
No Expression Divergence despite Transcriptional Interference between Nested Protein-Coding Genes in Mammals

Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of Nested and Unnested Gene Pairs
2.2. Inference of Gene Nesting and Unnesting Events
2.3. Gene Expression Analyses
2.4. Statistical Analyses
3. Results
3.1. Prevalence and Evolutionary Dynamics of Nested Protein-Coding Genes in Mammals
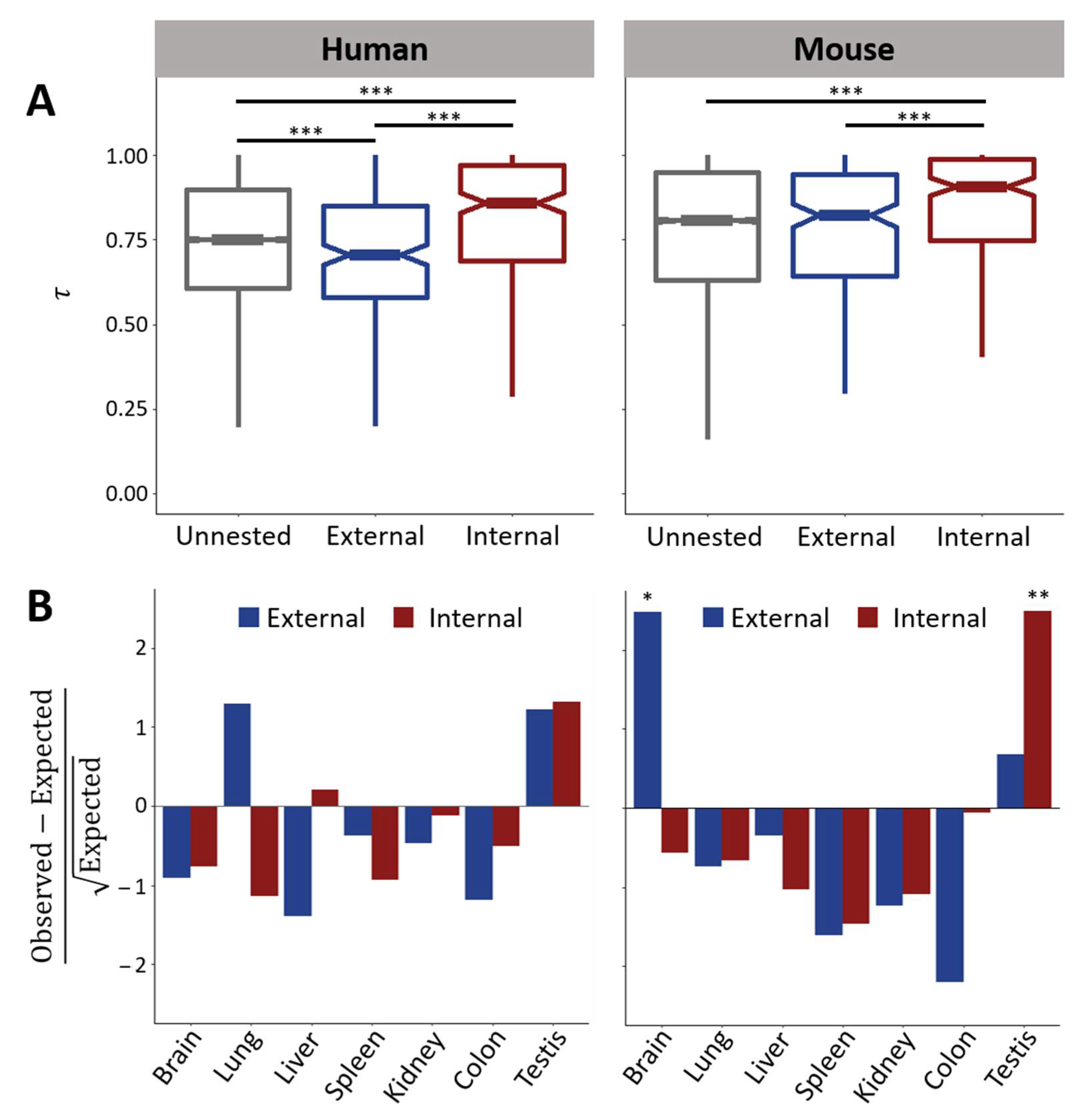
3.2. Genomic and Transcriptomic Properties of Nested Protein-Coding Genes in Mammals
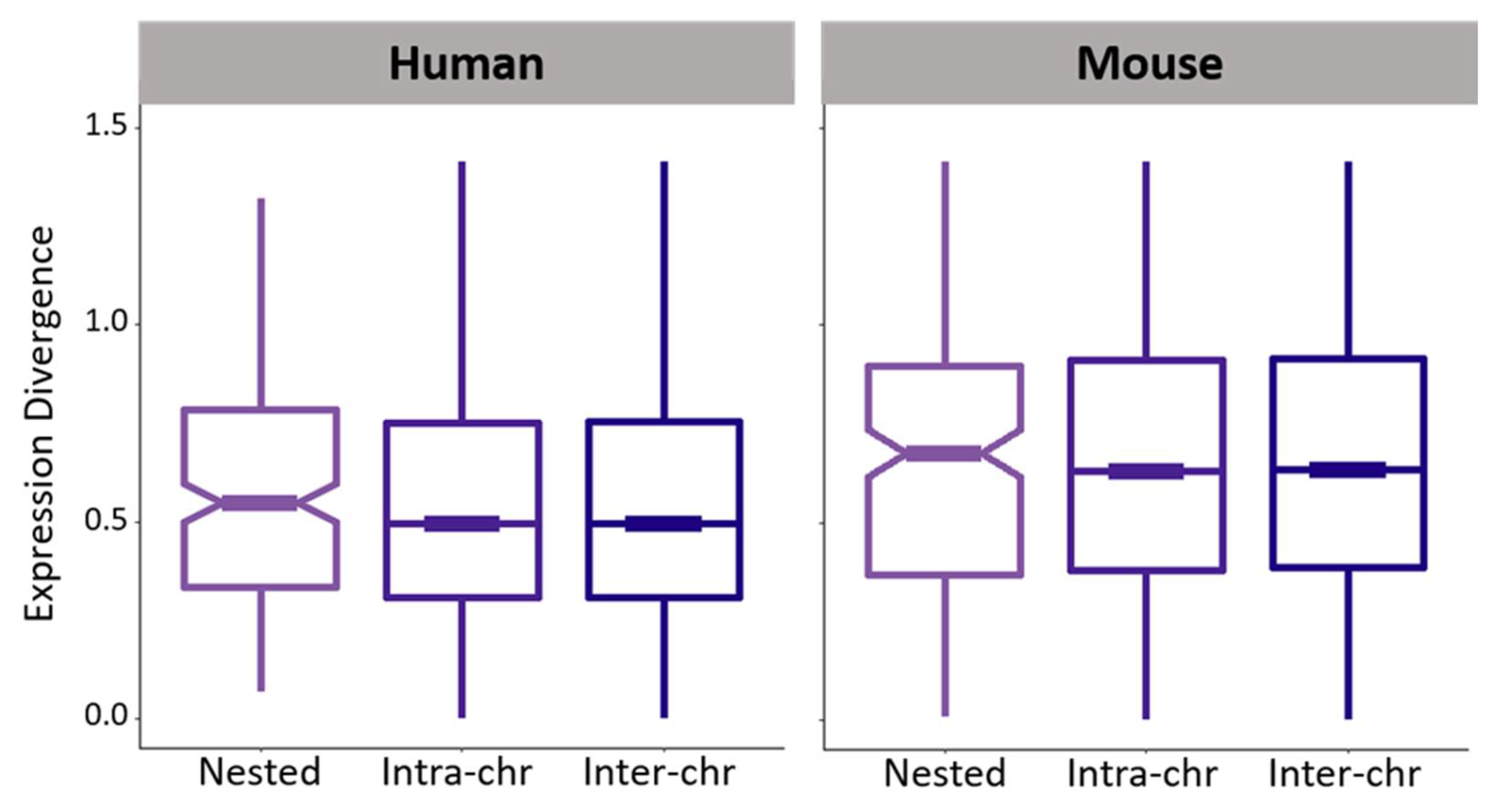
3.3. Expression Divergence between Nested Protein-Coding Genes in Mammals
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Veeramachaneni, V.; Makalowski, W.; Galdzicki, M.; Sood, R.; Makalowska, I. Mammalian Overlapping Genes: The Comparative Perspective. Genome Res. 2004, 14, 280–286. [Google Scholar] [CrossRef][Green Version]
- Yu, P.; Ma, D.; Xu, M. Nested genes in the human genome. Genomics 2005, 86, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Assis, R.; Kondrashov, A.S.; Koonin, E.V.; Kondrashov, F.A. Nested genes and increasing organizational complexity of metazoan genomes. Trends Genet. 2008, 24, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A. An Overview of Nested Genes in Eukaryotic Genomes. Eukaryot. Cell 2009, 8, 1321–1329. [Google Scholar] [CrossRef]
- Lee, Y.C.G.; Chang, H.-H. The Evolution and Functional Significance of Nested Gene Structures in Drosophila melanogaster. Genome Biol. Evol. 2013, 5, 1978–1985. [Google Scholar] [CrossRef]
- Assis, R. Transcriptional Interference Promotes Rapid Expression Divergence of Drosophila Nested Genes. Genome Biol. Evol. 2016, 8, 3149–3158. [Google Scholar] [CrossRef] [PubMed]
- Shearwin, K.E.; Callen, B.P.; Egan, J.B. Transcriptional interference—A crash course. Trends Genet 2005, 21, 339–345. [Google Scholar] [CrossRef]
- Liao, B.Y.; Zhang, J. Coexpression of linked genes in mammalian genomes is generally disadvantageous. Mol. Biol. Evol. 2008, 25, 1555–1565. [Google Scholar] [CrossRef][Green Version]
- Su, A.I.; Wiltshire, T.; Batalov, S.; Lapp, H.; Ching, K.A.; Block, D.; Zhang, J.; Soden, R.; Hayakawa, M.; Kreiman, G.; et al. A gene atlas of the mouse and human protein-encoding transcriptomes. Proc. Natl. Acad. Sci. USA 2004, 101, 6062–6067. [Google Scholar] [CrossRef]
- Chintapalli, V.R.; Wang, J.; Dow, J. Using FlyAtlas to identify better Drosophila melanogaster models of human disease. Nat. Genet. 2007, 39, 715–720. [Google Scholar] [CrossRef]
- Pereira, V.; Waxman, D.; Eyre-Walker, A. A Problem with the Correlation Coefficient as a Measure of Gene Expression Divergence. Genetics 2009, 183, 1597–1600. [Google Scholar] [CrossRef]
- Graveley, B.R.; Brooks, A.N.; Carlson, J.W.; Duff, M.O.; Landolin, J.M.; Yang, L.; Artieri, C.G.; van Baren, M.J.; Boley, N.; Booth, B.W.; et al. The developmental transcriptome of Drosophila melanogaster. Nature 2011, 471, 473–479. [Google Scholar] [CrossRef]
- Kaiser, V.B.; Zhou, Q.; Bachtrog, D. Non-random gene loss from the Drosophila miranda neo-Y chromosome. Genome Biol. Evol. 2011, 3, 1329–1337. [Google Scholar] [CrossRef]
- Merkin, J.; Russell, C.; Chen, P.; Burge, C.B. Evolutionary Dynamics of Gene and Isoform Regulation in Mammalian Tissues. Science 2012, 338, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Lin, Y.; Nery, J.R.; Urich, M.A.; Breschi, A.; Davis, C.A.; Dobin, A.; Zaleski, C.; Beer, M.A.; Chapman, W.C.; et al. Comparison of the transcriptional landscapes between human and mouse tissues. Proc. Natl. Acad. Sci. USA 2014, 111, 17224–17229. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Smedley, D.; Haider, S.; Ballester, B.; Holland, R.; London, D.; Thorisson, G.; Kasprzyk, A. BioMart—Biological queries made easy. BMC Genom. 2009, 10, 22. [Google Scholar] [CrossRef]
- Kapushesky, M.; Emam, I.; Holloway, E.; Kurnosov, P.; Zorin, A.; Malone, J.; Rustici, G.; Williams, E.; Parkinson, H.; Brazma, A. Gene Expression Atlas at the European Bioinformatics Institute. Nucleic Acids Res. 2010, 38, D690–D698. [Google Scholar] [CrossRef] [PubMed]
- Papatheodorou, I.; Fonseca, N.A.; Keays, M.; Tang, Y.A.; Barrera, E.; Bazant, W.; Burke, M.; Fullgrabe, A.; Fuentes, A.M.-P.; George, N.; et al. Expression Atlas: Gene and protein expression across multiple studies and organisms. Nucleic Acids Res. 2018, 46, D246–D251. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, Y.; Gordon, W.; Quan, J.; Xi, H.; Du, S.; Von Schack, D.; Zhang, B. Comparison of stranded and non-stranded RNA-seq transcriptome profiling and investigation of gene overlap. BMC Genom. 2015, 16, 675. [Google Scholar] [CrossRef] [PubMed]
- Yanai, I.; Benjamin, H.; Shmoish, M.; Chalifa-Caspi, V.; Shklar, M.; Ophir, R.; Bar-Even, A.; Horn-Saban, S.; Safran, M.; Domany, E.; et al. Genome-wide midrange transcription profiles reveal ex-pression level relationships in human tissue specification. Bioinformatics 2005, 21, 650–659. [Google Scholar] [CrossRef]
- Liao, B.-Y.; Zhang, J. Evolutionary Conservation of Expression Profiles Between Human and Mouse Orthologous Genes. Mol. Biol. Evol. 2005, 23, 530–540. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Fay, M.P.; Shaw, P.A. Exact and asymptotic weighted logrank tests for interval censored data: The interval R package. J. Stat. Soft 2010, 36, 1–34. [Google Scholar] [CrossRef]
- Itsara, A.; Wu, H.; Smith, J.D.; Nickerson, D.A.; Romieu, I.; London, S.J.; Eichler, E.E. De novo rates and selection of large copy number variation. Genome Res. 2010, 20, 1469–1481. [Google Scholar] [CrossRef]
- Schrider, D.R.; Houle, D.; Lynch, M.; Hahn, M. Rates and Genomic Consequences of Spontaneous Mutational Events in Drosophila melanogaster. Genetics 2013, 194, 937–954. [Google Scholar] [CrossRef]
- Zhang, J. Evolution by gene duplication: An update. Trends Ecol. Evol. 2003, 18, 292–298. [Google Scholar] [CrossRef]
- Francis, W.R.; Wörheide, G. Similar Ratios of Introns to Intergenic Sequence across Animal Genomes. Genome Biol. Evol. 2017, 9, 1582–1598. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Sung, W.; Morris, K.; Coffey, N.; Landry, C.R.; Dopman, E.B.; Dickinson, W.J.; Okamoto, K.; Kulkarni, S.; Hartl, D.L.; et al. A genome-wide view of the spectrum of spontaneous mutations in yeast. Proc. Natl. Acad. Sci. USA 2008, 105, 9272–9277. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, K.J.; Farslow, J.C.; Fitzpatrick, K.A.; Lynch, M.; Katju, V.; Bergthorsson, U. High Spontaneous Rate of Gene Duplication in Caenorhabditis elegans. Curr. Biol. 2011, 21, 306–310. [Google Scholar] [CrossRef]
- Keith, N.; Tucker, A.E.; Jackson, C.E.; Sung, W.; Lledó, J.I.L.; Schrider, D.R.; Schaack, S.; Dudycha, J.L.; Ackerman, M.; Younge, A.J.; et al. High mutational rates of large-scale duplication and deletion in Daphnia pulex. Genome Res. 2015, 26, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Konrad, A.; Flibotte, S.; Taylor, J.; Waterston, R.H.; Moerman, D.G.; Bergthorsson, U.; Katju, V. Mutational and transcriptional landscape of spontaneous gene duplications and deletions in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2018, 115, 7386–7391. [Google Scholar] [CrossRef] [PubMed]
- Betrán, E.; Thornton, K.; Long, M. Retroposed New Genes Out of the X in Drosophila. Genome Res. 2002, 12, 1854–1859. [Google Scholar] [CrossRef]
- Vinckenbosch, N.; Dupanloup, I.; Kaessmann, H. Evolutionary fate of retroposed gene copies in the human genome. Proc. Natl. Acad. Sci. USA 2006, 103, 3220–3225. [Google Scholar] [CrossRef]
- Kaessmann, H. Origins, evolution, and phenotypic impact of new genes. Genome Res. 2010, 20, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Assis, R.; Bachtrog, D. Neofunctionalization of young duplicate genes in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, 17409–17414. [Google Scholar] [CrossRef]
- Wu, N.-D.; Wang, X.; Li, Y.; Zeng, L.; Irwin, D.M.; Zhang, Y.-P. “Out of pollen” hypothesis for origin of new genes in flowering plants: Study from Arabidopsis thaliana. Genome Biol. Evol. 2014, 6, 2822–2829. [Google Scholar] [CrossRef]
- Assis, R.; Bachtrog, D. Rapid divergence and diversification of mammalian duplicate gene functions. BMC Evol. Biol. 2015, 15, 138. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Assis, R. Rapid functional divergence after small-scale gene duplication in grasses. BMC Evol. Biol. 2019, 19, 97. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B. Fundamental concepts in genetics: Effective population size and patterns of molecular evolution and variation. Nat. Rev. Genet. 2009, 10, 195–205. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Same | Opposite | Same vs. Opposite * | |
|---|---|---|---|
| Human | |||
| Mouse | |||
| Human vs. Mouse ** | |||
| External | Internal | External vs. Internal * | |
|---|---|---|---|
| Human | |||
| Mouse | |||
| Human vs. Mouse ** | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assis, R. No Expression Divergence despite Transcriptional Interference between Nested Protein-Coding Genes in Mammals. Genes 2021, 12, 1381. https://doi.org/10.3390/genes12091381
Assis R. No Expression Divergence despite Transcriptional Interference between Nested Protein-Coding Genes in Mammals. Genes. 2021; 12(9):1381. https://doi.org/10.3390/genes12091381
Chicago/Turabian StyleAssis, Raquel. 2021. "No Expression Divergence despite Transcriptional Interference between Nested Protein-Coding Genes in Mammals" Genes 12, no. 9: 1381. https://doi.org/10.3390/genes12091381
APA StyleAssis, R. (2021). No Expression Divergence despite Transcriptional Interference between Nested Protein-Coding Genes in Mammals. Genes, 12(9), 1381. https://doi.org/10.3390/genes12091381