
Genomic Imprinting at the Porcine PLAGL1 Locus and the Orthologous Locus in the Human


Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
2.2. Sample Acquisition
2.3. Whole-Genome Bisulfite Sequencing (WGBS)
2.4. RNA Sequencing (RNA-Seq)
2.5. Mining and Processing RNA-Seq, Whole-Genome Sequencing (WGS), Variant Call Format (VCF), and ChIP-Seq Data
2.6. DMR Calling
2.7. Differential Splicing Analysis
2.8. Data Visualization
3. Results
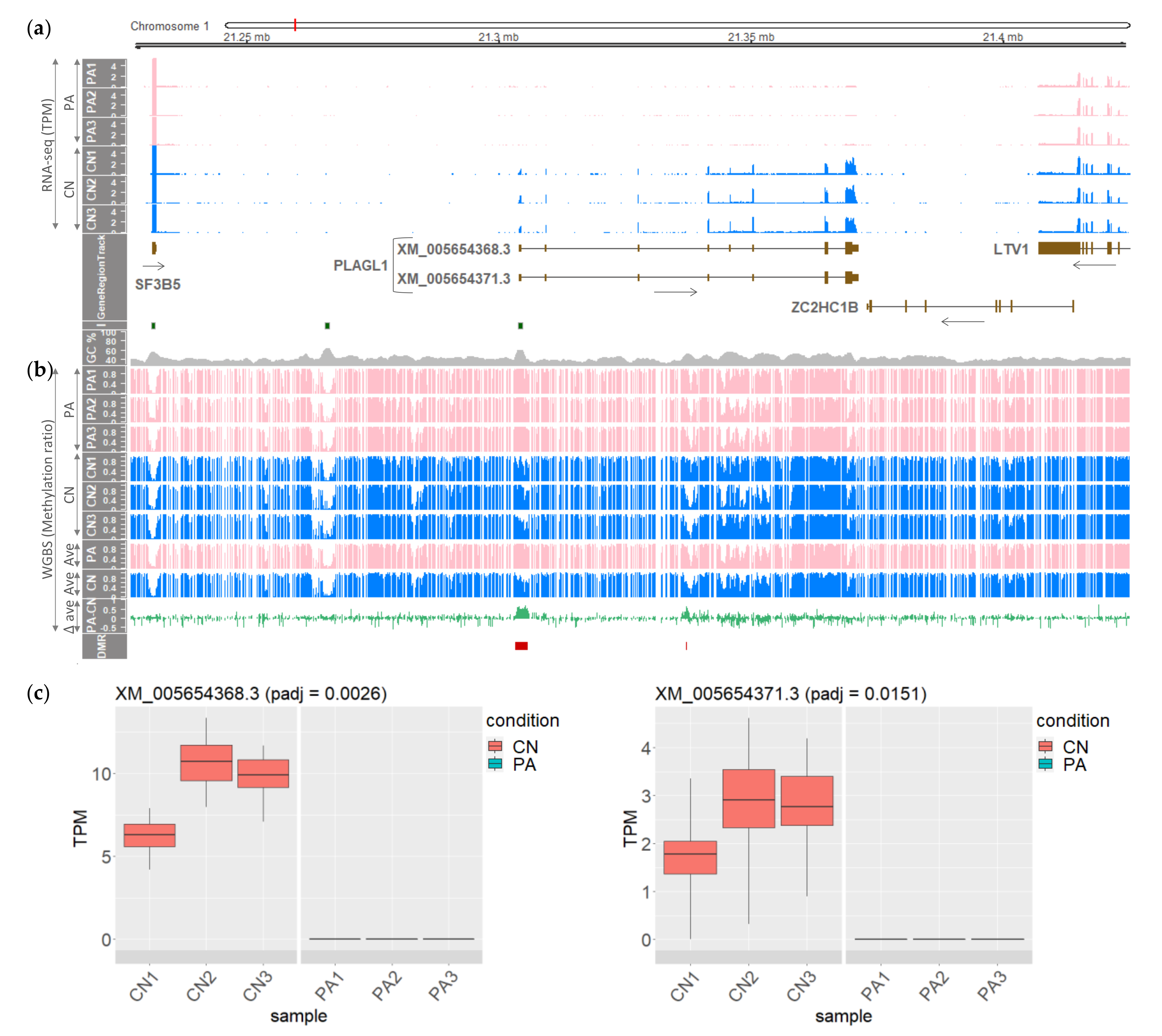
3.1. The PLAGL1 Locus in Pig Embryos Harboring Maternally Imprinted and Paternally Expressed Alternatively Spliced Transcripts
3.2. Differentially Methylated CpGs within the PLAGL1 Locus in the Porcine Genome
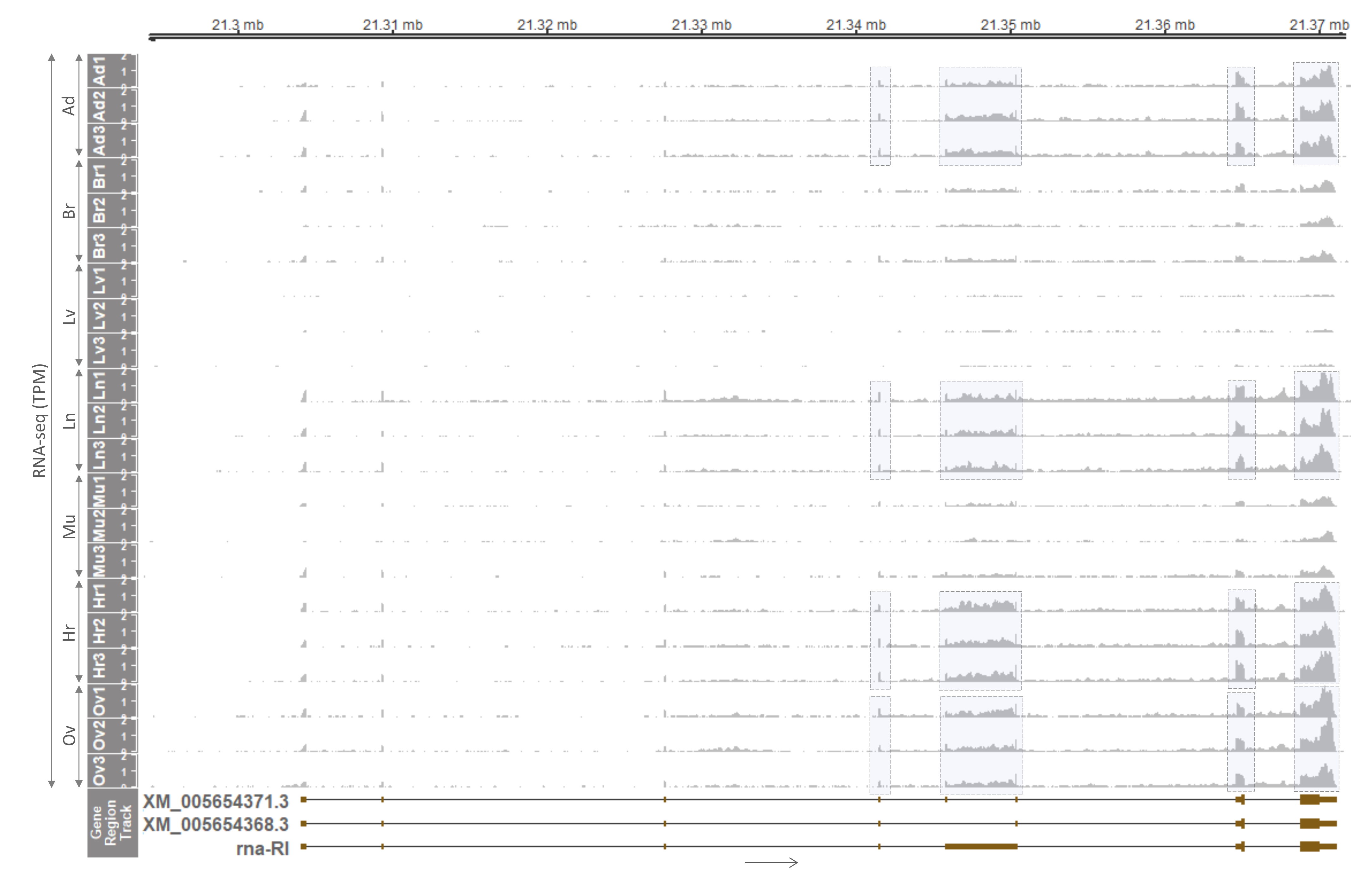
3.3. Tissue Distribution and Monoallelic Expression of the PLAGL1 Gene in Adult Pigs
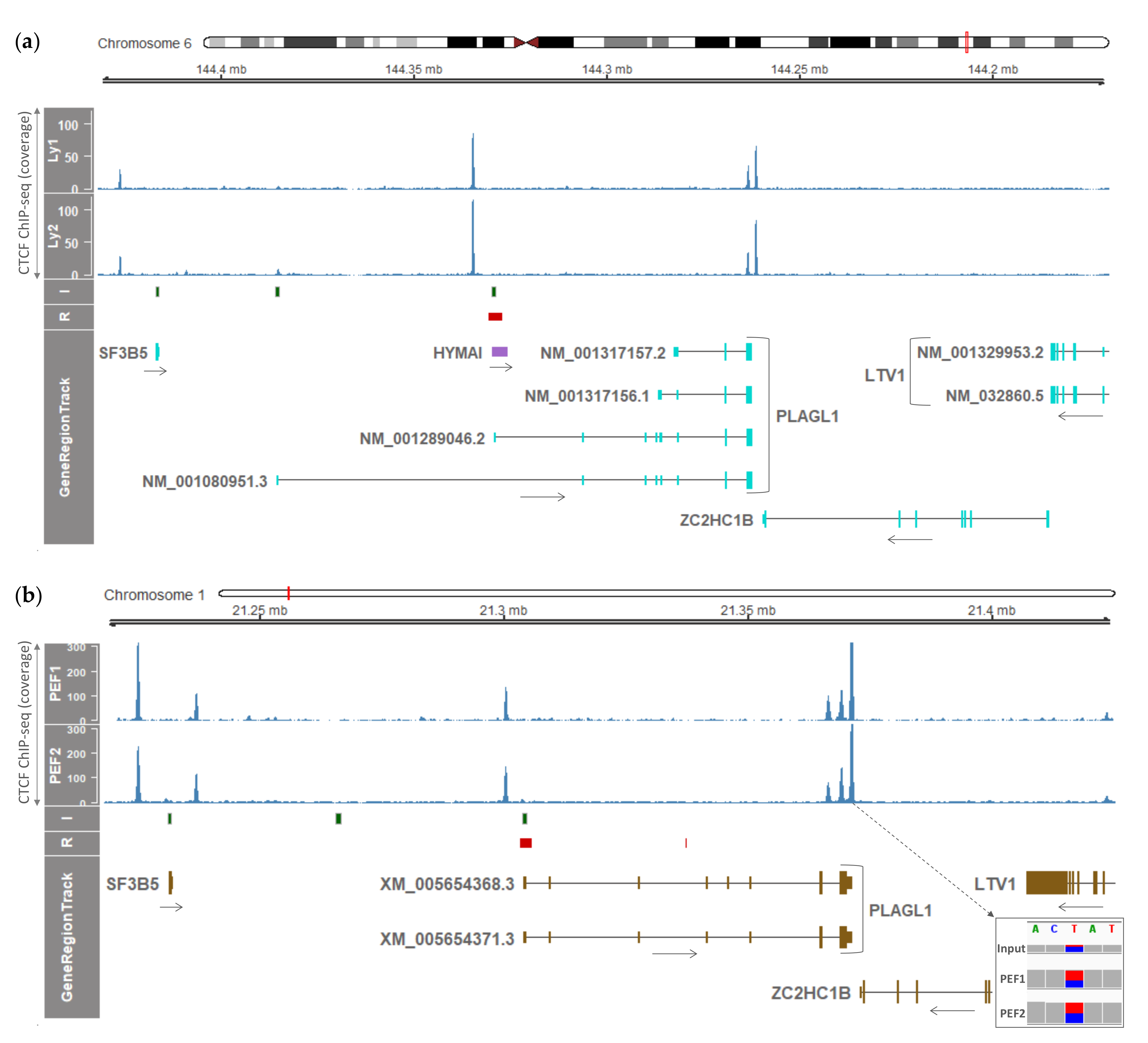
3.4. Conservation of CTCF Boundaries and Sequence Elements in Humans and Pigs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barlow, D.P.; Bartolomei, M.S. Genomic imprinting in mammals. Cold Spring Harb. Perspect. Biol. 2014, 6. [Google Scholar] [CrossRef]
- Davuluri, R.V.; Suzuki, Y.; Sugano, S.; Plass, C.; Huang, T.H. The functional consequences of alternative promoter use in mammalian genomes. Trends Genet. 2008, 24, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.M.; Ball, S.T.; Dawson, C.; Mehta, S.; Beechey, C.V.; Fray, M.; Teboul, L.; Dear, T.N.; Kelsey, G.; Peters, J. Uncoupling antisense-mediated silencing and DNA methylation in the imprinted Gnas cluster. PLoS Genet. 2011, 7, e1001347. [Google Scholar] [CrossRef] [PubMed]
- Kozasa, T.; Itoh, H.; Tsukamoto, T.; Kaziro, Y. Isolation and characterization of the human Gs alpha gene. Proc. Natl. Acad. Sci. USA 1988, 85, 2081–2085. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Wu, H.; Lee, J.; Hwang, I.S.; Yu, D.; Ahn, J.S.; Lee, J.W.; Hwang, S.; Lee, K. Identification of a Novel Imprinted Transcript in the Porcine GNAS Complex Locus Using Methylome and Transcriptome of Parthenogenetic Fetuses. Genes 2020, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Elhamamsy, A.R. Role of DNA methylation in imprinting disorders: An updated review. J. Assist. Reprod. Genet. 2017, 34, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Tucci, V.; Isles, A.R.; Kelsey, G.; Ferguson-Smith, A.C.; Erice Imprinting, G. Genomic Imprinting and Physiological Processes in Mammals. Cell 2019, 176, 952–965. [Google Scholar] [CrossRef]
- Franklin, G.C.; Adam, G.I.; Ohlsson, R. Genomic imprinting and mammalian development. Placenta 1996, 17, 3–14. [Google Scholar] [CrossRef]
- Ahn, J.; Hwang, I.S.; Park, M.R.; Cho, I.C.; Hwang, S.; Lee, K. The Landscape of Genomic Imprinting at the Porcine SGCE/PEG10 Locus from Methylome and Transcriptome of Parthenogenetic Embryos. G3 2020, 10, 4037–4047. [Google Scholar] [CrossRef]
- Kas, K.; Voz, M.L.; Hensen, K.; Meyen, E.; Van de Ven, W.J. Transcriptional activation capacity of the novel PLAG family of zinc finger proteins. J. Biol. Chem. 1998, 273, 23026–23032. [Google Scholar] [CrossRef]
- Spengler, D.; Villalba, M.; Hoffmann, A.; Pantaloni, C.; Houssami, S.; Bockaert, J.; Journot, L. Regulation of apoptosis and cell cycle arrest by Zac1, a novel zinc finger protein expressed in the pituitary gland and the brain. EMBO J. 1997, 16, 2814–2825. [Google Scholar] [CrossRef]
- Varrault, A.; Ciani, E.; Apiou, F.; Bilanges, B.; Hoffmann, A.; Pantaloni, C.; Bockaert, J.; Spengler, D.; Journot, L. hZAC encodes a zinc finger protein with antiproliferative properties and maps to a chromosomal region frequently lost in cancer. Proc. Natl. Acad. Sci. USA 1998, 95, 8835–8840. [Google Scholar] [CrossRef]
- Abdollahi, A.; Godwin, A.K.; Miller, P.D.; Getts, L.A.; Schultz, D.C.; Taguchi, T.; Testa, J.R.; Hamilton, T.C. Identification of a gene containing zinc-finger motifs based on lost expression in malignantly transformed rat ovarian surface epithelial cells. Cancer Res. 1997, 57, 2029–2034. [Google Scholar]
- Abdollahi, A.; Roberts, D.; Godwin, A.K.; Schultz, D.C.; Sonoda, G.; Testa, J.R.; Hamilton, T.C. Identification of a zinc-finger gene at 6q25: A chromosomal region implicated in development of many solid tumors. Oncogene 1997, 14, 1973–1979. [Google Scholar] [CrossRef]
- Abdollahi, A. LOT1 (ZAC1/PLAGL1) and its family members: Mechanisms and functions. J. Cell Physiol. 2007, 210, 16–25. [Google Scholar] [CrossRef]
- Huang, S.M.; Schonthal, A.H.; Stallcup, M.R. Enhancement of p53-dependent gene activation by the transcriptional coactivator Zac1. Oncogene 2001, 20, 2134–2143. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Arai, H.; Fujita, N. The novel metastasis promoter Merm1/Wbscr22 enhances tumor cell survival in the vasculature by suppressing Zac1/p53-dependent apoptosis. Cancer Res. 2011, 71, 1146–1155. [Google Scholar] [CrossRef]
- Bilanges, B.; Varrault, A.; Basyuk, E.; Rodriguez, C.; Mazumdar, A.; Pantaloni, C.; Bockaert, J.; Theillet, C.; Spengler, D.; Journot, L. Loss of expression of the candidate tumor suppressor gene ZAC in breast cancer cell lines and primary tumors. Oncogene 1999, 18, 3979–3988. [Google Scholar] [CrossRef]
- Human Protein Atlas. Available online: http://www.proteinatlas.org (accessed on 16 February 2021).
- GTExPortal. Available online: https://gtexportal.org (accessed on 16 February 2021).
- Kamiya, M.; Judson, H.; Okazaki, Y.; Kusakabe, M.; Muramatsu, M.; Takada, S.; Takagi, N.; Arima, T.; Wake, N.; Kamimura, K.; et al. The cell cycle control gene ZAC/PLAGL1 is imprinted—A strong candidate gene for transient neonatal diabetes. Hum. Mol. Genet. 2000, 9, 453–460. [Google Scholar] [CrossRef]
- Piras, G.; El Kharroubi, A.; Kozlov, S.; Escalante-Alcalde, D.; Hernandez, L.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Stewart, C.L. Zac1 (Lot1), a potential tumor suppressor gene, and the gene for epsilon-sarcoglycan are maternally imprinted genes: Identification by a subtractive screen of novel uniparental fibroblast lines. Mol. Cell Biol. 2000, 20, 3308–3315. [Google Scholar] [CrossRef]
- Iglesias-Platas, I.; Court, F.; Camprubi, C.; Sparago, A.; Guillaumet-Adkins, A.; Martin-Trujillo, A.; Riccio, A.; Moore, G.E.; Monk, D. Imprinting at the PLAGL1 domain is contained within a 70-kb CTCF/cohesin-mediated non-allelic chromatin loop. Nucleic Acids Res. 2013, 41, 2171–2179. [Google Scholar] [CrossRef]
- Ma, D.; Shield, J.P.; Dean, W.; Leclerc, I.; Knauf, C.; Burcelin, R.R.; Rutter, G.A.; Kelsey, G. Impaired glucose homeostasis in transgenic mice expressing the human transient neonatal diabetes mellitus locus, TNDM. J. Clin. Investig. 2004, 114, 339–348. [Google Scholar] [CrossRef]
- Zhang, F.W.; Cheng, H.C.; Deng, C.Y.; Xiong, Y.Z.; Li, F.E.; Lei, M.G. cDNA Cloning, Tissue Expression and Association of Porcine Pleiomorphic Adenoma Gene-like 1 (PLAGL1) Gene with Carcass Traits. Asian Australas. J. Anim. Sci. 2006, 19, 1257–1261. [Google Scholar] [CrossRef]
- Zhang, F.W.; Cheng, H.C.; Jiang, C.D.; Deng, C.Y.; Xiong, Y.Z.; Li, F.E.; Lei, M.G. Imprinted status of pleomorphic adenoma gene-like I and paternal expression gene 10 genes in pigs. J. Anim. Sci. 2007, 85, 886–890. [Google Scholar] [CrossRef]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; Hu, M.; Liu, J.S.; Ren, B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012, 485, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Li, W. BSMAP: Whole genome bisulfite sequence MAPping program. BMC Bioinform. 2009, 10, 232. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Ramirez, F.; Dundar, F.; Diehl, S.; Gruning, B.A.; Manke, T. deepTools: A flexible platform for exploring deep-sequencing data. Nucleic Acids Res. 2014, 42, W187–W191. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Yue, J.; Wei, X.; Wang, L.; Liu, X.; Gao, H.; Hou, X.; Zhao, F.; Yan, H.; et al. Genome-wide identification of RNA editing in seven porcine tissues by matched DNA and RNA high-throughput sequencing. J. Anim. Sci. Biotechnol. 2019, 10, 24. [Google Scholar] [CrossRef]
- Jühling, F.; Kretzmer, H.; Bernhart, S.H.; Otto, C.; Stadler, P.F.; Hoffmann, S. Metilene: Fast and sensitive calling of differentially methylated regions from bisulfite sequencing data. Genome Res. 2016, 26, 256–262. [Google Scholar] [CrossRef]
- Mehmood, A.; Laiho, A.; Venalainen, M.S.; McGlinchey, A.J.; Wang, N.; Elo, L.L. Systematic evaluation of differential splicing tools for RNA-seq studies. Brief. Bioinform. 2020, 21, 2052–2065. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, H.; Bray, N.L.; Puente, S.; Melsted, P.; Pachter, L. Differential analysis of RNA-seq incorporating quantification uncertainty. Nat. Methods 2017, 14, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Park, J.W.; Lu, Z.X.; Lin, L.; Henry, M.D.; Wu, Y.N.; Zhou, Q.; Xing, Y. rMATS: Robust and flexible detection of differential alternative splicing from replicate RNA-Seq data. Proc. Natl. Acad. Sci. USA 2014, 111, E5593–E5601. [Google Scholar] [CrossRef]
- Hahne, F.; Ivanek, R. Visualizing Genomic Data Using Gviz and Bioconductor. Methods Mol. Biol. 2016, 1418, 335–351. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef]
- Klett, H.; Balavarca, Y.; Toth, R.; Gigic, B.; Habermann, N.; Scherer, D.; Schrotz-King, P.; Ulrich, A.; Schirmacher, P.; Herpel, E.; et al. Robust prediction of gene regulation in colorectal cancer tissues from DNA methylation profiles. Epigenetics 2018, 13, 386–397. [Google Scholar] [CrossRef]
- Varrault, A.; Gueydan, C.; Delalbre, A.; Bellmann, A.; Houssami, S.; Aknin, C.; Severac, D.; Chotard, L.; Kahli, M.; Le Digarcher, A.; et al. Zac1 regulates an imprinted gene network critically involved in the control of embryonic growth. Dev. Cell 2006, 11, 711–722. [Google Scholar] [CrossRef]
- Varrault, A.; Dantec, C.; Le Digarcher, A.; Chotard, L.; Bilanges, B.; Parrinello, H.; Dubois, E.; Rialle, S.; Severac, D.; Bouschet, T.; et al. Identification of Plagl1/Zac1 binding sites and target genes establishes its role in the regulation of extracellular matrix genes and the imprinted gene network. Nucleic Acids Res. 2017, 45, 10466–10480. [Google Scholar] [CrossRef]
- Moore, T.; Haig, D. Genomic imprinting in mammalian development: A parental tug-of-war. Trends Genet. 1991, 7, 45–49. [Google Scholar] [CrossRef]
- Wilkins, J.F.; Haig, D. What good is genomic imprinting: The function of parent-specific gene expression. Nat. Rev. Genet. 2003, 4, 359–368. [Google Scholar] [CrossRef]
- Haig, D. Genomic imprinting and kinship: How good is the evidence? Annu. Rev. Genet. 2004, 38, 553–585. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Fung, M.R.; Barlow, D.P.; Wagner, E.F. Regulation of embryonic growth and lysosomal targeting by the imprinted Igf2/Mpr gene. Nature 1994, 372, 464–467. [Google Scholar] [CrossRef]
- Yu, Y.; Xu, F.; Peng, H.; Fang, X.; Zhao, S.; Li, Y.; Cuevas, B.; Kuo, W.L.; Gray, J.W.; Siciliano, M.; et al. NOEY2 (ARHI), an imprinted putative tumor suppressor gene in ovarian and breast carcinomas. Proc. Natl. Acad. Sci. USA 1999, 96, 214–219. [Google Scholar] [CrossRef]
- Xu, F.; Xia, W.; Luo, R.Z.; Peng, H.; Zhao, S.; Dai, J.; Long, Y.; Zou, L.; Le, W.; Liu, J.; et al. The human ARHI tumor suppressor gene inhibits lactation and growth in transgenic mice. Cancer Res. 2000, 60, 4913–4920. [Google Scholar]
- Sutton, M.N.; Lu, Z.; Li, Y.C.; Zhou, Y.; Huang, T.; Reger, A.S.; Hurwitz, A.M.; Palzkill, T.; Logsdon, C.; Liang, X.; et al. DIRAS3 (ARHI) Blocks RAS/MAPK Signaling by Binding Directly to RAS and Disrupting RAS Clusters. Cell Rep. 2019, 29, 3448–3459.e6. [Google Scholar] [CrossRef]
- Chamberlain, A.J.; Vander Jagt, C.J.; Hayes, B.J.; Khansefid, M.; Marett, L.C.; Millen, C.A.; Nguyen, T.T.; Goddard, M.E. Extensive variation between tissues in allele specific expression in an outbred mammal. BMC Genom. 2015, 16, 993. [Google Scholar] [CrossRef]
- Monk, D.; Arnaud, P.; Frost, J.; Hills, F.A.; Stanier, P.; Feil, R.; Moore, G.E. Reciprocal imprinting of human GRB10 in placental trophoblast and brain: Evolutionary conservation of reversed allelic expression. Hum. Mol. Genet. 2009, 18, 3066–3074. [Google Scholar] [CrossRef]
- Bilanges, B.; Varrault, A.; Mazumdar, A.; Pantaloni, C.; Hoffmann, A.; Bockaert, J.; Spengler, D.; Journot, L. Alternative splicing of the imprinted candidate tumor suppressor gene ZAC regulates its antiproliferative and DNA binding activities. Oncogene 2001, 20, 1246–1253. [Google Scholar] [CrossRef]
- Holwerda, S.J.; de Laat, W. CTCF: The protein, the binding partners, the binding sites and their chromatin loops. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120369. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genomic | Rs Release | Pig | gDNA (WGS) | Ad (RNA-Seq) | Ln (RNA-Seq) | Hr (RNA-Seq) | Ov (RNA-Seq) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Coordinate | rs ID | Ref | Alt | ID | Ref | Alt | Ref | Alt | Ref | Alt | Ref | Alt | Ref | Alt |
| chr1:21341446 | rs331051321 | C | T | P1 | 10 (45%) | 12 (55%) | 0 | 26 | 2 (5%) | 35 (95%) | 0 | 25 | 0 | 33 |
| P2 | 6 (38%) | 10 (63%) | 0 | 24 | 0 | 26 | 0 | 33 | 0 | 24 | ||||
| chr1:21349510 | rs331477147 | G | C | P1 | 7 (47%) | 8 (53%) | 0 | 19 | 0 | 11 | 0 | 35 | 0 | 22 |
| P2 | 10 (42%) | 14 (58%) | 0 | 11 | 0 | 11 | 0 | 30 | 0 | 20 | ||||
| P3 | 12 (32%) | 25 (68%) | 22 (100%) | 0 | 33 (100%) | 0 | 37 | 0 | 19 | 0 | ||||
| chr1:21365021 | NR | T | A | P2 | 9 (60%) | 6 (40%) | 51 (98%) | 1 (2%) | 46 (100%) | 0 | 60 | 0 | 40 | 0 |
| chr1:21368747 | rs327656939 | A | G | P3 | 8 (38%) | 13 (62%) | 37 (97%) | 1 (3%) | 48 (98%) | 1 (2%) | 52 | 0 | 63 (97%) | 2 (3%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, J.; Hwang, I.-S.; Park, M.-R.; Hwang, S.; Lee, K. Genomic Imprinting at the Porcine PLAGL1 Locus and the Orthologous Locus in the Human. Genes 2021, 12, 541. https://doi.org/10.3390/genes12040541
Ahn J, Hwang I-S, Park M-R, Hwang S, Lee K. Genomic Imprinting at the Porcine PLAGL1 Locus and the Orthologous Locus in the Human. Genes. 2021; 12(4):541. https://doi.org/10.3390/genes12040541
Chicago/Turabian StyleAhn, Jinsoo, In-Sul Hwang, Mi-Ryung Park, Seongsoo Hwang, and Kichoon Lee. 2021. "Genomic Imprinting at the Porcine PLAGL1 Locus and the Orthologous Locus in the Human" Genes 12, no. 4: 541. https://doi.org/10.3390/genes12040541
APA StyleAhn, J., Hwang, I.-S., Park, M.-R., Hwang, S., & Lee, K. (2021). Genomic Imprinting at the Porcine PLAGL1 Locus and the Orthologous Locus in the Human. Genes, 12(4), 541. https://doi.org/10.3390/genes12040541