
Allelic Diversity at Abiotic Stress Responsive Genes in Relationship to Ecological Drought Indices for Cultivated Tepary Bean, Phaseolus acutifolius A. Gray, and Its Wild Relatives


 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
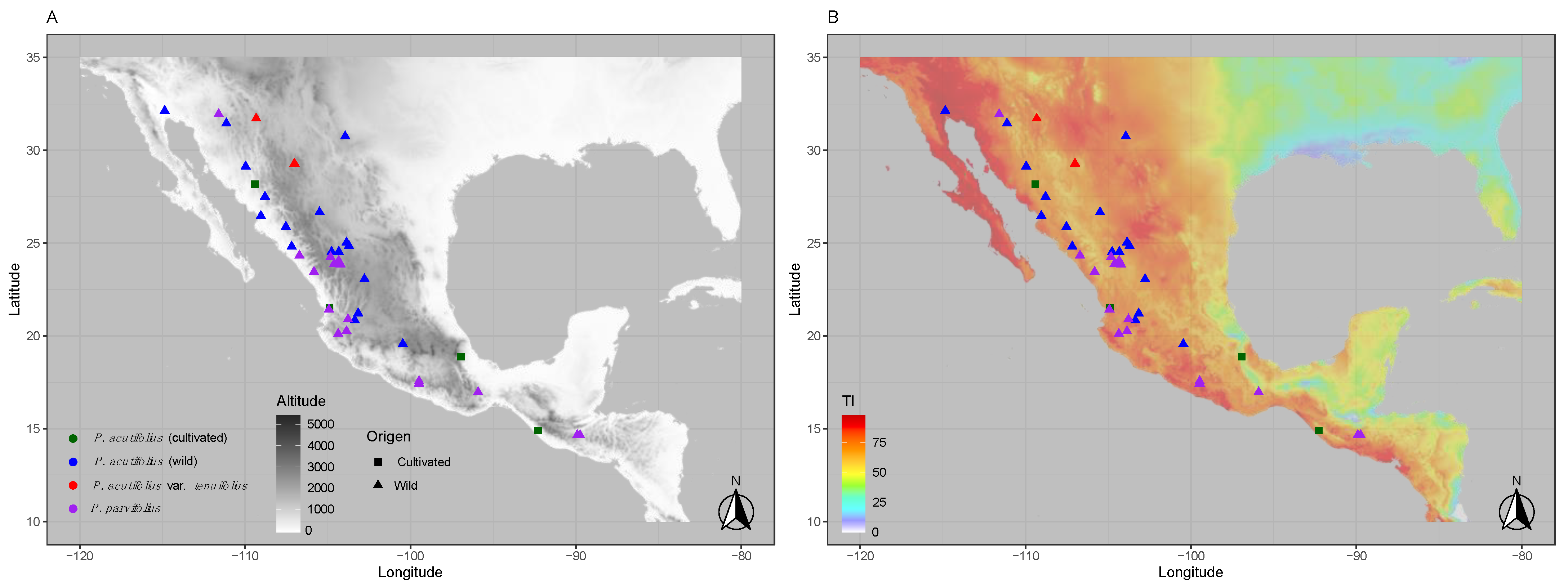
2.1. Plant Material
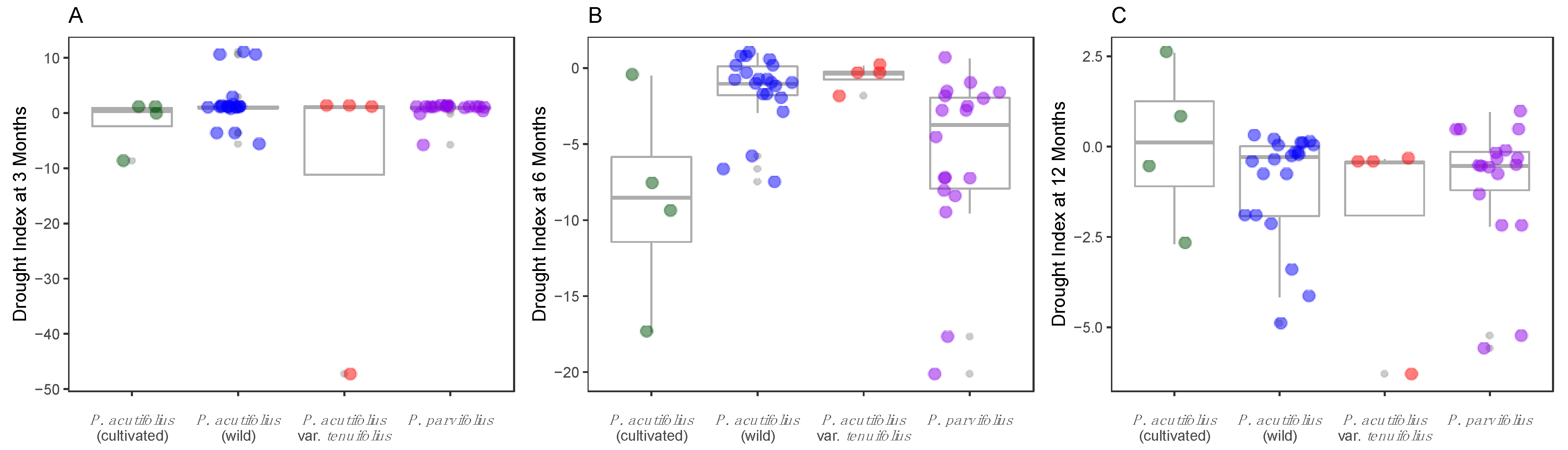
2.2. Habitat-Based Drought Stress Indices
2.3. DNA Extraction, Candidate Gene Amplification and Sequencing
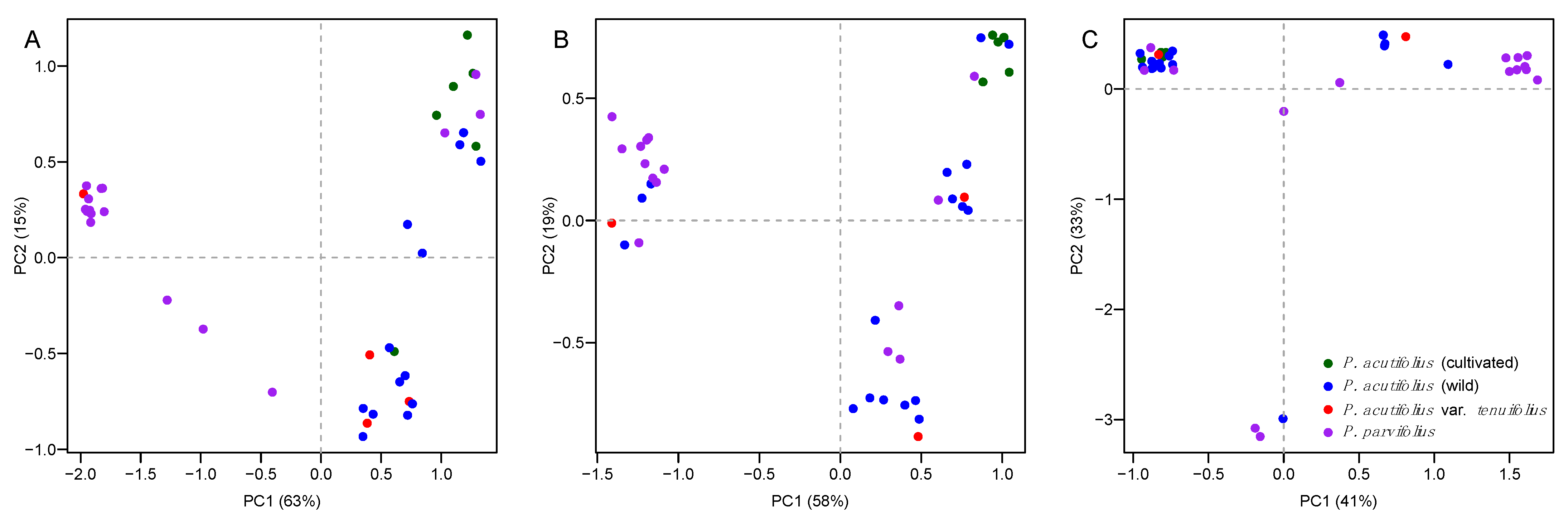
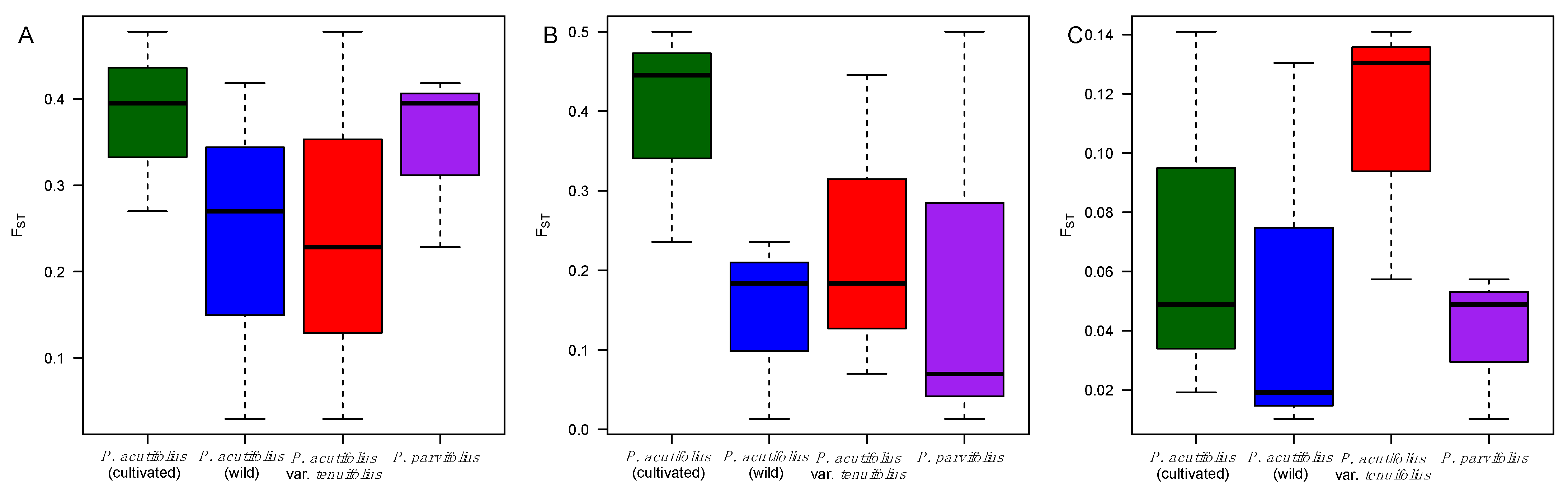
2.4. Patterns of Nucleotide Diversity and Environmental Correlations at Candidate Genes
3. Results
3.1. Pervasive Environmental and Allelic Diversity for Three Drought Candidate Genes in Tepary Bean s.l.
3.2. Allele Variants at Dreb2B and ERECTA-Encoding Candidate Genes Were Correlated with Drought Stress
4. Discussion
4.1. Tepary Bean s.l. Is a Reservoir of Novel Alleles at Candidate Genes for Drought Tolerance
4.2. Unlocking Useful Genetic Variation for Drought Tolerance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Barnard, P.; Moomaw, W.R. World Scientists’ Warning of a Climate Emergency. BioScience 2020, 70, 8–12. [Google Scholar] [CrossRef]
- Tai, A.P.K.; Martin, M.V.; Heald, C.L. Threat to future global food security from climate change and ozone air pollution. Nat. Clim. Chang. 2014, 4, 817–821. [Google Scholar] [CrossRef]
- Donoghue, M.J. Adaptation meets dispersal: Legumes in the land of succulents. New Phytol. 2019, 222, 1667–1669. [Google Scholar] [CrossRef] [PubMed]
- Coyne, C.J.; Kumar, S.; von Wettberg, E.J.B.; Marques, E.; Berger, J.D.; Redden, R.J.; Ellis, T.H.N.; Brus, J.; Zablatzká, L.; Smýkal, P. Potential and limits of exploitation of crop wild relatives for pea, lentil, and chickpea improvement. Legume Sci. 2020, 2, e36. [Google Scholar] [CrossRef]
- Borelli, T.; Hunter, D.; Powell, B.; Ulian, T.; Mattana, E.; Termote, C.; Pawera, L.; Beltrame, D.; Penafiel, D.; Tan, A.; et al. Born to Eat Wild: An Integrated Conservation Approach to Secure Wild Food Plants for Food Security and Nutrition. Plants 2020, 9, 1299. [Google Scholar] [CrossRef] [PubMed]
- Smýkal, P.; Coyne, C.J.; Ambrose, M.J.; Maxted, N.; Schaefer, H.; Blair, M.W.; Berger, J.; Greene, S.L.; Nelson, M.N.; Besharat, N.; et al. Legume Crops Phylogeny and Genetic Diversity for Science and Breeding. Crit. Rev. Plant. Sci. 2014, 34, 43–104. [Google Scholar] [CrossRef]
- Baudoin, J.-P.; Maquet, A. Improvement of protein and amino acid contents in seeds of food legumes. A case study in Phaseolus. Biotechnol. Agron. Soc. Environ. 1999, 3, 220–224. [Google Scholar]
- Gepts, P.; Aragão, F.; Barros, E.; Blair, M.; Brondani, R.; Broughton, W.; Galasso, I.; Hernandez, G.; Kami, J.; Lariguet, P.; et al. Genomics of Phaseolus Beans, a Major Source of Dietary Protein and Micronutrients in the Tropics. In Genomics of Tropical Crop Plants; Moore, P., Ming, R., Eds.; Springer: New York, NY, USA, 2008. [Google Scholar]
- Blair, M.W.; Pantoja, W.; Carmenza Munoz, L. First use of microsatellite markers in a large collection of cultivated and wild accessions of tepary bean (Phaseolus acutifolius A. Gray). Theor. Appl. Genet. 2012, 125, 1137–1147. [Google Scholar] [CrossRef]
- Mhlaba, Z.B.; Amelework, B.; Shimelis, H.; Modi, A.T.; Mashilo, J. Genetic differentiation among selected tepary bean collections revealed by morphological traits and simple sequence repeat markers. Acta Agric. Scand. Sect. B-Soil Plant. Sci. 2018, 68, 608–618. [Google Scholar] [CrossRef]
- Mhlaba, Z.B.; Mashilo, J.; Shimelis, H.; Assefa, A.B.; Modi, A.T. Progress in genetic analysis and breeding of tepary bean (Phaseolus acutifolius A. Gray): A review. Sci. Hortic. 2018, 237, 112–119. [Google Scholar] [CrossRef]
- Bitocchi, E.; Rau, D.; Bellucci, E.; Rodriguez, M.; Murgia, M.L.; Gioia, T.; Santo, D.; Nanni, L.; Attene, G.; Papa, R. Beans (Phaseolus ssp.) as a Model for Understanding Crop Evolution. Front. Plant. Sci 2017, 8, 722. [Google Scholar] [CrossRef] [PubMed]
- Debouck, D.G. Biodiversity, ecology and genetic resources of Phaseolus beans–Seven answered and unanswered questions. In Proceedings of the Wild Legumes, Tsukuba, Japan, 14 September 2009; pp. 95–123. [Google Scholar]
- Parsons, L.R.; Howe, T.K. Effects of water stress on the water relations of Phaseolus vulgaris and the drought resistant Phaseolus acutifolius. Physiol. Plant. 1984, 60, 197–202. [Google Scholar] [CrossRef]
- Cruz Jimenez-Galindo, J. Screening for Drought Tolerance in Tepary and Common Bean Based on Osmotic Potential Assays. Plant 2018, 6, 24. [Google Scholar] [CrossRef]
- Muñoz, L.C.; Duque, M.C.; Debouck, D.G.; Blair, M.W. Taxonomy of tepary bean and wild relatives as determined by amplified fragment length polymorphism (AFLP) markers. Crop. Sci. 2006, 46, 1744–1754. [Google Scholar] [CrossRef]
- Porch, T.; Beaver, J.; Debouck, D.; Jackson, S.; Kelly, J.; Dempewolf, H. Use of Wild Relatives and Closely Related Species to Adapt Common Bean to Climate Change. Agronomy 2013, 3, 433–461. [Google Scholar] [CrossRef]
- Singh, S.P. Broadening the genetic base of common bean cultivars: A review. Crop. Sci. 2001, 41, 1659–1675. [Google Scholar] [CrossRef]
- Muñoz, L.C.; Blair, M.W.; Duque, M.C.; Tohme, J.; Roca, W. Introgression in common bean x tepary bean interspecific congruity-backcross lines as measured by AFLP markers. Crop. Sci. 2003, 44, 637–645. [Google Scholar] [CrossRef]
- Mejía-Jiménez, A.; Muñoz, C.; Jacobsen, H.J.; Roca, W.M.; Singh, S.P. Interspecific hybridization between commonand tepary beans: Increased hybrid embryo growth, fertility, andefficiency of hybridization through recurrent and congruity backcrossing. Theor. Appl. Genet. 1994, 88, 324–331. [Google Scholar] [CrossRef]
- Souter, J.R.; Gurusamy, V.; Porch, T.G.; Bett, K.E. Successful Introgression of Abiotic Stress Tolerance from Wild Tepary Bean to Common Bean. Crop. Sci. 2017, 57, 1160–1171. [Google Scholar] [CrossRef]
- Crespo, S.; Rivera-Peña, M.; Rosero-Alpala, D.A.; Muñoz-Florez, J.E.; Rao, I.M.; Muñoz-Florez, L.C. Pollen viability of Tepary bean (Phaseolus acutifolius A. Gray.) mutant lines under water stress conditions and inoculation with rhizobia. Acta Agronómica 2018, 67, 319–325. [Google Scholar] [CrossRef]
- Gujaria-Verma, N.; Ramsay, L.; Sharpe, A.G.; Sanderson, L.-A.; Debouck, D.G.; Tar’an, B.; Bett, K.E. Gene-based SNP discovery in tepary bean (Phaseolus acutifolius) and common bean (P. vulgaris) for diversity analysis and comparative mapping. BMC Genom. 2016, 17, 1–16. [Google Scholar] [CrossRef]
- Beebe, S.E.; Rao, I.M.; Blair, M.W.; Acosta-Gallegos, J.A. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W.; Galeano, C.H.; Tovar, E.; Muñoz Torres, M.C.; Castrillón, A.V.; Beebe, S.E.; Rao, I.M. Development of a Mesoamerican intra-genepool genetic map for quantitative trait loci detection in a drought tolerant × susceptible common bean (Phaseolus vulgaris L.) cross. Mol. Breed. 2012, 29, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Asfaw, A.; Ambachew, D.; Shah, T.; Blair, M.W. Trait Associations in Diversity Panels of the Two Common Bean (Phaseolus vulgaris L.) Gene Pools Grown under Well-watered and Water-Stress Conditions. Front. Plant. Sci. 2017, 8, 733. [Google Scholar] [CrossRef] [PubMed]
- Gibson, M.J.S.; Moyle, L.C. Regional differences in the abiotic environment contribute to genomic divergence within a wild tomato species. Mol. Ecol. 2020. [Google Scholar] [CrossRef]
- Diaz, L.M.; Ricaurte, J.; Tovar, E.; Cajiao, C.; Teran, H.; Grajales, M.; Polania, J.; Rao, I.; Beebe, S.; Raatz, B. QTL analyses for tolerance to abiotic stresses in a common bean (Phaseolus vulgaris L.) population. PLoS ONE 2018, 13, e0202342. [Google Scholar] [CrossRef]
- Cortés, A.J.; Blair, M.W. Genotyping by Sequencing and Genome—Environment Associations in Wild Common Bean Predict Widespread Divergent Adaptation to Drought. Front. Plant. Sci. 2018, 9, 128. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant. Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Sachdeva, S.; Bharadwaj, C.; Singh, R.K.; Jain, P.K.; Patil, B.S.; Roorkiwal, M.; Varshney, R. Characterization of ASR gene and its role in drought tolerance in chickpea (Cicer arietinum L.). PLoS ONE 2020, 15, e0234550. [Google Scholar] [CrossRef] [PubMed]
- Frankel, N.; Carrari, F.; Hasson, E.; Lusem, N.D. Evolutionary history of the Asr gene family. Gene 2006, 378, 74–83. [Google Scholar] [CrossRef]
- Philippe, R.; Courtois, B.; Billot, C.; Rami, J.F.; This, D. Allelic diversity at orthologous candidate genes for drought tolerance in crops: Examples of the ASR gene family. In Plant Genomics European Meetings, Venice; Avenue Media: Bologne, Italy, 2006. [Google Scholar]
- Huber, S.C.; Huber, J.L. Role and regulation of sucrose-phosphate synthase in higher plants. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1996, 47, 431–444. [Google Scholar] [CrossRef]
- Cortés, A.J.; Chavarro, M.C.; Madriñán, S.; This, D.; Blair, M.W. Molecular ecology and selection in the drought-related Asr gene polymorphisms in wild and cultivated common bean (Phaseolus vulgaris L.). BMC Genet. 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Plant. Physiology, 4th ed.; Sinauer Associates: Sunderland, MA, USA, 2006. [Google Scholar]
- Agarwal, P.K.; Agarwal, P.; Reddy, M.K.; Sopory, S.K. Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant. Cell Rep. 2006, 25, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Meyerowitz, E.M. The AP2/EREBP family of plant transcription factors. Biol. Chem. 1998, 379, 633–646. [Google Scholar] [PubMed]
- Kim, J.-S.; Mizoi, J.; Yoshida, T.; Fujita, Y.; Nakajima, J.; Ohori, T.; Todaka, D.; Nakashima, K.; Hirayama, T.; Shinozaki, K.; et al. An ABRE promoter sequence is involved in osmotic stress-responsive expression of the DREB2A gene, which encodes a transcription factor regulating drought-inducible genes in Arabidopsis. Plant. Cell Physiol. 2011, 52, 2136–2146. [Google Scholar] [CrossRef]
- Cortés, A.J.; Monserrate, F.; Ramírez-Villegas, J.; Madriñán, S.; Blair, M.W. Drought Tolerance in Wild Plant Populations: The Case of Common Beans (Phaseolus vulgaris L.). PLoS ONE 2013, 8, e62898. [Google Scholar] [CrossRef]
- Cortés, A.J.; This, D.; Chavarro, C.; Madriñán, S.; Blair, M.W. Nucleotide diversity patterns at the drought-related DREB2 encoding genes in wild and cultivated common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2012, 125, 1069–1085. [Google Scholar] [CrossRef]
- Masle, J.; Gilmore, S.R.; Farquhar, G.D. The ERECTA gene regulates plant transpiration efficiency in Arabidopsis. Nature 2005, 436, 866–870. [Google Scholar] [CrossRef]
- Torii, K.U.; Mitsukawa, N.; Oosumi, T.; Matsuura, Y.; Yokayama, R.; Whittier, R.F.; Komeda, Y. The Arabidopsis ERECTA gene encodes a putative receptor protein kinase with extracellular leucine-rich repeats. Plant. Cell 1996, 8, 735–746. [Google Scholar]
- Tisne, S.; Barbier, F.; Granier, C. The ERECTA gene controls spatial and temporal patterns of epidermal cell number and size in successive developing leaves of Arabidopsis thaliana. Ann. Bot. 2011, 108, 159–168. [Google Scholar] [CrossRef]
- Zanten, M.; Snoek, L.B.; Proveniers, M.; Peeters, A.J. The many functions of ERECTA. Trends Plant. Sci. 2009, 14, 214–217. [Google Scholar] [CrossRef]
- Barghi, N.; Hermisson, J.; Schlötterer, C. Polygenic adaptation: A unifying framework to understand positive selection. Nat. Rev. Genet. 2020, 21, 769–781. [Google Scholar] [CrossRef]
- Burgarella, C.; Barnaud, A.; Kane, N.A.; Jankowski, F.; Scarcelli, N.; Billot, C.; Vigouroux, Y.; Berthouly-Salazar, C. Adaptive Introgression: An Untapped Evolutionary Mechanism for Crop Adaptation. Front. Plant. Sci. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W.; Cortés, A.J.; This, D. Identification of an ERECTA gene and its drought adaptation associations with wild and cultivated common bean. Plant. Sci. 2016, 242, 250–259. [Google Scholar] [CrossRef]
- Jiri, O.; Mafongoya, P.; Chivenge, P. Tepary Bean: A Climate Smart Crop for Food and Nutritional Security. J. Nutr. Food Sci. 2016, 6. [Google Scholar] [CrossRef]
- Mwale, S.E.; Shimelis, H.; Mafongoya, P.; Mashilo, J.; Link, W. Breeding tepary bean (Phaseolus acutifolius) for drought adaptation: A review. Plant. Breed. 2020, 139, 821–833. [Google Scholar] [CrossRef]
- Miklas, P.N.; Kelly, J.D.; Beebe, S.E.; Blair, M.W. Common bean breeding for resistance against biotic and abiotic stresses: From classical to MAS breeding. Euphytica 2006, 147, 105–131. [Google Scholar] [CrossRef]
- Herzog, E.; Frisch, M. Selection strategies for marker-assisted backcrossing with high-throughput marker systems. Theor. Appl. Genet. 2011, 123, 251–260. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Crossa, J.; Perez-Rodriguez, P.; Cuevas, J.; Montesinos-Lopez, O.; Jarquin, D.; de Los Campos, G.; Burgueno, J.; Gonzalez-Camacho, J.M.; Perez-Elizalde, S.; Beyene, Y.; et al. Genomic Selection in Plant Breeding: Methods, Models, and Perspectives. Trends Plant. Sci. 2017, 22, 961–975. [Google Scholar] [CrossRef]
- Thornthwaite, C.W.; Mather, J.R. The Water Balance. Climatology 1955, 8, 1–104. [Google Scholar]
- López-Hernández, F.; Cortés, A.J. Last-Generation Genome–Environment Associations Reveal the Genetic Basis of Heat Tolerance in Common Bean (Phaseolus vulgaris L.). Front. Genet. 2019, 10, 22. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant. Mol. Biol. Rep. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, R.M.; Ramdoss, Y.; Buckler, E.S. TASSELL Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 2011, 3, 109. [Google Scholar]
- Barton, N.; Hermisson, J.; Nordborg, M. Why structure matters. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Oord, E.J.C.G.v.d.; Sullivan, P.F. False discoveries and models for gene discovery. Trends Genet. 2003, 19, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Watterson, G.A. Number of segregating sites in genetic models without recombination. Theor. Popul. Biol. 1975, 7, 256–276. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Bellucci, E.; Bitocchi, E.; Ferrarini, A.; Benazzo, A.; Biagetti, E.; Klie, S.; Minio, A.; Rau, D.; Rodriguez, M.; Panziera, A.; et al. Decreased Nucleotide and Expression Diversity and Modified Coexpression Patterns Characterize Domestication in the Common Bean. Plant. Cell 2014, 26, 1901–1911. [Google Scholar] [CrossRef]
- Bitocchi, E.; Rau, D.; Benazzo, A.; Bellucci, E.; Goretti, D.; Biagetti, E.; Panziera, A.; Laido, G.; Rodriguez, M.; Gioia, T.; et al. High Level of Nonsynonymous Changes in Common Bean Suggests That Selection under Domestication Increased Functional Diversity at Target Traits. Front. Plant. Sci. 2016, 7, 2005. [Google Scholar] [CrossRef] [PubMed]
- Chacon-Sanchez, M.I.; Martinez-Castillo, J. Testing Domestication Scenarios of Lima Bean (Phaseolus lunatus L.) in Mesoamerica: Insights from Genome-Wide Genetic Markers. Front. Plant. Sci. 2017, 8, 1551. [Google Scholar] [CrossRef]
- Liu, A.; Ku, Y.-S.; Contador, C.A.; Lam, H.-M. The Impacts of Domestication and Agricultural Practices on Legume Nutrient Acquisition Through Symbiosis With Rhizobia and Arbuscular Mycorrhizal Fungi. Front. Genet. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yu, X.; Qin, Q.; Dinkins, R.D.; Zhu, H. The Impacts of Domestication and Breeding on Nitrogen Fixation Symbiosis in Legumes. Front. Genet. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Ku, Y.-S.; Contador, C.A.; Ng, M.-S.; Yu, J.; Chung, G.; Lam, H.-M. The Effects of Domestication on Secondary Metabolite Composition in Legumes. Front. Genet. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-W.; Lam, H.-M. The Modification of Circadian Clock Components in Soybean During Domestication and Improvement. Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Gepts, P.; Debouck, D. Origin, domestication and evolution of the common bean (Phaseolus vulgaris L.). In Common Beans: Research for Crop Improvement; Van Shoonhoven, A., Voysest, O., Eds.; Commonwealth Agricultural Bureau: Wallingford, UK, 1991; pp. 7–53. [Google Scholar]
- Gepts, P. Biochemical Evidence Bearing on the domestication of beans. Econ. Bot. 1990, 44, 28–38. [Google Scholar] [CrossRef]
- Cortés, A.J.; Chavarro, M.C.; Blair, M.W. SNP marker diversity in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2011, 123, 827–845. [Google Scholar] [CrossRef]
- Blair, M.W.; Soler, A.; Cortés, A.J. Diversification and Population Structure in Common Beans (Phaseolus vulgaris L.). PLoS ONE 2012, 7, e49488. [Google Scholar] [CrossRef]
- Broughton, W.J.; Hernandez, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model food legumes. Plant. Soil 2003, 252, 55–128. [Google Scholar] [CrossRef]
- Andermann, T.; Torres Jimenez, M.F.; Matos-Maravi, P.; Batista, R.; Blanco-Pastor, J.L.; Gustafsson, A.L.S.; Kistler, L.; Liberal, I.M.; Oxelman, B.; Bacon, C.D.; et al. A Guide to Carrying Out a Phylogenomic Target Sequence Capture Project. Front. Genet. 2019, 10, 1407. [Google Scholar] [CrossRef]
- Blair, M.W.; Cortes, A.J.; Farmer, A.D.; Huang, W.; Ambachew, D.; Penmetsa, R.V.; Carrasquilla-Garcia, N.; Assefa, T.; Cannon, S.B. Uneven recombination rate and linkage disequilibrium across a reference SNP map for common bean (Phaseolus vulgaris L.). PLoS ONE 2018, 13, e0189597. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, C.T.; Wilkin, J.; Zhuang, J.; Cortés, A.J.; Quintero, Á.L.P.; Gallagher, T.F.; Bohlmann, J.; Douglas, C.J.; Ellis, B.E.; Ritland, K. SNP discovery, gene diversity, and linkage disequilibrium in wild populations of Populus tremuloides. Tree Genet. Genomes 2012, 8, 821–829. [Google Scholar] [CrossRef]
- Cortés, A.J.; López-Hernández, F.; Osorio-Rodriguez, D. Predicting thermal adaptation by looking into populations’ genomic past. Front. Genet. 2020, 11, 564515. [Google Scholar] [CrossRef]
- Cortés, A.J.; Restrepo-Montoya, M.; Bedoya-Canas, L.E. Modern Strategies to Assess and Breed Forest Tree Adaptation to Changing Climate. Front. Plant. Sci. 2020, 11, 583323. [Google Scholar] [CrossRef] [PubMed]
- Madriñán, S.; Cortés, A.J.; Richardson, J.E. Páramo is the world’s fastest evolving and coolest biodiversity hotspot. Front. Genet. 2013, 4, 192. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, C.D.; Wakeley, J.; Sawyer, S.; Hartl, D.L. Directional Selection and the Site-Frequency Spectrum. Genetics 2001, 159, 1779–1788. [Google Scholar] [PubMed]
- Yeaman, S.; Otto, S.P. Establishment and maintenance of adaptive genetic divergence under migration, selection, and drift. Evolution 2011, 65, 2123–2129. [Google Scholar] [CrossRef]
- Wakeley, J. Coalescent Theory: An. Introduction; Harvard University: Cambridge, MA, USA, 2008. [Google Scholar]
- Barrett, R.D.H.; Hoekstra, H.E. Molecular spandrels: Tests of adaptation at the genetic level. Nat. Rev. Genet. 2011, 12, 767–780. [Google Scholar] [CrossRef]
- Schmutz, J.; McClean, P.E.; Mamidi, S.; Wu, G.A.; Cannon, S.B.; Grimwood, J.; Jenkins, J.; Shu, S.; Song, Q.; Chavarro, C.; et al. A reference genome for common bean and genome-wide analysis of dual domestications. Nat. Genet. 2014, 46, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; Skeen, P.; Blair, M.W.; Chacón-Sánchez, M.I. Does the genomic landscape of species divergence in Phaseolus beans coerce parallel signatures of adaptation and domestication? Front. Plant. Sci 2018, 9, 1816. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J. On the Origin of the Common Bean (Phaseolus vulgaris L.). Am. J. Plant. Sci. 2013, 4, 1998–2000. [Google Scholar] [CrossRef]
- Caliebe, A. Mathematical models in population genetics. Med. Genet. 2008, 20, 282–287. [Google Scholar] [CrossRef]
- Bryant, D.; Bouckaert, R.; Felsenstein, J.; Rosenberg, N.A.; RoyChoudhury, A. Inferring species trees directly from biallelic genetic markers: Bypassing gene trees in a full coalescent analysis. Mol. Biol. Evol. 2012, 29, 1917–1932. [Google Scholar] [CrossRef] [PubMed]
- Cornuet, J.M.; Pudlo, P.; Veyssier, J.; Dehne-Garcia, A.; Gautier, M.; Leblois, R.; Marin, J.M.; Estoup, A. DIYABC v2.0: A software to make approximate Bayesian computation inferences about population history using single nucleotide polymorphism, DNA sequence and microsatellite data. Bioinformatics 2014, 30, 1187–1189. [Google Scholar] [CrossRef]
- Cortés, A.J.; Garzón, L.N.; Valencia, J.B.; Madriñán, S. On the Causes of Rapid Diversification in the Páramos: Isolation by Ecology and Genomic Divergence in Espeletia. Front. Plant. Sci. 2018, 9, 1700. [Google Scholar] [CrossRef] [PubMed]
- Arenas, S.; Cortés, A.J.; Mastretta-Yanes, A.; Jaramillo-Correa, J.P. Evaluating the accuracy of genomic prediction for the management and conservation of relictual natural tree populations. Tree Genet. Genomes 2021, 17. [Google Scholar] [CrossRef]
- Capblancq, T.; Fitzpatrick, M.C.; Bay, R.A.; Exposito-Alonso, M.; Keller, S.R. Genomic Prediction of (Mal)Adaptation Across Current and Future Climatic Landscapes. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 245–269. [Google Scholar] [CrossRef]
- Reyes-Herrera, P.H.; Muñoz-Baena, L.; Velásquez-Zapata, V.; Patiño, L.; Delgado-Paz, O.A.; Díaz-Diez, C.A.; Navas-Arboleda, A.A.; Cortés, A.J. Inheritance of Rootstock Effects in Avocado (Persea americana Mill.) cv. Hass. Front. Plant. Sci. 2020, 11, 555071. [Google Scholar] [CrossRef] [PubMed]
- Valencia, J.B.; Mesa, J.; León, J.G.; Madriñán, S.; Cortés, A.J. Climate Vulnerability Assessment of the Espeletia Complex on Páramo Sky Islands in the Northern Andes. Front. Ecol. Evol. 2020, 8. [Google Scholar] [CrossRef]
- Cortés, A.J.; Blair, M.W. Lessons from Common Bean on How Wild Relatives and Landraces Can Make Tropical Crops More Resistant to Climate Change. In Rediscovery of Landraces as a Resource for the Future; Grillo, O., Ed.; InTech: Rijeka, Croatia, 2018. [Google Scholar] [CrossRef]
- Cortés, A.J.; Wheeler, J.A. The Environmental Heterogeneity of Mountains at a Fine Scale in a Changing World. In Mountains, Climate, and Biodiversity; Hoorn, C., Perrigo, A., Antonelli, A., Eds.; Wiley: New York, NY, USA, 2018. [Google Scholar]
- Hendrick, M.F.; Finseth, F.R.; Mathiasson, M.E.; Palmer, K.A.; Broder, E.M.; Breigenzer, P.; Fishman, L. The genetics of extreme microgeographic adaptation: An integrated approach identifies a major gene underlying leaf trichome divergence in Yellowstone Mimulus guttatus. Mol. Ecol. 2016, 25, 5647–5662. [Google Scholar] [CrossRef]
- Galeano, C.H.; Cortés, A.J.; Fernandez, A.C.; Soler, A.; Franco-Herrera, N.; Makunde, G.; Vanderleyden, J.; Blair, M.W. Gene-Based Single Nucleotide Polymorphism Markers for Genetic and Association Mapping in Common Bean. BMC Genet. 2012, 13, 48. [Google Scholar] [CrossRef] [PubMed]
- Cortés, A.J.; Wheeler, J.A.; Sedlacek, J.; Lexer, C.; Karrenberg, S. Genome-wide patterns of microhabitat-driven divergence in the alpine dwarf shrub Salix herbacea L. In On The Big Challenges of a Small Shrub: Ecological Genetics of Salix herbacea L.; Acta Universitatis Upsaliensis: Uppsala, Switzerland, 2015. [Google Scholar]
- Blair, M.W.; Cortés, A.J.; Penmetsa, R.V.; Farmer, A.; Carrasquilla-Garcia, N.; Cook, D.R. A high-throughput SNP marker system for parental polymorphism screening, and diversity analysis in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2013, 126, 535–548. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Hirschhorn, J.N.; Daly, M.J. Genome-wide association studies for common diseases and complex traits. Nat. Rev. Genet. 2005, 6, 95–108. [Google Scholar] [CrossRef]
- Rafalski, J.A. Association genetics in crop improvement. Curr. Opin. Plant. Biol. 2010, 13, 174–180. [Google Scholar] [CrossRef]
- Manolio, T.A.; Collins, F.S.; Cox, N.J.; Goldstein, D.B.; Hindorff, L.A.; Hunter, D.J.; McCarthy, M.I.; Ramos, E.M.; Cardon, L.R.; Chakravarti, A.; et al. Finding the missing heritability of complex diseases. Nature 2009, 461, 747–753. [Google Scholar] [CrossRef]
- Yang, J.; Benyamin, B.; McEvoy, B.P.; Gordon, S.; Henders, A.K.; Nyholt, D.R.; Madden, P.A.; Heath, A.C.; Martin, N.G.; Montgomery, G.W.; et al. Common SNPs explain a large proportion of the heritability for human height. Nature Genet. 2010, 42, 565–569. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
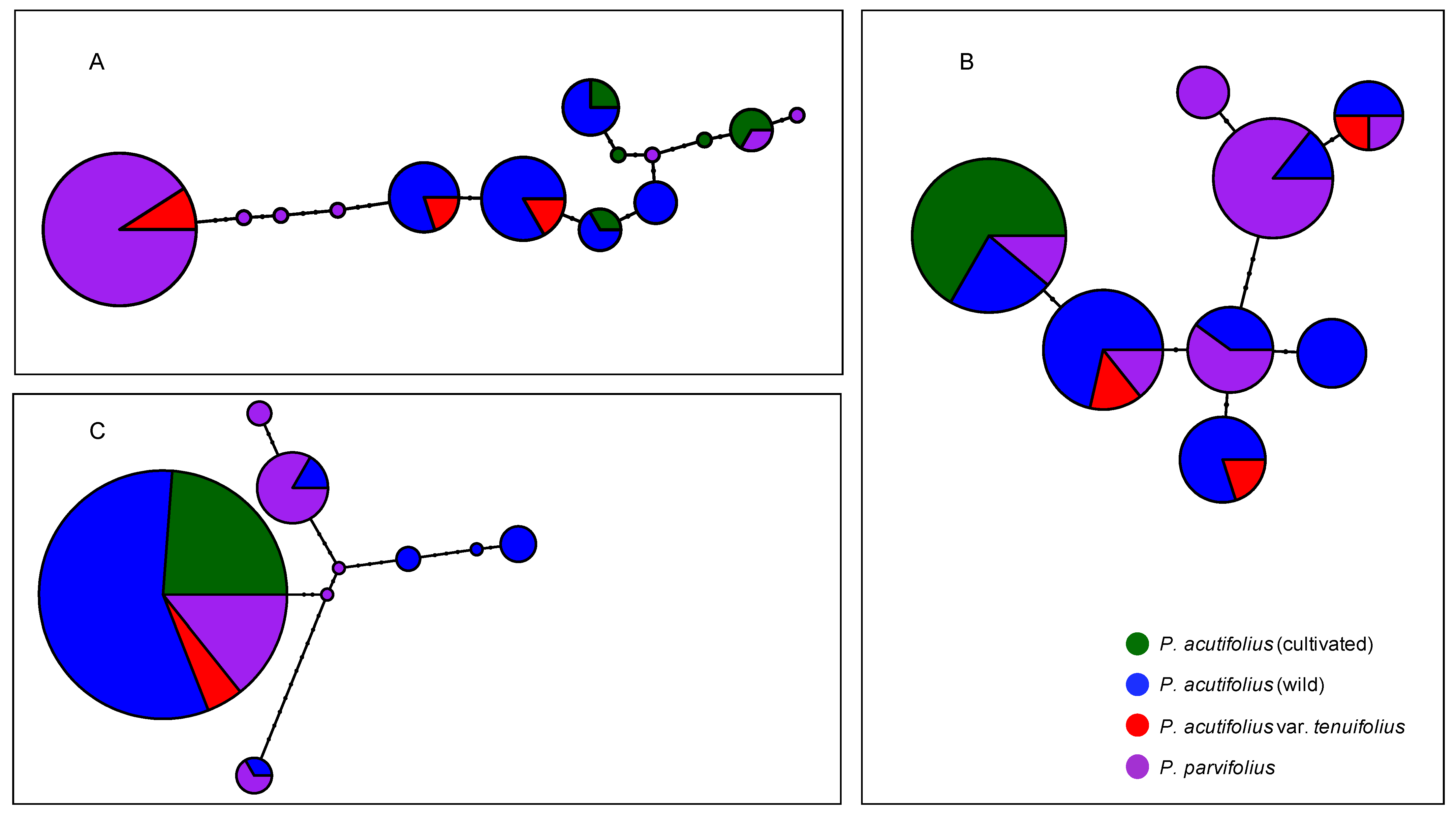{kind=link}
| Gene Name | Position (bp) | Major Allele | Major Allele Frequency | Minor Allele | Minor Allele Frequency | He |
|---|---|---|---|---|---|---|
| Asr2 | 1 | C | 0.5 | T | 0.5 | 0.5 |
| Asr2 | 101 | A | 0.67 | G | 0.33 | 0.44 |
| Asr2 | 185 | T | 0.73 | A | 0.27 | 0.4 |
| Asr2 | 188 | T | 0.55 | A | 0.45 | 0.5 |
| Asr2 | 189 | A | 0.67 | T | 0.33 | 0.44 |
| Asr2 | 190 | A | 0.74 | T | 0.26 | 0.39 |
| Asr2 | 191 | T | 0.69 | A | 0.31 | 0.43 |
| Asr2 | 192 | T | 0.69 | A | 0.31 | 0.43 |
| Asr2 | 200 | A | 0.88 | T | 0.12 | 0.21 |
| Asr2 | 246 | T | 0.88 | A | 0.12 | 0.21 |
| Asr2 | 407 | G | 0.67 | A | 0.33 | 0.44 |
| Asr2 | 455 | C | 0.74 | A | 0.26 | 0.39 |
| Asr2 | 486 | C | 0.9 | T | 0.1 | 0.17 |
| Dreb2B | 24 | C | 0.89 | G | 0.11 | 0.2 |
| Dreb2B | 33 | C | 0.64 | G | 0.36 | 0.46 |
| Dreb2B | 81 | A | 0.93 | T | 0.07 | 0.13 |
| Dreb2B | 134 | A | 0.68 | G | 0.32 | 0.43 |
| Dreb2B | 135 | C | 0.68 | T | 0.32 | 0.43 |
| Dreb2B | 136 | A | 0.68 | G | 0.32 | 0.43 |
| Dreb2B | 342 | C | 0.8 | T | 0.2 | 0.33 |
| Dreb2B | 357 | G | 0.82 | C | 0.18 | 0.3 |
| ERECTA | 32 | C | 0.85 | T | 0.15 | 0.26 |
| ERECTA | 33 | T | 0.53 | A | 0.48 | 0.5 |
| ERECTA | 46 | C | 0.8 | T | 0.2 | 0.32 |
| ERECTA | 47 | A | 0.8 | G | 0.2 | 0.32 |
| ERECTA | 55 | T | 0.93 | A | 0.08 | 0.14 |
| ERECTA | 137 | G | 0.93 | A | 0.08 | 0.14 |
| ERECTA | 172 | G | 0.93 | A | 0.08 | 0.14 |
| ERECTA | 187 | G | 0.95 | A | 0.05 | 0.1 |
| ERECTA | 189 | C | 0.93 | T | 0.08 | 0.14 |
| ERECTA | 223 | T | 0.95 | C | 0.05 | 0.1 |
| ERECTA | 228 | C | 0.93 | T | 0.08 | 0.14 |
| ERECTA | 285 | T | 0.93 | C | 0.08 | 0.14 |
| ERECTA | 286 | G | 0.93 | A | 0.08 | 0.14 |
| ERECTA | 311 | C | 0.63 | A | 0.38 | 0.47 |
| ERECTA | 449 | C | 0.93 | G | 0.08 | 0.14 |
| ERECTA | 476 | T | 0.65 | A | 0.35 | 0.46 |
| ERECTA | 480 | A | 0.93 | G | 0.08 | 0.14 |
| ERECTA | 615 | T | 0.93 | C | 0.08 | 0.14 |
| ERECTA | 637 | G | 0.9 | A | 0.1 | 0.18 |
| ERECTA | 683 | T | 0.93 | A | 0.08 | 0.14 |
| ERECTA | 725 | C | 0.63 | A | 0.38 | 0.47 |
| ERECTA | 734 | T | 0.95 | G | 0.05 | 0.1 |
| Gene Name | Taxa | Length (bp) | S | maf | He | π | θW | Tajima’s D |
|---|---|---|---|---|---|---|---|---|
| Asr2 | All | 547 | 13 | 0.29 | 0.38 | 0.017 | 0.014 | 0.873 |
| Asr2 | P. acutifolius (cultivated) | 548 | 13 | 0.21 | 0.33 | 0.009 | 0.010 | −0.816 |
| Asr2 | P. acutifolius (wild) | 548 | 9 | 0.23 | 0.36 | 0.005 | 0.005 | 0.314 |
| Asr2 | P. parvifolius | 548 | 15 | 0.17 | 0.29 | 0.008 | 0.008 | −0.079 |
| Dreb2B | All | 373 | 8 | 0.24 | 0.34 | 0.012 | 0.016 | −0.814 |
| Dreb2B | P. acutifolius (wild) | 374 | 9 | 0.18 | 0.29 | 0.007 | 0.007 | 0.128 |
| Dreb2B | P. parvifolius | 374 | 11 | 0.16 | 0.27 | 0.008 | 0.009 | −0.512 |
| ERECTA | All | 750 | 22 | 0.15 | 0.22 | 0.009 | 0.012 | −0.974 |
| ERECTA | P. acutifolius (cultivated) | 751 | 8 | 0.25 | 0.38 | 0.005 | 0.005 | 0.142 |
| ERECTA | P. acutifolius (wild) | 751 | 22 | 0.10 | 0.18 | 0.005 | 0.009 | −1.519 |
| ERECTA | P. parvifolius | 751 | 25 | 0.18 | 0.30 | 0.009 | 0.010 | −0.447 |
| SNP | GLM p-Value | MLM p-Value | GLM p-Value | MLM p-Value | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | bp | 3 m | 6 m | 12 m | 3 m | 6 m | 12 m | T 1 | T 2 | T 3 | T 4 | T 1 | T 2 | T 3 | T 4 |
| Asr | 1 | 0.421 | 0.050 | 0.183 | 0.594 | 0.456 | 0.751 | 0.467 | 0.066 | 0.969 | 0.033 | 0.594 | 0.422 | 0.505 | 0.094 |
| Dreb2B | 81 | 0.450 | 0.921 | 0.180 | 0.893 | 0.810 | 0.050 | 0.451 | 0.852 | 0.002 | 0.144 | 0.893 | 0.762 | 0.009 | 0.487 |
| Dreb2B | 134 | 0.379 | 0.672 | 0.100 | 0.297 | 0.465 | 0.400 | 0.380 | 0.597 | 0.026 | 0.144 | 0.297 | 0.488 | 0.737 | 0.464 |
| Dreb2B | 135 | 0.379 | 0.672 | 0.100 | 0.297 | 0.465 | 0.400 | 0.380 | 0.597 | 0.026 | 0.144 | 0.297 | 0.488 | 0.737 | 0.464 |
| Dreb2B | 136 | 0.379 | 0.672 | 0.100 | 0.297 | 0.465 | 0.400 | 0.380 | 0.597 | 0.026 | 0.144 | 0.297 | 0.488 | 0.737 | 0.464 |
| Dreb2B | 342 | 0.134 | 0.162 | 0.064 | 0.276 | 0.048 | 0.219 | 0.135 | 0.215 | 0.729 | 0.504 | 0.276 | 0.054 | 0.851 | 0.234 |
| ERECTA | 32 | 0.384 | 0.010 | 0.342 | 0.274 | 0.040 | 0.530 | 0.384 | 0.010 | 0.654 | 0.923 | 0.274 | 0.077 | 0.943 | 0.922 |
| ERECTA | 46 | 0.519 | 0.009 | 0.145 | 0.891 | 0.202 | 0.089 | 0.520 | 0.005 | 0.344 | 0.768 | 0.891 | 0.410 | 0.122 | 0.765 |
| ERECTA | 47 | 0.519 | 0.009 | 0.145 | 0.891 | 0.202 | 0.089 | 0.520 | 0.005 | 0.344 | 0.768 | 0.891 | 0.410 | 0.122 | 0.765 |
| ERECTA | 187 | 0.891 | 0.001 | 0.449 | 0.984 | 0.010 | 0.556 | 0.892 | 0.001 | 0.891 | 0.885 | 0.984 | 0.012 | 0.947 | 0.883 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buitrago-Bitar, M.A.; Cortés, A.J.; López-Hernández, F.; Londoño-Caicedo, J.M.; Muñoz-Florez, J.E.; Muñoz, L.C.; Blair, M.W. Allelic Diversity at Abiotic Stress Responsive Genes in Relationship to Ecological Drought Indices for Cultivated Tepary Bean, Phaseolus acutifolius A. Gray, and Its Wild Relatives. Genes 2021, 12, 556. https://doi.org/10.3390/genes12040556
Buitrago-Bitar MA, Cortés AJ, López-Hernández F, Londoño-Caicedo JM, Muñoz-Florez JE, Muñoz LC, Blair MW. Allelic Diversity at Abiotic Stress Responsive Genes in Relationship to Ecological Drought Indices for Cultivated Tepary Bean, Phaseolus acutifolius A. Gray, and Its Wild Relatives. Genes. 2021; 12(4):556. https://doi.org/10.3390/genes12040556
Chicago/Turabian StyleBuitrago-Bitar, María A., Andrés J. Cortés, Felipe López-Hernández, Jorge M. Londoño-Caicedo, Jaime E. Muñoz-Florez, L. Carmenza Muñoz, and Matthew Wohlgemuth Blair. 2021. "Allelic Diversity at Abiotic Stress Responsive Genes in Relationship to Ecological Drought Indices for Cultivated Tepary Bean, Phaseolus acutifolius A. Gray, and Its Wild Relatives" Genes 12, no. 4: 556. https://doi.org/10.3390/genes12040556
APA StyleBuitrago-Bitar, M. A., Cortés, A. J., López-Hernández, F., Londoño-Caicedo, J. M., Muñoz-Florez, J. E., Muñoz, L. C., & Blair, M. W. (2021). Allelic Diversity at Abiotic Stress Responsive Genes in Relationship to Ecological Drought Indices for Cultivated Tepary Bean, Phaseolus acutifolius A. Gray, and Its Wild Relatives. Genes, 12(4), 556. https://doi.org/10.3390/genes12040556