
Epigenetic Regulation of Circadian Clocks and Its Involvement in Drug Addiction


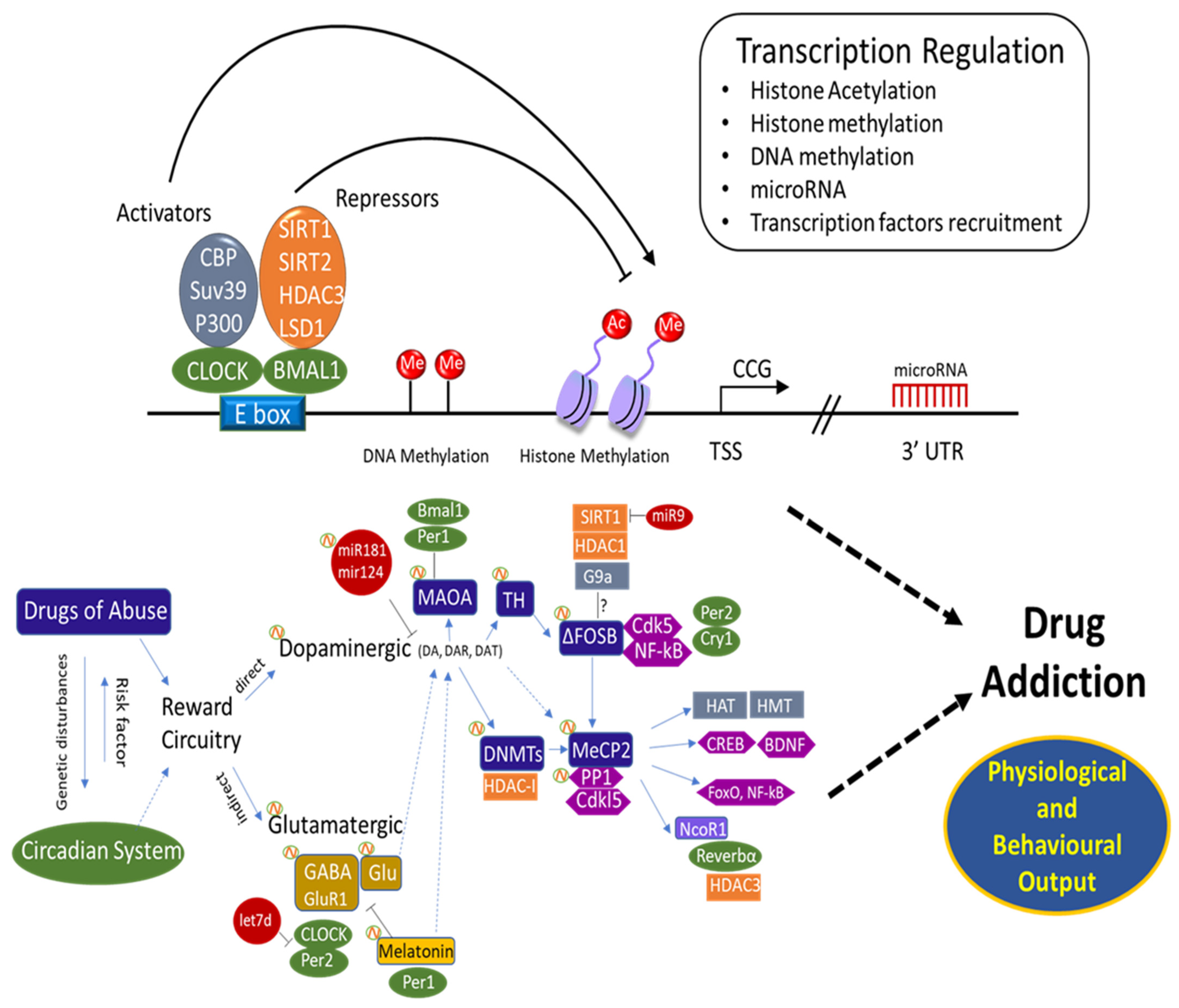{kind=link}
Abstract
1. The Basics of Epigenetics
2. The Circadian Timing System
3. Epigenetics, Circadian Timing and Addiction
4. Epigenetics in Circadian System
5. Addiction and Epigenetics
6. Clock Genes and Addiction
7. Epigenetic Connection between Circadian Rhythms and Addiction
8. Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dupont, C.; Armant, D.R.; Brenner, C.A. Epigenetics: Definition, mechanisms and clinical perspective. Semin. Reprod. Med. 2009, 27, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. Epigenetic mechanisms of drug addiction. Neuropharmacology 2014, 76, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Anglard, P.; Zwiller, J. Chapter 9-Cocaine and Epigenetics: An Overview. In The Neuroscience of Cocaine; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 81–88. ISBN 978-0-12-803750-8. [Google Scholar]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef]
- Robison, A.J.; Nestler, E.J. Transcriptional and Epigenetic Mechanisms of Addiction. Nat. Rev. Neurosci. 2012, 12, 623–637. [Google Scholar] [CrossRef]
- Zwiller, J. Addiction et régulations épigénétiques. Med. Sci. 2015, 31, 439–446. [Google Scholar] [CrossRef]
- Gräff, J.; Tsai, L.-H. Histone acetylation: Molecular mnemonics on the chromatin. Nat. Rev. Neurosci. 2013, 14, 97–111. [Google Scholar] [CrossRef]
- Bastle, R.M.; Neisewander, J.L. Epigenetics and Drug Abuse. In Recent Advances in Drug Addiction Research and Clinical Applications; InTech: London, UK, 2016. [Google Scholar]
- de Ruijter, A.J.M.; van Gennip, A.H.; Caron, H.N.; Kemp, S.; van Kuilenburg, A.B.P. Histone deacetylases (HDACs): Characterization of the classical HDAC family. Biochem. J. 2003, 370, 737–749. [Google Scholar] [CrossRef]
- Rice, J.C.; Allis, C.D. Histone methylation versus histone acetylation: New insights into epigenetic regulation. Curr. Opin. Cell Biol. 2001, 13, 263–273. [Google Scholar] [CrossRef]
- Greer, E.L.; Shi, Y. Histone methylation: A dynamic mark in health, disease and inheritance. Nat. Rev. Genet. 2012, 13, 343–357. [Google Scholar] [CrossRef]
- Bannister, A.J.; Schneider, R.; Kouzarides, T. Histone Methylation: Dynamic or Static? Cell 2002, 109, 801–806. [Google Scholar] [CrossRef]
- Cadet, J.L.; McCoy, M.T.; Jayanthi, S. Epigenetics and addiction. Clin. Pharmacol. Ther. 2016, 99, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Falkenberg, K.J.; Johnstone, R.W. Histone deacetylases and their inhibitors in cancer, neurological diseases and immune disorders. Nat. Rev. Drug Discov. 2014, 13, 673–691. [Google Scholar] [CrossRef]
- DiTacchio, L.; Le, H.D.; Vollmers, C.; Hatori, M.; Witcher, M.; Secombe, J.; Panda, S. Histone lysine demethylase JARID1a activates CLOCK-BMAL1 and influences the circadian clock. Science 2011, 333, 1881–1885. [Google Scholar] [CrossRef] [PubMed]
- Challen, G.A.; Sun, D.; Mayle, A.; Jeong, M.; Luo, M.; Rodriguez, B.; Mallaney, C.; Celik, H.; Yang, L.; Xia, Z.; et al. Dnmt3a and Dnmt3b have overlapping and distinct functions in hematopoietic stem cells. Cell Stem Cell 2014, 15, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Karnik, R.; Gu, H.; Ziller, M.J.; Clement, K.; Tsankov, A.M.; Akopian, V.; Gifford, C.A.; Donaghey, J.; Galonska, C.; et al. Targeted disruption of DNMT1, DNMT3A and DNMT3B in human embryonic stem cells. Nat. Genet. 2015, 47, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Riggs, A.D. DNA methylation and demethylation in mammals. J. Biol. Chem. 2011, 286, 18347–18353. [Google Scholar] [CrossRef]
- Rasmussen, K.D.; Helin, K. Role of TET enzymes in DNA methylation, development, and cancer. Genes Dev. 2016, 30, 733–750. [Google Scholar] [CrossRef]
- Tan, L.; Shi, Y.G. Tet family proteins and 5-hydroxymethylcytosine in development and disease. Development 2012, 139, 1895–1902. [Google Scholar] [CrossRef]
- Guo, J.U.; Su, Y.; Zhong, C.; Ming, G.; Song, H. Emerging roles of TET proteins and 5-hydroxymethylcytosines in active DNA demethylation and beyond. Cell Cycle 2011, 10, 2662–2668. [Google Scholar] [CrossRef]
- Goll, M.G.; Bestor, T.H. Eukaryotic cytosine methyltransferases. Annu. Rev. Biochem. 2005, 74, 481–514. [Google Scholar] [CrossRef]
- Guo, J.U.; Su, Y.; Zhong, C.; Ming, G.; Song, H. Hydroxylation of 5-Methylcytosine by TET1 Promotes Active DNA Demethylation in the Adult Brain. Cell 2011, 145, 423–434. [Google Scholar] [CrossRef]
- Khare, T.; Pai, S.; Koncevicius, K.; Pal, M.; Kriukiene, E.; Liutkeviciute, Z.; Irimia, M.; Jia, P.; Ptak, C.; Xia, M.; et al. 5-hmC in the brain is abundant in synaptic genes and shows differences at the exon-intron boundary. Nat. Struct. Mol. Biol. 2012, 19, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Kriaucionis, S.; Heintz, N. The nuclear DNA base 5-hydroxymethylcytosine is present in purkinje neurons and the brain. Science 2009, 324, 929–930. [Google Scholar] [CrossRef]
- Doerfler, W. Patterns of de novo DNA methylation and promoter inhibition: Studies on the adenovirus and the human genomes. EXS 1993, 64, 262–299. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.M., 3rd; Kim, D.; Berquist, B.R.; Sigurdson, A.J. Variation in base excision repair capacity. Mutat. Res. 2011, 711, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Kenny, P.J. Epigenetics, microRNA, and addiction. Dialogues Clin. Neurosci. 2014, 16, 335–344. [Google Scholar]
- Bali, P.; Kenny, P.J. MicroRNAs and drug addiction. Front. Genet. 2013, 4, 1–8. [Google Scholar] [CrossRef]
- Surendran, S.; Jideonwo, V.N.; Merchun, C.; Ahn, M.; Murray, J.; Ryan, J.; Dunn, K.W.; Kota, J.; Morral, N. Gene targets of mouse miR-709: Regulation of distinct pools. Sci. Rep. 2016, 6, 18958. [Google Scholar] [CrossRef]
- Yao, Q.; Chen, Y.; Zhou, X. The roles of microRNAs in epigenetic regulation. Curr. Opin. Chem. Biol. 2019, 51, 11–17. [Google Scholar] [CrossRef]
- Mosig, R.A.; Kojima, S. Timing without coding: How do long non-coding RNAs regulate circadian rhythms? Semin. Cell Dev. Biol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Challet, E.; Kalsbeek, A. Editorial: Circadian Rhythms and Metabolism. Front. Endocrinol. 2017, 8, 201. [Google Scholar] [CrossRef] [PubMed]
- Forde, L.A.; Kalsi, G. Addiction and the role of circadian genes. J. Stud. Alcohol Drugs 2017, 78, 645–653. [Google Scholar] [CrossRef]
- Gamble, K.L.; Berry, R.; Frank, S.J.; Young, M.E. Circadian clock control of endocrine factors. Nat. Rev. Endocrinol. 2014, 10, 466–475. [Google Scholar] [CrossRef]
- Feng, D.; Lazar, M.A. Clocks, metabolism, and the epigenome. Mol. Cell 2012, 47, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S.; Hong, H.-K.; Ko, C.H.; McDearmon, E.L. The Genetics of Mammalian Circadian Order and Disorder: Implications for Physiology and Disease. Nat. Rev. Genet. 2008, 9, 764–775. [Google Scholar] [CrossRef]
- Ozburn, A.R.; Larson, E.B.; Self, D.W.; McClung, C.A. Cocaine self-administration behaviors in ClockΔ19 mice. Psychopharmacology 2012, 223, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; Yi, C.-X.; Cailotto, C.; la Fleur, S.E.; Fliers, E.; Buijs, R.M. Mammalian clock output mechanisms. Essays Biochem. 2011, 49, 137–151. [Google Scholar] [CrossRef]
- Liu, C.; Chung, M. Genetics and epigenetics of circadian rhythms and their potential roles in neuropsychiatric disorders. Neurosci. Bull. 2015, 31, 141–159. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Merrow, M.; Roenneberg, T.; Foster, R.G. Neurobiology of Circadian Timing. Preface. Prog. Brain Res. 2012, 199, xi–xii. [Google Scholar]
- Balsalobre, A. Clock genes in mammalian peripheral tissues. Cell Tissue Res. 2002, 309, 193–199. [Google Scholar] [CrossRef]
- Abe, M.; Herzog, E.D.; Yamazaki, S.; Straume, M.; Tei, H.; Sakaki, Y.; Menaker, M.; Block, G.D. Circadian rhythms in isolated brain regions. J. Neurosci. 2002, 22, 350–356. [Google Scholar] [CrossRef]
- Crosio, C.; Cermakian, N.; Allis, C.D.; Sassone-Corsi, P. Light induces chromatin modification in cells of the mammalian circadian clock. Nat. Neurosci. 2000, 3, 1241–1247. [Google Scholar] [CrossRef]
- Chaix, A.; Zarrinpar, A.; Panda, S. The circadian coordination of cell biology. J. Cell Biol. 2016, 215, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, J.C. Molecular Bases for Circadian Clocks. Cell 1999, 96, 271–290. [Google Scholar] [CrossRef]
- Masri, S.; Sassone-Corsi, P. Plasticity and specificity of the circadian epigenome. Nat. Neurosci. 2010, 13, 1324–1329. [Google Scholar] [CrossRef]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef]
- Wager-Smith, K.; Kay, S.A. Circadian rhythm genetics: From flies to mice to humans. Nat. Genet. 2000, 26, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, S.; Yamaguchi, S.; Matsuo, T.; Ishida, Y.; Okamura, H. Antagonistic role of E4BP4 and PAR proteins in the circadian oscillatory mechanism. Genes Dev. 2001, 15, 995–1006. [Google Scholar] [CrossRef]
- Gachon, F.; Olela, F.F.; Schaad, O.; Descombes, P.; Schibler, U. The circadian PAR-domain basic leucine zipper transcription factors DBP, TEF, and HLF modulate basal and inducible xenobiotic detoxification. Cell Metab. 2006, 4, 25–36. [Google Scholar] [CrossRef]
- Koike, N.; Yoo, S.H.; Huang, H.C.; Kumar, V.; Lee, C.; Kim, T.K.; Takahashi, J.S. Transcriptional architecture and chromatin landscape of the core circadian clock in mammals. Science 2012, 338, 349–354. [Google Scholar] [CrossRef]
- Ripperger, J.A.; Schibler, U. Rhythmic CLOCK-BMAL1 binding to multiple E-box motifs drives circadian Dbp transcription and chromatin transitions. Nat. Genet. 2006, 38, 369–374. [Google Scholar] [CrossRef]
- Doi, M.; Hirayama, J.; Sassone-Corsi, P. Circadian Regulator CLOCK Is a Histone Acetyltransferase. Cell 2006, 125, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Etchegaray, J.P.; Lee, C.; Wade, P.A.; Reppert, S.M. Rhythmic histone acetylation underlies transcription in the mammalian circadian clock. Nature 2003, 421, 177–182. [Google Scholar] [CrossRef]
- Hirayama, J.; Sahar, S.; Grimaldi, B.; Tamaru, T.; Takamatsu, K.; Nakahata, Y.; Sassone-Corsi, P. CLOCK-mediated acetylation of BMAL1 controls circadian function. Nature 2007, 450, 1086–1090. [Google Scholar] [CrossRef]
- Asher, G.; Gatfield, D.; Stratmann, M.; Reinke, H.; Dibner, C.; Kreppel, F.; Mostoslavsky, R.; Alt, F.W.; Schibler, U. SIRT1 Regulates Circadian Clock Gene Expression through PER2 Deacetylation. Cell 2008, 134, 317–328. [Google Scholar] [CrossRef]
- Nakahata, Y.; Kaluzova, M.; Grimaldi, B.; Sahar, S.; Hirayama, J.; Chen, D.; Guarente, L.P.; Sassone-Corsi, P. The NAD+-Dependent Deacetylase SIRT1 Modulates CLOCK-Mediated Chromatin Remodeling and Circadian Control. Cell 2008, 134, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, I.A.; Mehler, M.F. Epigenetics of sleep and chronobiology. Curr. Neurol. Neurosci. Rep. 2014, 14, 1–16. [Google Scholar] [CrossRef]
- Curtis, A.M.; Seo, S.B.; Westgate, E.J.; Rudic, R.D.; Smyth, E.M.; Chakravarti, D.; FitzGerald, G.A.; McNamara, P. Histone Acetyltransferase-dependent Chromatin Remodeling and the Vascular Clock. J. Biol. Chem. 2004, 279, 7091–7097. [Google Scholar] [CrossRef]
- Johansson, A.S.; Brask, J.; Owe-Larsson, B.; Hetta, J.; Lundkvist, G.B.S. Valproic acid phase shifts the rhythmic expression of PERIOD2::LUCIFERASE. J. Biol. Rhythms 2011, 26, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Alenghat, T.; Meyers, K.; Mullican, S.E.; Leitner, K.; Adeniji-Adele, A.; Avila, J.; Bućan, M.; Ahima, R.S.; Kaestner, K.H.; Lazar, M.A. Nuclear receptor corepressor and histone deacetylase 3 govern circadian metabolic physiology. Nature 2008, 456, 997–1000. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Liu, T.; Sun, Z.; Bugge, A.; Mullican, S.E.; Alenghat, T.; Liu, X.S.; Lazar, M.A. A circadian rhythm orchestrated by histone deacetylase 3 controls hepatic lipid metabolism. Science 2011, 331, 1315–1319. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Mucke, L. Paths of Convergence: Sirtuins in Aging and Neurodegeneration. Neuron 2008, 58, 10–14. [Google Scholar] [CrossRef]
- Chang, H.C.; Guarente, L. XSIRT1 mediates central circadian control in the SCN by a mechanism that decays with aging. Cell 2013, 153, 1448. [Google Scholar] [CrossRef]
- Michán, S.; Li, Y.; Chou, M.M.H.; Parrella, E.; Ge, H.; Long, J.M.; Allard, J.S.; Lewis, K.; Miller, M.; Xu, W.; et al. SIRT1 is essential for normal cognitive function and synaptic plasticity. J. Neurosci. 2010, 30, 9695–9707. [Google Scholar] [CrossRef]
- Gao, J.; Wang, W.Y.; Mao, Y.W.; Gräff, J.; Guan, J.S.; Pan, L.; Mak, G.; Kim, D.; Su, S.C.; Tsai, L.H. A novel pathway regulates memory and plasticity via SIRT1 and miR-134. Nature 2010, 466, 1105–1109. [Google Scholar] [CrossRef]
- Papazyan, R.; Zhang, Y.; Lazar, M.A. Genetic and epigenomic mechanisms of mammalian circadian transcription. Nat. Struct. Mol. Biol. 2016, 23, 1045–1052. [Google Scholar] [CrossRef]
- Duong, H.A.; Weitz, C.J. Temporal orchestration of repressive chromatin modifiers by circadian clock Period complexes. Nat. Struct. Mol. Biol. 2014, 21, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Etchegaray, J.P.; Yang, X.; Debruyne, J.P.; Peters, A.H.F.M.; Weaver, D.R.; Jenuwein, T.; Reppert, S.M. The polycomb group protein EZH2 is required for mammalian circadian clock function. J. Biol. Chem. 2006, 281, 21209–21215. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Yang, Y.; Guo, G.L.; Qin, X.M. Genetic and epigeneticregulations of mammalian circadian rhythms. Yi Chuan Hered 2017, 39, 1122–1137. [Google Scholar]
- Nam, H.J.; Boo, K.; Kim, D.; Han, D.H.; Choe, H.K.; Kim, C.R.; Sun, W.; Kim, H.; Kim, K.; Lee, H.; et al. Phosphorylation of LSD1 by PKCα Is Crucial for Circadian Rhythmicity and Phase Resetting. Mol. Cell 2014, 53, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.A.; Covington, M.F.; DiTacchio, L.; Vollmers, C.; Panda, S.; Harmer, S.L. Jumonji domain protein JMJD5 functions in both the plant and human circadian systems. Proc. Natl. Acad. Sci. USA 2010, 107, 21623–21628. [Google Scholar] [CrossRef]
- Pastor, W.A.; Pape, U.J.; Huang, Y.; Henderson, H.R.; Lister, R.; Ko, M.; McLoughlin, E.M.; Brudno, Y.; Mahapatra, S.; Kapranov, P.; et al. Genome-wide mapping of 5-hydroxymethylcytosine in embryonic stem cells. Nature 2011, 473, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Ficz, G.; Branco, M.R.; Seisenberger, S.; Santos, F.; Krueger, F.; Hore, T.A.; Marques, C.J.; Andrews, S.; Reik, W. Dynamic regulation of 5-hydroxymethylcytosine in mouse ES cells and during differentiation. Nature 2011, 473, 398–404. [Google Scholar] [CrossRef]
- Kim, J.K.; Samaranayake, M.; Pradhan, S. Epigenetic mechanisms in mammals. Cell. Mol. Life Sci. 2009, 66, 596–612. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.S.P.; Srivastava, G.P.; Yu, L.; Chibnik, L.B.; Xu, J.; Buchman, A.S.; Schneider, J.A.; Myers, A.J.; Bennett, D.A.; De Jager, P.L. 24-Hour Rhythms of DNA Methylation and Their Relation with Rhythms of RNA Expression in the Human Dorsolateral Prefrontal Cortex. PLoS Genet. 2014, 10, e1004792. [Google Scholar] [CrossRef]
- Azzi, A.; Dallmann, R.; Casserly, A.; Rehrauer, H.; Patrignani, A.; Maier, B.; Kramer, A.; Brown, S.A. Circadian behavior is light-reprogrammed by plastic DNA methylation. Nat. Neurosci. 2014, 17, 377–382. [Google Scholar] [CrossRef]
- Vollmers, C.; Schmitz, R.J.; Nathanson, J.; Yeo, G.; Ecker, J.R.; Panda, S. Circadian oscillations of protein-coding and regulatory RNAs in a highly dynamic mammalian liver epigenome. Cell Metab. 2012, 16, 833–845. [Google Scholar] [CrossRef]
- Bönsch, D.; Hothorn, T.; Krieglstein, C.; Koch, M.; Nehmer, C.; Lenz, B.; Reulbach, U.; Kornhuber, J.; Bleich, S. Daily variations of homocysteine concentration may influence methylation of DNA in normal healthy individuals. Chronobiol. Int. 2007, 24, 315–326. [Google Scholar] [CrossRef]
- Guéant, J.-L.; Caillerez-Fofou, M.; Battaglia-Hsu, S.; Alberto, J.-M.; Freund, J.-N.; Dulluc, I.; Adjalla, C.; Maury, F.; Merle, C.; Nicolas, J.-P.; et al. Molecular and cellular effects of vitamin B12 in brain, myocardium and liver through its role as co-factor of methionine synthase. Biochimie 2013, 95, 1033–1040. [Google Scholar] [CrossRef]
- Geoffroy, A.; Kerek, R.; Pourié, G.; Helle, D.; Guéant, J.-L.; Daval, J.-L.; Bossenmeyer-Pourié, C. Late Maternal Folate Supplementation Rescues from Methyl Donor Deficiency-Associated Brain Defects by Restoring Let-7 and miR-34 Pathways. Mol. Neurobiol. 2017, 54, 5017–5033. [Google Scholar] [CrossRef]
- Krishna, S.M.; Dear, A.; Craig, J.M.; Norman, P.E.; Golledge, J. The potential role of homocysteine mediated DNA methylation and associated epigenetic changes in abdominal aortic aneurysm formation. Atherosclerosis 2013, 228, 295–305. [Google Scholar] [CrossRef]
- Masoomi, M.; Azdaki, N.; Shahouzehi, B. Elevated Plasma Homocysteine Concentration in Opium-Addicted Individuals. Addict. Health 2015, 7, 149–156. [Google Scholar]
- Lutz, U.C. Alterations in homocysteine metabolism among alcohol dependent patients—Clinical, pathobiochemical and genetic aspects. Curr. Drug Abuse Rev. 2008, 1, 47–55. [Google Scholar] [CrossRef]
- Massart, R.; Freyburger, M.; Suderman, M.; Paquet, J.; El Helou, J.; Belanger-Nelson, E.; Rachalski, A.; Koumar, O.C.; Carrier, J.; Szyf, M.; et al. The genome-wide landscape of DNA methylation and hydroxymethylation in response to sleep deprivation impacts on synaptic plasticity genes. Transl. Psychiatry 2014, 4, e347. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Stevens, R.G.; Hoffman, A.E.; Tjonneland, A.; Vogel, U.B.; Zheng, T.; Hansen, J. Epigenetic impact of long-term shiftwork: Pilot evidence from circadian genes and whole-genome methylation analysis. Chronobiol. Int. 2011, 28, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Ma, S.; Zhang, Y.; Wang, T.; Zhou, M.; Wang, Z.; Zhang, J. Daily variation in global and local DNA methylation in mouse livers. PLoS ONE 2015, 10, e0118101. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, F.; Shimba, S.; Takumi, S.; Sano, T.; Suzuki, T.; Bao, J.; Ohwada, M.; Ehara, T.; Ogawa, Y.; Nohara, K. Diurnal expression of Dnmt3b mRNA in mouse liver is regulated by feeding and hepatic clockwork. Epigenetics 2012, 7, 1046–1056. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, K.; Kumar, V.; Koike, N.; Kim, T.K.; Chong, J.; Buhr, E.D.; Whiteley, A.R.; Low, S.S.; Omura, C.; Fenner, D.; et al. Usf1, a suppressor of the circadian Clock mutant, reveals the nature of the DNA-binding of the CLOCK:BMAL1 complex in mice. Elife 2013, 2013, e00426. [Google Scholar] [CrossRef]
- Hu, S.; Wan, J.; Su, Y.; Song, Q.; Zeng, Y.; Nguyen, H.N.; Shin, J.; Cox, E.; Rho, H.S.; Woodard, C.; et al. DNA methylation presents distinct binding sites for human transcription factors. Elife 2013, 2013, e00726. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Miller, C.; Miraglia, L.J.; Romero, A.; Mure, L.S.; Panda, S.; Kay, S.A. A genome-wide microRNA screen identifies the microRNA-183/96/182 cluster as a modulator of circadian rhythms. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Sehgal, A. Regulation of circadian behavioral output via a MicroRNA-JAK/STAT circuit. Cell 2012, 148, 765–779. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.Y.M.; Papp, J.W.; Varlamova, O.; Dziema, H.; Russell, B.; Curfman, J.P.; Nakazawa, T.; Shimizu, K.; Okamura, H.; Impey, S.; et al. microRNA Modulation of Circadian-Clock Period and Entrainment. Neuron 2007, 54, 813–829. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhu, J.; Liu, Y.; Chen, Y.; Li, Y.; Chen, S.; Li, T.; Dang, Y.; Chen, T. Chronic methamphetamine regulates the expression of MicroRNAs and putative target genes in the nucleus accumbens of mice. J. Neurosci. Res. 2015, 93, 1600–1610. [Google Scholar] [CrossRef] [PubMed]
- Darcq, E.; Warnault, V.; Phamluong, K.; Besserer, G.M.; Liu, F.; Ron, D. MicroRNA-30a-5p in the prefrontal cortex controls the transition from moderate to excessive alcohol consumption. Mol. Psychiatry 2015, 20, 1240–1250. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; D’Alessandro, M.; Lee, C. MiRNAs are required for generating a time delay critical for the circadian oscillator. Curr. Biol. 2013, 23, 1959–1968. [Google Scholar] [CrossRef]
- Kiessling, S.; Ucar, A.; Chowdhury, K.; Oster, H.; Eichele, G. Genetic background-dependent effects of murine micro RNAs on circadian clock function. PLoS ONE 2017, 12, e0176547. [Google Scholar] [CrossRef]
- Wanet, A.; Tacheny, A.; Arnould, T.; Renard, P. miR-212/132 expression and functions: Within and beyond the neuronal compartment. Nucleic Acids Res. 2012, 40, 4742–4753. [Google Scholar] [CrossRef]
- Alvarez-Saavedra, M.; Antoun, G.; Yanagiya, A.; Oliva-Hernandez, R.; Cornejo-Palma, D.; Perez-Iratxeta, C.; Sonenberg, N.; Cheng, H.Y.M. miRNA-132 orchestrates chromatin remodeling and translational control of the circadian clock. Hum. Mol. Genet. 2011, 20, 731–751. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, C.; Okamoto, Y.; Aoyama, K.; Nakaki, T. MicroRNA: A Key Player for the Interplay of Circadian Rhythm Abnormalities, Sleep Disorders and Neurodegenerative Diseases. Clocks Sleep 2020, 2, 22. [Google Scholar] [CrossRef]
- Chen, W.; Liu, Z.; Li, T.; Zhang, R.; Xue, Y.; Zhong, Y.; Bai, W.; Zhou, D.; Zhao, Z. Regulation of Drosophila circadian rhythms by miRNA let-7 is mediated by a regulatory cycle. Nat. Commun. 2014, 5, 5549. [Google Scholar] [CrossRef]
- Figueredo, D.D.S.; Barbosa, M.R.; Gitaí, D.L.G.; De Andrade, T.G. Predicted microRNAs for mammalian circadian Rhythm. J. Biol. Rhythm 2013, 28, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Coon, S.L.; Munson, P.J.; Cherukuri, P.F.; Sugden, D.; Rath, M.F.; Møller, M.; Clokie, S.J.H.; Fu, C.; Olanich, M.E.; Rangel, Z.; et al. Circadian changes in long noncoding RNAs in the pineal gland. Proc. Natl. Acad. Sci. USA 2012, 109, 13319–13324. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Ye, Q.; Anson, S.R.; Yang, J.; Xiao, G.; Kowbel, D.; Glass, N.L.; Crosthwaite, S.K.; Liu, Y. Transcriptional interference by antisense RNA is required for circadian clock function. Nature 2014, 514, 650–653. [Google Scholar] [CrossRef]
- Hasin, D.S.; O’Brien, C.P.; Auriacombe, M.; Borges, G.; Bucholz, K.; Budney, A.; Compton, W.M.; Crowley, T.; Ling, W.; Petry, N.M.; et al. DSM-5 criteria for substance use disorders: Recommendations and rationale. Am. J. Psychiatry 2013, 170, 834–851. [Google Scholar] [CrossRef]
- Kalivas, P.W. Cocaine and amphetamine-like psychostimulants: Neuro circuitry and glutamate neuroplasticity. Dialogues Clin. Neurosci. 2007, 9, 389–397. [Google Scholar]
- Imbesi, M.; Yildiz, S.; Dirim Arslan, A.; Sharma, R.; Manev, H.; Uz, T. Dopamine receptor-mediated regulation of neuronal “clock” gene expression. Neuroscience 2009, 158, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.E.; Berridge, K.C. The neural basis of drug craving: An incentive-sensitization theory of addiction. Brain Res. Brain Res. Rev. 1993, 18, 247–291. [Google Scholar] [CrossRef]
- Nestler, E.J. Molecular basis of long-term plasticity underlying addiction. Nat. Rev. Neurosci. 2001, 2, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, J.-B.; Takemori, H.; Grosch-Dirrig, S.; Bertorello, A.; Zwiller, J. Cocaine induces the expression of MEF2C transcription factor in rat striatum through activation of SIK1 and phosphorylation of the histone deacetylase HDAC5. Synapse 2012, 66, 61–70. [Google Scholar] [CrossRef]
- Nestler, E.J. Transcriptional mechanisms of addiction: Role of ΔFosB. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 3245–3255. [Google Scholar] [CrossRef] [PubMed]
- Perrotti, L.I.; Weaver, R.R.; Robison, B.; Renthal, W.; Maze, I.; Yazdani, S.; Elmore, R.G.; Knapp, D.J.; Selley, D.E.; Martin, B.R.; et al. Distinct patterns of ΔFosB induction in brain by drugs of abuse. Synapse 2008, 62, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Hiroi, N.; Brown, J.R.; Haile, C.N.; Ye, H.; Greenberg, M.E.; Nestler, E.J. FosB mutant mice: Loss of chronic cocaine induction of Fos-related proteins and heightened sensitivity to cocaine’s psychomotor and rewarding effects. Proc. Natl. Acad. Sci. USA 1997, 94, 10397–10402. [Google Scholar] [CrossRef]
- Nestler, E.J. Transcriptional mechanisms of drug addiction. Clin. Psychopharmacol. Neurosci. Off. Sci. J. Korean Coll. Neuropsychopharmacol. 2012, 10, 136–143. [Google Scholar] [CrossRef]
- Kelz, M.B.; Chen, J.; Carlezon, W.A.; Whisler, K.; Gilden, L.; Beckmann, A.M.; Steffen, C.; Zhang, Y.J.; Marotti, L.; Self, D.W.; et al. Expression of the transcription factor ΔFosB in the brain controls sensitivity to cocaine. Nature 1999, 401, 272–276. [Google Scholar] [CrossRef]
- Zachariou, V.; Bolanos, C.A.; Selley, D.E.; Theobald, D.; Cassidy, M.P.; Kelz, M.B.; Shaw-Lutchman, T.; Berton, O.; Sim-Selley, L.J.; Dileone, R.J.; et al. An essential role for ΔFosB in the nucleus accumbens in morphine action. Nat. Neurosci. 2006, 9, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Vialou, V.; Robison, A.J.; Laplant, Q.C.; Covington, H.E.; Dietz, D.M.; Ohnishi, Y.N.; Mouzon, E.; Rush, A.J.; Watts, E.L.; Wallace, D.L.; et al. ΔfosB in brain reward circuits mediates resilience to stress and antidepressant responses. Nat. Neurosci. 2010, 13, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.E.; Ferrario, C.R. AMPA receptor plasticity in the nucleus accumbens after repeated exposure to cocaine. Neurosci. Biobehav. Rev. 2010, 35, 185–211. [Google Scholar] [CrossRef]
- Russo, S.J.; Dietz, D.M.; Dumitriu, D.; Morrison, J.H.; Malenka, R.C.; Nestler, E.J. The addicted synapse: Mechanisms of synaptic and structural plasticity in nucleus accumbens. Trends Neurosci. 2010, 33, 267–276. [Google Scholar] [CrossRef]
- Kalivas, P.W. The glutamate homeostasis hypothesis of addiction. Nat. Rev. Neurosci. 2009, 10, 561–572. [Google Scholar] [CrossRef]
- Robison, A.J.; Vialou, V.; Mazei-Robison, M.; Feng, J.; Kourrich, S.; Collins, M.; Wee, S.; Koob, G.; Turecki, G.; Neve, R.; et al. Behavioral and structural responses to chronic cocaine require a feedforward loop involving Δ FosB and calcium/calmodulin-dependent protein kinase II in the nucleus accumbens shell. J. Neurosci. 2013, 33, 4295–4307. [Google Scholar] [CrossRef]
- Koob, G.F.; Volkow, N.D. Neurocircuitry of addiction. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2010, 35, 217–238. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.A.; Bozarth, M.A. A psychomotor stimulant theory of addiction. Psychol. Rev. 1987, 94, 469–492. [Google Scholar] [CrossRef] [PubMed]
- Di Chiara, G.; Imperato, A. Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc. Natl. Acad. Sci. USA 1988, 85, 5274–5278. [Google Scholar] [CrossRef]
- Koob, G.F.; Sanna, P.P.; Bloom, F.E. Neuroscience of addiction. Neuron 1998, 21, 467–476. [Google Scholar] [CrossRef]
- Nutt, D.J.; Lingford-Hughes, A.; Erritzoe, D.; Stokes, P.R.A. The dopamine theory of addiction: 40 years of highs and lows. Nat. Rev. Neurosci. 2015, 16, 305–312. [Google Scholar] [CrossRef]
- Uz, T.; Ahmed, R.; Akhisaroglu, M.; Kurtuncu, M.; Imbesi, M.; Dirim Arslan, A.; Manev, H. Effect of fluoxetine and cocaine on the expression of clock genes in the mouse hippocampus and striatum. Neuroscience 2005, 134, 1309–1316. [Google Scholar] [CrossRef]
- Cassel, S.; Carouge, D.; Gensburger, C.; Anglard, P.; Burgun, C.; Dietrich, J.B.; Aunis, D.; Zwiller, J. Fluoxetine and cocaine induce the epigenetic factors MeCP2 and MBD1 in adult rat brain. Mol. Pharmacol. 2006, 70, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Lanteri, C.; Salomon, L.; Torrens, Y.; Glowinski, J.; Tassin, J.-P. Drugs of abuse specifically sensitize noradrenergic and serotonergic neurons via a non-dopaminergic mechanism. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2008, 33, 1724–1734. [Google Scholar] [CrossRef]
- Peng, X.-Q.; Li, X.; Gilbert, J.G.; Pak, A.C.; Ashby, C.R.J.; Brodie, J.D.; Dewey, S.L.; Gardner, E.L.; Xi, Z.-X. Gamma-vinyl GABA inhibits cocaine-triggered reinstatement of drug-seeking behavior in rats by a non-dopaminergic mechanism. Drug Alcohol Depend. 2008, 97, 216–225. [Google Scholar] [CrossRef]
- De Backer, J.-F.; Monlezun, S.; Detraux, B.; Gazan, A.; Vanopdenbosch, L.; Cheron, J.; Cannazza, G.; Valverde, S.; Cantacorps, L.; Nassar, M.; et al. Deletion of Maged1 in mice abolishes locomotor and reinforcing effects of cocaine. EMBO Rep. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Badiani, A.; Belin, D.; Epstein, D.; Calu, D.; Shaham, Y. Opiate versus psychostimulant addiction: The differences do matter. Nat. Rev. Neurosci. 2011, 12, 685–700. [Google Scholar] [CrossRef]
- Hersey, M.; Bacon, A.K.; Bailey, L.G.; Coggiano, M.A.; Newman, A.H.; Leggio, L.; Tanda, G. Psychostimulant Use Disorder, an Unmet Therapeutic Goal: Can Modafinil Narrow the Gap? Front. Neurosci. 2021, 15, 656475. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, M.K. Anti-stress neuropharmacological mechanisms and targets for addiction treatment: A translational framework. Neurobiol. Stress 2018, 9, 84–104. [Google Scholar] [CrossRef] [PubMed]
- Martinotti, G.; Chiappini, S.; Pettorruso, M.; Mosca, A.; Miuli, A.; Di Carlo, F.; D’Andrea, G.; Collevecchio, R.; Di Muzio, I.; Sensi, S.L.; et al. Therapeutic Potentials of Ketamine and Esketamine in Obsessive-Compulsive Disorder (OCD), Substance Use Disorders (SUD) and Eating Disorders (ED): A Review of the Current Literature. Brain Sci. 2021, 11, 856. [Google Scholar] [CrossRef] [PubMed]
- Boutrel, B.; Steiner, N.; Halfon, O. The hypocretins and the reward function: What have we learned so far? Front. Behav. Neurosci. 2013, 7, 59. [Google Scholar] [CrossRef]
- James, M.H.; Mahler, S.V.; Moorman, D.E.; Aston-Jones, G. A Decade of Orexin/Hypocretin and Addiction: Where Are We Now? Curr. Top. Behav. Neurosci. 2017, 33, 247–281. [Google Scholar] [CrossRef] [PubMed]
- Gentile, T.A.; Simmons, S.J.; Barker, D.J.; Shaw, J.K.; España, R.A.; Muschamp, J.W. Suvorexant, an orexin/hypocretin receptor antagonist, attenuates motivational and hedonic properties of cocaine. Addict. Biol. 2018, 23, 247–255. [Google Scholar] [CrossRef]
- Levy, K.A.; Brodnik, Z.D.; Shaw, J.K.; Perrey, D.A.; Zhang, Y.; España, R.A. Hypocretin receptor 1 blockade produces bimodal modulation of cocaine-associated mesolimbic dopamine signaling. Psychopharmacology. 2017, 234, 2761–2776. [Google Scholar] [CrossRef]
- Keck, T.M.; Zou, M.-F.; Bi, G.-H.; Zhang, H.-Y.; Wang, X.-F.; Yang, H.-J.; Srivastava, R.; Gardner, E.L.; Xi, Z.-X.; Newman, A.H. A novel mGluR5 antagonist, MFZ 10-7, inhibits cocaine-taking and cocaine-seeking behavior in rats. Addict. Biol. 2014, 19, 195–209. [Google Scholar] [CrossRef]
- Zaniewska, M.; Przegaliński, E.; Filip, M.; Pilc, A.; Doller, D. Inhibitory actions of mGlu4 receptor ligands on cocaine−, but not nicotine−, induced sensitizing and conditioning locomotor responses in rats. Pharmacol. Rep. 2014, 66, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Gobin, C.; Schwendt, M. The cognitive cost of reducing relapse to cocaine-seeking with mGlu5 allosteric modulators. Psychopharmacology 2020, 237, 115–125. [Google Scholar] [CrossRef]
- Hajasova, Z.; Canestrelli, C.; Acher, F.; Noble, F.; Marie, N. Role of mGlu7 receptor in morphine rewarding effects is uncovered by a novel orthosteric agonist. Neuropharmacology 2018, 131, 424–430. [Google Scholar] [CrossRef] [PubMed]
- Sondheimer, I.; Knackstedt, L.A. Ceftriaxone prevents the induction of cocaine sensitization and produces enduring attenuation of cue- and cocaine-primed reinstatement of cocaine-seeking. Behav. Brain Res. 2011, 225, 252–258. [Google Scholar] [CrossRef]
- Schwendt, M.; Shallcross, J.; Hadad, N.A.; Namba, M.D.; Hiller, H.; Wu, L.; Krause, E.G.; Knackstedt, L.A. A novel rat model of comorbid PTSD and addiction reveals intersections between stress susceptibility and enhanced cocaine seeking with a role for mGlu5 receptors. Transl. Psychiatry 2018, 8, 209. [Google Scholar] [CrossRef] [PubMed]
- Noble, F.; Marie, N. Management of Opioid Addiction With Opioid Substitution Treatments: Beyond Methadone and Buprenorphine. Front. psychiatry 2018, 9, 742. [Google Scholar] [CrossRef]
- Tesone-Coelho, C.; Morel, L.J.; Bhatt, J.; Estevez, L.; Naudon, L.; Giros, B.; Zwiller, J.; Daugé, V. Vulnerability to opiate intake in maternally deprived rats: Implication of MeCP2 and of histone acetylation. Addict. Biol. 2015, 20, 120–131. [Google Scholar] [CrossRef]
- Baker-Andresen, D.; Zhao, Q.; Li, X.; Jupp, B.; Chesworth, R.; Lawrence, A.J.; Bredy, T. Persistent variations in neuronal DNA methylation following cocaine self-administration and protracted abstinence in mice. Neuroepigenetics 2015, 4, 1–11. [Google Scholar] [CrossRef]
- Ploense, K.L.; Li, X.; Baker-Andresen, D.; Carr, A.E.; Woodward, N.; Bagley, J.; Szumlinski, K.K.; Bredy, T.W.; Kippin, T.E. Prolonged-access to cocaine induces distinct Homer2 DNA methylation, hydroxymethylation, and transcriptional profiles in the dorsomedial prefrontal cortex of Male Sprague-Dawley rats. Neuropharmacology 2018, 143, 299–305. [Google Scholar] [CrossRef]
- Bibb, J.A.; Chen, J.; Taylor, J.R.; Svenningsson, P.; Nishi, A.; Snyder, G.L.; Yan, Z.; Sagawa, Z.K.; Ouimet, C.C.; Nairn, A.C.; et al. Effects of chronic exposure to cocaine are regulated by the neuronal protein Cdk5. Nature 2001, 410, 376–380. [Google Scholar] [CrossRef]
- McQuown, S.C.; Wood, M.A. Epigenetic regulation in substance use disorders. Curr. Psychiatry Rep. 2010, 12, 145–153. [Google Scholar] [CrossRef][Green Version]
- Kumar, A.; Choi, K.H.; Renthal, W.; Tsankova, N.M.; Theobald, D.E.H.; Truong, H.T.; Russo, S.J.; LaPlant, Q.; Sasaki, T.S.; Whistler, K.N.; et al. Chromatin remodeling is a key mechanism underlying cocaine-induced plasticity in striatum. Neuron 2005, 48, 303–314. [Google Scholar] [CrossRef]
- Renthal, W.; Kumar, A.; Xiao, G.; Wilkinson, M.; Covington, H.E.; Maze, I.; Sikder, D.; Robison, A.J.; LaPlant, Q.; Dietz, D.M.; et al. Genome-wide Analysis of Chromatin Regulation by Cocaine Reveals a Role for Sirtuins. Neuron 2009, 62, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Renthal, W.; Nestler, E.J. Histone acetylation in drug addiction. Semin. Cell Dev. Biol. 2009, 20, 387–394. [Google Scholar] [CrossRef]
- Pandey, S.C.; Ugale, R.; Zhang, H.; Tang, L.; Prakash, A. Brain chromatin remodeling: A novel mechanism of alcoholism. J. Neurosci. 2008, 28, 3729–3737. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Krishnan, H.R.; Ghezzi, A.; Yin, J.C.P.; Atkinson, N.S. Drug-induced epigenetic changes produce drug tolerance. PLoS Biol. 2007, 5, 2342–2353. [Google Scholar] [CrossRef] [PubMed]
- Khare, M.; Taylor, A.H.; Konje, J.C.; Bell, S.C. Δ9-Tetrahydrocannabinol inhibits cytotrophoblast cell proliferation and modulates gene transcription. Mol. Hum. Reprod. 2006, 12, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Olfson, E.; Bierut, L.J. Convergence of Genome-Wide Association and Candidate Gene Studies for Alcoholism. Alcohol. Clin. Exp. Res. 2012, 36, 2086–2094. [Google Scholar] [CrossRef] [PubMed]
- Renthal, W.; Maze, I.; Krishnan, V.; Covington, H.E.; Xiao, G.; Kumar, A.; Russo, S.J.; Graham, A.; Tsankova, N.; Kippin, T.E.; et al. Histone Deacetylase 5 Epigenetically Controls Behavioral Adaptations to Chronic Emotional Stimuli. Neuron 2007, 56, 517–529. [Google Scholar] [CrossRef]
- Romieu, P.; Host, L.; Gobaille, S.; Sandner, G.; Aunis, D.; Zwiller, J. Histone deacetylase inhibitors decrease cocaine but not sucrose self-administration in rats. J. Neurosci. 2008, 28, 9342–9348. [Google Scholar] [CrossRef]
- Kim, W.Y.; Kim, S.; Kim, J.H. Chronic microinjection of valproic acid into the nucleus accumbens attenuates amphetamine-induced locomotor activity. Neurosci. Lett. 2008, 432, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.; Shao, N.; Heller, E.; Feng, J.; Neve, R.; Kim, H.D.; Call, T.; Magazu, S.; Shen, L.; Nestler, E.J. SIRT1-FOXO3a regulate cocaine actions in the nucleus accumbens. J. Neurosci. 2015, 35, 3100–3111. [Google Scholar] [CrossRef]
- Sassone-Corsi, P. Minireview: NAD +, a circadian metabolite with an epigenetic twist. Endocrinology 2012, 153, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bellet, M.M.; Sassone-Corsi, P. Mammalian circadian clock and metabolism—The epigenetic link. J. Cell Sci. 2010, 123, 3837–3848. [Google Scholar] [CrossRef]
- Finkel, T.; Deng, C.X.; Mostoslavsky, R. Recent progress in the biology and physiology of sirtuins. Nature 2009, 460, 587–591. [Google Scholar] [CrossRef]
- Brunet, A.; Sweeney, L.B.; Sturgill, J.F.; Chua, K.F.; Greer, P.L.; Lin, Y.; Tran, H.; Ross, S.E.; Mostoslavsy, R.; Cohen, H.Y.; et al. Stress-Dependent Regulation of FOXO Transcription Factors by the SIRT1 Deacetylase. Science 2004, 303, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Herskovits, A.Z.; Guarente, L. SIRT1 in Neurodevelopment and Brain Senescence. Neuron 2014, 81, 471–483. [Google Scholar] [CrossRef]
- Wang, F.; Tong, Q. SIRT2 suppresses adipocyte differentiation by deacetylating FOXO1 and enhancing FOXO1’s repressive interaction with PPARγ. Mol. Biol. Cell 2009, 20, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Renthal, W.; Carle, T.L.; Maze, I.; Covington, H.E.; Truong, H.T.; Alibhai, I.; Kumar, A.; Montgomery, R.L.; Olson, E.N.; Nestler, E.J. ΔFosB mediates epigenetic desensitization of the c-fos gene after chronic amphetamine exposure. J. Neurosci. 2008, 28, 7344–7349. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.; Koo, J.W.; Feng, J.; Heller, E.; Rabkin, J.; Heshmati, M.; Renthal, W.; Neve, R.; Liu, X.; Shao, N.; et al. Essential role of SIRT1 signaling in the nucleus accumbens in cocaine and morphine action. J. Neurosci. 2013, 33, 16088–16098. [Google Scholar] [CrossRef]
- Maze, I.; Nestler, E.J. The epigenetic landscape of addiction. Ann. N. Y. Acad. Sci. 2011, 1216, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Maze, I.; Covingtoni, H.E.; Dietz, D.M.; Laplant, Q.; Renthal, W.; Russo, S.J.; Mechanic, M.; Mouzon, E.; Neve, R.L.; Haggarty, S.J.; et al. Essential role of the histone methyltransferase G9a in cocaine-induced plasticity. Science 2010, 327, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Russo, S.J.; Wilkinson, M.B.; Mazei-Robison, M.S.; Dietz, D.M.; Maze, I.; Krishnan, V.; Renthal, W.; Graham, A.; Birnbaum, S.G.; Green, T.A.; et al. Nuclear factor kB signaling regulates neuronal morphology and cocaine reward. J. Neurosci. 2009, 29, 3529–3537. [Google Scholar] [CrossRef] [PubMed]
- Norrholm, S.D.; Bibb, J.A.; Nestler, E.J.; Ouimet, C.C.; Taylor, J.R.; Greengard, P. Cocaine-induced proliferation of dendritic spines in nucleus accumbens is dependent on the activity of cyclin-dependent kinase-5. Neuroscience 2003, 116, 19–22. [Google Scholar] [CrossRef]
- Pulipparacharuvil, S.; Renthal, W.; Hale, C.F.; Taniguchi, M.; Xiao, G.; Kumar, A.; Russo, S.J.; Sikder, D.; Dewey, C.M.; Davis, M.M.; et al. Cocaine Regulates MEF2 to Control Synaptic and Behavioral Plasticity. Neuron 2008, 59, 621–633. [Google Scholar] [CrossRef]
- Brenna, A.; Olejniczak, I.; Chavan, R.; Ripperger, J.A.; Langmesser, S.; Cameroni, E.; Hu, Z.; De Virgilio, C.; Dengjel, J.; Albrecht, U. Cyclin-dependent kinase 5 (CDK5) regulates the circadian clock. Elife 2019, 8. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Akumuo, R.C.; España, R.A.; Yan, C.X.; Gao, W.J.; Li, Y.C. The histone demethylase KDM6B in the medial prefrontal cortex epigenetically regulates cocaine reward memory. Neuropharmacology 2018, 141, 113–125. [Google Scholar] [CrossRef]
- Johnstone, A.L.; Andrade, N.S.; Barbier, E.; Khomtchouk, B.B.; Rienas, C.A.; Lowe, K.; Van Booven, D.J.; Domi, E.; Esanov, R.; Vilca, S.; et al. Dysregulation of the histone demethylase KDM6B in alcohol dependence is associated with epigenetic regulation of inflammatory signaling pathways. Addict. Biol. 2019, e12816. [Google Scholar] [CrossRef]
- Walker, D.M.; Cates, H.M.; Heller, E.A.; Nestler, E.J. Regulation of chromatin states by drugs of abuse. Curr. Opin. Neurobiol. 2015, 30, 112–121. [Google Scholar] [CrossRef]
- Aguilar-Valles, A.; Vaissière, T.; Griggs, E.M.; Mikaelsson, M.A.; Takács, I.F.; Young, E.J.; Rumbaugh, G.; Miller, C.A. Methamphetamine-associated memory is regulated by a writer and an eraser of permissive histone methylation. Biol. Psychiatry 2014, 76, 57–65. [Google Scholar] [CrossRef]
- Pisera-Fuster, A.; Zwiller, J.; Bernabeu, R. Methionine Supplementation Abolishes Nicotine-Induced Place Preference in Zebrafish: A Behavioral and Molecular Analysis. Mol. Neurobiol. 2021. [Google Scholar] [CrossRef]
- Carouge, D.; Host, L.; Aunis, D.; Zwiller, J.; Anglard, P. CDKL5 is a brain MeCP2 target gene regulated by DNA methylation. Neurobiol. Dis. 2010, 38, 414–424. [Google Scholar] [CrossRef]
- Neul, J.L. The relationship of Rett syndrome and MECP2 disorders to autism. Dialogues Clin. Neurosci. 2012, 14, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Anier, K.; Malinovskaja, K.; Aonurm-Helm, A.; Zharkovsky, A.; Kalda, A. DNA methylation regulates cocaine-induced behavioral sensitization in mice. Neuropsychopharmacology 2010, 35, 2450–2461. [Google Scholar] [CrossRef]
- Duman, R.S.; Newton, S.S. Epigenetic marking and neuronal plasticity. Biol. Psychiatry 2007, 62, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Im, H.I.; Hollander, J.A.; Bali, P.; Kenny, P.J. MeCP2 controls BDNF expression and cocaine intake through homeostatic interactions with microRNA-212. Nat. Neurosci. 2010, 13, 1120–1127. [Google Scholar] [CrossRef]
- Lyst, M.J.; Ekiert, R.; Ebert, D.H.; Merusi, C.; Nowak, J.; Selfridge, J.; Guy, J.; Kastan, N.R.; Robinson, N.D.; De Lima Alves, F.; et al. Rett syndrome mutations abolish the interaction of MeCP2 with the NCoR/SMRT co-repressor. Nat. Neurosci. 2013, 16, 898–902. [Google Scholar] [CrossRef] [PubMed]
- Host, L.; Dietrich, J.B.; Carouge, D.; Aunis, D.; Zwiller, J. Cocaine self-administration alters the expression of chromatin-remodelling proteins; Modulation by histone deacetylase inhibition. J. Psychopharmacol. 2011, 25, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Shiota, K. Methyl-CpG-binding protein, MeCP2, is a target molecule for maintenance DNA methyltransferase, Dnmt1. J. Biol. Chem. 2003, 278, 4806–4812. [Google Scholar] [CrossRef] [PubMed]
- Mari, F.; Azimonti, S.; Bertani, I.; Bolognese, F.; Colombo, E.; Caselli, R.; Scala, E.; Longo, I.; Grosso, S.; Pescucci, C.; et al. CDKL5 belongs to the same molecular pathway of MeCP2 and it is responsible for the early-onset seizure variant of Rett syndrome. Hum. Mol. Genet. 2005, 14, 1935–1946. [Google Scholar] [CrossRef]
- Moretti, P.; Levenson, J.M.; Battaglia, F.; Atkinson, R.; Teague, R.; Antalffy, B.; Armstrong, D.; Arancio, O.; Sweatt, J.D.; Zoghbi, H.Y. Learning and memory and synaptic plasticity are impaired in a mouse model of Rett syndrome. J. Neurosci. 2006, 26, 319–327. [Google Scholar] [CrossRef]
- Genoux, D.; Haditsch, U.; Knobloch, M.; Michalon, A.; Storm, D.; Mansuy, I.M. Protein phosphatase 1 is a molecular constraint on learning and memory. Nature 2002, 418, 970–975. [Google Scholar] [CrossRef]
- Miller, C.A.; Sweatt, J.D. Covalent modification of DNA regulates memory formation. Neuron 2007, 53, 857–869. [Google Scholar] [CrossRef]
- Bodetto, S.P.; Carouge, D.; Fonteneau, M.; Dietrich, J.B.; Zwiller, J.; Anglard, P. Cocaine represses protein phosphatase-1Cβ through DNA methylation and Methyl-CpG Binding Protein-2 recruitment in adult rat brain. Neuropharmacology 2013, 73, 31–40. [Google Scholar] [CrossRef]
- Pol Bodetto, S.; Romieu, P.; Sartori, M.; Tesone-Coelho, C.; Majchrzak, M.; Barbelivien, A.; Zwiller, J.; Anglard, P. Differential regulation of MeCP2 and PP1 in passive or voluntary administration of cocaine or food. Int. J. Neuropsychopharmacol. 2014, 17, 2031–2044. [Google Scholar] [CrossRef]
- Dong, Y.; Nestler, E.J. The neural rejuvenation hypothesis of cocaine addiction. Trends Pharmacol. Sci. 2014, 35, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Shao, N.; Szulwach, K.E.; Vialou, V.; Huynh, J.; Zhong, C.; Le, T.; Ferguson, D.; Cahill, M.E.; Li, Y.; et al. Role of Tet1 and 5-hydroxymethylcytosine in cocaine action. Nat. Neurosci. 2015, 18, 536–544. [Google Scholar] [CrossRef] [PubMed]
- Hopf, F.W.; Bonci, A. Dnmt3a: Addiction’s molecular forget-me-not? Nat. Neurosci. 2010, 13, 1041–1043. [Google Scholar] [CrossRef]
- Laplant, Q.; Vialou, V.; Covington, H.E.; Dumitriu, D.; Feng, J.; Warren, B.L.; Maze, I.; Dietz, D.M.; Watts, E.L.; Iñiguez, S.D.; et al. Dnmt3a regulates emotional behavior and spine plasticity in the nucleus accumbens. Nat. Neurosci. 2010, 13, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- Saad, L.; Sartori, M.; Pol Bodetto, S.; Romieu, P.; Kalsbeek, A.; Zwiller, J.; Anglard, P. Regulation of Brain DNA Methylation Factors and of the Orexinergic System by Cocaine and Food Self-Administration. Mol. Neurobiol. 2019, 56, 5315–5331. [Google Scholar] [CrossRef]
- Saad, L.; Kalsbeek, A.; Zwiller, J.; Anglard, P. Rhythmic regulation of DNA methylation factors and core-clock genes in brain structures activated by cocaine or sucrose. Genes 2021, 12, 1195. [Google Scholar] [CrossRef]
- Tian, W.; Zhao, M.; Li, M.; Song, T.; Zhang, M.; Quan, L.; Li, S.; Sun, Z.S. Reversal of cocaine-conditioned place preference through methyl supplementation in mice: Altering global DNA methylation in the prefrontal cortex. PLoS ONE 2012, 7, e33435. [Google Scholar] [CrossRef]
- Fragou, D.; Zanos, P.; Kouidou, S.; Njau, S.; Kitchen, I.; Bailey, A.; Kovatsi, L. Effect of chronic heroin and cocaine administration on global DNA methylation in brain and liver. Toxicol. Lett. 2013, 218, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.N.; Hollis, F.; Duclot, F.; Dossat, A.M.; Strong, C.E.; Francis, T.C.; Mercer, R.; Feng, J.; Dietz, D.M.; Lobo, M.K.; et al. Methyl supplementation attenuates cocaine-seeking behaviors and cocaine-induced c-Fos activation in a DNA methylation-dependent manner. J. Neurosci. 2015, 35, 8948–8958. [Google Scholar] [CrossRef] [PubMed]
- Massart, R.; Barnea, R.; Dikshtein, Y.; Suderman, M.; Meir, O.; Hallett, M.; Kennedy, P.; Nestler, E.J.; Szyf, M.; Yadid, G. Role of DNA methylation in the nucleus accumbens in incubation of cocaine craving. J. Neurosci. 2015, 35, 8042–8058. [Google Scholar] [CrossRef] [PubMed]
- Fonteneau, M.; Filliol, D.; Anglard, P.; Befort, K.; Romieu, P.; Zwiller, J. Inhibition of DNA methyltransferases regulates cocaine self-administration by rats: A genome-wide DNA methylation study. Genes Brain Behav. 2017, 16, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.V.; Rodriguiz, R.M.; Hutchinson, A.N.; Kim, I.H.; Wetsel, W.C.; West, A.E. MeCP2 in the nucleus accumbens contributes to neural and behavioral responses to psychostimulants. Nat. Neurosci. 2010, 13, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- Kelz, M.B.; Nestler, E.J. ΔFosB: A molecular switch underlying long-term neural plasticity. Curr. Opin. Neurol. 2000, 13, 715–720. [Google Scholar] [CrossRef]
- Peters, R.V.; Aronin, N.; Schwartz, W.J. Circadian regulation of Fos B is different from c-Fos in the rat suprachiasmatic nucleus. Mol. Brain Res. 1994, 27, 243–248. [Google Scholar] [CrossRef]
- Schwartz, W.J.; Carpino, A.; de la Iglesia, H.O.; Baler, R.; Klein, D.C.; Nakabeppu, Y.; Aronin, N. Differential regulation of fos family genes in the ventrolateral and dorsomedial subdivisions of the rat suprachiasmatic nucleus. Neuroscience 2000, 98, 535–547. [Google Scholar] [CrossRef]
- Li, M.D.; Van Der Vaart, A.D. MicroRNAs in addiction: Adaptation’s middlemen. Mol. Psychiatry 2011, 16, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.J.; Zhang, C.; Zhong, Y.; Luo, J.; Zhang, C.Y.; Zhang, C.; Wang, C. Altered serum microRNA expression profile in subjects with heroin and methamphetamine use disorder. Biomed. Pharmacother. 2020, 125, 109918. [Google Scholar] [CrossRef] [PubMed]
- Du, H.Y.; Cao, D.N.; Chen, Y.; Wang, L.; Wu, N.; Li, J. Alterations of prefrontal cortical microRNAs in methamphetamine self-administering rats: From controlled drug intake to escalated drug intake. Neurosci. Lett. 2016, 611, 21–27. [Google Scholar] [CrossRef]
- Michelhaugh, S.K.; Lipovich, L.; Blythe, J.; Jia, H.; Kapatos, G.; Bannon, M.J. Mining Affymetrix microarray data for long non-coding RNAs: Altered expression in the nucleus accumbens of heroin abusers. J. Neurochem. 2011, 116, 459–466. [Google Scholar] [CrossRef]
- Young, J.I.; Hong, E.P.; Castle, J.C.; Crespo-Barreto, J.; Bowman, A.B.; Rose, M.F.; Kang, D.; Richman, R.; Johnson, J.M.; Berget, S.; et al. Regulation of RNA splicing by the methylation-dependent transcriptional repressor methyl-CpG binding protein 2. Proc. Natl. Acad. Sci. USA 2005, 102, 17551–17558. [Google Scholar] [CrossRef]
- Cheng, T.-L.; Wang, Z.; Liao, Q.; Zhu, Y.; Zhou, W.-H.; Xu, W.; Qiu, Z. MeCP2 suppresses nuclear microRNA processing and dendritic growth by regulating the DGCR8/Drosha complex. Dev. Cell 2014, 28, 547–560. [Google Scholar] [CrossRef]
- Chandrasekar, V.; Dreyer, J.L. microRNAs miR-124, let-7d and miR-181a regulate Cocaine-induced Plasticity. Mol. Cell. Neurosci. 2009, 42, 350–362. [Google Scholar] [CrossRef]
- Chandrasekar, V.; Dreyer, J.L. Regulation of MiR-124, Let-7d, and MiR-181a in the accumbens affects the expression, extinction, and reinstatement of cocaine-induced conditioned place preference. Neuropsychopharmacology 2011, 36, 1149–1164. [Google Scholar] [CrossRef]
- Sanchez-Simon, F.M.; Zhang, X.X.; Loh, H.H.; Law, P.Y.; Rodriguez, R.E. Morphine regulates dopaminergic neuron differentiation via miR-133b. Mol. Pharmacol. 2010, 78, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zeng, Y.; Zhang, X.; Chu, J.; Loh, H.H.; Law, P.Y. μ-opioid receptor agonists differentially regulate the expression of miR-190 and NeuroD. Mol. Pharmacol. 2010, 77, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Pietrzykowski, A.Z. The Role of microRNAs in Drug Addiction. A Big Lesson from Tiny Molecules. Int. Rev. Neurobiol. 2010, 91, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Hammer, S.B.; Ruby, C.L.; Brager, A.J.; Prosser, R.A.; Glass, J.D. Environmental modulation of alcohol intake in hamsters: Effects of wheel running and constant light exposure. Alcohol. Clin. Exp. Res. 2010, 34, 1651–1658. [Google Scholar] [CrossRef] [PubMed]
- Kosobud, A.E.K.; Gillman, A.G.; Leffel, J.K.; Pecoraro, N.C.; Rebec, G.V.; Timberlake, W. Drugs of abuse can entrain circadian rhythms. Sci. World J. 2007, 7, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Logan, R.W.; Williams, W.P., 3rd; McClung, C.A. Circadian rhythms and addiction: Mechanistic insights and future directions. Behav. Neurosci. 2014, 128, 387–412. [Google Scholar] [CrossRef] [PubMed]
- Partonen, T. Clock genes in human alcohol abuse and comorbid conditions. Alcohol 2015, 49, 359–365. [Google Scholar] [CrossRef]
- Gamble, K.L.; Motsinger-Reif, A.A.; Hida, A.; Borsetti, H.M.; Servick, S.V.; Ciarleglio, C.M.; Robbins, S.; Hicks, J.; Carver, K.; Hamilton, N.; et al. Shift work in nurses: Contribution of phenotypes and genotypes to adaptation. PLoS ONE 2011, 6, e18395. [Google Scholar] [CrossRef]
- Adan, A.; Archer, S.N.; Hidalgo, M.P.; Di Milia, L.; Natale, V.; Randler, C. Circadian typology: A comprehensive review. Chronobiol. Int. 2012, 29, 1153–1175. [Google Scholar] [CrossRef]
- Thakkar, M.M.; Sharma, R.; Sahota, P. Alcohol disrupts sleep homeostasis. Alcohol 2015, 49, 299–310. [Google Scholar] [CrossRef]
- Modulation, E.; Alcohol, O.F.; In, I.; Exposure, L. Hamsters: Effects of Wheel-Running and Constant. Alcohol Clin. Exp. Res. 2011, 34, 1651–1658. [Google Scholar] [CrossRef]
- Parekh, P.K.; Ozburn, A.R.; McClung, C.A. Circadian clock genes: Effects on dopamine, reward and addiction. Alcohol 2015, 49, 341–349. [Google Scholar] [CrossRef] [PubMed]
- McClung, C.A.; Sidiropoulou, K.; Vitaterna, M.; Takahashi, J.S.; White, F.J.; Cooper, D.C.; Nestler, E.J. Regulation of dopaminergic transmission and cocaine reward by the Clock gene. Proc. Natl. Acad. Sci. USA 2005, 102, 9377–9381. [Google Scholar] [CrossRef]
- Falcón, E.; McClung, C.A. A role for the circadian genes in drug addiction. Neuropharmacology 2009, 56, 91–96. [Google Scholar] [CrossRef]
- Grandin, L.D.; Alloy, L.B.; Abramson, L.Y. The social zeitgeber theory, circadian rhythms, and mood disorders: Review and evaluation. Clin. Psychol. Rev. 2006, 26, 679–694. [Google Scholar] [CrossRef] [PubMed]
- McClung, C.A. Circadian Genes, Rhythms and the Biology of Mood Disorders. Pharmacol. Ther. 2007, 114, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Lynch, W.J.; Girgenti, M.J.; Breslin, F.J.; Newton, S.S.; Taylor, J.R. Gene profiling the response to repeated cocaine self-administration in dorsal striatum: A focus on circadian genes. Brain Res. 2008, 1213, 166–177. [Google Scholar] [CrossRef]
- Roberts, D.C.S.; Brebner, K.; Vincler, M.; Lynch, W.J. Patterns of cocaine self-administration in rats produced by various access conditions under a discrete trials procedure. Drug Alcohol Depend. 2002, 67, 291–299. [Google Scholar] [CrossRef]
- Abarca, C.; Albrecht, U.; Spanagel, R. Cocaine sensitization and reward are under the influence of circadian genes and rhythm. Proc. Natl. Acad. Sci. USA 2002, 99, 9026–9030. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Wan, C.; Zhou, W.; Peng, T.; Liu, Y.; Wang, Z.; Li, G.; Cornelisson, G.; Halberg, F. The role of mPer1 in morphine dependence in mice. Neuroscience 2005, 130, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Linde, K.V.D.; Lyons, L.C. Circadian modulation of acute alcohol sensitivity but not acute tolerance in drosophila. Chronobiol. Int. 2011, 28, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Andretic, R.; Chaney, S.; Hirsh, J. Requirement of circadian genes for cocaine sensitization in Drosophila. Science 1999, 285, 1066–1068. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.A.; Zhang, D.; Estill, S.J.; Michnoff, C.; Rutter, J.; Reick, M.; Scott, K.; Diaz-Arrastia, R.; McKnight, S.L. Impaired cued and contextual memory in NPAS2-deficient mice. Science 2000, 288, 2226–2230. [Google Scholar] [CrossRef]
- Dudley, C.A.; Erbel-Sieler, C.; Estill, S.J.; Reick, M.; Franken, P.; Pitts, S.N.; McKnight, S.L. Altered patterns of sleep and behavioral adaptability in NPAS2-deficient mice. Science 2003, 301, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Franken, P.; Dudley, C.A.; Estill, S.J.; Barakat, M.; Thomason, R.; O’Hara, B.F.; McKnight, S.L. NPAS2 as a transcriptional regulator of non-rapid eye movement sleep: Genotype and sex interactions. Proc. Natl. Acad. Sci. USA 2006, 103, 7118–7123. [Google Scholar] [CrossRef] [PubMed]
- Hampp, G.; Ripperger, J.A.; Houben, T.; Schmutz, I.; Blex, C.; Perreau-Lenz, S.; Brunk, I.; Spanagel, R.; Ahnert-Hilger, G.; Meijer, J.H.; et al. Regulation of Monoamine Oxidase A by Circadian-Clock Components Implies Clock Influence on Mood. Curr. Biol. 2008, 18, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wang, S.; Zhou, Z.; Guo, L.; Yu, F.; Wu, B. Bmal1 regulates circadian expression of cytochrome P450 3a11 and drug metabolism in mice. Commun. Biol. 2019, 2, 378. [Google Scholar] [CrossRef] [PubMed]
- Perreau-Lenz, S.; Zghoul, T.; De Fonseca, F.R.; Spanagel, R.; Bilbao, A. Circadian regulation of central ethanol sensitivity by the mPer2 gene. Addict. Biol. 2009, 14, 253–259. [Google Scholar] [CrossRef]
- Iijima, M.; Nikaido, T.; Akiyama, M.; Moriya, T.; Shibata, S. Methamphetamine-induced, suprachiasmatic nucleus-independent circadian rhythms of activity and mPer gene expression in the striatum of the mouse. Eur. J. Neurosci. 2002, 16, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Jiang, Z.; Wan, C.; Zhou, W.; Wang, Z. The extracellular signal-regulated kinase signaling pathway is involved in the modulation of morphine-induced reward by mPer1. Neuroscience 2007, 146, 265–271. [Google Scholar] [CrossRef]
- Spanagel, R.; Pendyala, G.; Abarca, C.; Zghoul, T.; Sanchis-Segura, C.; Magnone, M.C.; Lascorz, J.; Depner, M.; Holzberg, D.; Soyka, M.; et al. The clock gene Per2 influences the glutamatergic system and modulates alcohol consumption. Nat. Med. 2005, 11, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Brager, A.J.; Prosser, R.A.; Glass, J.D. Circadian and acamprosate modulation of elevated ethanol drinking in mPer2 clock gene mutant mice. Chronobiol. Int. 2011, 28, 664–672. [Google Scholar] [CrossRef][Green Version]
- McClung, C.A. Circadian rhythms, the mesolimbic dopaminergic circuit, and drug addiction. Sci. World J. 2007, 7, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Roybal, K.; Theobold, D.; Graham, A.; DiNieri, J.A.; Russo, S.J.; Krishnan, V.; Chakravarty, S.; Peevey, J.; Oehrlein, N.; Birnbaum, S.; et al. Mania-like behavior induced by disruption of CLOCK. Proc. Natl. Acad. Sci. USA 2007, 104, 6406–6411. [Google Scholar] [CrossRef]
- Sleipness, E.P.; Sorg, B.A.; Jansen, H.T. Diurnal differences in dopamine transporter and tyrosine hydroxylase levels in rat brain: Dependence on the suprachiasmatic nucleus. Brain Res. 2007, 1129, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Lauterburg, T.; Tobler, I.; Burgunder, J.M. Circadian patterns of neurotransmitter related gene expression in motor regions of the rat brain. Neurosci. Lett. 2004, 358, 17–20. [Google Scholar] [CrossRef]
- Sleipness, E.P.; Sorg, B.A.; Jansen, H.T. Contribution of the suprachiasmatic nucleus to day:night variation in cocaine-seeking behavior. Physiol. Behav. 2007, 91, 523–530. [Google Scholar] [CrossRef]
- Beaulé, C.; Swanstrom, A.; Leone, M.J.; Herzog, E.D. Circadian modulation of gene expression, but not glutamate uptake, in mouse and rat cortical astrocytes. PLoS ONE 2009, 4, e7476. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, T.R.; de Prado, B.M.; Prieto, D.; Mora, F. Circadian rhythms of dopamine, glutamate and GABA in the striatum and nucleus accumbens of the awake rat: Modulation by light. J. Pineal Res. 2004, 36, 177–185. [Google Scholar] [CrossRef]
- De Marquez Prado, B.; Castañeda, T.R.; Galindo, A.; Del Arco, A.; Segovia, G.; Reiter, R.J.; Mora, F. Melatonin disrupts circadian rhythms of glutamate and GABA in the neostriatum of the awake rat: A microdialysis study. J. Pineal Res. 2000, 29, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Hnasko, T.S.; Chuhma, N.; Zhang, H.; Goh, G.Y.; Sulzer, D.; Palmiter, R.D.; Rayport, S.; Edwards, R.H. Vesicular Glutamate Transport Promotes Dopamine Storage and Glutamate Corelease In Vivo. Neuron 2010, 65, 643–656. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.M.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef]
- Nikaido, T.; Akiyama, M.; Moriya, T.; Shibata, S. Sensitized increase of period gene expression in the mouse caudate/putamen caused by repeated injection of methamphetamine. Mol. Pharmacol. 2001, 59, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Amir, S.; Harbour, V.L.; Robinson, B. Pinealectomy does not affect diurnal PER2 expression in the rat limbic forebrain. Neurosci. Lett. 2006, 399, 147–150. [Google Scholar] [CrossRef]
- Imbesi, M.; Uz, T.; Yildiz, S.; Arslan, A.D.; Manev, H. Drug- and region-specific effects of protracted antidepressant and cocaine treatment on the content of melatonin MT1 and MT2 receptor mRNA in the mouse brain. Int. J. Neuroprot. Neuroregener. 2006, 2, 185–189. [Google Scholar] [PubMed]
- Robinson, S.L.; Thiele, T.E. The Role of Neuropeptide Y (NPY) in Alcohol and Drug Abuse Disorders. Int. Rev. Neurobiol. 2017, 136, 177–197. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Li, H.; Wu, Z.; Yan, J.; Liu, Q.; Ou, C.; Chen, M. A Promising Therapeutic Target for Metabolic Diseases: Neuropeptide Y Receptors in Humans. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 45, 88–107. [Google Scholar] [CrossRef] [PubMed]
- Feillet, C.A.; Bainier, C.; Mateo, M.; Blancas-Velázquez, A.; Salaberry, N.L.; Ripperger, J.A.; Albrecht, U.; Mendoza, J. Rev-erbα modulates the hypothalamic orexinergic system to influence pleasurable feeding behaviour in mice. Addict. Biol. 2017, 22, 411–422. [Google Scholar] [CrossRef]
- Bayerlein, K.; Kraus, T.; Leinonen, I.; Pilniok, D.; Rotter, A.; Hofner, B.; Schwitulla, J.; Sperling, W.; Kornhuber, J.; Biermann, T. Orexin A expression and promoter methylation in patients with alcohol dependence comparing acute and protracted withdrawal. Alcohol 2011, 45, 541–547. [Google Scholar] [CrossRef]
- Rotter, A.; Bayerlein, K.; Hansbauer, M.; Weiland, J.; Sperling, W.; Kornhuber, J.; Biermann, T. Orexin A expression and promoter methylation in patients with cannabis dependence in comparison to nicotine-dependent cigarette smokers and nonsmokers. Neuropsychobiology 2012, 66, 126–133. [Google Scholar] [CrossRef]
- Dehan, P.; Canon, C.; Trooskens, G.; Rehli, M.; Munaut, C.; Van Criekinge, W.; Delvenne, P. Expression of type 2 orexin receptor in human endometrium and its epigenetic silencing in endometrial cancer. J. Clin. Endocrinol. Metab. 2013, 98, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Hirosawa, M.; Tabei, Y.; Arai, D.; Tanaka, S.; Murakami, N.; Yagi, S.; Shiota, K. Epigenetic switching by the metabolism-sensing factors in the generation of orexin neurons from mouse embryonic stem cells. J. Biol. Chem. 2013, 288, 17099–17110. [Google Scholar] [CrossRef]
- Wiskerke, J.; James, M.H.; Aston-Jones, G. The orexin-1 receptor antagonist SB-334867 reduces motivation, but not inhibitory control, in a rat stop signal task. Brain Res. 2020, 1731, 146222. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, G.; Wörtwein, G.; Fink-Jensen, A.; Woldbye, D.P.D. Neuropeptide Y Y5 receptor antagonism causes faster extinction and attenuates reinstatement in cocaine-induced place preference. Pharmacol. Biochem. Behav. 2013, 105, 151–156. [Google Scholar] [CrossRef]
- Manev, H.; Uz, T. Clock genes: Influencing and being influenced by psychoactive drugs. Trends Pharmacol. Sci. 2006, 27, 186–189. [Google Scholar] [CrossRef]
- Masubuchi, S.; Honma, S.; Abe, H.; Ishizaki, K.; Namihira, M.; Ikeda, M.; Honma, K.I. Clock genes outside the suprachiasmatic nucleus involved in manifestation of locomotor activity rhythm in rats. Eur. J. Neurosci. 2000, 12, 4206–4214. [Google Scholar] [CrossRef]
- Schade, R.; Vick, K.; Ott, T.; Sohr, R.; Pfister, C.; Bellach, J.; Golor, G.; Lemmer, B. Circadian rhythms of dopamine and cholecystokinin in nucleus accumbens and striatum of rats—Influence on dopaminergic stimulation. Chronobiol. Int. 1995, 12, 87–99. [Google Scholar] [CrossRef]
- Sellix, M.T.; Egli, M.; Poletini, M.O.; McKee, D.T.; Bosworth, M.D.; Fitch, C.A.; Freeman, M.E. Anatomical and functional characterization of clock gene expression in neuroendocrine dopaminergic neurons. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R1309–R1323. [Google Scholar] [CrossRef]
- Yelamanchili, S.V.; Pendyala, G.; Brunk, I.; Darna, M.; Albrecht, U.; Ahnert-Hilger, G. Differential sorting of the vesicular glutamate transporter 1 into a defined vesicular pool is regulated by light signaling involving the clock gene Period2. J. Biol. Chem. 2006, 281, 15671–15679. [Google Scholar] [CrossRef]
- Dykens, E.; Shah, B. Psychiatric disorders in Prader-Willi syndrome: Epidemiology and management. CNS Drugs 2003, 17, 167–178. [Google Scholar] [CrossRef]
- Sahoo, T.; Del Gaudio, D.; German, J.R.; Shinawi, M.; Peters, S.U.; Person, R.E.; Garnica, A.; Cheung, S.W.; Beaudet, A.L. Prader-Willi phenotype caused by paternal deficiency for the HBII-85 C/D box small nucleolar RNA cluster. Nat. Genet. 2008, 40, 719–721. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.T.; Coulson, R.L.; Crary, F.K.; Wong, S.S.; Ach, R.A.; Tsang, P.; Yamada, N.A.; Yasui, D.H.; LaSalle, J.M. A Prader-Willi locus lncRNA cloud modulates diurnal genes and energy expenditure. Hum. Mol. Genet. 2013, 22, 4318–4328. [Google Scholar] [CrossRef] [PubMed]
- Saus, E.; Soria, V.; Escaramís, G.; Vivarelli, F.; Crespo, J.M.; Kagerbauer, B.; Menchòn, J.M.; Urretavizcaya, M.; Gratacòs, M.; Estivill, X. Genetic variants and abnormal processing of pre-miR-182, a circadian clock modulator, in major depression patients with late insomnia. Hum. Mol. Genet. 2010, 19, 4017–4025. [Google Scholar] [CrossRef]
- Rumbaugh, G.; Miller, C.A. Epigenetic changes in the brain: Measuring global histone modifications. Methods Mol. Biol. 2011, 670, 263–274. [Google Scholar] [CrossRef]
- Martínez de Paz, A.; Sanchez-Mut, J.V.; Samitier-Martí, M.; Petazzi, P.; Sáez, M.; Szczesna, K.; Huertas, D.; Esteller, M.; Ausió, J. Circadian cycle-dependent MeCP2 and brain chromatin changes. PLoS ONE 2015, 10, e0123693. [Google Scholar] [CrossRef]
- Schmutz, I.; Wendt, S.; Schnell, A.; Kramer, A.; Mansuy, I.M.; Albrecht, U. Protein phosphatase 1 (PP1) is a post-translational regulator of the mammalian circadian clock. PLoS ONE 2011, 6, e21325. [Google Scholar] [CrossRef] [PubMed]
- Gallego, M.; Kang, H.; Virshup, D.M. Protein phosphatase 1 regulates the stability of the circadian protein PER2. Biochem. J. 2006, 399, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Sathyanarayanan, S.; Sehgal, A. Post-translational regulation of the Drosophila circadian clock requires protein phosphatase 1 (PP1). Genes Dev. 2007, 21, 1506–1518. [Google Scholar] [CrossRef] [PubMed]
- Garbe, D.S.; Fang, Y.; Zheng, X.; Sowcik, M.; Anjum, R.; Gygi, S.P.; Sehgal, A. Cooperative interaction between phosphorylation sites on PERIOD maintains circadian period in Drosophila. PLoS Genet. 2013, 9, e1003749. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.G.; Chan, D.W.; Reynolds, A.B.; Qin, J.; Wong, J. N-CoR mediates DNA methylation-dependent repression through a methyl CpG binding protein Kaiso. Mol. Cell 2003, 12, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Lazar, M.A. The orphan nuclear receptor Rev-erbα recruits the N-CoR/histone deacetylase 3 corepressor to regulate the circadian Bmal1 gene. Mol. Endocrinol. 2005, 19, 1452–1459. [Google Scholar] [CrossRef]
- Mottis, A.; Mouchiroud, L.; Auwerx, J. Emerging roles of the corepressors NCoR1 and SMRT in homeostasis. Genes Dev. 2013, 27, 819–835. [Google Scholar] [CrossRef]
- Schroeder, F.A.; Penta, K.L.; Matevossian, A.; Jones, S.R.; Konradi, C.; Tapper, A.R.; Akbarian, S. Drug-induced activation of dopamine D1 receptor signaling and inhibition of class I/II histone deacetylase induce chromatin remodeling in reward circuitry and modulate cocaine-related behaviors. Neuropsychopharmacology 2008, 33, 2981–2992. [Google Scholar] [CrossRef] [PubMed]
- Bertran-Gonzalez, J.; Bosch, C.; Maroteaux, M.; Matamales, M.; Hervé, D.; Valjent, E.; Girault, J.A. Opposing patterns of signaling activation in dopamine D1 and D2 receptor-expressing striatal neurons in response to cocaine and haloperidol. J. Neurosci. 2008, 28, 5671–5685. [Google Scholar] [CrossRef]
- De Sa Nogueira, D.; Merienne, K.; Befort, K. Neuroepigenetics and addictive behaviors: Where do we stand? Neurosci. Biobehav. Rev. 2019, 106, 58–72. [Google Scholar] [CrossRef]
- Heyer, M.P.; Kenny, P.J. Corticostriatal microRNAs in addiction. Brain Res. 2015, 1628, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Sadakierska-Chudy, A.; Frankowska, M.; Miszkiel, J.; Wydra, K.; Jastrzębska, J.; Filip, M. Prolonged Induction of miR-212/132 and REST Expression in Rat Striatum Following Cocaine Self-Administration. Mol. Neurobiol. 2017, 54, 2241–2254. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.L.; Edwards, S.; Bachtell, R.K.; DiLeone, R.J.; Rios, M.; Self, D.W. Dynamic BDNF activity in nucleus accumbens with cocaine use increases self-administration and relapse. Nat. Neurosci. 2007, 10, 1029–1037. [Google Scholar] [CrossRef]
- Shahid, Z.; Simpson, B.; Miao, K.H.; Singh, G. Genetics, Histone Code; StatPearls: Tampa/St. Petersburg, FL, USA, 2021. [Google Scholar]
- Shimaji, K.; Tomida, S.; Yamaguchi, M. Regulation of animal behavior by epigenetic regulators. Front. Biosci. 2019, 24, 1071–1084. [Google Scholar] [CrossRef]
- Bjorness, T.E.; Greene, R.W. Interaction between cocaine use and sleep behavior: A comprehensive review of cocaine’s disrupting influence on sleep behavior and sleep disruptions influence on reward seeking. Pharmacol. Biochem. Behav. 2021, 206, 173194. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saad, L.; Zwiller, J.; Kalsbeek, A.; Anglard, P. Epigenetic Regulation of Circadian Clocks and Its Involvement in Drug Addiction. Genes 2021, 12, 1263. https://doi.org/10.3390/genes12081263
Saad L, Zwiller J, Kalsbeek A, Anglard P. Epigenetic Regulation of Circadian Clocks and Its Involvement in Drug Addiction. Genes. 2021; 12(8):1263. https://doi.org/10.3390/genes12081263
Chicago/Turabian StyleSaad, Lamis, Jean Zwiller, Andries Kalsbeek, and Patrick Anglard. 2021. "Epigenetic Regulation of Circadian Clocks and Its Involvement in Drug Addiction" Genes 12, no. 8: 1263. https://doi.org/10.3390/genes12081263
APA StyleSaad, L., Zwiller, J., Kalsbeek, A., & Anglard, P. (2021). Epigenetic Regulation of Circadian Clocks and Its Involvement in Drug Addiction. Genes, 12(8), 1263. https://doi.org/10.3390/genes12081263