
Emerging Role of microRNAs and Long Non-Coding RNAs in Sjögren’s Syndrome

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Literature Search Strategy
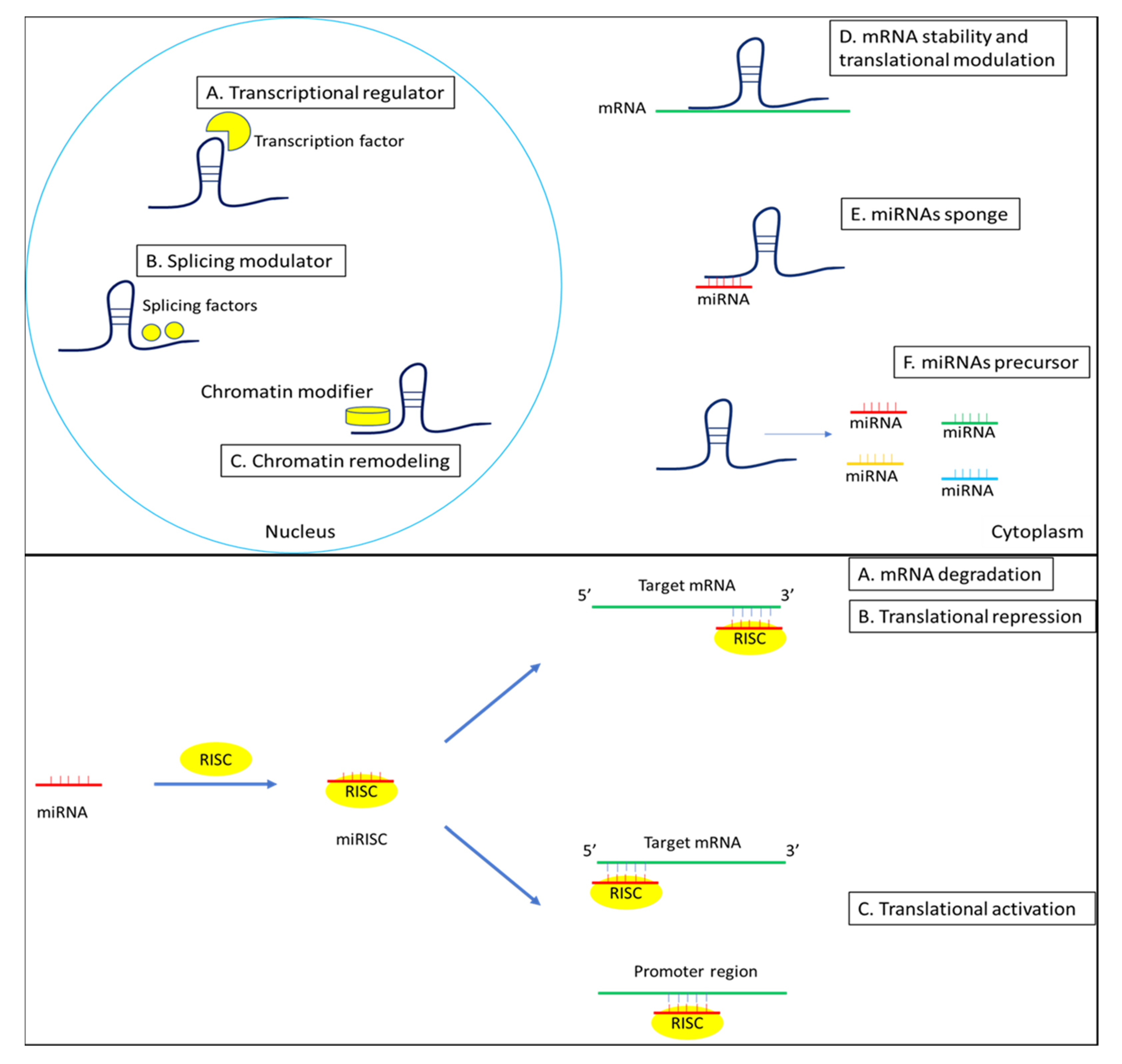
3. Roles of miRNAs in SS
3.1. miRNAs Dysregulated in PBMCs
3.2. miRNAs Dysregulated in Salivary Glands
3.3. miRNAs Dysregulated in Other Tissues
4. Roles of lncRNAs in SS
4.1. lncRNAs Dysregulated in PBMCs
4.2. lncRNAs Dysregulated in Salivary Gland
4.3. Genetics of lncRNAs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chivasso, C.; Sarrand, J.; Perret, J.; Delporte, C.; Soyfoo, M.S. The Involvement of Innate and Adaptive Immunity in the Initiation and Perpetuation of Sjögren’s Syndrome. Int. J. Mol. Sci. 2021, 22, 658. [Google Scholar] [CrossRef] [PubMed]
- Delli, K.; Vissink, A.; Spijkervet, F.K. Salivary gland biopsy for Sjögren’s syndrome. Oral. Maxillofac. Surg. Clin. N. Am. 2014, 26, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Cafaro, G.; Croia, C.; Argyropoulou, O.D.; Leone, M.C.; Orlandi, M.; Finamore, F.; Cecchettini, A.; Ferro, F.; Baldini, C.; Bartoloni, E. One year in review 2019: Sjögren’s syndrome. Clin. Exp. Rheumatol. 2019, 37, 3–15. [Google Scholar]
- Mavragani, C.P.; Moutsopoulos, H.M. Primary versus Secondary Sjögren Syndrome: Is It Time To Reconsider These Terms? J. Rheumatol. 2019, 46, 665–666. [Google Scholar] [CrossRef]
- Shiboski, C.H.; Shiboski, S.C.; Seror, R.; Criswell, L.A.; Labetoulle, M.; Lietman, T.M.; Rasmussen, A.; Scofield, H.; Vitali, C.; Bowman, S.J.; et al. 2016 American College of Rheumatology/European League Against Rheumatism Classification Criteria for Primary Sjögren’s Syndrome: A Consensus and Data-Driven Methodology Involving Three International Patient Cohorts. Arthritis Rheumatol. 2017, 69, 35–45. [Google Scholar] [CrossRef]
- Imgenberg-Kreuz, J.; Rasmussen, A.; Sivils, K.; Nordmark, G. Genetics and epigenetics in primary Sjögren’s syndrome. Rheumatology 2019, 60, 2085–2098. [Google Scholar] [CrossRef]
- Lessard, C.J.; Registry, F.U.P.S.S.; Li, H.; Adrianto, I.; Ice, J.A.; Rasmussen, A.; Grundahl, K.M.; A Kelly, J.; Dozmorov, M.; Miceli-Richard, C.; et al. Variants at multiple loci implicated in both innate and adaptive immune responses are associated with Sjögren’s syndrome. Nat. Genet. 2013, 45, 1284–1292. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, K.; Chen, H.; Sun, F.; Xu, J.; Wu, Z.; Li, P.; Zhang, L.; Du, Y.; Luan, H.; et al. A genome-wide association study in Han Chinese identifies a susceptibility locus for primary Sjögren’s syndrome at 7q11.23. Nat. Genet. 2013, 45, 1361–1365. [Google Scholar] [CrossRef]
- Yuan, X.; Berg, N.; Lee, J.W.; Le, T.-T.; Neudecker, V.; Jing, N.; Eltzschig, H. MicroRNA miR-223 as regulator of innate immunity. J. Leukoc. Biol. 2018, 104, 515–524. [Google Scholar] [CrossRef]
- Pauley, K.M.; Cha, S.; Chan, E.K. MicroRNA in autoimmunity and autoimmune diseases. J. Autoimmun. 2009, 32, 189–194. [Google Scholar] [CrossRef]
- Wei, Y.; Li, N.; Zhao, L.; Yang, C.; Ma, B.; Li, X.; Wei, R.; Nian, H. MicroRNAs and Autoimmune-Mediated Eye Diseases. Front. Cell Dev. Biol. 2020, 8, 818. [Google Scholar] [CrossRef] [PubMed]
- Hur, K.; Kim, S.H.; Kim, J.M. Potential Implications of Long Noncoding RNAs in Autoimmune Diseases. Immune Netw. 2019, 19, e4. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Liu, C.; Zhang, L.; Luo, G.; Liang, S. Research progress on the interactions between long non-coding RNAs and microRNAs in human cancer (Review). Oncol. Lett. 2019, 19, 595–605. [Google Scholar] [CrossRef]
- Rusca, N.; Monticelli, S. MiR-146a in Immunity and Disease. Mol. Biol. Int. 2011, 2011, 437301. [Google Scholar] [CrossRef] [PubMed]
- Pauley, K.M.; Stewart, C.M.; Gauna, A.E.; Dupre, L.C.; Kuklani, R.; Chan, A.L.; Pauley, B.A.; Reeves, W.H.; Chan, E.K.L.; Cha, S. Altered miR-146a expression in Sjögren’s syndrome and its functional role in innate immunity. Eur. J. Immunol. 2011, 41, 2029–2039. [Google Scholar] [CrossRef] [PubMed]
- Zilahi, E.; Tarr, T.; Papp, G.; Griger, Z.; Sipka, S.; Zeher, M. Increased microRNA-146a/b, TRAF6 gene and decreased IRAK1 gene expressions in the peripheral mononuclear cells of patients with Sjögren’s syndrome. Immunol. Lett. 2012, 141, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Zheng, L.Y.; Zhang, P.; Yu, C.Q. miR-146a and miR-155 expression in PBMCs from patients with Sjögren’s syndrome. J. Oral Pathol. Med. 2014, 43, 792–797. [Google Scholar] [CrossRef]
- Peng, L.; Ma, W.; Yi, F.; Yang, Y.-J.; Lin, W.; Chen, H.; Zhang, X.; Zhang, L.-H.; Zhang, F.; Du, Q. MicroRNA Profiling in Chinese Patients with Primary Sjögren Syndrome Reveals Elevated miRNA-181a in Peripheral Blood Mononuclear Cells. J. Rheumatol. 2014, 41, 2208–2213. [Google Scholar] [CrossRef]
- Wang-Renault, S.F.; Boudaoud, S.; Nocturne, G.; Roche, E.; Sigrist, N.; Daviaud, C.; Bugge Tinggaard, A.; Renault, V.; Deleuze, J.F.; Mariette, X.; et al. Deregulation of microRNA expression in purified T and B lymphocytes from patients with primary Sjögren’s syndrome. Ann. Rheum. Dis. 2018, 77, 133–140. [Google Scholar] [CrossRef]
- Varin, M.M.; Le Pottier, L.; Youinou, P.; Saulep, D.; Mackay, F.; Pers, J.O. B-cell tolerance breakdown in Sjögren’s syndrome: Focus on BAFF. Autoimmun. Rev. 2010, 9, 604–608. [Google Scholar] [CrossRef]
- Kiripolsky, J.; McCabe, L.G.; Kramer, J.M. Innate immunity in Sjögren’s syndrome. Clin. Immunol. 2017, 182, 4–13. [Google Scholar] [CrossRef]
- Lopes, A.P.; Van Roon, J.A.G.; Blokland, S.L.M.; Wang, M.; Chouri, E.; Hartgring, S.A.Y.; Van Der Wurff-Jacobs, K.M.G.; Kruize, A.A.; Burgering, B.M.T.; Rossato, M.; et al. MicroRNA-130a Contributes to Type-2 Classical DC-activation in Sjögren’s Syndrome by Targeting Mitogen- and Stress-Activated Protein Kinase-1. Front. Immunol. 2019, 10, 1335. [Google Scholar] [CrossRef]
- Starnes, T.; Broxmeyer, H.E.; Robertson, M.J.; Hromas, R. Cutting edge: IL-17D, a novel member of the IL-17 family, stimulates cytokine production and inhibits hemopoiesis. J. Immunol. 2002, 169, 642–646. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Wang, L.; Sun, C.; Xie, C.; Li, Z. MiR-let-7d-3p regulates IL-17 expression through targeting AKT1/mTOR signaling in CD4+ T cells. In Vitro Cell Dev. Biol. Anim. 2020, 56, 67–74. [Google Scholar] [CrossRef]
- Kassan, S.S.; Thomas, T.L.; Moutsopoulos, H.M.; Hoover, R.; Kimberly, R.; Budman, D.R.; Costa, J.; Decker, J.L.; Chused, T.M. Increased Risk of Lymphoma in Sicca Syndrome. Ann. Intern. Med. 1978, 89, 888–892. [Google Scholar] [CrossRef]
- Kapsogeorgou, E.K.; Papageorgiou, A.; Protogerou, A.D.; Voulgarelis, M.; Tzioufas, A.G. Low miR200b-5p levels in minor salivary glands: A novel molecular marker predicting lymphoma development in patients with Sjögren’s syndrome. Ann. Rheum. Dis. 2018, 77, 1200–1207. [Google Scholar] [CrossRef]
- Jang, S.I.; Tandon, M.; Teos, L.; Zheng, C.; Warner, B.M.; Alevizos, I. Dual function of miR-1248 links interferon induction and calcium signaling defects in Sjögren’s syndrome. EBioMedicine 2019, 48, 526–538. [Google Scholar] [CrossRef]
- Yan, T.; Shen, J.; Chen, J.; Zhao, M.; Guo, H.; Wang, Y. Differential expression of miR-17-92 cluster among varying histological stages of minor salivary gland in patients with primary Sjögren’s syndrome. Clin. Exp. Rheumatol. 2019, 37 (Suppl. 118), 49–54. [Google Scholar] [PubMed]
- Cortes-Troncoso, J.; Jang, S.I.; Perez, P.; Hidalgo, J.; Ikeuchi, T.; Greenwell-Wild, T.; Warner, B.M.; Moutsopoulos, N.M.; Alevizos, I. T cell exosome-derived miR-142-3p impairs glandular cell function in Sjögren’s syndrome. JCI Insight 2020, 5, e133497. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, P.; Fietta, P. Apoptosis and Sjögren syndrome. Semin. Arthritis Rheum. 2003, 33, 49–65. [Google Scholar] [CrossRef]
- Yang, Y.; Hou, Y.; Li, J.; Zhang, F.; Du, Q. Characterization of antiapoptotic microRNAs in primary Sjögren’s syndrome. Cell Biochem. Funct. 2020, 38, 1111–1118. [Google Scholar] [CrossRef]
- Sembler-Møller, M.L.; Belstrøm, D.; Locht, H.; Pedersen, A.M.L. Distinct microRNA expression profiles in saliva and salivary gland tissue differentiate patients with primary Sjögren’s syndrome from non-Sjögren’s sicca patients. J. Oral Pathol. Med. 2020, 49, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Yeon, Y.; Lee, W.J.; Shin, Y.U.; Cho, H.; Sung, Y.-K.; Kim, D.R.; Lim, H.W.; Kang, M.H. Comparison of MicroRNA Expression in Tears of Normal Subjects and Sjögren Syndrome Patients. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4889–4895. [Google Scholar] [CrossRef]
- Pilson, Q.; Smith, S.; Jefferies, C.A.; Ní Gabhann-Dromgoole, J.; Murphy, C.C. miR-744-5p contributes to ocular inflammation in patients with primary Sjogrens Syndrome. Sci. Rep. 2020, 10, 7484. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hao, Y.; Zhang, D.; Fu, R.; Liu, W.; Zhang, X.; Xue, F.; Yang, R. Aberrant expression of long noncoding RNA TMEVPG1 in patients with primary immune thrombocytopenia. Autoimmunity 2016, 49, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Peng, H.; Tian, J.; Ma, J.; Tang, X.; Rui, K.; Tian, X.; Wang, Y.; Chen, J.; Lu, L.; et al. Upregulation of long noncoding RNA TMEVPG1 enhances T helper type 1 cell response in patients with Sjögren syndrome. Immunol. Res. 2016, 64, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Inamo, J.; Suzuki, K.; Takeshita, M.; Kassai, Y.; Takiguchi, M.; Kurisu, R.; Okuzono, Y.; Tasaki, S.; Yoshimura, A.; Takeuchi, T. Identification of novel genes associated with dysregulation of B cells in patients with primary Sjögren’s syndrome. Arthritis Res. Ther. 2020, 22, 153. [Google Scholar] [CrossRef]
- Nocturne, G.; Mariette, X. B cells in the pathogenesis of primary Sjögren syndrome. Nat. Rev. Rheumatol. 2018, 14, 133–145. [Google Scholar] [CrossRef]
- Ye, L.; Shi, H.; Yu, C.; Fu, J.; Chen, C.; Wu, S.; Zhan, T.; Wang, B.; Zheng, L. LncRNA Neat1 positively regulates MAPK signaling and is involved in the pathogenesis of Sjögren’s syndrome. Int. Immunopharmacol. 2020, 88, 106992. [Google Scholar] [CrossRef]
- Singh, N.; Cohen, P.L. The T cell in Sjogren’s syndrome: Force majeure, not spectateur. J. Autoimmun. 2012, 39, 229–233. [Google Scholar] [CrossRef]
- Singh, N.; Cohen, P.L. LncRNA PVT1 links Myc to glycolytic metabolism upon CD4+ T cell activation and Sjögren’s syndrome-like autoimmune response. J. Autoimmun. 2020, 107, 102358. [Google Scholar]
- Peng, Y.; Luo, X.; Chen, Y.; Peng, L.; Deng, C.; Fei, Y.; Zhang, W.; Zhao, Y. LncRNA and mRNA expression profile of peripheral blood mononuclear cells in primary Sjögren’s syndrome patients. Sci. Rep. 2020, 10, 19629. [Google Scholar] [CrossRef]
- Dolcino, M.; Tinazzi, E.; Vitali, C.; Del Papa, N.; Puccetti, A.; Lunardi, C. Long Non-Coding RNAs Modulate Sjögren’s Syndrome Associated Gene Expression and Are Involved in the Pathogenesis of the Disease. J. Clin. Med. 2019, 8, 1349. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Cao, N.; Pu, Y.; Xie, L.; Zheng, L.; Yu, C. Long non-coding RNA expression profile in minor salivary gland of primary Sjögren’s syndrome. Arthritis Res. Ther. 2016, 18, 109. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.F.; Su, L.C.; Jia, H.; Liu, Y.; Xu, W.D. No association of single nucleotide polymorphisms within H19 and HOX transcript antisense RNA (HOTAIR) with genetic susceptibility to systemic lupus erythematosus, rheumatoid arthritis, and primary Sjögren’s syndrome in a Chinese Han population. Clin. Rheumatol. 2017, 36, 2447–2453. [Google Scholar] [CrossRef]
- Colafrancesco, S.; Ciccacci, C.; Priori, R.; Latini, A.; Picarelli, G.; Arienzo, F.; Novelli, G.; Valesini, G.; Perricone, C.; Borgiani, P. STAT4, TRAF3IP2, IL10, and HCP5 Polymorphisms in Sjögren’s Syndrome: Association with Disease Susceptibility and Clinical Aspects. J. Immunol. Res. 2019, 2019, 1–8. [Google Scholar] [CrossRef]


{kind=link}
| Tissues | miRNAs | Evidence | Refs. |
| PBMCs | hsa-miR-146a | up-regulated | [15,16,17] |
| hsa-miR-181a | up-regulated | [18] | |
| hsa-miR-30b | down-regulated | [19] | |
| hsa-miR-130a | down-regulated | [22] | |
| hsa-miR-708 | down-regulated | [22] | |
| hsa-let-7d | up-regulated | [24] | |
| Salivary glands | hsa-miR-200b | down-regulated in patients who have or will develop lymphoma | [26] |
| hsa-miR-1248 | up-regulated | [27] | |
| hsa-miR-18a | up-regulated | [28] | |
| hsa-miR-92a | down-regulated | [28] | |
| hsa-miR-142 | up-regulated | [29] | |
| hsa-miR-1207 | down-regulated | [31] | |
| hsa-miR-4695 | down-regulated | [31] | |
| hsa-miR-29a | up-regulated | [32] | |
| Saliva | hsa-miR-17 family | down-regulated | [32] |
| Lacrimal glands | hsa-miR-16 | up-regulated | [33] |
| hsa-miR-34a | up-regulated | [33] | |
| hsa-miR-142 | up-regulated | [33] | |
| hsa-miR-223 | up-regulated | [33] | |
| hsa-miR-30b,c,d | down-regulated | [33] | |
| hsa-miR-92a | down-regulated | [33] | |
| hsa-miR-134 | down-regulated | [33] | |
| hsa-miR-137 | down-regulated | [33] | |
| hsa-miR-302d | down-regulated | [33] | |
| hsa-miR-365b | down-regulated | [33] | |
| hsa-miR-374c | down-regulated | [33] | |
| hsa-miR-487b | down-regulated | [33] | |
| Conjunctiva | hsa-miR-744 | up-regulated | [34] |
| Tissues | lncRNAs | Evidence | Refs. |
| PBMCs | TMEVPG1 | up-regulated | [36] |
| LINC00487 | up-regulated | [37] | |
| NEAT1 | up-regulated | [39] | |
| PTV1 | up-regulated | [41] | |
| NRIR | up-regulated | [42] | |
| BISPR | up-regulated | [42] | |
| LINC0042 | up-regulated | [42] | |
| CYTOR | up-regulated | [42] | |
| TPTEP1 | down-regulated | [42] | |
| LINC00657 | up-regulated | [43] | |
| LINC00511 | down-regulated | [43] | |
| CTD-2020K17.1 | up-regulated | [43] | |
| Salivary glands | LINC00426-003 | up-regulated | [44] |
| AC017002.1 | up-regulated | [44] | |
| n336161 | up-regulated | [44] | |
| NR_002712 | up-regulated | [44] | |
| LINC02384 | up-regulated | [44] | |
| lnc-UTS2D-1:1 | up-regulated | [44] | |
| n340599 | up-regulated | [44] | |
| TCONS_l2_00014794 | up-regulated | [44] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Benedittis, G.; Ciccacci, C.; Latini, A.; Novelli, L.; Novelli, G.; Borgiani, P. Emerging Role of microRNAs and Long Non-Coding RNAs in Sjögren’s Syndrome. Genes 2021, 12, 903. https://doi.org/10.3390/genes12060903
De Benedittis G, Ciccacci C, Latini A, Novelli L, Novelli G, Borgiani P. Emerging Role of microRNAs and Long Non-Coding RNAs in Sjögren’s Syndrome. Genes. 2021; 12(6):903. https://doi.org/10.3390/genes12060903
Chicago/Turabian StyleDe Benedittis, Giada, Cinzia Ciccacci, Andrea Latini, Lucia Novelli, Giuseppe Novelli, and Paola Borgiani. 2021. "Emerging Role of microRNAs and Long Non-Coding RNAs in Sjögren’s Syndrome" Genes 12, no. 6: 903. https://doi.org/10.3390/genes12060903
APA StyleDe Benedittis, G., Ciccacci, C., Latini, A., Novelli, L., Novelli, G., & Borgiani, P. (2021). Emerging Role of microRNAs and Long Non-Coding RNAs in Sjögren’s Syndrome. Genes, 12(6), 903. https://doi.org/10.3390/genes12060903