
Single-Cell Sequencing Confirms Transcripts and VHDJH Rearrangements of Immunoglobulin Genes in Human Podocytes

 ,
,
Abstract
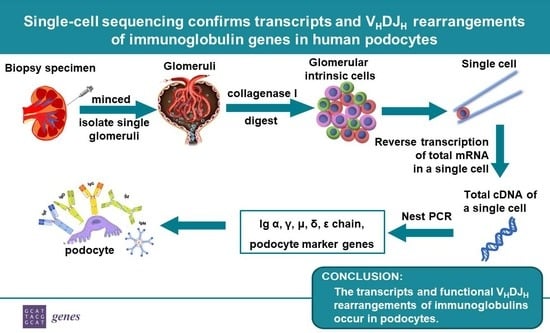
{kind=link}
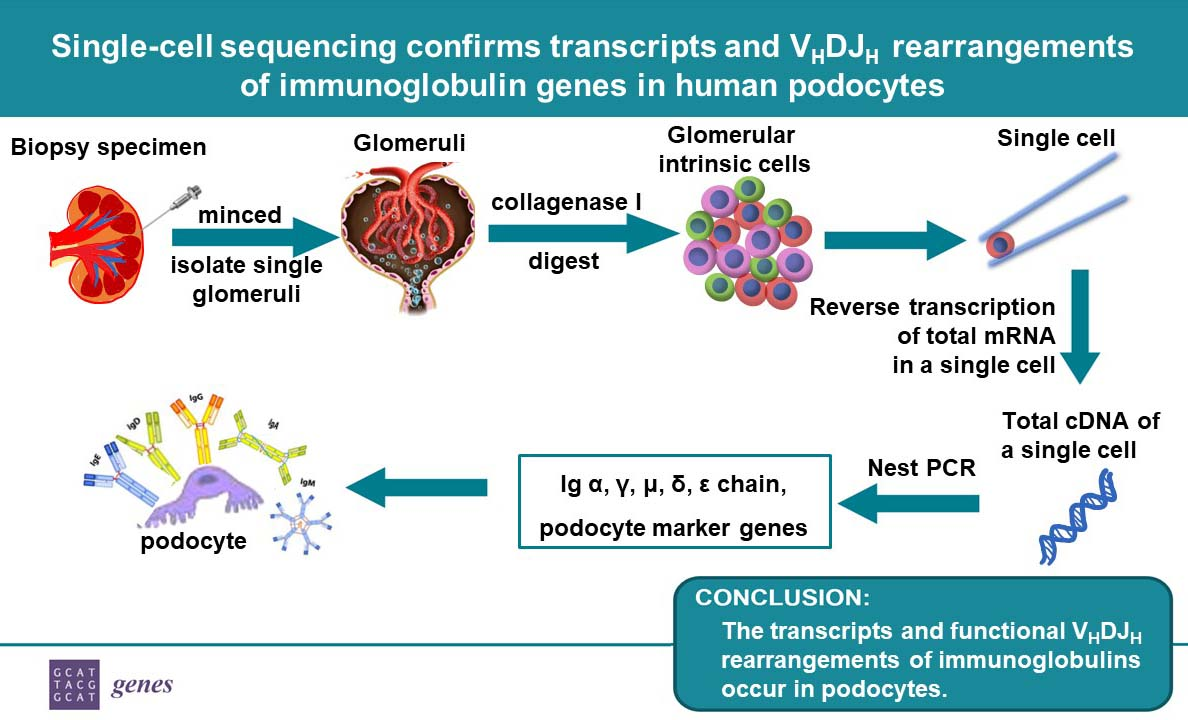
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Patients and Control Subjects
2.2. 10× Library Preparation and Sequencing
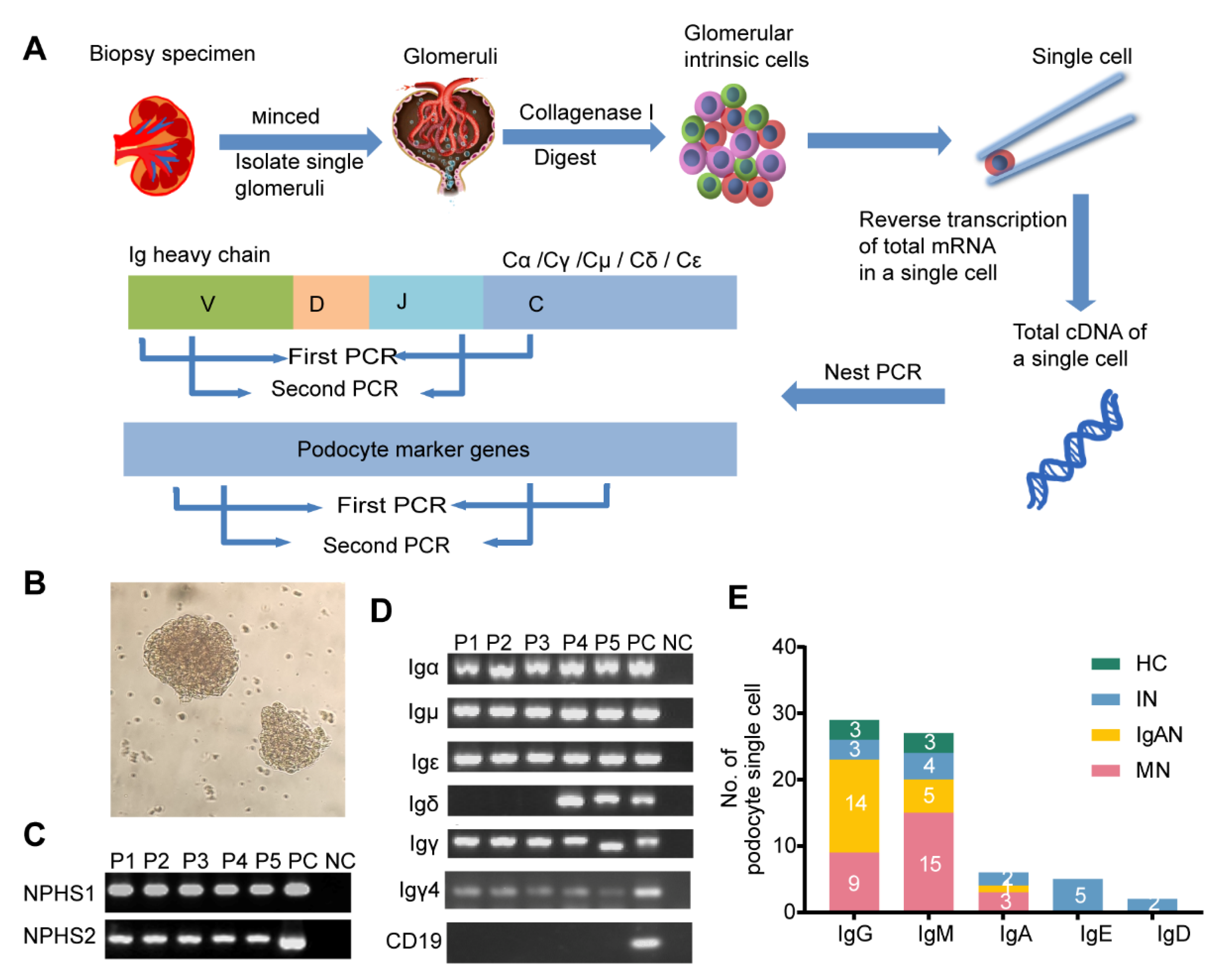
2.3. Tissue Processing and Single-Cell Dissociation
2.4. RNA Extraction and cDNA Synthesis of Single Cells
2.5. PCR Amplification
2.6. Sanger Sequencing and Analyses of the Sequencing Data
2.7. Data Mining of Single-Cell RNA Sequencing Datasets
2.8. Statistical Analysis
3. Results
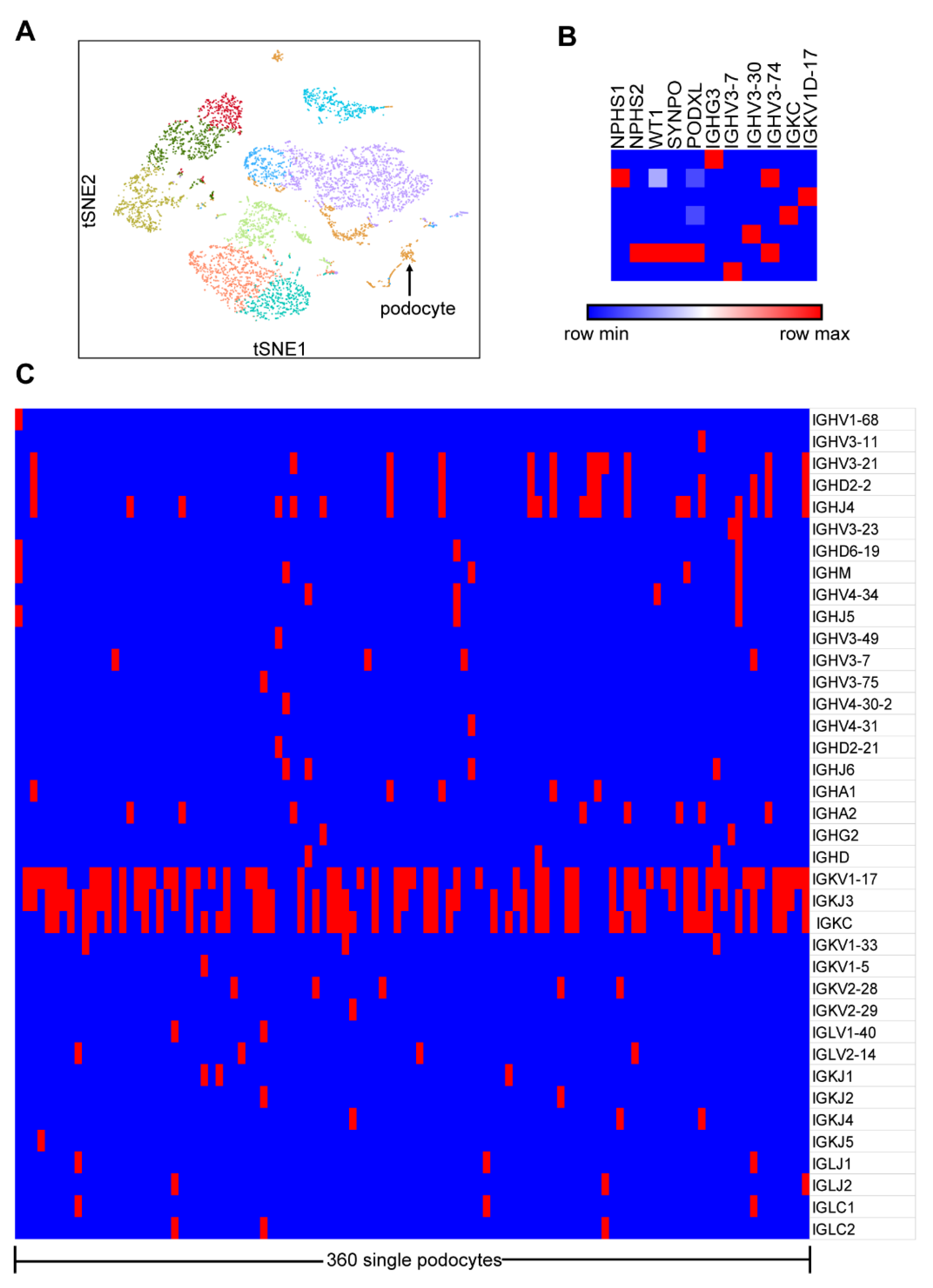
3.1. Few Ig Gene Transcripts and Lots of V, D or J Gene Segments Were Detected in Single Podocytes by the 10×Genomics Chromium System
3.2. Transcripts of Five Classes of Ig Heavy Chain Genes Were Amplified in Single Podocytes by Nested PCR and SANGER Sequencing
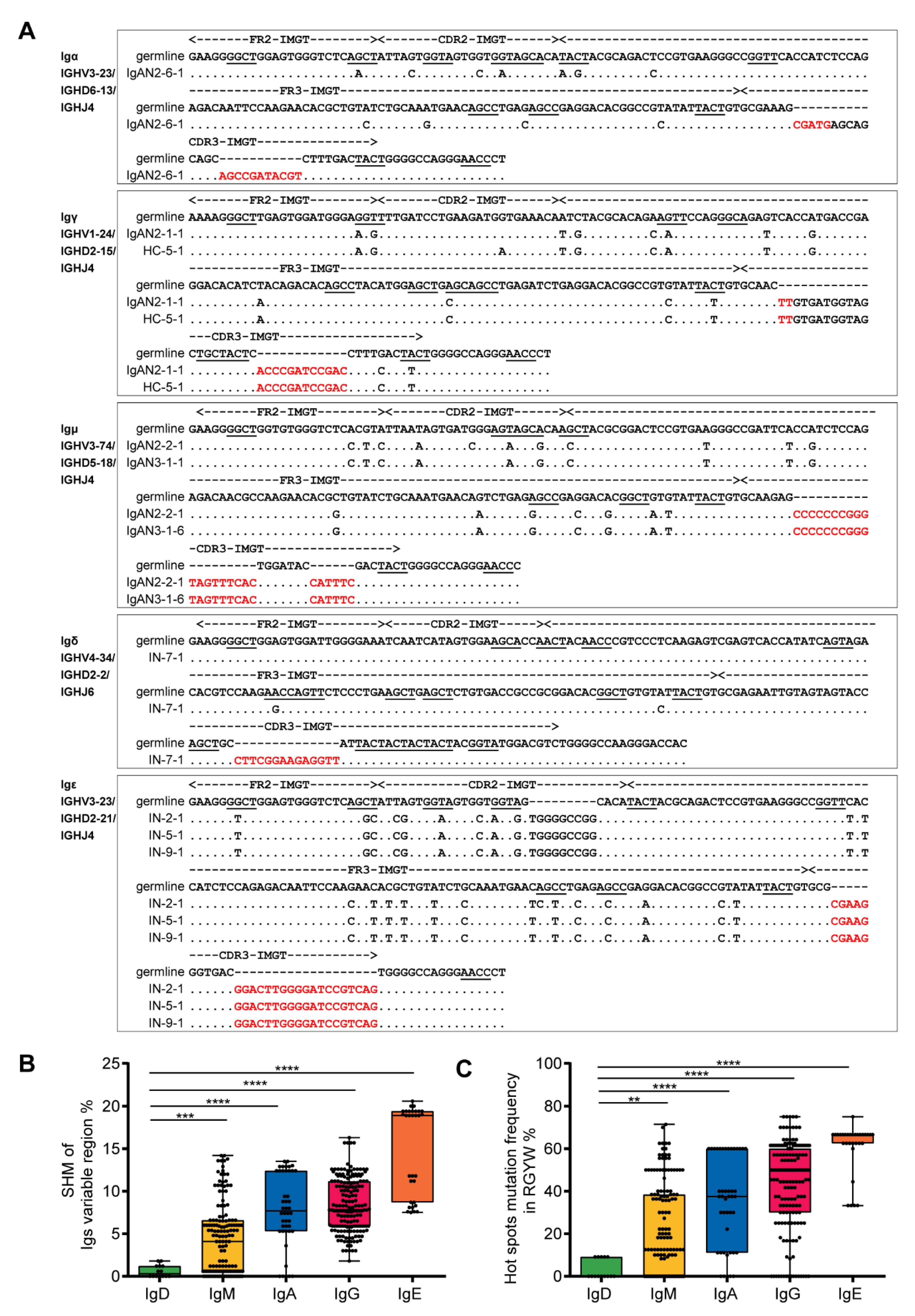
3.3. Podocyte-Derived Igs Presented Some Basic Characteristics Similar to Those of B-Igs
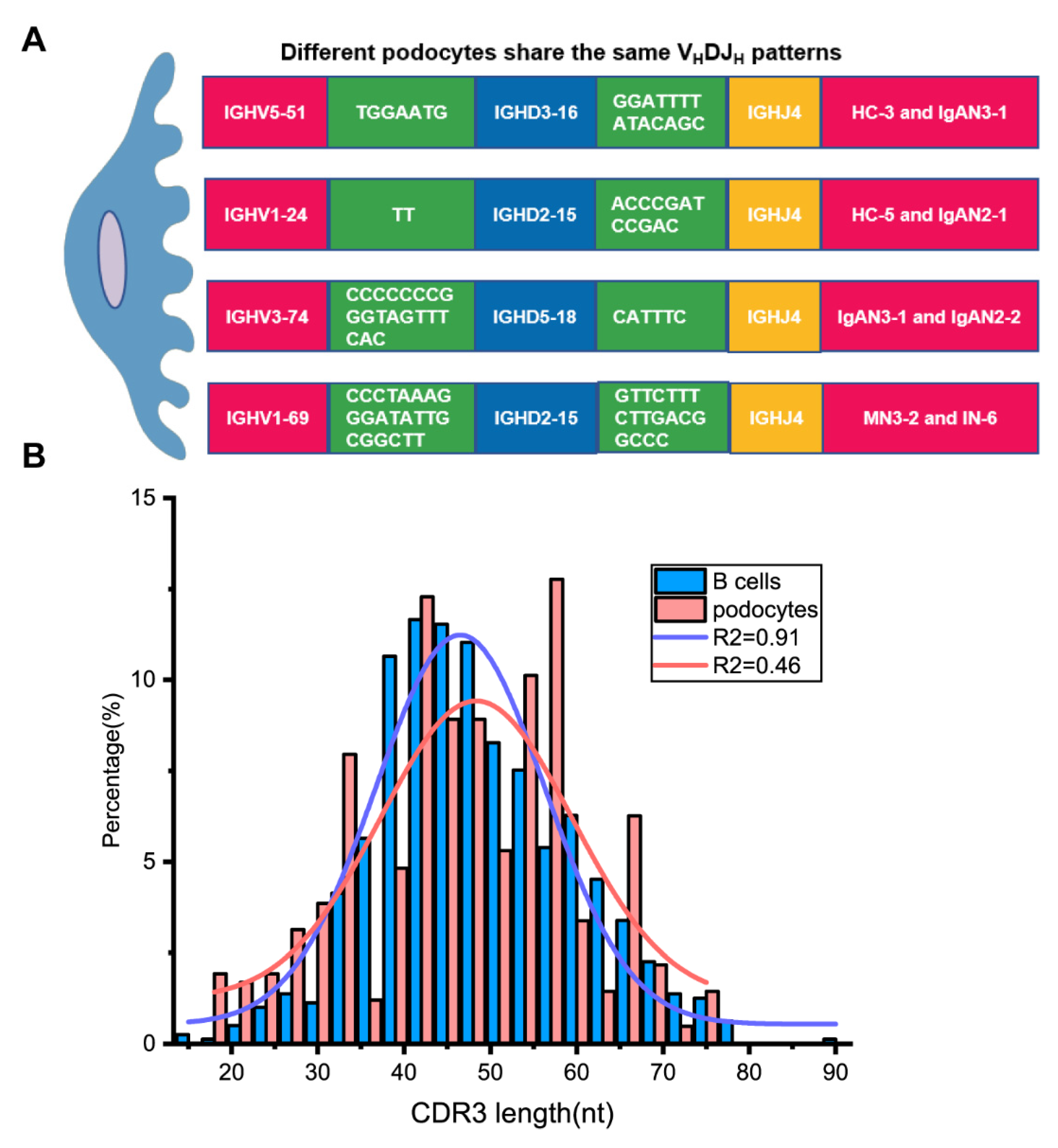
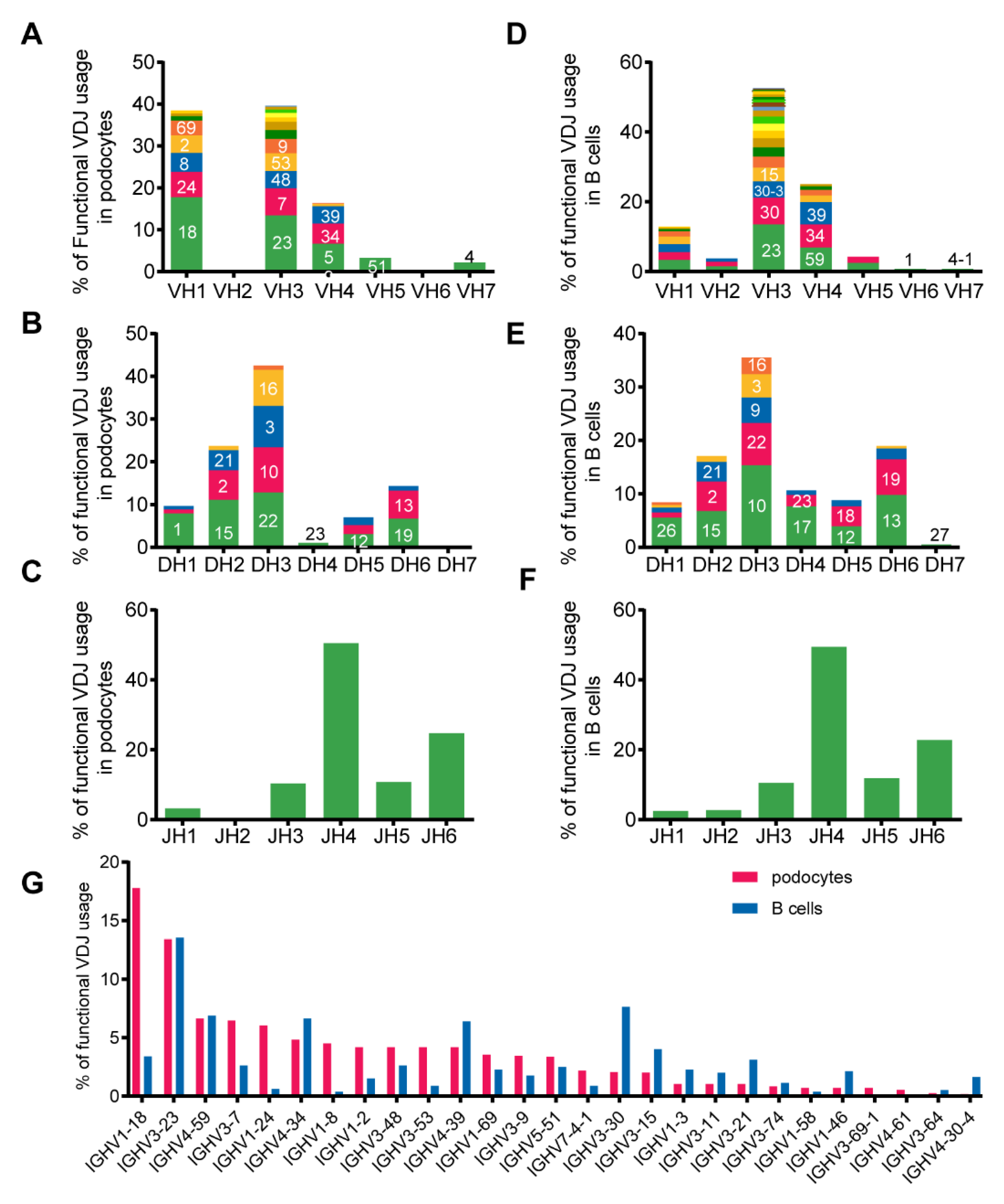
3.4. Podocyte-Derived Igs Displayed VH1 Bias and Lower Diversity Than B-Igs
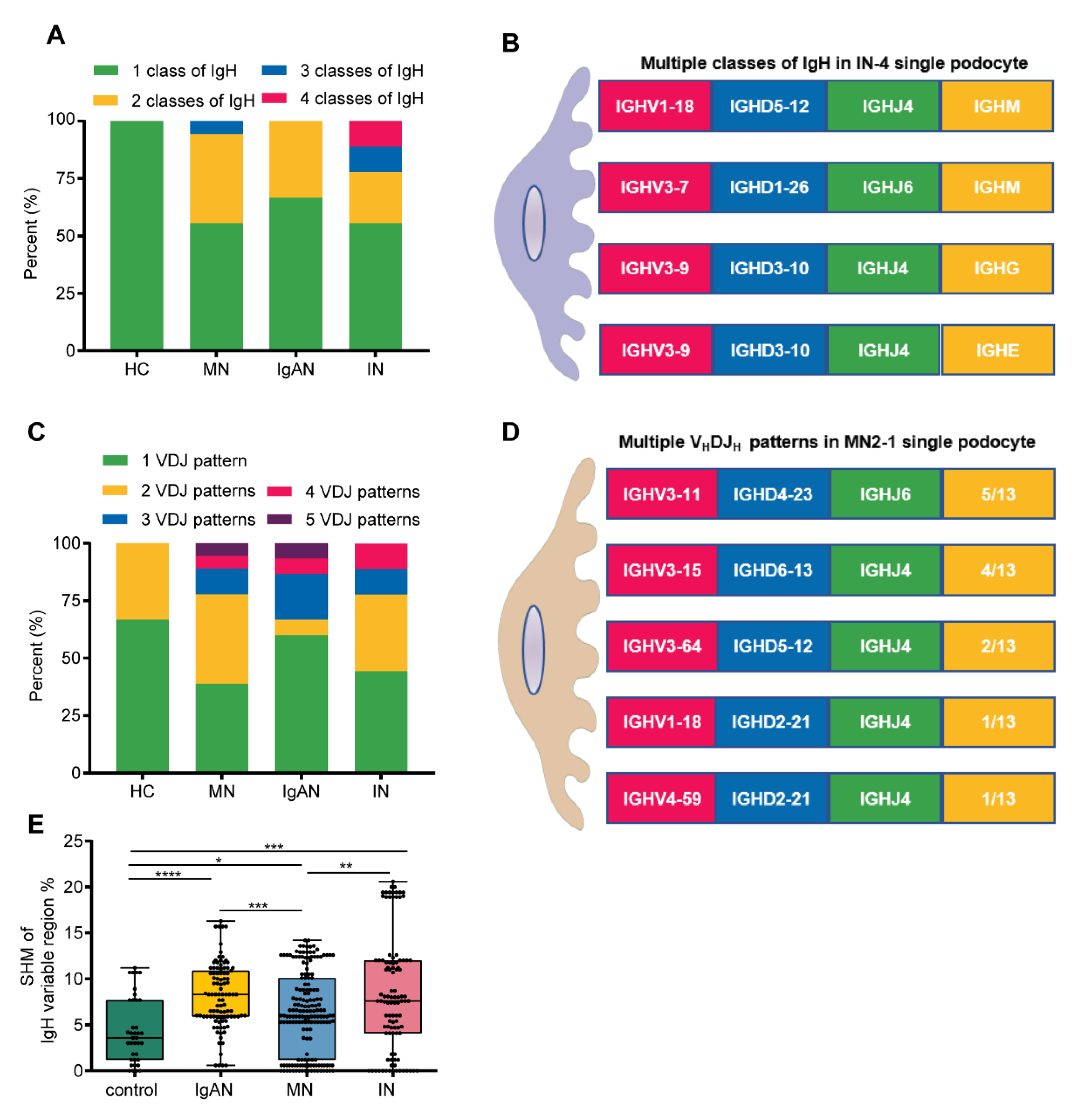
3.5. Podocytes from Kidney Patients Exhibited More Ig Classes, More VHDJH Patterns and Higher SHM
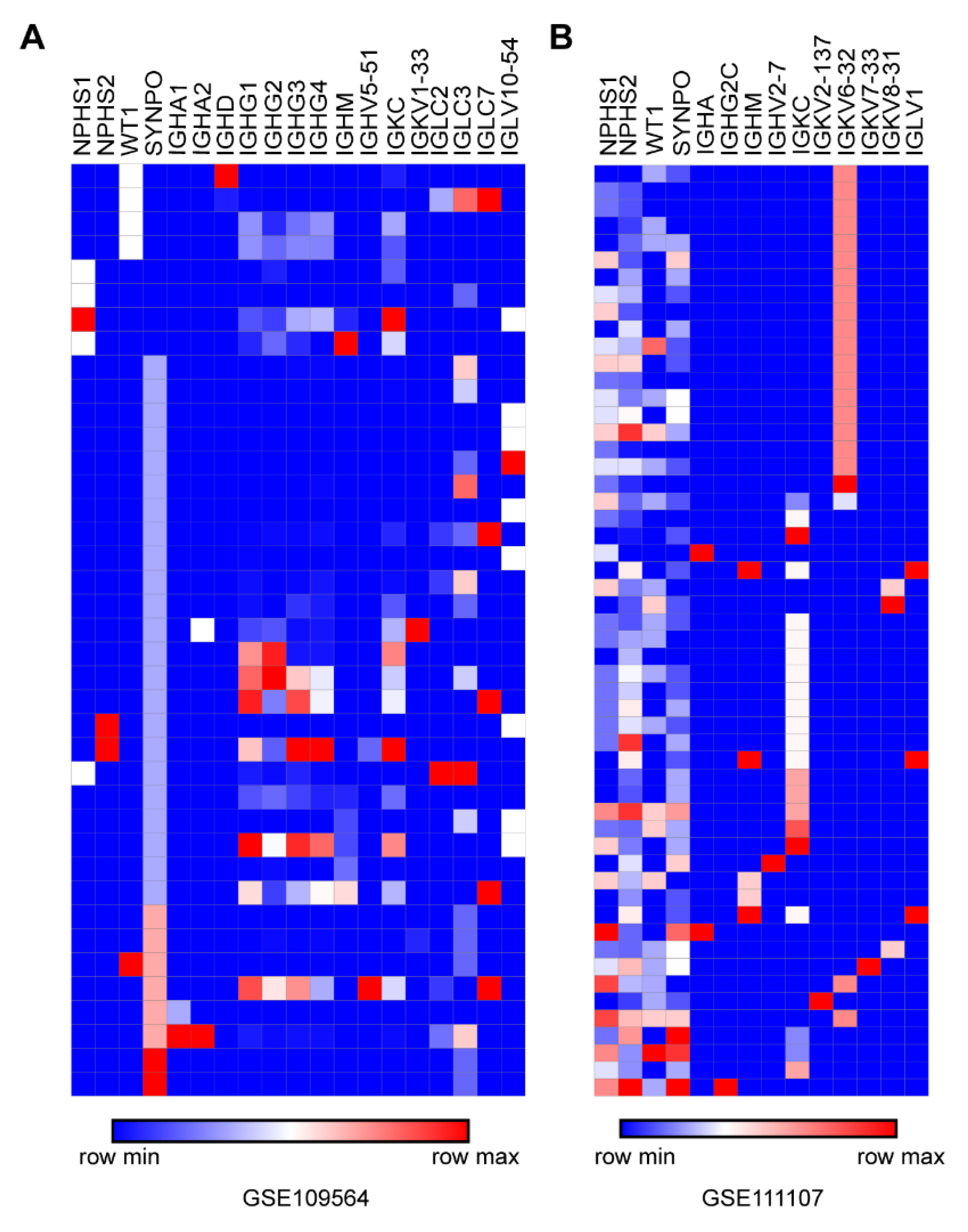
3.6. Confirmation of Ig Transcription in Podocytes by Published scRNA-seq Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhargava, R.; Tsokos, G.C. The immune podocyte. Curr. Opin. Rheumatol. 2019, 31, 167–174. [Google Scholar] [CrossRef]
- Kimoto, Y. Expression of heavy-chain constant region of immunoglobulin and T-cell receptor gene transcripts in human non-hematopoietic tumor cell lines. Genes Chromosom. Cancer 1998, 22, 83–86. [Google Scholar] [CrossRef]
- Qiu, X.Y.; Zhu, X.H.; Zhang, L.; Mao, Y.T.; Zhang, J.; Hao, P.; Li, G.H.; Lv, P.; Li, Z.X.; Sun, X.; et al. Human epithelial cancers secrete immunoglobulin g with unidentified specificity to promote growth and survival of tumor cells. Cancer Res. 2003, 63, 6488–6495. [Google Scholar]
- Chen, Z.; Gu, J. Immunoglobulin G expression in carcinomas and cancer cell lines. FASEB J. 2007, 21, 2931–2938. [Google Scholar] [CrossRef]
- Niu, N.; Zhang, J.; Sun, Y.; Wang, S.; Sun, Y.; Korteweg, C.; Gao, W.; Gu, J. Expression and distribution of immunoglobulin G and its receptors in an immune privileged site: The eye. Cell. Mol. Life Sci. 2010, 68, 2481–2492. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Sun, X.; Mao, Y.T.; Zhu, X.H.; Zhang, P.; Zhang, L.; Du, J.; Qiu, X.Y. Expression of immunoglobulin gene with classical V-(D)-J rearrangement in mouse brain neurons. Int. J. Biochem. Cell Biol. 2008, 40, 1604–1615. [Google Scholar] [CrossRef] [PubMed]
- Niu, N.; Zhang, J.; Guo, Y.; Zhao, Y.; Korteweg, C.; Gu, J. Expression and distribution of immunoglobulin G and its receptors in the human nervous system. Int. J. Biochem. Cell Biol. 2011, 43, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Korteweg, C.; Qiu, Y.; Luo, J.; Chen, Z.; Huang, G.; Li, W.; Gu, J. Two Ultrastructural Distribution Patterns of Immunoglobulin G in Human Placenta and Functional Implications1. Biol. Reprod. 2014, 91, 128. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, L.; Ma, T.; Zhang, P.; Qiu, X. Expression of Immunoglobulin Gene With Classical V-(D)-J Rearrangement in Mouse Testis and Epididymis. J. Histochem. Cytochem. 2008, 57, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Mao, Y.; Huang, J.; Ma, T.; Zhang, L.; Zhu, X.; Zheng, J.; Wu, L.; Yin, C.C.; Qiu, X. Immunoglobulin gene locus events in epithelial cells of lactating mouse mammary glands. Cell. Mol. Life Sci. 2009, 67, 985–994. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, X.; Ye, J.; Pan, P.; Cao, Q.; Yang, B.; Li, Z.; Su, M.; Huang, C.; Gu, J. Immunoglobulin G is present in a wide variety of soft tissue tumors and correlates well with proliferation markers and tumor grades. Cancer 2010, 116, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Ru, H.; Zhang, P.; Wu, H. Structural gymnastics of RAG-mediated DNA cleavage in V(D)J recombination. Curr. Opin. Struct. Biol. 2018, 53, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Ge, J.; Liao, Q.; Ma, J.; Liu, Y.; Huang, J.; Wang, C.; Xu, W.; Zheng, J.; Shao, W.; et al. IgG and IgA with Potential Microbial-Binding Activity Are Expressed by Normal Human Skin Epidermal Cells. Int. J. Mol. Sci. 2015, 16, 2574–2590. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Hu, F.; Ma, J.; Zhang, C.; Liao, Q.; Zhu, Z.; Liu, E.; Qiu, X. Epithelial cells are a source of natural IgM that contribute to innate immune responses. Int. J. Biochem. Cell Biol. 2016, 73, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Zhang, J.; Liu, Y.; Liao, Q.; Huang, J.; Geng, Z.; Xu, W.; Sheng, Z.; Lee, G.; Zhang, Y.; et al. Lung squamous cell carcinoma cells express non-canonically glycosylated IgG that activates integrin-FAK signaling. Cancer Lett. 2018, 430, 148–159. [Google Scholar] [CrossRef]
- Deng, H.; Ma, J.; Jing, Z.; Deng, Z.; Liang, Y.; Lata, A.; Liu, Y.; Qiu, X.; Wang, Y. Expression of immunoglobulin A in human mesangial cells and its effects on cell apoptosis and adhesion. Mol. Med. Rep. 2018, 17, 5272–5282. [Google Scholar] [CrossRef]
- Jing, Z.; Deng, H.; Ma, J.; Guo, Y.; Liang, Y.; Wu, R.; Lata, A.; Geng, Z.; Qiu, X.; Wang, Y. Expression of immunoglobulin G in human podocytes, and its role in cell viability and adhesion. Int. J. Mol. Med. 2018, 41, 3296–3306. [Google Scholar] [CrossRef]
- Deng, Z.; Wang, X.; Liu, Y.; Tian, X.; Deng, S.; Sun, Y.; Wang, S.; Zheng, D.; Cui, Z.; Pan, Y.; et al. Single-cell RNA sequencing confirms IgG transcription and limited diversity of VHDJH rearrangements in proximal tubular epithelial cells. Sci. Rep. 2020, 10, 19657. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zhang, Q.; Yan, H.; Yang, Y.; Wang, P.; Zhang, Y.; Deng, Z.; Yu, M.; Zhou, W.; Wang, Q.; et al. More than one antibody of individual B cells revealed by single-cell immune profiling. Cell Discov. 2019, 5, 64. [Google Scholar] [CrossRef]
- Tang, F.; Barbacioru, C.; Nordman, E.; Li, B.; Xu, N.; I Bashkirov, V.; Lao, K.; Surani, M.A. RNA-Seq analysis to capture the transcriptome landscape of a single cell. Nat. Protoc. 2010, 5, 516–535. [Google Scholar] [CrossRef]
- Van Dongen, J.J.M.; Langerak, A.; Brüggemann, M.; Evans, P.A.S.; Hummel, M.; Lavender, F.; Delabesse, E.; Davi, F.; Schuuring, E.; García-Sanz, R.; et al. Design and standardization of PCR primers and protocols for detection of clonal immunoglobulin and T-cell receptor gene recombinations in suspect lymphoproliferations: Report of the BIOMED-2 Concerted Action BMH4-CT98-3936. Leukemia 2003, 17, 2257–2317. [Google Scholar] [CrossRef] [PubMed]
- 10× Genomics. Available online: https://support.10xgenomics.com/single-cell-vdj/datasets/2.2.0/vdj_v1_hs_pbmc_b (accessed on 20 August 2019).
- Wu, H.; Malone, A.F.; Donnelly, E.L.; Kirita, Y.; Uchimura, K.; Ramakrishnan, S.M.; Gaut, J.P.; Humphreys, B.D. Single-Cell Transcriptomics of a Human Kidney Allograft Biopsy Specimen Defines a Diverse Inflammatory Response. J. Am. Soc. Nephrol. 2018, 29, 2069–2080. [Google Scholar] [CrossRef] [PubMed]
- Karaiskos, N.; Rahmatollahi, M.; Boltengagen, A.; Liu, H.; Hoehne, M.; Rinschen, M.; Schermer, B.; Benzing, T.; Rajewsky, N.; Kocks, C.; et al. A Single-Cell Transcriptome Atlas of the Mouse Glomerulus. J. Am. Soc. Nephrol. 2018, 29, 2060–2068. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Z.; Yan, H.; Shi, Z.; Tian, X.; Cui, Z.; Sun, Y.; Wang, S.; Zheng, D.; Qiu, X.; Wang, Y. Single-Cell Sequencing Confirms Transcripts and VHDJH Rearrangements of Immunoglobulin Genes in Human Podocytes. Genes 2021, 12, 472. https://doi.org/10.3390/genes12040472
Deng Z, Yan H, Shi Z, Tian X, Cui Z, Sun Y, Wang S, Zheng D, Qiu X, Wang Y. Single-Cell Sequencing Confirms Transcripts and VHDJH Rearrangements of Immunoglobulin Genes in Human Podocytes. Genes. 2021; 12(4):472. https://doi.org/10.3390/genes12040472
Chicago/Turabian StyleDeng, Zhenling, Huige Yan, Zhan Shi, Xinyu Tian, Zhuan Cui, Yingchun Sun, Song Wang, Danxia Zheng, Xiaoyan Qiu, and Yue Wang. 2021. "Single-Cell Sequencing Confirms Transcripts and VHDJH Rearrangements of Immunoglobulin Genes in Human Podocytes" Genes 12, no. 4: 472. https://doi.org/10.3390/genes12040472
APA StyleDeng, Z., Yan, H., Shi, Z., Tian, X., Cui, Z., Sun, Y., Wang, S., Zheng, D., Qiu, X., & Wang, Y. (2021). Single-Cell Sequencing Confirms Transcripts and VHDJH Rearrangements of Immunoglobulin Genes in Human Podocytes. Genes, 12(4), 472. https://doi.org/10.3390/genes12040472