
Effects of BPA, BPS, and BPF on Oxidative Stress and Antioxidant Enzyme Expression in Bovine Oocytes and Spermatozoa



Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cumulus-Oocyte-Complex (COC) Collection and Maturation
2.3. Dose–Response Curve and In Vitro Embryo Production
2.4. Spermatozoa Preparation and Evaluation of Motility and Morphology
2.5. Measurement of Reactive Oxygen Species (ROS)
2.6. RNA Extraction and Reverse Transcription
2.7. Quantitative Polymerase Chain Reaction (qPCR)
2.8. Western Blotting
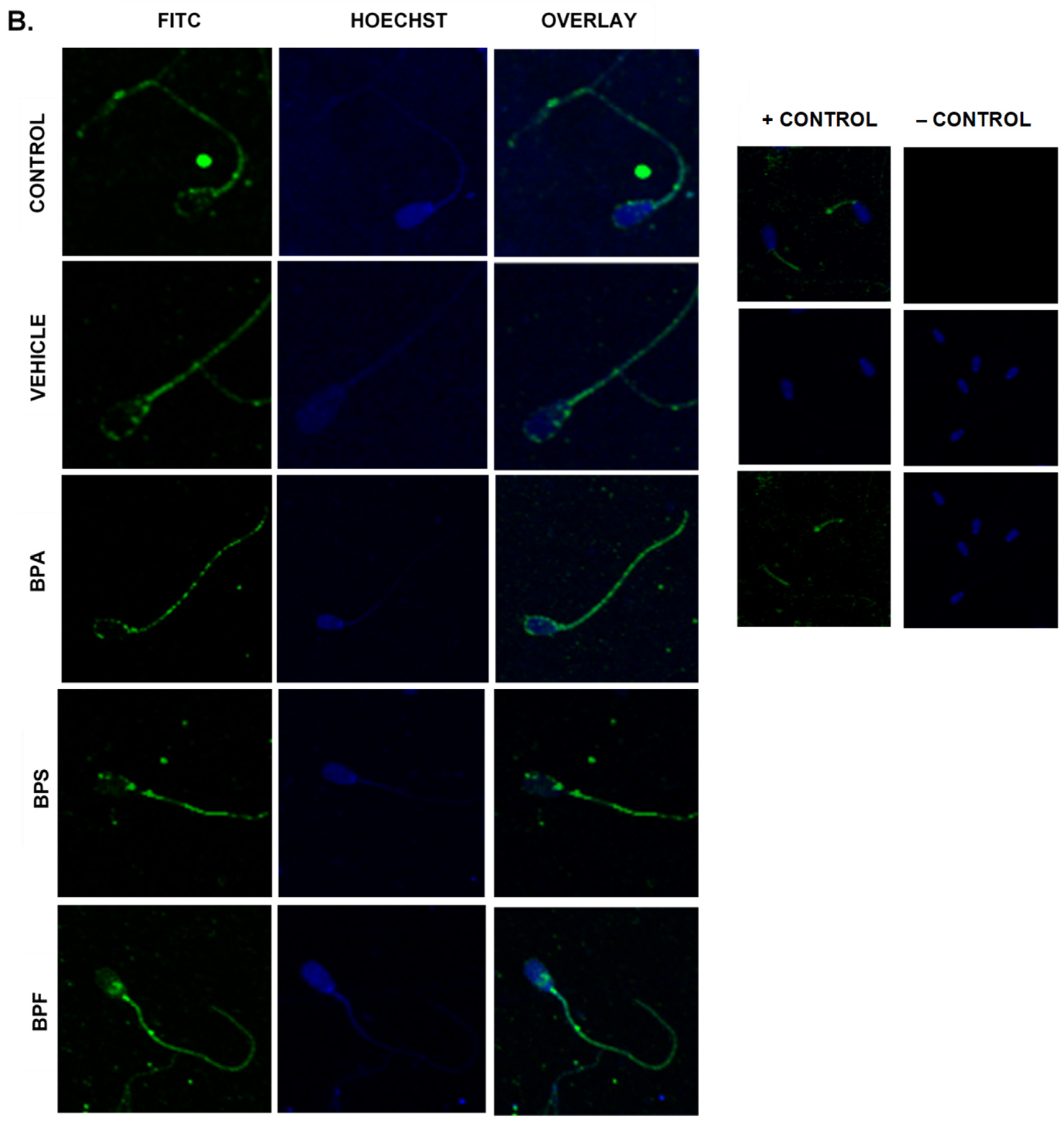
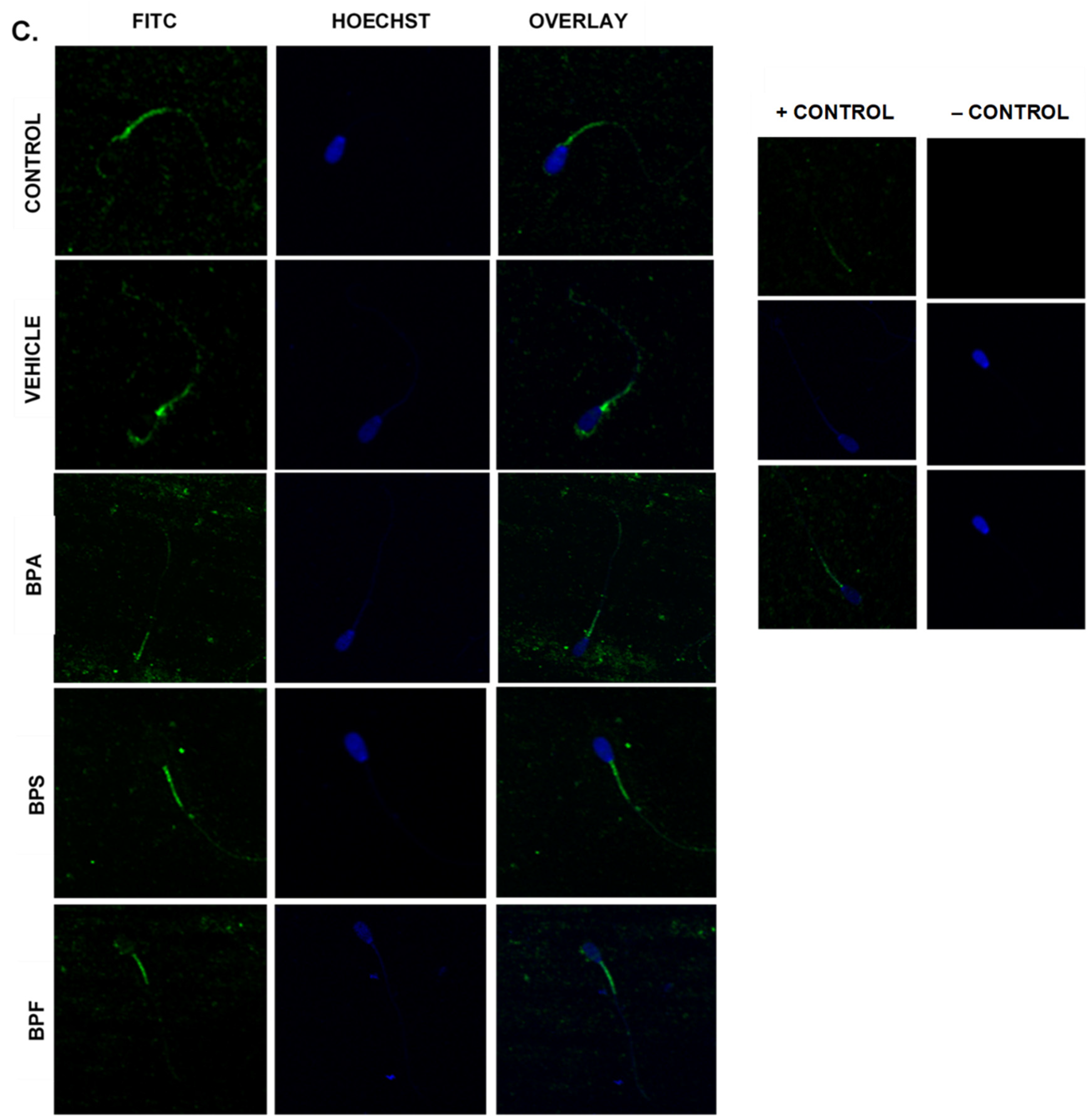
2.9. Immunofluorescent Staining of Sperm
2.10. Statistical Analysis
3. Results
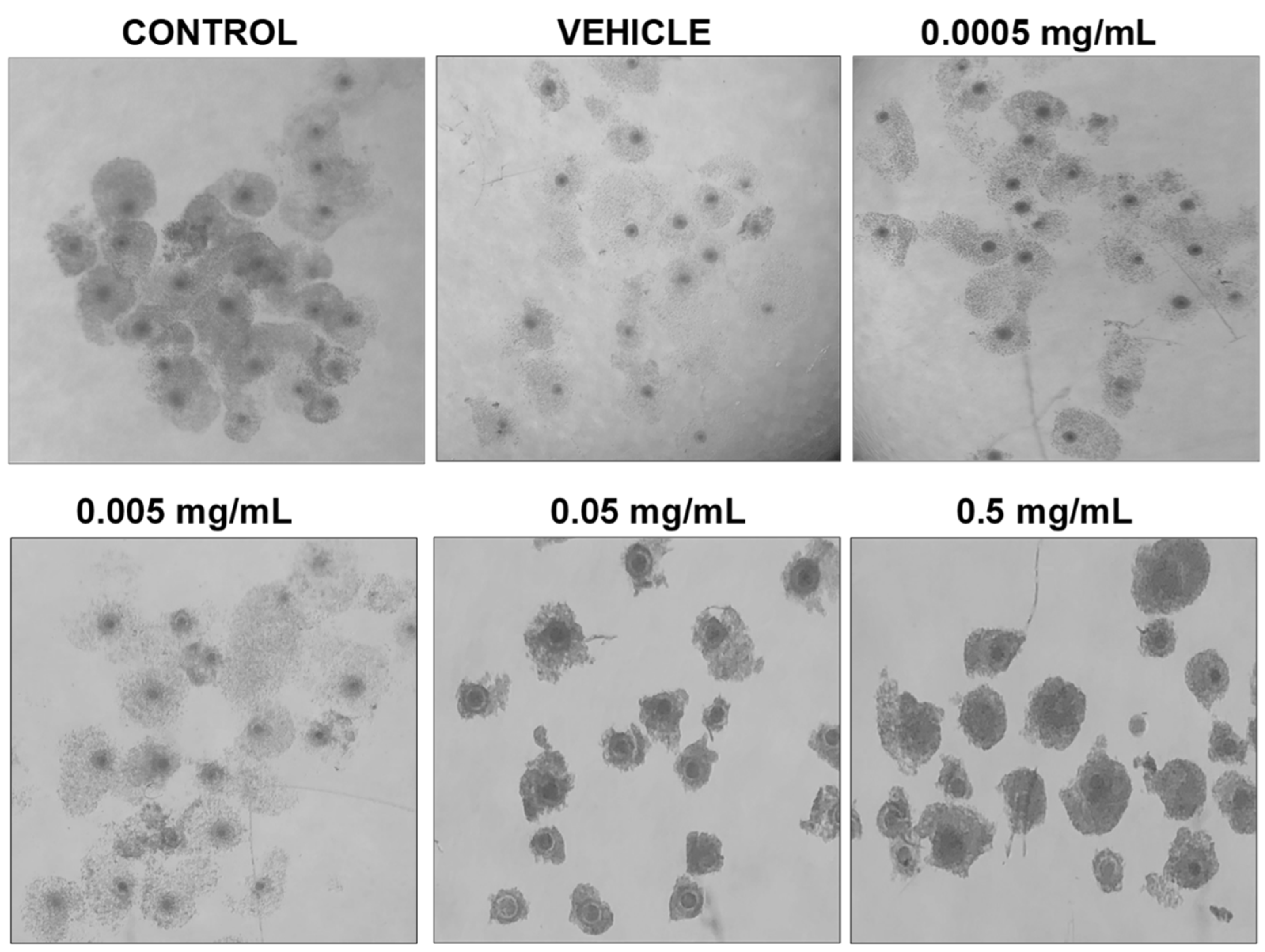
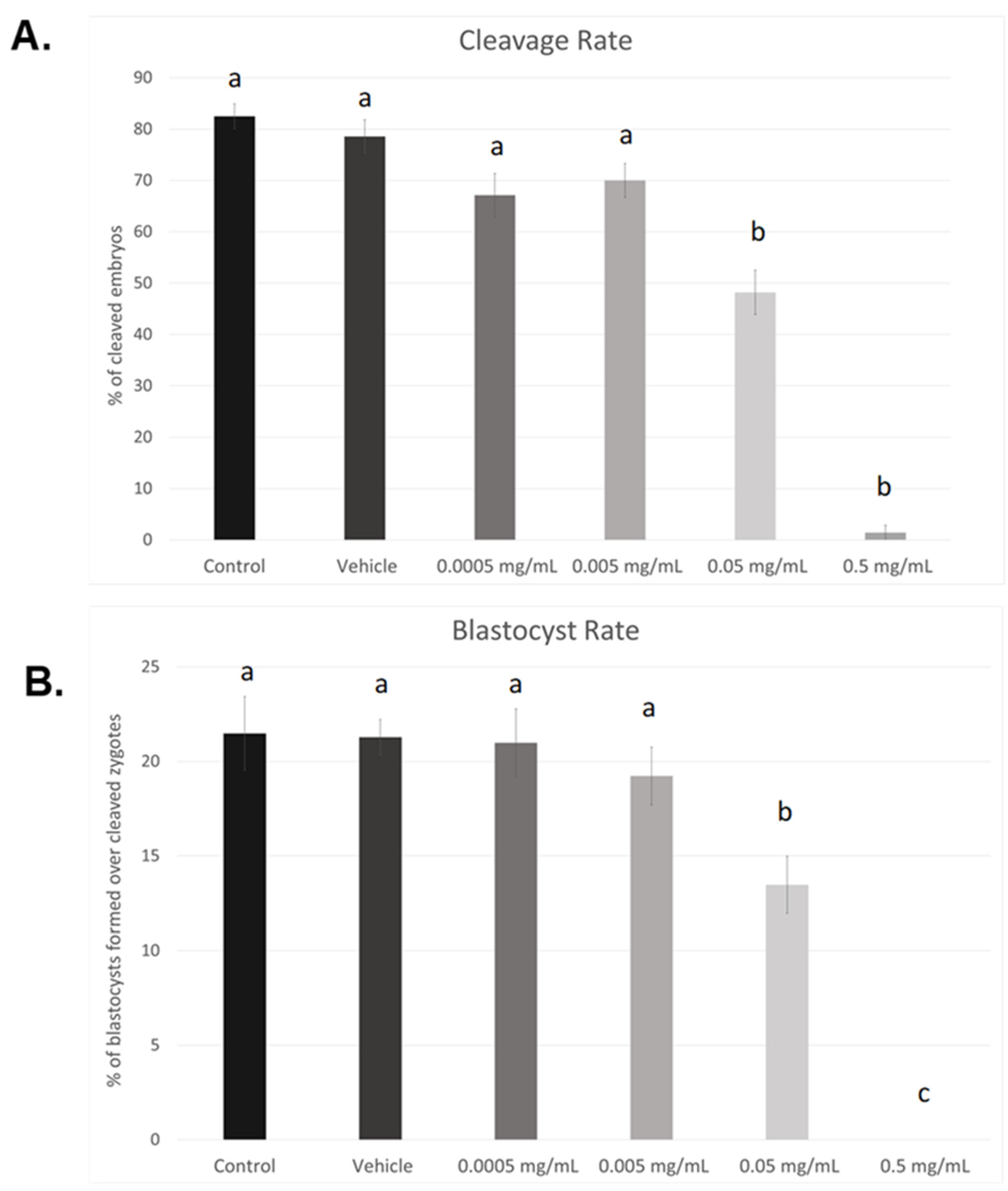
3.1. Dose–Response Curve for BPF
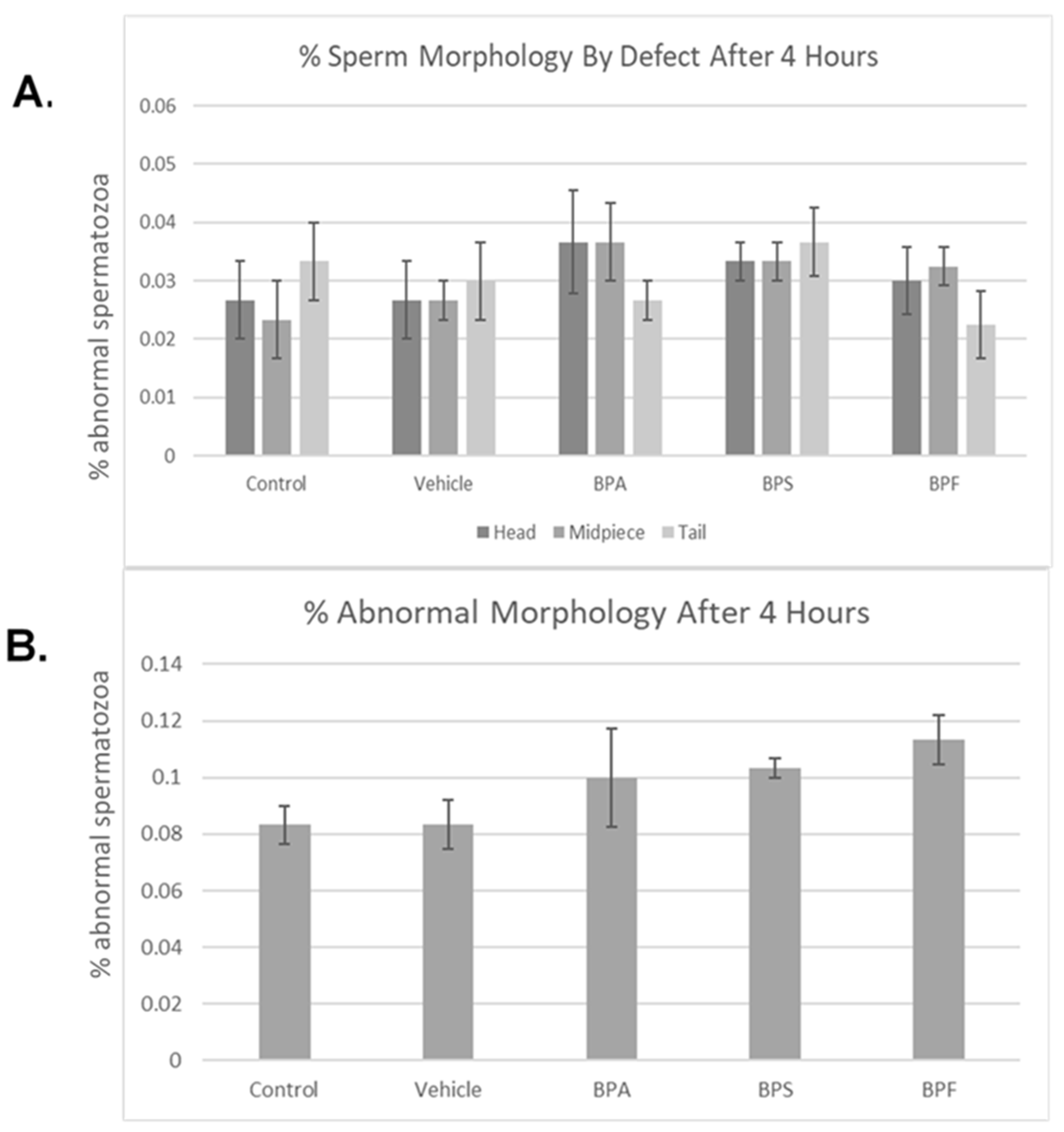
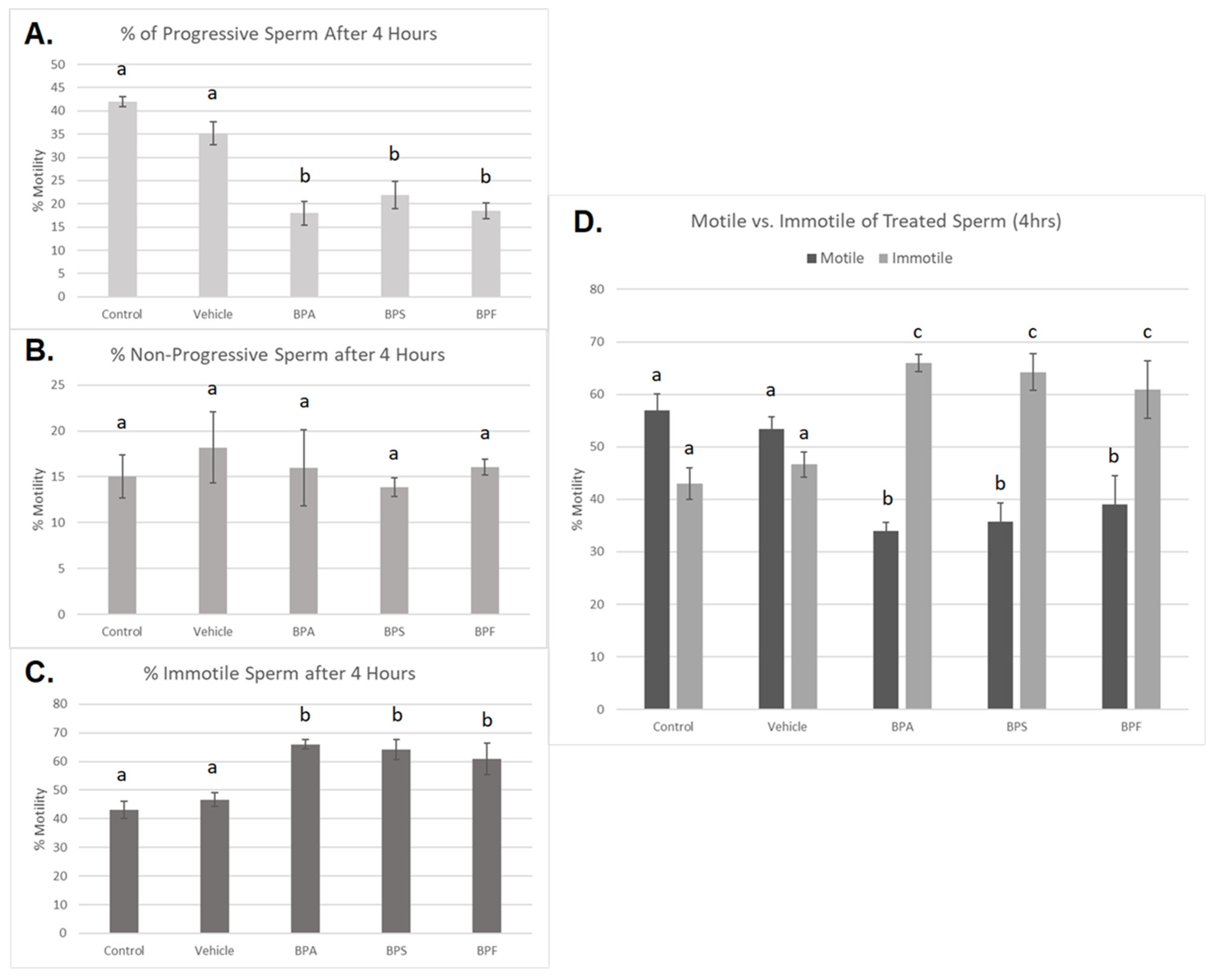
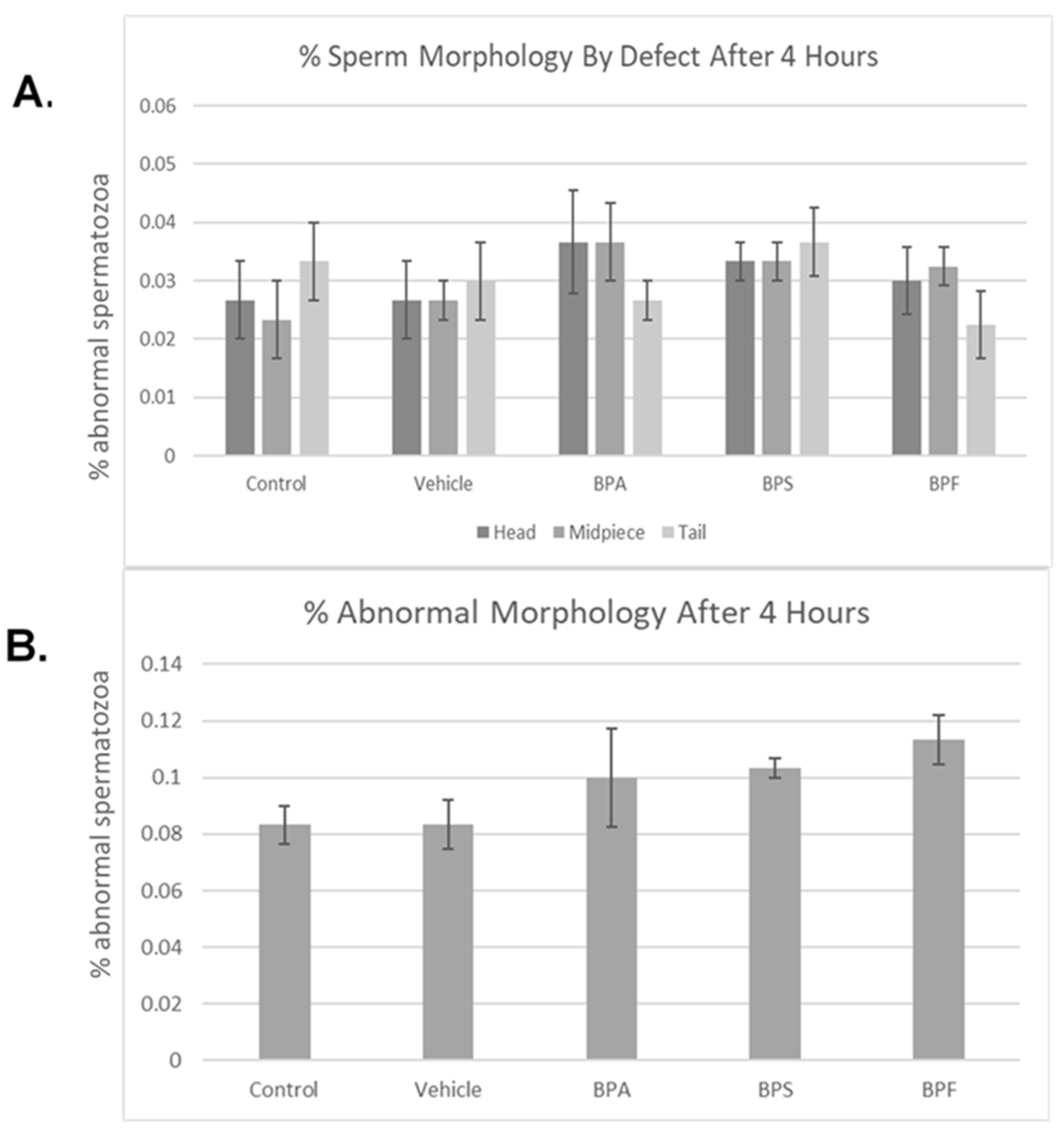
3.2. Sperm Motility and Morphology
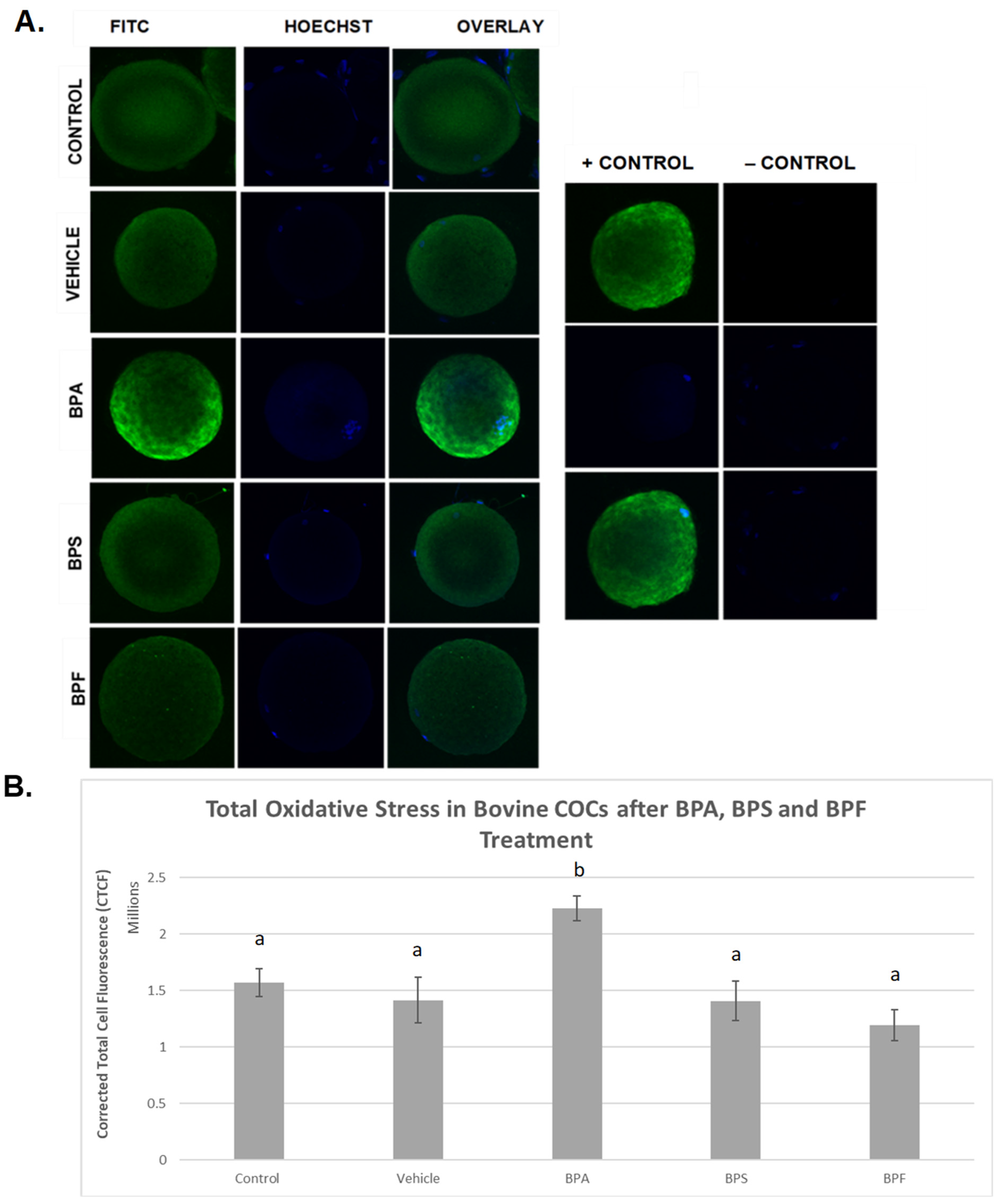
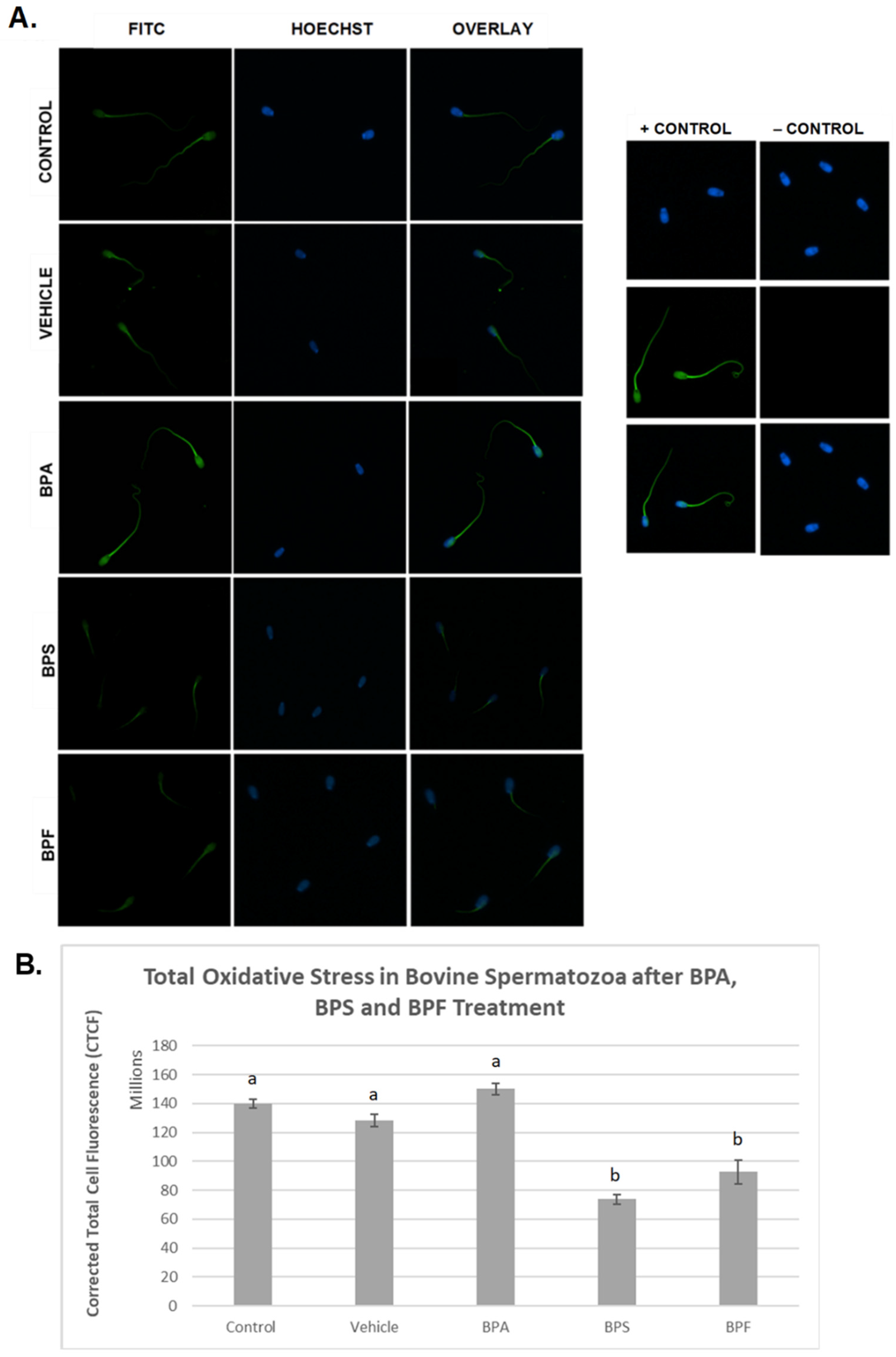
3.3. Oxidative Stress
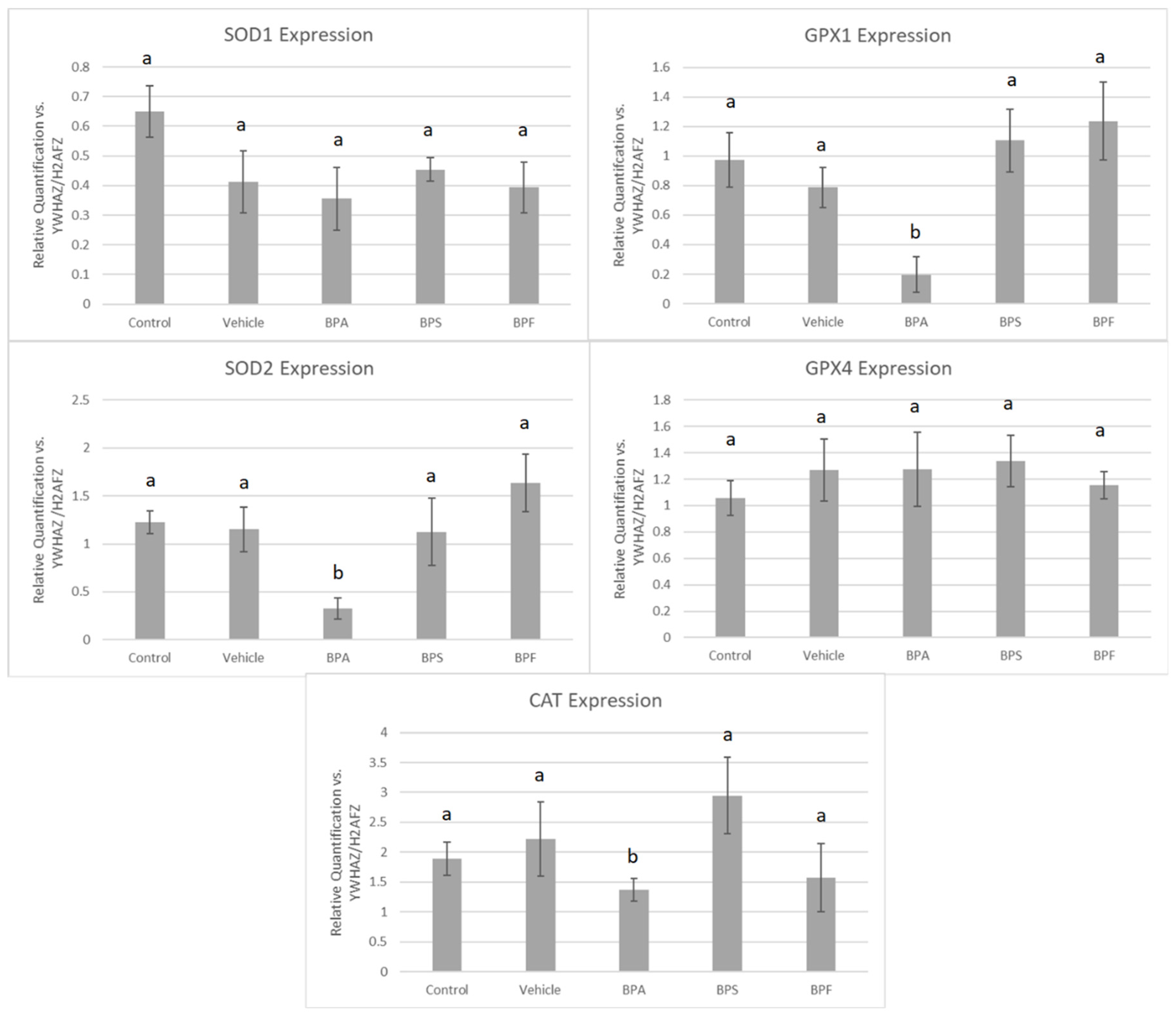
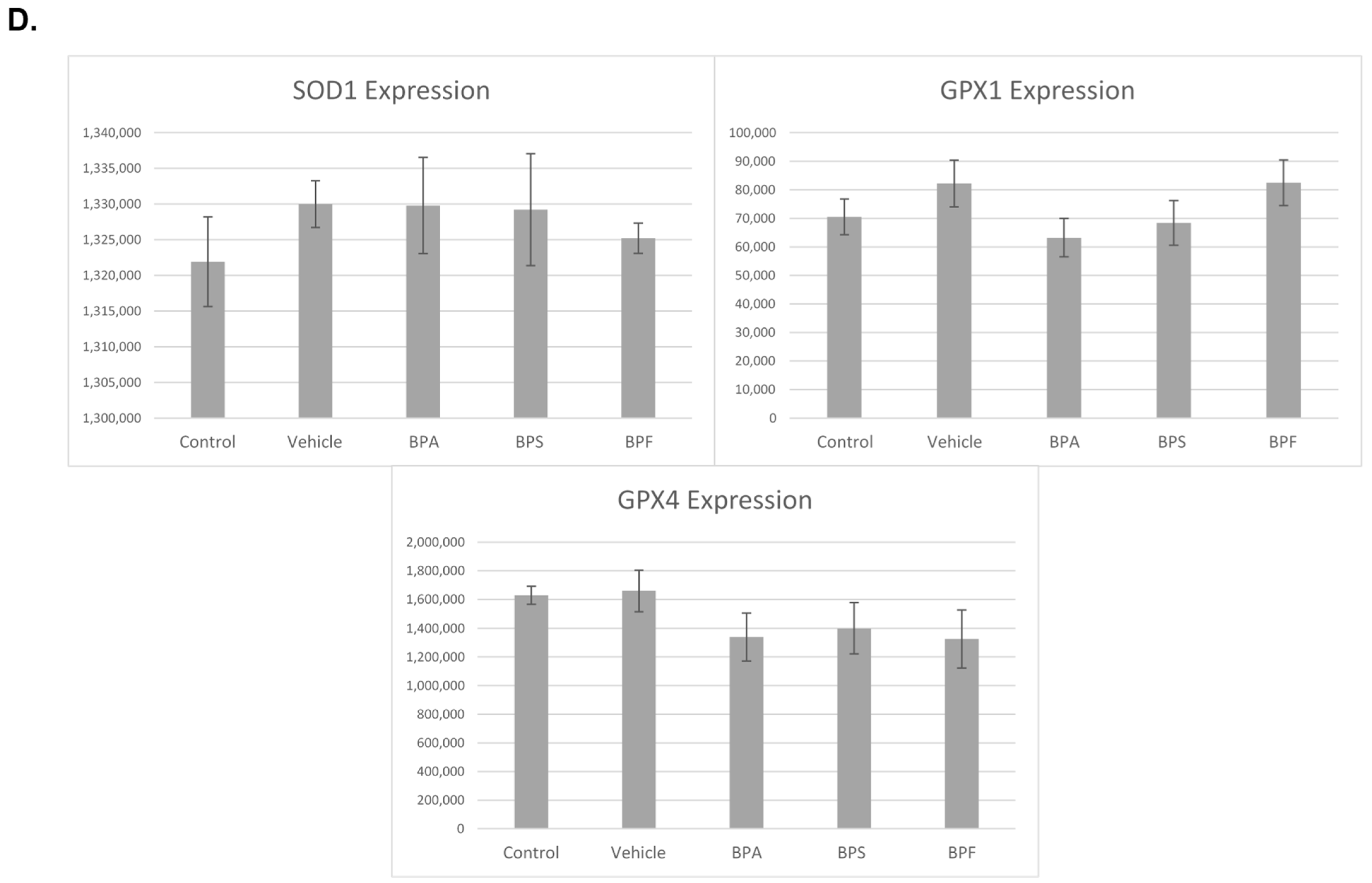
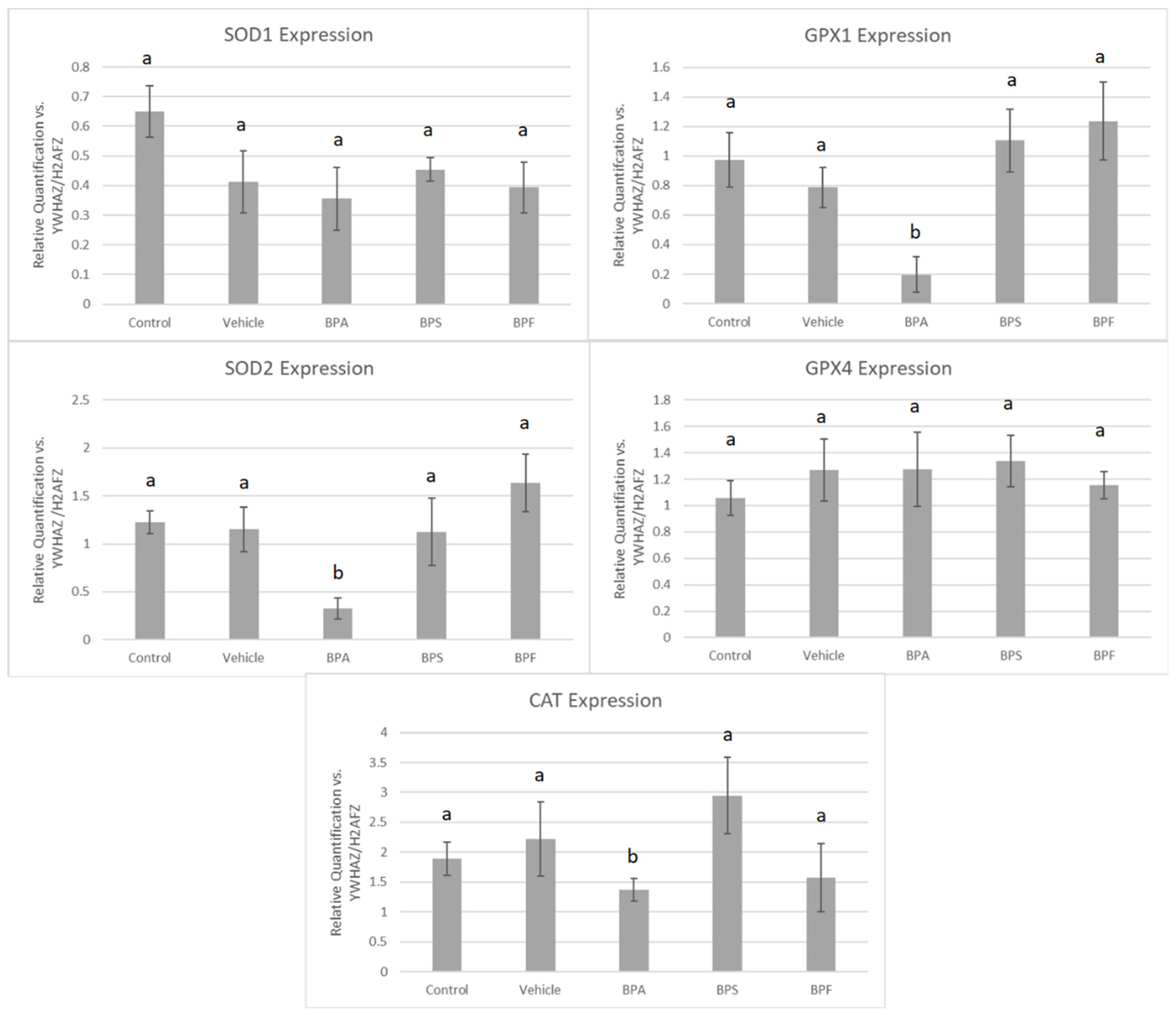
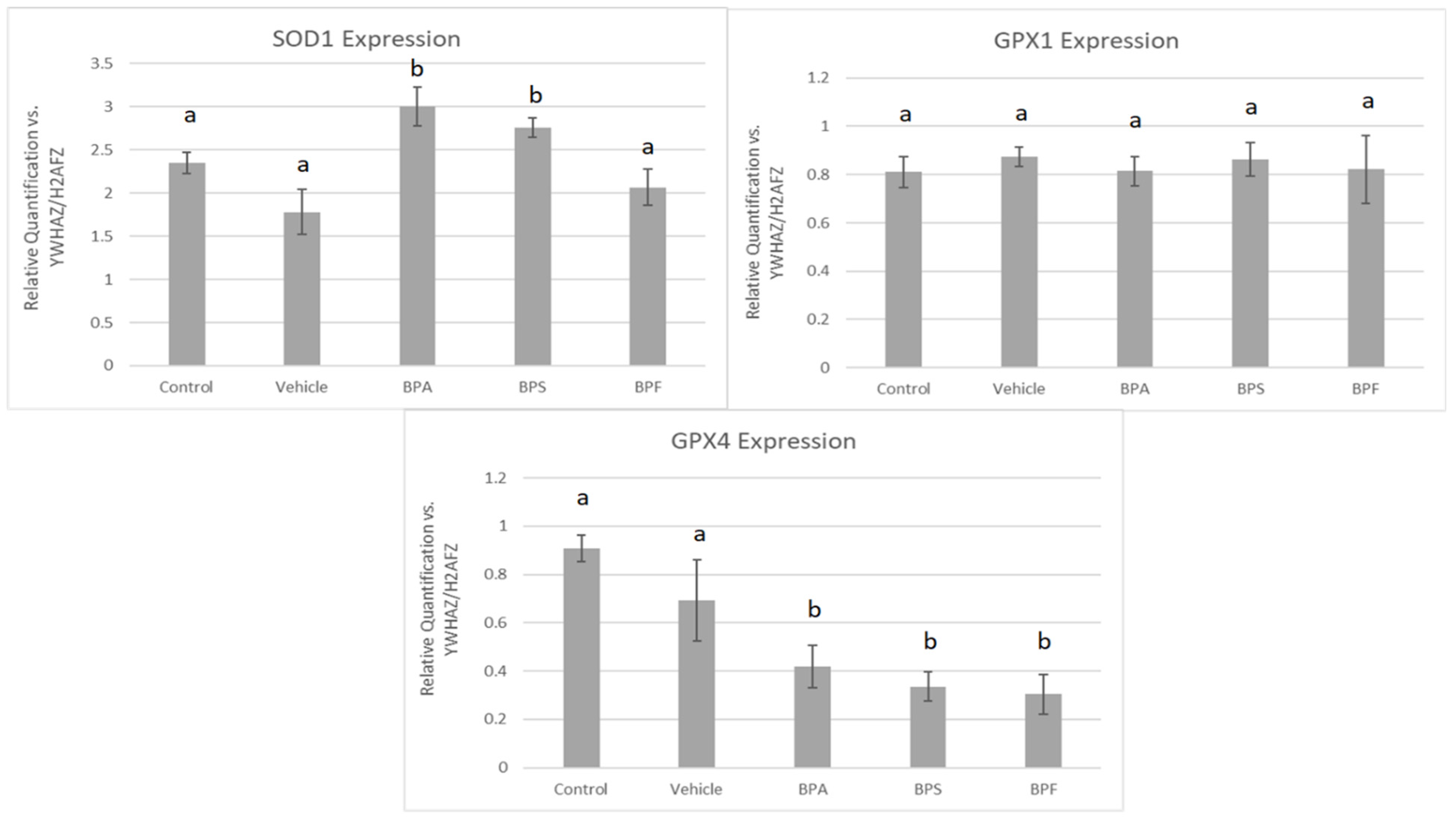
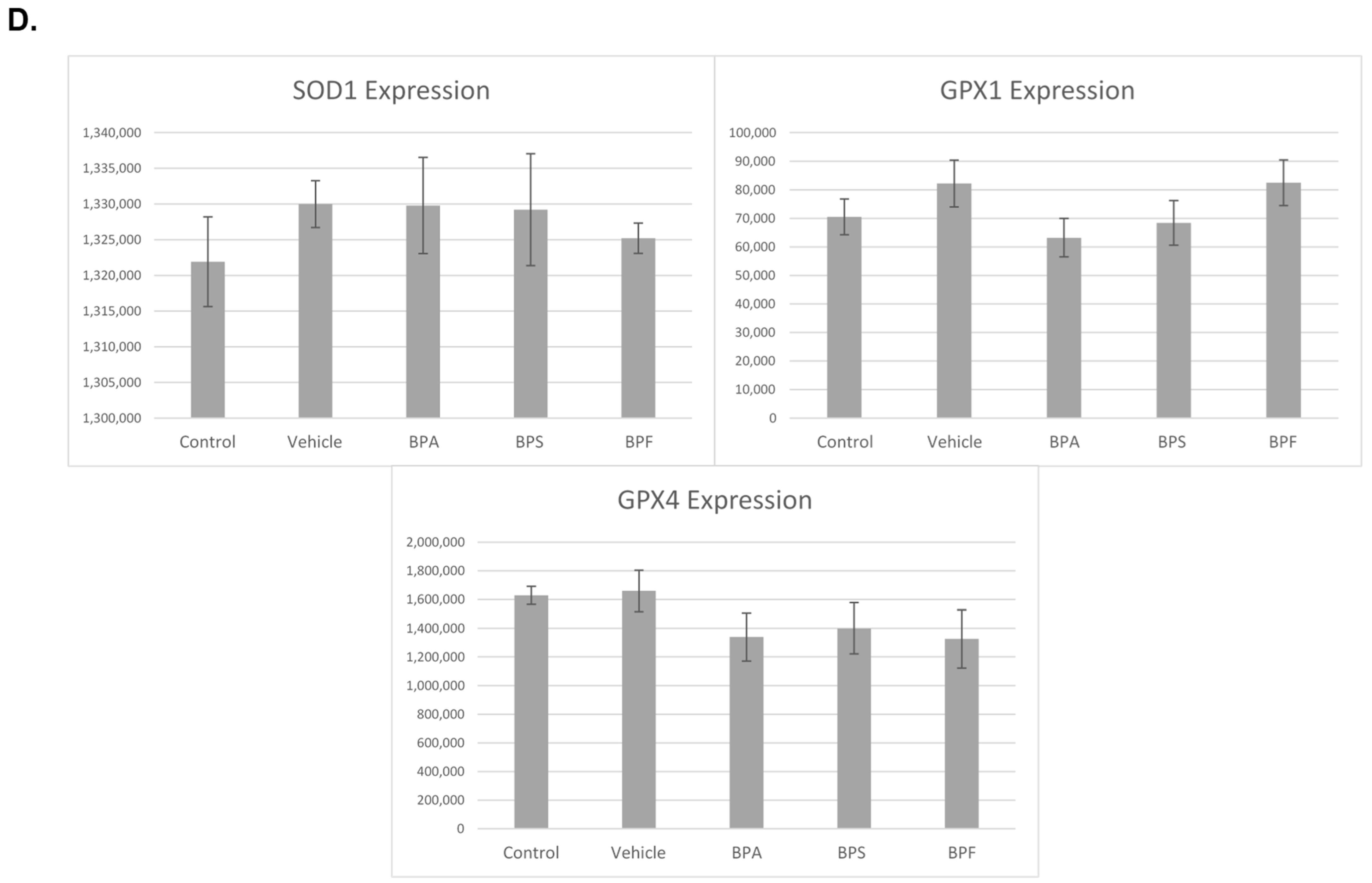
3.4. mRNA Expression in Treated Oocytes and Sperm
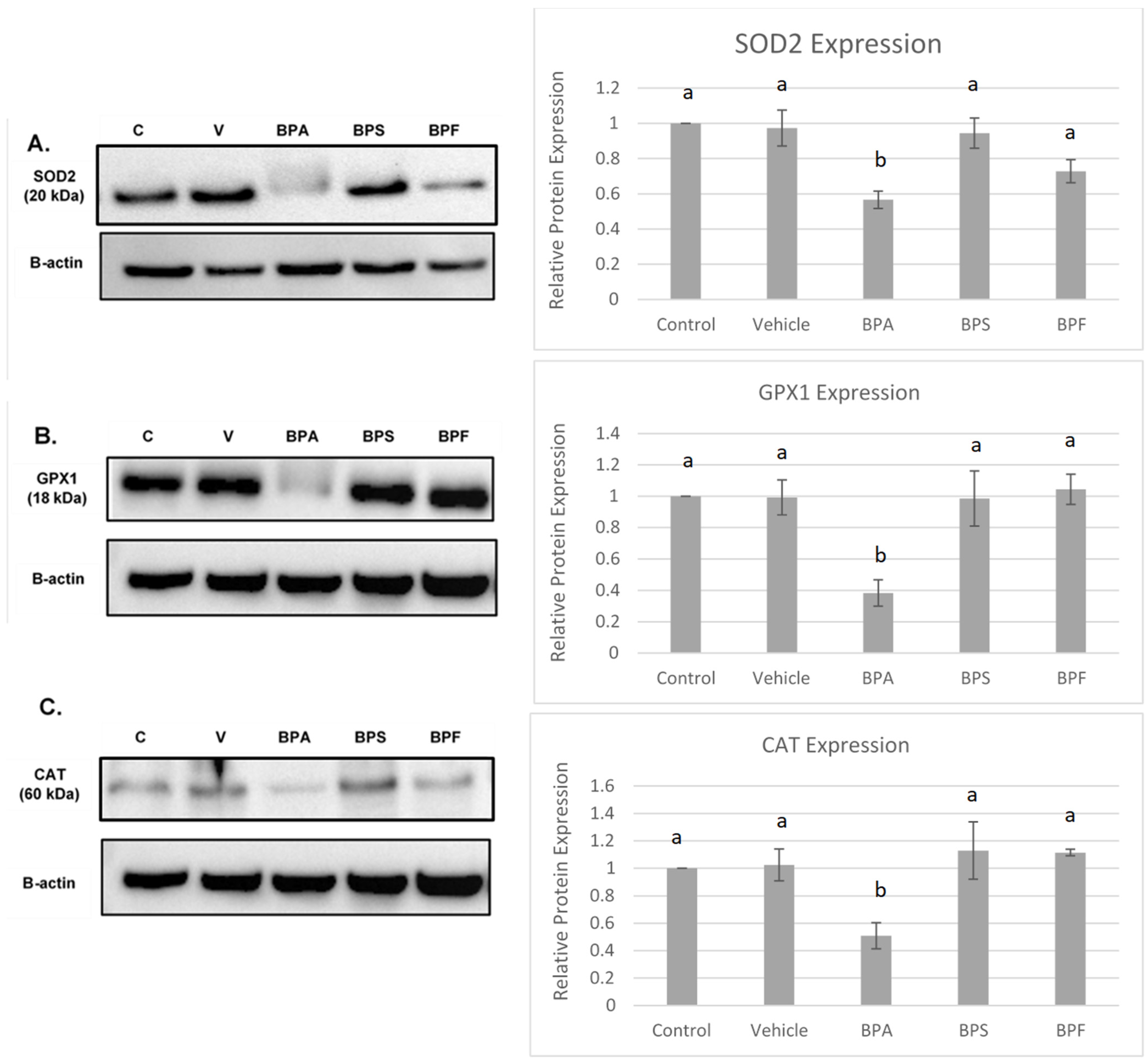
3.5. Protein Expression of COCs after BPA, BPS, and BPF Treatment
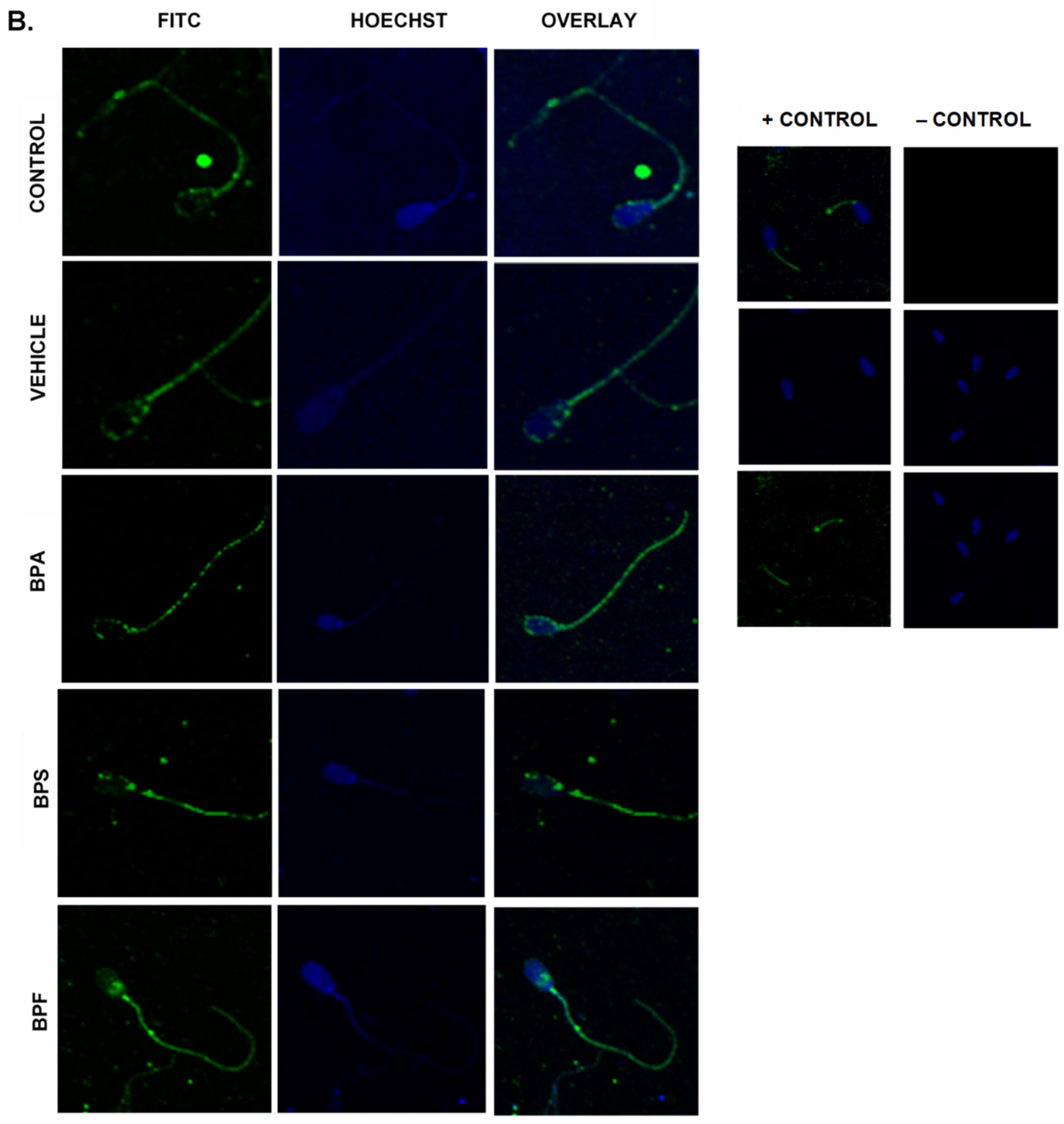
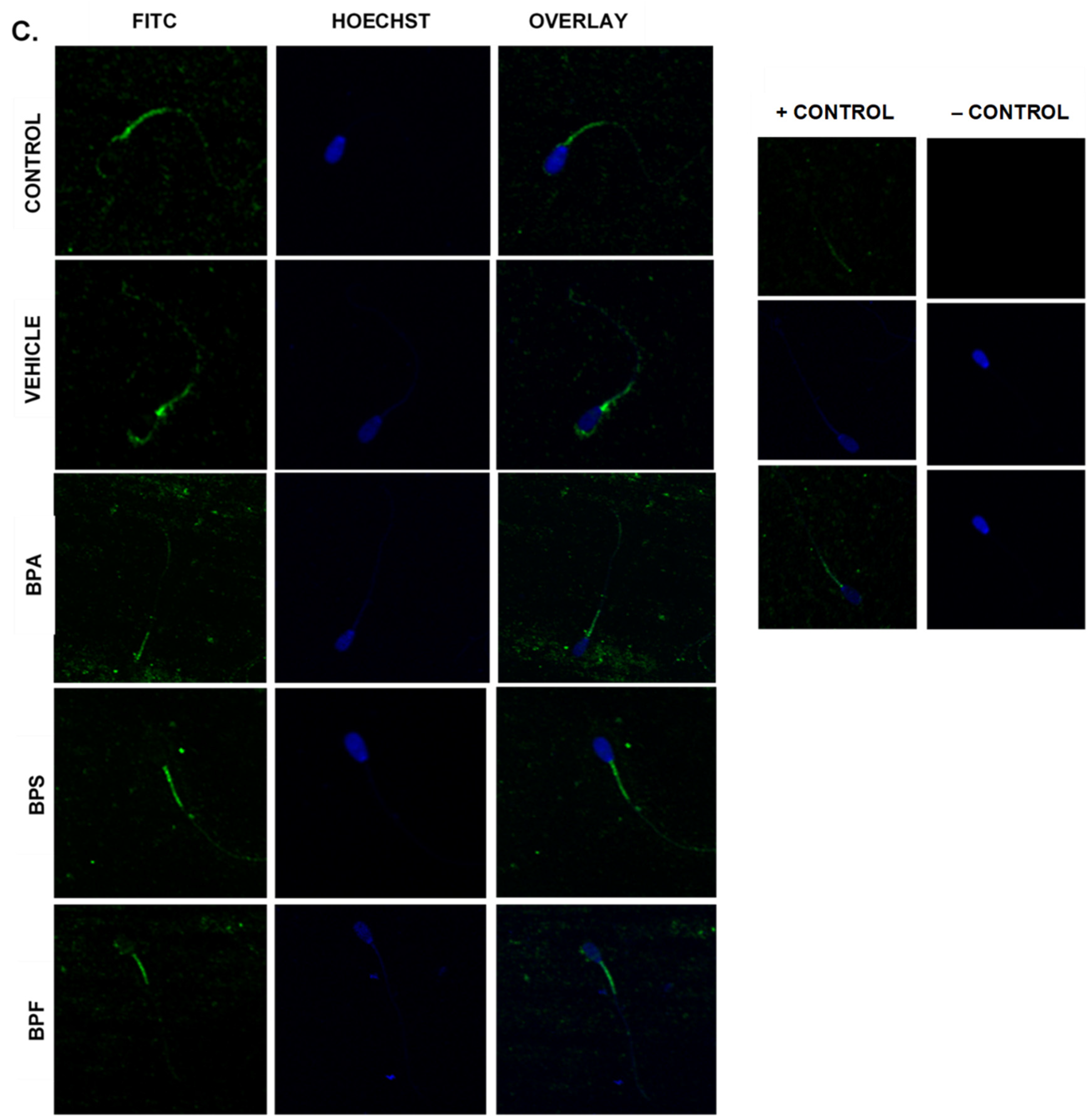
3.6. Protein Expression and Localization of Sperm
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Encarnação, T.; Pais, A.A.; Campos, M.G.; Burrows, H.D. Endocrine Disrupting Chemicals: Impact on Human Health, Wildlife and the Environment. Sci. Prog. 2019, 102, 3–42. [Google Scholar] [CrossRef] [PubMed]
- Corrales, J.; Kristofco, L.A.; Steele, W.B.; Yates, B.S.; Breed, C.S.; Williams, E.S.; Brooks, B.W. Global Assessment of Bisphenol A in the Environment: Review and Analysis of Its Occurrence and Bioaccumulation. Dose-Response 2015, 13, 1559325815598308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalal, N.; Surendranath, A.R.; Pathak, J.L.; Yu, S.; Chung, C.Y. Bisphenol A (BPA) the Mighty and the Mutagenic. Toxicol. Rep. 2018, 5, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Hauser, R.; Marcus, M.; Olea, N.; Welshons, W.V. Human Exposure to Bisphenol A (BPA). Reprod. Toxicol. 2007, 24, 139–177. [Google Scholar] [CrossRef]
- Meli, R.; Monnolo, A.; Annunziata, C.; Pirozzi, C.; Ferrante, M.C. Oxidative Stress and BPA Toxicity: An Antioxidant Approach for Male and Female Reproductive Dysfunction. Antioxidants 2020, 9, 405. [Google Scholar] [CrossRef] [PubMed]
- Téteau, O.; Jaubert, M.; Desmarchais, A.; Papillier, P.; Binet, A.; Maillard, V.; Elis, S. Bisphenol A and S Impaired Ovine Granulosa Cell Steroidogenesis. Reproduction 2020, 159, 571–583. [Google Scholar] [CrossRef]
- Thilagavathi, S.; Pugalendhi, P.; Rajakumar, T.; Vasudevan, K. Monotonic Dose Effect of Bisphenol-A, an Estrogenic Endocrine Disruptor, on Estrogen Synthesis in Female Sprague-Dawley Rats. Indian J. Clin. Biochem. 2018, 33, 387–396. [Google Scholar] [CrossRef]
- Santangeli, S.; Maradonna, F.; Olivotto, I.; Piccinetti, C.C.; Gioacchini, G.; Carnevali, O. Effects of BPA on Female Reproductive Function: The Involvement of Epigenetic Mechanism. Gen. Comp. Endocrinol. 2017, 245, 122–126. [Google Scholar] [CrossRef]
- Peretz, J.; Gupta, R.K.; Singh, J.; Hernández-Ochoa, I.; Flaws, J.A. Bisphenol A Impairs Follicle Growth, Inhibits Steroidogenesis, and Downregulates Rate-Limiting Enzymes in the Estradiol Biosynthesis Pathway. Toxicol. Sci. Off. J. Soc. Toxicol. 2011, 119, 209–217. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Z.; Liu, F. Chronic Exposure of BPA Impairs Male Germ Cell Proliferation and Induces Lower Sperm Quality in Male Mice. Chemosphere 2021, 262, 127880. [Google Scholar] [CrossRef]
- Lan, H.-C.; Wu, K.-Y.; Lin, I.-W.; Yang, Z.-J.; Chang, A.-A.; Hu, M.-C. Bisphenol A Disrupts Steroidogenesis and Induces a Sex Hormone Imbalance through C-Jun Phosphorylation in Leydig Cells. Chemosphere 2017, 185, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Luo, C.; Li, Q.; Chen, S.; Hu, Y. Mitochondrion-Mediated Apoptosis Is Involved in Reproductive Damage Caused by BPA in Male Rats. Environ. Toxicol. Pharmacol. 2014, 38, 1025–1033. [Google Scholar] [CrossRef]
- Moon, M.K. Concern about the Safety of Bisphenol A Substitutes. Diabetes Metab. J. 2019, 43, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Tucker, D.K.; Hayes, B.S.; Brar, S.S.; Kissling, G.E.; Fenton, S.E. Evaluation of Prenatal Exposure to Bisphenol Analogues on Development and Long-Term Health of the Mammary Gland in Female Mice. Environ. Health Perspect. 2018, 126, 087003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreman, J.; Lee, O.; Trznadel, M.; David, A.; Kudoh, T.; Tyler, C.R. Acute Toxicity, Teratogenic, and Estrogenic Effects of Bisphenol A and Its Alternative Replacements Bisphenol S, Bisphenol F, and Bisphenol AF in Zebrafish Embryo-Larvae. Environ. Sci. Technol. 2017, 51, 12796–12805. [Google Scholar] [CrossRef]
- Žalmanová, T.; Hošková, K.; Nevoral, J.; Prokešová, Š.; Zámostná, K.; Kott, T.; Petr, J. Bisphenol S Instead of Bisphenol A: A Story of Reproductive Disruption by Regretable Substitution—A Review. Czech J. Anim. Sci. 2016, 61, 433–449. [Google Scholar] [CrossRef] [Green Version]
- Rochester, J.R.; Bolden, A.L. Bisphenol S and F: A Systematic Review and Comparison of the Hormonal Activity of Bisphenol A Substitutes. Environ. Health Perspect. 2015, 123, 643–650. [Google Scholar] [CrossRef]
- Aitken, R.J. Impact of Oxidative Stress on Male and Female Germ Cells: Implications for Fertility. Reproduction 2020, 159, R189–R201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; He, J.; Xu, T.; Han, H.; Zhu, Z.; Meng, L.; Pang, Q.; Fan, R. Bisphenol A(BPA), BPS and BPB-Induced Oxidative Stress and Apoptosis Mediated by Mitochondria in Human Neuroblastoma Cell Lines. Ecotoxicol. Environ. Saf. 2021, 207, 111299. [Google Scholar] [CrossRef]
- Lu, J.; Wang, Z.; Cao, J.; Chen, Y.; Dong, Y. A Novel and Compact Review on the Role of Oxidative Stress in Female Reproduction. Reprod. Biol. Endocrinol. 2018, 16, 80. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.L.M.; Day, M.L.; Morris, M.B. Redox Regulation and Oxidative Stress in Mammalian Oocytes and Embryos Developed In Vivo and In Vitro. Int. J. Environ. Res. Public Health 2021, 18, 11374. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of Oxidative Stress on Male Reproduction. World J. Mens Health 2014, 32, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bindhumol, V.; Chitra, K.C.; Mathur, P.P. Bisphenol A Induces Reactive Oxygen Species Generation in the Liver of Male Rats. Toxicology 2003, 188, 117–124. [Google Scholar] [CrossRef]
- Aboul Ezz, H.S.; Khadrawy, Y.A.; Mourad, I.M. The Effect of Bisphenol A on Some Oxidative Stress Parameters and Acetylcholinesterase Activity in the Heart of Male Albino Rats. Cytotechnology 2015, 67, 145–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Han, J.; Duan, X.; Xiong, B.; Cui, X.-S.; Kim, N.-H.; Liu, H.-L.; Sun, S.-C. The Toxic Effects and Possible Mechanisms of Bisphenol A on Oocyte Maturation of Porcine in Vitro. Oncotarget 2016, 7, 32554–32565. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-J.; Park, S.-Y.; Kim, J.-W.; Yang, S.-G.; Kim, M.-J.; Jegal, H.-G.; Kim, I.-S.; Choo, Y.-K.; Koo, D.-B. Melatonin Improves Oocyte Maturation and Mitochondrial Functions by Reducing Bisphenol A-Derived Superoxide in Porcine Oocytes In Vitro. Int. J. Mol. Sci. 2018, 19, 3422. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Zhao, M.-H.; Shin, K.-T.; Niu, Y.-J.; Ahn, Y.-D.; Kim, N.-H.; Cui, X.-S. The Possible Molecular Mechanisms of Bisphenol A Action on Porcine Early Embryonic Development. Sci. Rep. 2017, 7, 8632. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.; Mahboubi, K.; MacLusky, N.; King, W.A.; Favetta, L.A. BPA Exposure during in Vitro Oocyte Maturation Results in Dose-Dependent Alterations to Embryo Development Rates, Apoptosis Rate, Sex Ratio and Gene Expression. Reprod. Toxicol. 2016, 59, 128–138. [Google Scholar] [CrossRef]
- Alahmar, A.T. Role of Oxidative Stress in Male Infertility: An Updated Review. J. Hum. Reprod. Sci. 2019, 12, 4–18. [Google Scholar] [CrossRef]
- Fang, Y.; Zhong, R. Effects of Oxidative Stress on Spermatozoa and Male Infertility; IntechOpen: London, UK, 2019; ISBN 978-1-78985-144-1. [Google Scholar]
- Kaur, S.; Saluja, M.; Bansal, M.P. Bisphenol A Induced Oxidative Stress and Apoptosis in Mice Testes: Modulation by Selenium. Andrologia 2018, 50, e12834. [Google Scholar] [CrossRef]
- Langbeen, A.; De Porte, H.F.M.; Bartholomeus, E.; Leroy, J.L.M.R.; Bols, P.E.J. Bovine in Vitro Reproduction Models Can Contribute to the Development of (Female) Fertility Preservation Strategies. Theriogenology 2015, 84, 477–489. [Google Scholar] [CrossRef]
- Santos, R.R.; Schoevers, E.J.; Roelen, B.A. Usefulness of Bovine and Porcine IVM/IVF Models for Reproductive Toxicology. Reprod. Biol. Endocrinol. 2014, 12, 117. [Google Scholar] [CrossRef] [Green Version]
- Santonicola, S.; Ferrante, M.C.; Murru, N.; Gallo, P.; Mercogliano, R. Hot Topic: Bisphenol A in Cow Milk and Dietary Exposure at the Farm Level. J. Dairy Sci. 2019, 102, 1007–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgescu, B.; Georgescu, C. Bisphenol A Levels in Commercial Milk, Infant Formula and Dairy Products. Anim. Biol. Anim. Husb. 2013, 5, 171–174. [Google Scholar]
- Rhind, S.M.; Kyle, C.E.; Mackie, C.; Yates, K.; Duff, E.I. Geographic Variation in Tissue Accumulation of Endocrine Disrupting Compounds (EDCs) in Grazing Sheep. Environ. Pollut. 1987 2011, 159, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Sabry, R.; Saleh, A.C.; Stalker, L.; LaMarre, J.; Favetta, L.A. Effects of Bisphenol A and Bisphenol S on MicroRNA Expression during Bovine (Bos Taurus) Oocyte Maturation and Early Embryo Development. Reprod. Toxicol. 2021, 99, 96–108. [Google Scholar] [CrossRef]
- Saleh, A.C.; Sabry, R.; Mastromonaco, G.F.; Favetta, L.A. BPA and BPS Affect the Expression of Anti-Mullerian Hormone (AMH) and Its Receptor during Bovine Oocyte Maturation and Early Embryo Development. Reprod. Biol. Endocrinol. 2021, 19, 119. [Google Scholar] [CrossRef] [PubMed]
- Davis, O.; Hickey, K.; Favetta, L. 113 The Effects of Bisphenols on Cryopreserved Bovine Spermatozoa in Vitro. Reprod. Fertil. Dev. 2021, 34, 293–294. [Google Scholar] [CrossRef]
- Li, N.; Kang, H.; Peng, Z.; Wang, H.; Weng, S.; Zeng, X. Physiologically Detectable Bisphenol A Impairs Human Sperm Functions by Reducing Protein-Tyrosine Phosphorylation. Ecotoxicol. Environ. Saf. 2021, 221, 112418. [Google Scholar] [CrossRef]
- WHO Laboratory Manual for the Examination and Processing of Human Semen. Available online: https://www.who.int/publications-detail-redirect/9789240030787 (accessed on 1 December 2021).
- Tiwari, M.; Chaube, S.K. Moderate Increase of Reactive Oxygen Species Triggers Meiotic Resumption in Rat Follicular Oocytes. J. Obstet. Gynaecol. Res. 2016, 42, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Sangalli, J.R.; Sampaio, R.V.; del Collado, M.; da Silveira, J.C.; De Bem, T.H.C.; Perecin, F.; Smith, L.C.; Meirelles, F.V. Metabolic Gene Expression and Epigenetic Effects of the Ketone Body β-Hydroxybutyrate on H3K9ac in Bovine Cells, Oocytes and Embryos. Sci. Rep. 2018, 8, 13766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Li, Y.; Zhao, Y.; Gao, Q.; Jin, Q.; Yan, C.; Xu, Y. L-Carnitine Supplementation during in Vitro Culture Regulates Oxidative Stress in Embryos from Bovine Aged Oocytes. Theriogenology 2020, 143, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Misirlioglu, M.; Page, G.P.; Sagirkaya, H.; Kaya, A.; Parrish, J.J.; First, N.L.; Memili, E. Dynamics of Global Transcriptome in Bovine Matured Oocytes and Preimplantation Embryos. Proc. Natl. Acad. Sci. USA 2006, 103, 18905–18910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Loos, F.; van Vliet, C.; van Maurik, P.; Kruip, T.A. Morphology of Immature Bovine Oocytes. Gamete Res. 1989, 24, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Franciosi, F. Acquisition of Oocyte Competence to Develop as an Embryo: Integrated Nuclear and Cytoplasmic Events. Hum. Reprod. Update 2018, 24, 245–266. [Google Scholar] [CrossRef]
- Albertini, D.F.; Sanfins, A.; Combelles, C.M.H. Origins and Manifestations of Oocyte Maturation Competencies. Reprod. Biomed. Online 2003, 6, 410–415. [Google Scholar] [CrossRef]
- Molina-Molina, J.-M.; Amaya, E.; Grimaldi, M.; Sáenz, J.-M.; Real, M.; Fernández, M.F.; Balaguer, P.; Olea, N. In Vitro Study on the Agonistic and Antagonistic Activities of Bisphenol-S and Other Bisphenol-A Congeners and Derivatives via Nuclear Receptors. Toxicol. Appl. Pharmacol. 2013, 272, 127–136. [Google Scholar] [CrossRef]
- Maćczak, A.; Duchnowicz, P.; Sicińska, P.; Koter-Michalak, M.; Bukowska, B.; Michałowicz, J. The in Vitro Comparative Study of the Effect of BPA, BPS, BPF and BPAF on Human Erythrocyte Membrane; Perturbations in Membrane Fluidity, Alterations in Conformational State and Damage to Proteins, Changes in ATP Level and Na+/K+ ATPase and AChE Activities. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2017, 110, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Turathum, B.; Gao, E.-M.; Chian, R.-C. The Function of Cumulus Cells in Oocyte Growth and Maturation and in Subsequent Ovulation and Fertilization. Cells 2021, 10, 2292. [Google Scholar] [CrossRef]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and Consequences of Oxidative Stress in Spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Agnihotri, S.K.; Agrawal, A.K.; Hakim, B.A.; Vishwakarma, A.L.; Narender, T.; Sachan, R.; Sachdev, M. Mitochondrial Membrane Potential (MMP) Regulates Sperm Motility. Vitr. Cell. Dev. Biol. Anim. 2016, 52, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Chabory, E.; Damon, C.; Lenoir, A.; Kauselmann, G.; Kern, H.; Zevnik, B.; Garrel, C.; Saez, F.; Cadet, R.; Henry-Berger, J.; et al. Epididymis Seleno-Independent Glutathione Peroxidase 5 Maintains Sperm DNA Integrity in Mice. J. Clin. Investig. 2009, 119, 2074–2085. [Google Scholar] [CrossRef]
- Itach, S.B.-S.; Finklestein, M.; Etkovitz, N.; Breitbart, H. Hyper-Activated Motility in Sperm Capacitation Is Mediated by Phospholipase D-Dependent Actin Polymerization. Dev. Biol. 2012, 362, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naresh, S.; Atreja, S.K. The Protein Tyrosine Phosphorylation during in Vitro Capacitation and Cryopreservation of Mammalian Spermatozoa. Cryobiology 2015, 70, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhao, Z. Influence of N-Acetyl-L-Cysteine against Bisphenol a on the Maturation of Mouse Oocytes and Embryo Development: In Vitro Study. BMC Pharmacol. Toxicol. 2019, 20, 43. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. Reactive Oxygen Species as Mediators of Sperm Capacitation and Pathological Damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Castellini, C.; Totaro, M.; Parisi, A.; D’Andrea, S.; Lucente, L.; Cordeschi, G.; Francavilla, S.; Francavilla, F.; Barbonetti, A. Bisphenol A and Male Fertility: Myths and Realities. Front. Endocrinol. 2020, 11, 353. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, X.-F.; Ji, Y.-L.; Wang, H.; Liu, P.; Zhang, C.; Zhang, Y.; Xu, D.-X. Mitochondrial Signaling Pathway Is Also Involved in Bisphenol A Induced Germ Cell Apoptosis in Testes. Toxicol. Lett. 2010, 199, 129–135. [Google Scholar] [CrossRef]
- Koppers, A.J.; Mitchell, L.A.; Wang, P.; Lin, M.; Aitken, R.J. Phosphoinositide 3-Kinase Signalling Pathway Involvement in a Truncated Apoptotic Cascade Associated with Motility Loss and Oxidative DNA Damage in Human Spermatozoa. Biochem. J. 2011, 436, 687–698. [Google Scholar] [CrossRef]
- Combelles, C.M.; Gupta, S.; Agarwal, A. Could Oxidative Stress Influence the In-Vitro Maturation of Oocytes? Reprod. Biomed. Online 2009, 18, 864–880. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Gur, Y.; Breitbart, H. Mammalian Sperm Translate Nuclear-Encoded Proteins by Mitochondrial-Type Ribosomes. Genes Dev. 2006, 20, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Ribas-Maynou, J.; Yeste, M. Oxidative Stress in Male Infertility: Causes, Effects in Assisted Reproductive Techniques, and Protective Support of Antioxidants. Biology 2020, 9, 77. [Google Scholar] [CrossRef] [Green Version]
- Anjum, S.; Rahman, S.; Kaur, M.; Ahmad, F.; Rashid, H.; Ansari, R.A.; Raisuddin, S. Melatonin Ameliorates Bisphenol A-Induced Biochemical Toxicity in Testicular Mitochondria of Mouse. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2011, 49, 2849–2854. [Google Scholar] [CrossRef] [PubMed]
- Papas, M.; Catalan, J.; Barranco, I.; Arroyo, L.; Bassols, A.; Yeste, M.; Miró, J. Total and Specific Activities of Superoxide Dismutase (SOD) in Seminal Plasma Are Related with the Cryotolerance of Jackass Spermatozoa. Cryobiology 2020, 92, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Noblanc, A.; Kocer, A.; Chabory, E.; Vernet, P.; Saez, F.; Cadet, R.; Conrad, M.; Drevet, J.R. Glutathione Peroxidases at Work on Epididymal Spermatozoa: An Example of the Dual Effect of Reactive Oxygen Species on Mammalian Male Fertilizing Ability. J. Androl. 2011, 32, 641–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, M.; Förster, H.; Boersma, A.; Seiler, A.; Wehnes, H.; Sinowatz, F.; Neumüller, C.; Deutsch, M.J.; Walch, A.; Hrabé de Angelis, M.; et al. Mitochondrial Glutathione Peroxidase 4 Disruption Causes Male Infertility. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2009, 23, 3233–3242. [Google Scholar] [CrossRef] [Green Version]
- El-Taieb, M.A.; Ali, M.A.; Nada, E.A. Oxidative Stress and Acrosomal Morphology: A Cause of Infertility in Patients with Normal Semen Parameters. Middle East Fertil. Soc. J. 2015, 20, 79–85. [Google Scholar] [CrossRef]
- Nguyen, M.; Sabry, R.; Younes, S.; Favetta, L.A. BPA and Its Analogs Increase Oxidative Stress Levels in in Vitro Cultured Granulosa Cells by Altering Anti-Oxidant Enzymes Expression. Mol. Cell. Endocrinol. 2021; submitted manuscript. [Google Scholar]
- Sasaki, H.; Hamatani, T.; Kamijo, S.; Iwai, M.; Kobanawa, M.; Ogawa, S.; Miyado, K.; Tanaka, M. Impact of Oxidative Stress on Age-Associated Decline in Oocyte Developmental Competence. Front. Endocrinol. 2019, 10, 811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Mih, N.; Anand, A.; Park, J.H.; Tan, J.; Yurkovich, J.T.; Monk, J.M.; Lloyd, C.J.; Sandberg, T.E.; Seo, S.W.; et al. Cellular Responses to Reactive Oxygen Species Are Predicted from Molecular Mechanisms. Proc. Natl. Acad. Sci. USA 2019, 116, 14368–14373. [Google Scholar] [CrossRef] [Green Version]
- Ugarte, N.; Petropoulos, I.; Friguet, B. Oxidized Mitochondrial Protein Degradation and Repair in Aging and Oxidative Stress. Antioxid. Redox Signal. 2010, 13, 539–549. [Google Scholar] [CrossRef]
- Barciszewski, J.; Barciszewska, M.Z.; Siboska, G.; Rattan, S.I.; Clark, B.F. Some Unusual Nucleic Acid Bases Are Products of Hydroxyl Radical Oxidation of DNA and RNA. Mol. Biol. Rep. 1999, 26, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.M.; Parker, R. Stressing out over TRNA Cleavage. Cell 2009, 138, 215–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Q.; Xia, W.; Liu, J.-C.; Yang, J.-Y.; Lee, D.-F.; Xia, J.; Bartholomeusz, G.; Li, Y.; Pan, Y.; Li, Z.; et al. Erk Associates with and Primes GSK-3beta for Its Inactivation Resulting in Upregulation of β-Catenin. Mol. Cell 2005, 19, 159–170. [Google Scholar] [CrossRef]
- Honda, K.; Smith, M.A.; Zhu, X.; Baus, D.; Merrick, W.C.; Tartakoff, A.M.; Hattier, T.; Harris, P.L.; Siedlak, S.L.; Fujioka, H.; et al. Ribosomal RNA in Alzheimer Disease Is Oxidized by Bound Redox-Active Iron. J. Biol. Chem. 2005, 280, 20978–20986. [Google Scholar] [CrossRef] [Green Version]
- Reichmann, D.; Voth, W.; Jakob, U. Maintaining a Healthy Proteome during Oxidative Stress. Mol. Cell 2018, 69, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, J.-U.; Gray, M.J.; Jakob, U. Protein Quality Control under Oxidative Stress Conditions. J. Mol. Biol. 2015, 427, 1549–1563. [Google Scholar] [CrossRef] [Green Version]
- Mesika, R.; Reichmann, D. When Safeguarding Goes Wrong: Impact of Oxidative Stress on Protein Homeostasis in Health and Neurodegenerative Disorders. Adv. Protein Chem. Struct. Biol. 2019, 114, 221–264. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Accession # | Primer Sequence (5′–3′) | Source |
|---|---|---|---|---|
| SOD1 | Superoxide dismutase 1 | NM_174615.2 | F: AAGATGAAGAGAGGCATGTTGGA R: GATGGCAACACCGTTTTTGTC | [43] |
| SOD2 | Superoxide dismutase 2 | NM_201527.2 | F: TCTGTTGGTGTCCAAGGCTC R: AGCAGGGGGATAAGACCTGT | |
| CAT | Catalase | NM_1035386.2 | F: CTATGGCCTCCGCGATCTTT R: CGTGAGGCCAAACCTTGGTA | |
| GPX1 | Glutathione peroxidase 1 | NM_174076.3 | F: CGGGTTCGAGCCCAACT R: GCGCCTTCTCGCCATTC | |
| GPX4 | Glutathione peroxidase 4 | NM_1753024.3 | F: TGTGGTTTACGGATCCTGGC R: CCCTTGGGCTGGACTTTCAT | [44] |
| YWHAZ | tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta | NM_174814.2 | F: GCATCCCACAGACTATTTCC R: GCAAAGACAATGACAGACCA | [37] |
| H2AFZ | H2A histone family, member Z | NM_174809.2 | F: CTCACCGCAGAGGTACTTGAATT R: AGTCCAATTCTTCATCTCCACGA | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, M.; Sabry, R.; Davis, O.S.; Favetta, L.A. Effects of BPA, BPS, and BPF on Oxidative Stress and Antioxidant Enzyme Expression in Bovine Oocytes and Spermatozoa. Genes 2022, 13, 142. https://doi.org/10.3390/genes13010142
Nguyen M, Sabry R, Davis OS, Favetta LA. Effects of BPA, BPS, and BPF on Oxidative Stress and Antioxidant Enzyme Expression in Bovine Oocytes and Spermatozoa. Genes. 2022; 13(1):142. https://doi.org/10.3390/genes13010142
Chicago/Turabian StyleNguyen, Mimi, Reem Sabry, Ola S. Davis, and Laura A. Favetta. 2022. "Effects of BPA, BPS, and BPF on Oxidative Stress and Antioxidant Enzyme Expression in Bovine Oocytes and Spermatozoa" Genes 13, no. 1: 142. https://doi.org/10.3390/genes13010142
APA StyleNguyen, M., Sabry, R., Davis, O. S., & Favetta, L. A. (2022). Effects of BPA, BPS, and BPF on Oxidative Stress and Antioxidant Enzyme Expression in Bovine Oocytes and Spermatozoa. Genes, 13(1), 142. https://doi.org/10.3390/genes13010142