scholar] [CrossRef]
- Dolinoy, D.C.; Jirtle, R.L. Environmental epigenomics in human health and disease. Environ. Mol. Mutagen. 2008, 49, 4–8. [Google Scholar] [CrossRef]
- Dominguez-Salas, P.; Moore, S.E.; Baker, M.S.; Bergen, A.; Cox, S.; Dyer, R.A.; Fulford, A.J.; Guan, Y.; Laritsky, E.; Silver, M.; et al. Maternal nutrition at conception modulates DNA methylation of human metastable epialleles. Nat. Commun. 2014, 5, 3746. [Google Scholar] [CrossRef]
- Khulan, B.; Cooper, W.N.; Skinner, B.; Bauer, J.; Owens, S.; Prentice, A.M.; Belteki, G.; Constancia, M.; Dunger, P.D.; Affara, N.A. Periconceptional maternal micronutrient supplementation is associated with widespread gender related changes in the epigenome: A study of a unique resource in the Gambia. Hum. Mol. Genet. 2012, 21, 2086–2101. [Google Scholar] [CrossRef]
- Moore, S.E. Early life nutritional programming of health and disease in the Gambia. J. Dev. Orig. Health Dis. 2016, 7, 123–131. [Google Scholar] [CrossRef]
- Fleming, T.P.; Eckert, J.J.; Denisenko, O. The Role of Maternal Nutrition during the Periconceptional Period and Its Effect on Offspring Phenotype. Adv. Exp. Med. Biol. 2017, 1014, 87–105. [Google Scholar] [CrossRef]
- Bar-Sadeh, B.; Rudnizky, S.; Pnueli, L.; Bentley, G.R.; Stöger, R.; Kaplan, A.; Melamed, P. Unravelling the role of epigenetics in reproductive adaptations to early-life environment. Nat. Rev. Endocrinol. 2020, 16, 519–533. [Google Scholar] [CrossRef]
- Chamson-Reig, A.; Arany, E.J.; Summers, K.; Hill, D.J. A low protein diet in early life delays the onset of diabetes in the non-obese diabetic mouse. J. Endocrinol. 2009, 201, 231–239. [Google Scholar] [CrossRef]
- Eckert, J.J.; Porter, R.; Watkins, A.J.; Burt, E.; Brooks, S.; Leese, H.J.; Humpherson, P.G.; Cameron, I.T.; Fleming, T.P. Metabolic Induction and Early Responses of Mouse Blastocyst Developmental Programming following Maternal Low Protein Diet Affecting Life-Long Health. PLoS ONE 2012, 7, e52791. [Google Scholar] [CrossRef]
- Fleming, T.P.; Watkins, A.J.; Sun, C.; Velazquez, M.A.; Smyth, N.R.; Eckert, J.J. Do little embryos make big decisions? How maternal dietary protein restriction can permanently change an embryo’s potential, affecting adult health. Reprod. Fertil. Dev. 2015, 27, 684–692. [Google Scholar] [CrossRef]
- Velazquez, M.A.; Sheth, B.; Smith, S.J.; Eckert, J.J.; Osmond, C.; Fleming, T.P. Insulin and branched-chain amino acid depletion during mouse preimplantation embryo culture programmes body weight gain and raised blood pressure during early postnatal life. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1864, 590–600. [Google Scholar] [CrossRef]
- Gould, J.; Smith, P.J.; Airey, C.J.; Mort, E.J.; Airey, L.E.; Warricker, F.D.M.; Pearson-Farr, J.E.; Weston, E.C.; Gould, P.J.W.; Semmence, O.G.; et al. Mouse maternal protein restriction during preimplantation alone permanently alters brain neuron proportion and adult short-term memory. Proc. Natl. Acad. Sci. USA 2018, 115, E7398–E7407. [Google Scholar] [CrossRef]
- Jousse, C.; Parry, L.; Lambert-Langlais, S.; Maurin, A.; Averous, J.; Bruhat, A.; Carraro, V.; Tost, J.; Letteron, P.; Chen, P.; et al. Perinatal undernutrition affects the methylation and expression of the leptin gene in adults: Implication for the understanding of metabolic syndrome. FASEB J. 2011, 25, 3271–3278. [Google Scholar] [CrossRef]
- Gruzdeva, O.; Borodkina, D.; Uchasova, E.; Dyleva, Y.; Barbarash, O. Leptin resistance: Underlying mechanisms and diagnosis. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 191–198. [Google Scholar] [CrossRef]
- Painter, R.C.; De Rooij, S.R.; Bossuyt, P.M.; Simmers, T.A.; Osmond, C.; Barker, D.J.; Bleker, O.P.; Roseboom, T.J. Early onset of coronary artery disease after prenatal exposure to the Dutch famine. Am. J. Clin. Nutr. 2006, 84, 322–327. [Google Scholar] [CrossRef]
- Hossain, P.; Kawar, B.; El Nahas, M. Obesity and Diabetes in the Developing World—A Growing Challenge. N. Engl. J. Med. 2007, 356, 213–215. [Google Scholar] [CrossRef]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef]
- Kominiarek, M.A.; Peaceman, A.M. Gestational weight gain. Am. J. Obstet. Gynecol. 2017, 217, 642–651. [Google Scholar] [CrossRef]
- Grissom, N.; Bowman, N.; Reyes, T.M. Epigenetic programming of reward function in offspring: A role for maternal diet. Mamm. Genome 2014, 25, 41–48. [Google Scholar] [CrossRef]
- Şanlı, E.; Kabaran, S. Maternal Obesity, Maternal Overnutrition and Fetal Programming: Effects of Epigenetic Mechanisms on the Development of Metabolic Disorders. Curr. Genom. 2019, 20, 419–427. [Google Scholar] [CrossRef]
- Sarker, G.; Litwan, K.; Kastli, R.; Peleg-Raibstein, D. Maternal overnutrition during critical developmental periods leads to different health adversities in the offspring: Relevance of obesity, addiction and schizophrenia. Sci. Rep. 2019, 9, 17322. [Google Scholar] [CrossRef] [PubMed]
- Khalyfa, A.; Carreras, A.; Hakim, F.; Cunningham, J.M.; Wang, Y.; Gozal, D. Effects of late gestational high-fat diet on body weight, metabolic regulation and adipokine expression in offspring. Int. J. Obes. 2013, 37, 1481–1489. [Google Scholar] [CrossRef]
- Chen, J.; Wang, D.-Z. microRNAs in cardiovascular development. J. Mol. Cell. Cardiol. 2012, 52, 949–957. [Google Scholar] [CrossRef]
- Anderson, O.S.; Sant, K.; Dolinoy, D.C. Nutrition and epigenetics: An interplay of dietary methyl donors, one-carbon metabolism and DNA methylation. J. Nutr. Biochem. 2012, 23, 853–859. [Google Scholar] [CrossRef]
- Chamberlain, J.A.; Dugue, P.-A.; Bassett, J.K.; Hodge, A.; Brinkman, M.T.; Joo, J.E.; Jung, C.-H.; Makalic, E.; Schmidt, D.F.; Hopper, J.L.; et al. Dietary intake of one-carbon metabolism nutrients and DNA methylation in peripheral blood. Am. J. Clin. Nutr. 2018, 108, 611–621. [Google Scholar] [CrossRef]
- Steegers-Theunissen, R.P.; Twigt, J.; Pestinger, V.; Sinclair, K.D. The periconceptional period, reproduction and long-term health of offspring: The importance of one-carbon metabolism. Hum. Reprod. Update 2013, 19, 640–655. [Google Scholar] [CrossRef]
- Friso, S.; Udali, S.; De Santis, D.; Choi, S.-W. One-carbon metabolism and epigenetics. Mol. Asp. Med. 2017, 54, 28–36. [Google Scholar] [CrossRef]
- Kalhan, S.C. One carbon metabolism in pregnancy: Impact on maternal, fetal and neonatal health. Mol. Cell. Endocrinol. 2016, 435, 48–60. [Google Scholar] [CrossRef]
- Dominguez-Salas, P.; Moore, S.E.; Cole, D.; Da Costa, K.-A.; Cox, S.; Dyer, R.A.; Fulford, A.J.C.; Innis, S.M.; Waterland, R.A.; Zeisel, S.H.; et al. DNA methylation potential: Dietary intake and blood concentrations of one-carbon metabolites and cofactors in rural African women. Am. J. Clin. Nutr. 2013, 97, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Kalhan, S.C.; Marczewski, S.E. Methionine, homocysteine, one carbon metabolism and fetal growth. Rev. Endocr. Metab. Disord. 2012, 13, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, K.D.; Allegrucci, C.; Singh, R.; Gardner, D.S.; Sebastian, S.; Bispham, J.; Thurston, A.; Huntley, J.F.; Rees, W.D.; Maloney, C.; et al. DNA methylation, insulin resistance, and blood pressure in offspring determined by maternal periconceptional B vitamin and methionine status. Proc. Natl. Acad. Sci. USA 2007, 104, 19351–19356. [Google Scholar] [CrossRef]
- Blom, H.J. Folic acid, methylation and neural tube closure in humans. Birth Defects Res. Part A Clin. Mol. Teratol. 2009, 85, 295–302. [Google Scholar] [CrossRef]
- Imbard, A.; Benoist, J.-F.; Blom, H.J. Neural Tube Defects, Folic Acid and Methylation. Int. J. Environ. Res. Public Health 2013, 10, 4352–4389. [Google Scholar] [CrossRef]
- Steegers-Theunissen, R.P.; Obermann-Borst, S.A.; Kremer, D.; Lindemans, J.; Siebel, C.; Steegers, E.A.; Slagboom, P.E.; Heijmans, B.T. Periconceptional Maternal Folic Acid Use of 400 µg per Day Is Related to Increased Methylation of the IGF2 Gene in the Very Young Child. PLoS ONE 2009, 4, e7845. [Google Scholar] [CrossRef]
- Cappuccilli, M.; Bergamini, C.; Giacomelli, F.A.; Cianciolo, G.; Donati, G.; Conte, D.; Natali, T.; La Manna, G.; Capelli, I. Vitamin B Supplementation and Nutritional Intake of Methyl Donors in Patients with Chronic Kidney Disease: A Critical Review of the Impact on Epigenetic Machinery. Nutrients 2020, 12, 1234. [Google Scholar] [CrossRef]
- Crider, K.S.; Yang, T.P.; Berry, R.; Bailey, L.B. Folate and DNA Methylation: A Review of Molecular Mechanisms and the Evidence for Folate’s Role. Adv. Nutr. 2012, 3, 21–38. [Google Scholar] [CrossRef]
- Ono, H.; Iwasaki, M.; Kuchiba, A.; Kasuga, Y.; Yokoyama, S.; Onuma, H.; Nishimura, H.; Kusama, R.; Ohnami, S.; Sakamoto, H.; et al. Association of dietary and genetic factors related to one-carbon metabolism with global methylation level of leukocyte DNA. Cancer Sci. 2012, 103, 2159–2164. [Google Scholar] [CrossRef]
- Parisi, F.; Rousian, M.; Koning, A.H.; Willemsen, S.P.; Cetin, I.; Steegers, E.A.; Steegers-Theunissen, R.P. Periconceptional maternal biomarkers of one-carbon metabolism and embryonic growth trajectories: The Rotterdam Periconceptional Cohort (Predict Study). Fertil. Steril. 2017, 107, 691–698. [Google Scholar] [CrossRef]
- Twigt, J.M.; Bezstarosti, K.; Demmers, J.; Lindemans, J.; Laven, J.S.E.; Steegers-Theunissen, R.P. Preconception folic acid use influences the follicle fluid proteome. Eur. J. Clin. Investig. 2015, 45, 833–841. [Google Scholar] [CrossRef]
- Van Mil, N.H.; Bouwland-Both, M.I.; Stolk, L.; Verbiest, M.M.P.J.; Hofman, A.; Jaddoe, V.W.V.; Verhulst, F.C.; Eilers, P.H.C.; Uitterlinden, A.G.; Steegers, E.A.P.; et al. Determinants of maternal pregnancy one-carbon metabolism and newborn human DNA methylation profiles. Reproduction 2014, 148, 581–592. [Google Scholar] [CrossRef]
- Caton, J.S.; Crouse, M.; Reynolds, L.P.; Neville, T.L.; Dahlen, C.R.; Ward, A.K.; Swanson, K.C. Maternal nutrition and programming of offspring energy requirements. Transl. Anim. Sci. 2019, 3, 976–990. [Google Scholar] [CrossRef] [PubMed]
- Diniz, W.; Reynolds, L.; Borowicz, P.; Ward, A.; Sedivec, K.; McCarthy, K.; Kassetas, C.; Baumgaertner, F.; Kirsch, J.; Dorsam, S.; et al. Maternal Vitamin and Mineral Supplementation and Rate of Maternal Weight Gain Affects Placental Expression of Energy Metabolism and Transport-Related Genes. Genes 2021, 12, 385. [Google Scholar] [CrossRef]
- Canovas, S.; Ross, P.J. Epigenetics in preimplantation mammalian development. Theriogenology 2016, 86, 69–79. [Google Scholar] [CrossRef]
- Fleming, T.P.; Watkins, A.J.; Velazquez, M.A.; Mathers, J.C.; Prentice, A.M.; Stephenson, J.; Barker, M.; Saffery, R.; Yajnik, C.S.; Eckert, J.J.; et al. Origins of lifetime health around the time of conception: Causes and consequences. Lancet 2018, 391, 1842–1852. [Google Scholar] [CrossRef]
- Sinclair, K.D.; Karamitri, A.; Gardner, D.S. Dietary regulation of developmental programming in ruminants: Epigenetic modifications in the germline. Soc. Reprod. Fertil. Suppl. 2010, 67, 59–72. [Google Scholar] [CrossRef]
- Neel, J.V. Diabetes mellitus: A “thrifty” genotype rendered detrimental by “progress”? Am. J. Hum. Genet. 1962, 14, 353–362. [Google Scholar]
- Paradies, Y.; Montoya, M.J.; Fullerton, S.M. Racialized Genetics and the Study of Complex Diseases: The Thrifty Genotype Revisited. Perspect. Biol. Med. 2007, 50, 203–227. [Google Scholar] [CrossRef]
- O’Rourke, R.W. Metabolic Thrift and the Genetic Basis of Human Obesity. Ann. Surg. 2014, 259, 642–648. [Google Scholar] [CrossRef]
- Venniyoor, A. PTEN: A Thrifty Gene That Causes Disease in Times of Plenty? Front. Nutr. 2020, 7, 81. [Google Scholar] [CrossRef]
- Gensous, N.; Ravaioli, F.; Pirazzini, C.; Gramignoli, R.; Ellis, E.; Storci, G.; Capri, M.; Strom, S.; Laconi, E.; Franceschi, C.; et al. Aging and Caloric Restriction Modulate the DNA Methylation Profile of the Ribosomal RNA Locus in Human and Rat Liver. Nutrients 2020, 12, 277. [Google Scholar] [CrossRef] [PubMed]
- Kusnadi, E.P.; Hannan, K.; Hicks, R.; Hannan, R.; Pearson, R.B.; Kang, J. Regulation of rDNA transcription in response to growth factors, nutrients and energy. Gene 2015, 556, 27–34. [Google Scholar] [CrossRef]
- Denisenko, O.; Lucas, E.S.; Sun, C.; Watkins, A.J.; Mar, D.; Bomsztyk, K.; Fleming, T.P. Regulation of ribosomal RNA expression across the lifespan is fine-tuned by maternal diet before implantation. Biochim. Biophys. Acta (BBA)-Gene Regul. Mech. 2016, 1859, 906–913. [Google Scholar] [CrossRef]
- Holland, M.L.; Lowe, R.; Caton, P.W.; Gemma, C.; Carbajosa, G.; Danson, A.F.; Carpenter, A.A.M.; Loche, E.; Ozanne, S.E.; Rakyan, V.K. Early-life nutrition modulates the epigenetic state of specific rDNA genetic variants in mice. Science 2016, 353, 495–498. [Google Scholar] [CrossRef]
- Bonakdar, E.; Edriss, M.A.; Bakhtari, A.; Jafarpour, F.; Asgari, V.; Hosseini, S.M.; Boroujeni, N.S.; Hajian, M.; Rahmani, H.R.; Nasr-Esfahani, M.H. A physiological, rather than a superovulated, post-implantation environment can attenuate the compromising effect of assisted reproductive techniques on gene expression in developing mice embryos. Mol. Reprod. Dev. 2015, 82, 191–206. [Google Scholar] [CrossRef]
- Farquhar, C.; Marjoribanks, J. Assisted reproductive technology: An overview of Cochrane Reviews. Cochrane Database Syst. Rev. 2018, 8, CD010537. [Google Scholar] [CrossRef]
- Mani, S.; Ghosh, J.; Coutifaris, C.; Sapienza, C.; Mainigi, M. Epigenetic changes and assisted reproductive technologies. Epigenetics 2020, 15, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.; Brown, L.; Brown, G.; Casson, P.; Brown, S. Microarray assessment of methylation in individual mouse blastocyst stage embryos shows that in vitro culture may have widespread genomic effects. Hum. Reprod. 2011, 26, 2576–2585. [Google Scholar] [CrossRef][Green Version]
- Santos, F.; Hyslop, L.; Stojkovic, P.; Leary, C.; Murdoch, A.; Reik, W.; Stojkovic, M.; Herbert, M.; Dean, W. Evaluation of epigenetic marks in human embryos derived from IVF and ICSI. Hum. Reprod. 2010, 25, 2387–2395. [Google Scholar] [CrossRef] [PubMed]
- Fauque, P.; Jouannet, P.; Lesaffre, C.; Ripoche, M.-A.; Dandolo, L.; Vaiman, D.; Jammes, H. Assisted Reproductive Technology affects developmental kinetics, H19 Imprinting Control Region methylation and H19 gene expression in individual mouse embryos. BMC Dev. Biol. 2007, 7, 116. [Google Scholar] [CrossRef]
- Li, B.; Chen, S.; Tang, N.; Xiao, X.; Huang, J.; Jiang, F.; Huang, X.; Sun, F.; Wang, X. Assisted Reproduction Causes Reduced Fetal Growth Associated with Downregulation of Paternally Expressed Imprinted Genes That Enhance Fetal Growth in Mice. Biol. Reprod. 2016, 94, 45. [Google Scholar] [CrossRef] [PubMed]
- Hattori, H.; Hiura, H.; Kitamura, A.; Miyauchi, N.; Kobayashi, N.; Takahashi, S.; Okae, H.; Kyono, K.; Kagami, M.; Ogata, T.; et al. Association of four imprinting disorders and ART. Clin. Epigenet. 2019, 11, 21. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ma, Y.; Wen, L.; Lei, H.; Chen, S.; Wang, X. Changes in DNA methylation and imprinting disorders in E9.5 mouse fetuses and placentas derived from vitrified eight-cell embryos. Mol. Reprod. Dev. 2019, 86, 404–415. [Google Scholar] [CrossRef]
- Melamed, N.; Choufani, S.; Wilkins-Haug, L.E.; Koren, G.; Weksberg, R. Comparison of genome-wide and gene-specific DNA methylation between ART and naturally conceived pregnancies. Epigenetics 2015, 10, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Chi, F.; Zhao, M.; Li, K.; Lin, A.-Q.; Li, Y.; Teng, X. DNA methylation status of imprinted H19 and KvDMR1 genes in human placentas after conception using assisted reproductive technology. Ann. Transl. Med. 2020, 8, 854. [Google Scholar] [CrossRef]
- Tobi, E.W.; Almqvist, C.; Hedman, A.; Andolf, E.; Holte, J.; Olofsson, J.I.; Wramsby, H.; Wramsby, M.; Pershagen, G.; Heijmans, B.T.; et al. DNA methylation differences at birth after conception through ART. Hum. Reprod. 2021, 36, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Ghosh, J.; Mainigi, M.; Turan, N.; Weinerman, R.; Truongcao, M.; Coutifaris, C.; Sapienza, C. DNA methylation differences between in vitro- and in vivo-conceived children are associated with ART procedures rather than infertility. Clin. Epigenet. 2015, 7, 1–10. [Google Scholar] [CrossRef]
- Shi, X.; Chen, S.; Zheng, H.; Wang, L.; Wu, Y. Aberrant DNA methylation of imprinted loci in human in vitro matured oocytes after long agonist stimulation. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 167, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Chen, S.; Zheng, H.; Wang, L.; Wu, Y. Abnormal DNA Methylation of Imprinted Loci in Human Preimplantation Embryos. Reprod. Sci. 2014, 21, 978–983. [Google Scholar] [CrossRef]
- Binder, G.; Begemann, M.; Eggermann, T.; Kannenberg, K. Silver–Russell syndrome. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 153–160. [Google Scholar] [CrossRef]
- Cocchi, G.; Marsico, C.; Cosentino, A.; Spadoni, C.; Rocca, A.; De Crescenzo, A.; Riccio, A. Silver-Russell syndrome due to paternal H19/IGF2 hypomethylation in a twin girl born after in vitro fertilization. Am. J. Med. Genet. Part A 2013, 161, 2652–2655. [Google Scholar] [CrossRef][Green Version]
- Krzyzewska, I.; Alders, M.; Maas, S.M.; Bliek, J.; Venema, A.; Henneman, P.; Rezwan, F.I.; Lip, K.V.D.; Mul, A.N.; Mackay, D.J.; et al. Genome-wide methylation profiling of Beckwith-Wiedemann syndrome patients without molecular confirmation after routine diagnostics. Clin. Epigenet. 2019, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Pandita, A.; Gupta, S.; Gupta, G.; Panghal, A. Beckwith-Weidemann syndrome with IC2 (KvDMR1) hypomethylation defect: A novel mutation. BMJ Case Rep. 2018, 2018, bcr-2017. [Google Scholar] [CrossRef]
- Elalaoui, S.; Garin, I.; Sefiani, A.; De Nanclares, G.P. Maternal Hypomethylation of KvDMR in a Monozygotic Male Twin Pair Discordant for Beckwith-Wiedemann Syndrome. Mol. Syndr. 2014, 5, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Choux, C.; Binquet, C.; Carmignac, V.; Bruno, C.; Chapusot, C.; Barberet, J.; LaMotte, M.; Sagot, P.; Bourc’His, D.; Fauque, P. The epigenetic control of transposable elements and imprinted genes in newborns is affected by the mode of conception: ART versus spontaneous conception without underlying infertility. Hum. Reprod. 2018, 33, 331–340. [Google Scholar] [CrossRef]
- Vincent, R.N.; Gooding, L.D.; Louie, K.; Wong, E.C.; Ma, S. Altered DNA methylation and expression of PLAGL1 in cord blood from assisted reproductive technology pregnancies compared with natural conceptions. Fertil. Steril. 2016, 106, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Novakovic, B.; Lewis, S.; Halliday, J.; Kennedy, J.; Burgner, D.P.; Czajko, A.; Kim, B.; Sexton-Oates, A.; Juonala, M.; Hammarberg, K.; et al. Assisted reproductive technologies are associated with limited epigenetic variation at birth that largely resolves by adulthood. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Rexhaj, E.; Pireva, A.; Paoloni-Giacobino, A.; Allemann, Y.; Cerny, D.; Dessen, P.; Sartori, C.; Scherrer, U.; Rimoldi, S.F. Prevention of vascular dysfunction and arterial hypertension in mice generated by assisted reproductive technologies by addition of melatonin to culture media. Am. J. Physiol. Circ. Physiol. 2015, 309, H1151–H1156. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Y.; Huang, H.; Guan, Y.; Li, M.; Jiang, X.; Yu, M.; Yang, X. Effects of superovulation, in vitro fertilization, and oocyte in vitro maturation on imprinted gene Grb10 in mouse blastocysts. Arch. Gynecol. Obstet. 2018, 298, 1219–1227. [Google Scholar] [CrossRef]
- Bakhtari, A.; Rahmani, H.-R.; Bonakdar, E.; Jafarpour, F.; Asgari, V.; Hosseini, S.-M.; Hajian, M.; Edriss, M.-A.; Nasr-Esfahani, M.-H. The interfering effects of superovulation and vitrification upon some important epigenetic biomarkers in mouse blastocyst. Cryobiology 2014, 69, 419–427. [Google Scholar] [CrossRef]
- Li, T.; Vu, T.H.; Ulaner, G.A.; Littman, E.; Ling, J.-Q.; Chen, H.-L.; Hu, J.-F.; Behr, B.; Giudice, L.; Hoffman, A.R. IVF results in de novo DNA methylation and histone methylation at an Igf2-H19 imprinting epigenetic switch. Mol. Hum. Reprod. 2005, 11, 631–640. [Google Scholar] [CrossRef]
- De Waal, E.; Vrooman, L.A.; Fischer, E.; Ord, T.; Mainigi, M.; Coutifaris, C.; Schultz, R.M.; Bartolomei, M.S. The cumulative effect of assisted reproduction procedures on placental development and epigenetic perturbations in a mouse model. Hum. Mol. Genet. 2015, 24, 6975–6985. [Google Scholar] [CrossRef]
- Rahimi, S.; Martel, J.; Karahan, G.; Angle, C.; Behan, N.A.; Chan, D.; Macfarlane, A.J.; Trasler, J.M. Moderate maternal folic acid supplementation ameliorates adverse embryonic and epigenetic outcomes associated with assisted reproduction in a mouse model. Hum. Reprod. 2019, 34, 851–862. [Google Scholar] [CrossRef]
- Chen, S.; Sun, F.-Z.; Huang, X.; Wang, X.; Tang, N.; Zhu, B.; Li, B. Assisted reproduction causes placental maldevelopment and dysfunction linked to reduced fetal weight in mice. Sci. Rep. 2015, 5, srep10596. [Google Scholar] [CrossRef]
- Rexhaj, E.; Paoloni-Giacobino, A.; Rimoldi, S.F.; Fuster, D.G.; Anderegg, M.; Somm, E.; Bouillet, E.; Allemann, Y.; Sartori, C.; Scherrer, U. Mice generated by in vitro fertilization exhibit vascular dysfunction and shortened life span. J. Clin. Investig. 2013, 123, 5052–5060. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Alcaraz, B.; Crispi, F.; Bijnens, B.; Cruz-Lemini, M.; Creus, M.; Sitges, M.; Bartrons, J.; Civico, S.; Balasch, J.; Gratacós, E. Assisted Reproductive Technologies Are Associated with Cardiovascular Remodeling in Utero That Persists Postnatally. Circulation 2013, 128, 1442–1450. [Google Scholar] [CrossRef] [PubMed]
- Le, F.; Wang, N.; Wang, Q.; Yang, X.; Li, L.; Wang, L.; Liu, X.; Hu, M.; Jin, F.; Lou, H. Long-Term Disturbed Expression and DNA Methylation of SCAP/SREBP Signaling in the Mouse Lung From Assisted Reproductive Technologies. Front. Genet. 2021, 12, 566168. [Google Scholar] [CrossRef] [PubMed]
- Panth, N.; Gavarkovs, A.; Tamez, M.; Mattei, J. The Influence of Diet on Fertility and the Implications for Public Health Nutrition in the United States. Front. Public Health 2018, 6, 211. [Google Scholar] [CrossRef]
- Chavarro, J.E.; Rich-Edwards, J.W.; Rosner, B.A.; Willett, W.C. Protein intake and ovulatory infertility. Am. J. Obstet. Gynecol. 2008, 198, 210.e1–210.e7. [Google Scholar] [CrossRef]
- Gaskins, A.J.; Afeiche, M.C.; Wright, D.L.; Toth, T.L.; Williams, P.L.; Gillman, M.W.; Hauser, R.; Chavarro, J. Dietary Folate and Reproductive Success among Women Undergoing Assisted Reproduction. Obstet. Gynecol. 2014, 124, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Lin, Y.; Lin, D.; Zou, C.; Zou, X.; Fu, L.; Meng, F.; Qian, W. Mediterranean diet improves embryo yield in IVF: A prospective cohort study. Reprod. Biol. Endocrinol. 2019, 17, 73. [Google Scholar] [CrossRef]
- Alibeigi, Z.; Jafari-Dehkordi, E.; Kheiri, S.; Nemati, M.; Mohammadi-Farsani, G.; Tansaz, M. Auswirkungen einer auf traditioneller Medizin basierenden Ernahrung und Lebensfuhrung auf die Infertilitatsbehandlung bei Frauen, die sich Masnahmen zur assistierten Reproduktion unterziehen: Eine randomisierte kontrollierte Studie. Complement. Med. Res. 2020, 27, 230–241. [Google Scholar] [CrossRef]
- Jahangirifar, M.; Askari, G.; Taebi, M. Dietary Patterns and the Outcomes of Assisted Reproductive Techniques in Women with Primary Infertility: A Prospective Cohort Study. Int. J. Fertil. Steril. 2019, 12, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Gaskins, A.J.; Nassan, F.L.; Chiu, Y.-H.; Arvizu, M.; Williams, P.L.; Keller, M.G.; Souter, I.; Hauser, R.; Chavarro, J.E. Dietary patterns and outcomes of assisted reproduction. Am. J. Obstet. Gynecol. 2019, 220, 567.e1–567.e18. [Google Scholar] [CrossRef]
- Karayiannis, D.; Kontogianni, M.; Mendorou, C.; Mastrominas, M.; Yiannakouris, N. Adherence to the Mediterranean diet and IVF success rate among non-obese women attempting fertility. Hum. Reprod. 2018, 33, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic, M.; De Vries, J.H.; Lindemans, J.; Macklon, N.S.; Van Der Spek, P.J.; Steegers, E.A.; Steegers-Theunissen, R.P. The preconception Mediterranean dietary pattern in couples undergoing in vitro fertilization/intracytoplasmic sperm injection treatment increases the chance of pregnancy. Fertil. Steril. 2010, 94, 2096–2101. [Google Scholar] [CrossRef]
- Twigt, J.M.; Bolhuis, M.E.C.; Steegers, E.A.P.; Hammiche, F.; Van Inzen, W.G.; Laven, J.S.E.; Steegers-Theunissen, R.P.M. The preconception diet is associated with the chance of ongoing pregnancy in women undergoing IVF/ICSI treatment. Hum. Reprod. 2012, 27, 2526–2531. [Google Scholar] [CrossRef]
- Gaskins, A.J.; Chiu, Y.-H.; Williams, P.L.; Keller, M.G.; Toth, T.L.; Hauser, R.; Chavarro, J. Maternal whole grain intake and outcomes of in vitro fertilization. Fertil. Steril. 2016, 105, 1503–1510.e4. [Google Scholar] [CrossRef]
- Machtinger, R.; Gaskins, A.; Mansur, A.; Adir, M.; Racowsky, C.; Baccarelli, A.; Hauser, R.; Chavarro, J. The association between preconception maternal caffeinated and non-caffeinated beverage intake on IVF outcomes. Hum. Reprod. 2017, 32, 284. [Google Scholar]
- Sohrabi, M.; Roushandeh, A.M.; Alizadeh, Z.; Vahidinia, A.; Vahabian, M.; Hosseini, M. Effect of a high fat diet on ovary morphology, in vitro development, in vitro fertilisation rate and oocyte quality in mice. Singap. Med. J. 2015, 56, 573–579. [Google Scholar] [CrossRef]
- Li, M.-C.; Nassan, F.; Chiu, Y.-H.; Mínguez-Alarcón, L.; Williams, P.L.; Souter, I.; Hauser, R.; Chavarro, J. Intake of Antioxidants in Relation to Infertility Treatment Outcomes with Assisted Reproductive Technologies. Epidemiology 2019, 30, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kutateladze, T.G. Diet and the epigenome. Nat. Commun. 2018, 9, 3375. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Tao, J.; Wu, H.; Guan, S.; Liu, L.; Zhang, L.; Deng, S.-L.; He, C.; Ji, P.; Liu, J.; et al. Aanat Knockdown and Melatonin Supplementation in Embryo Development: Involvement of Mitochondrial Function and DNA Methylation. Antioxid. Redox Signal. 2019, 30, 2050–2065. [Google Scholar] [CrossRef]
- Andreas, E.; Reid, M.; Zhang, W.; Moley, K.H. The effect of maternal high-fat/high-sugar diet on offspring oocytes and early embryo development. Mol. Hum. Reprod. 2019, 25, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.; Wong, F.Y.; Horne, R.S.; Yiallourou, S. Intrauterine growth restriction: Impact on cardiovascular development and function throughout infancy. Pediatr. Res. 2016, 79, 821–830. [Google Scholar] [CrossRef]
- Jaddoe, V.W.V.; De Jonge, L.L.; Hofman, A.; Franco, O.; Steegers, E.A.P.; Gaillard, R. First trimester fetal growth restriction and cardiovascular risk factors in school age children: Population based cohort study. BMJ 2014, 348, g14. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).