
Landscape of Chimeric RNAs in Non-Cancerous Cells



Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RNA Extraction and qRT-PCR
2.3. Real-Time PCR (RT-PCR) and Sanger Sequencing
2.4. Cell Fractionation
2.5. Preparation of Peripheral Blood Buffy Coat
2.6. RNA-Seq and Bioinformatics Analysis
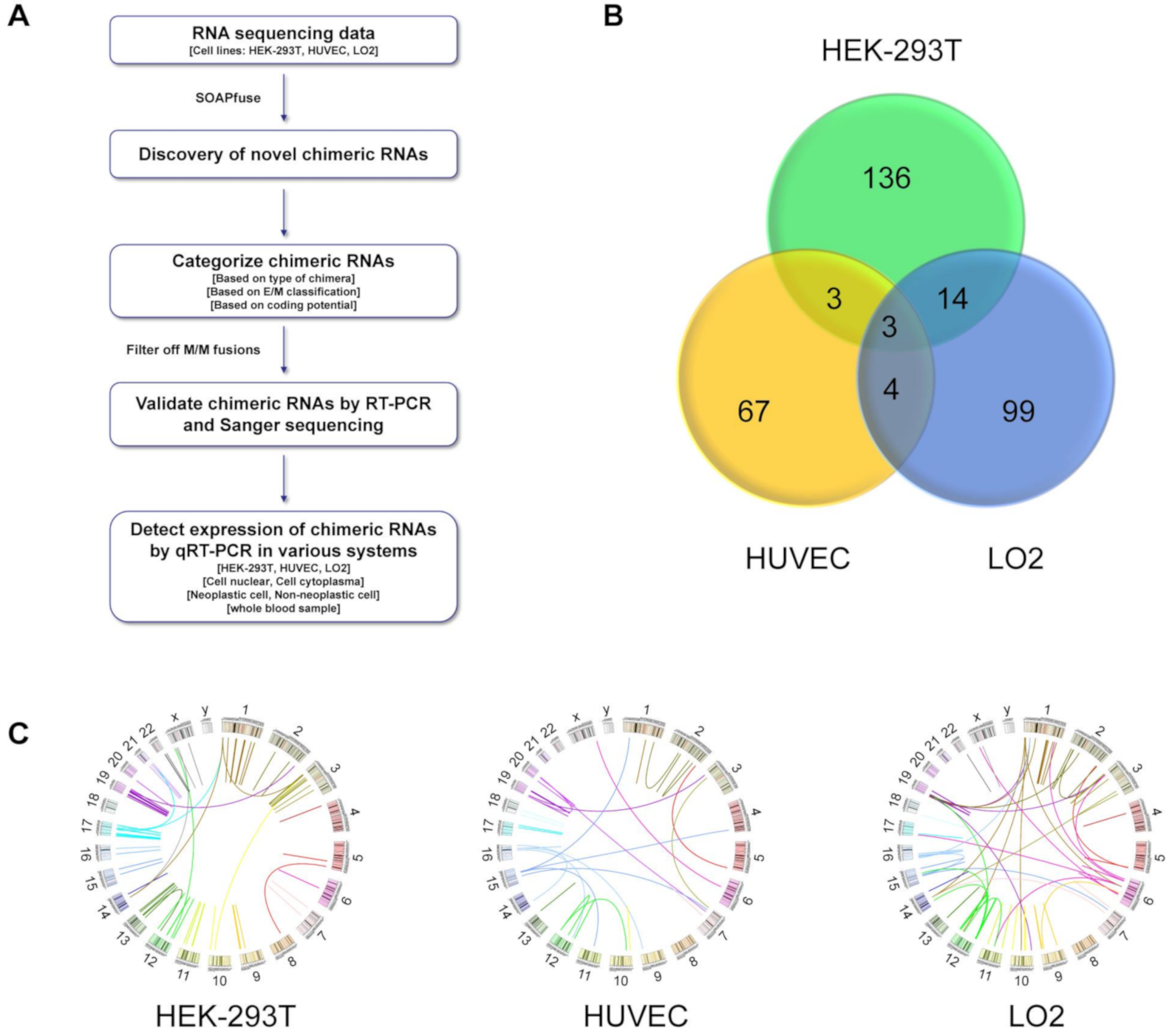
3. Results
3.1. Discovery of the Chimeric RNAs in HEK-293T, HUVEC, and LO2 Cell Lines
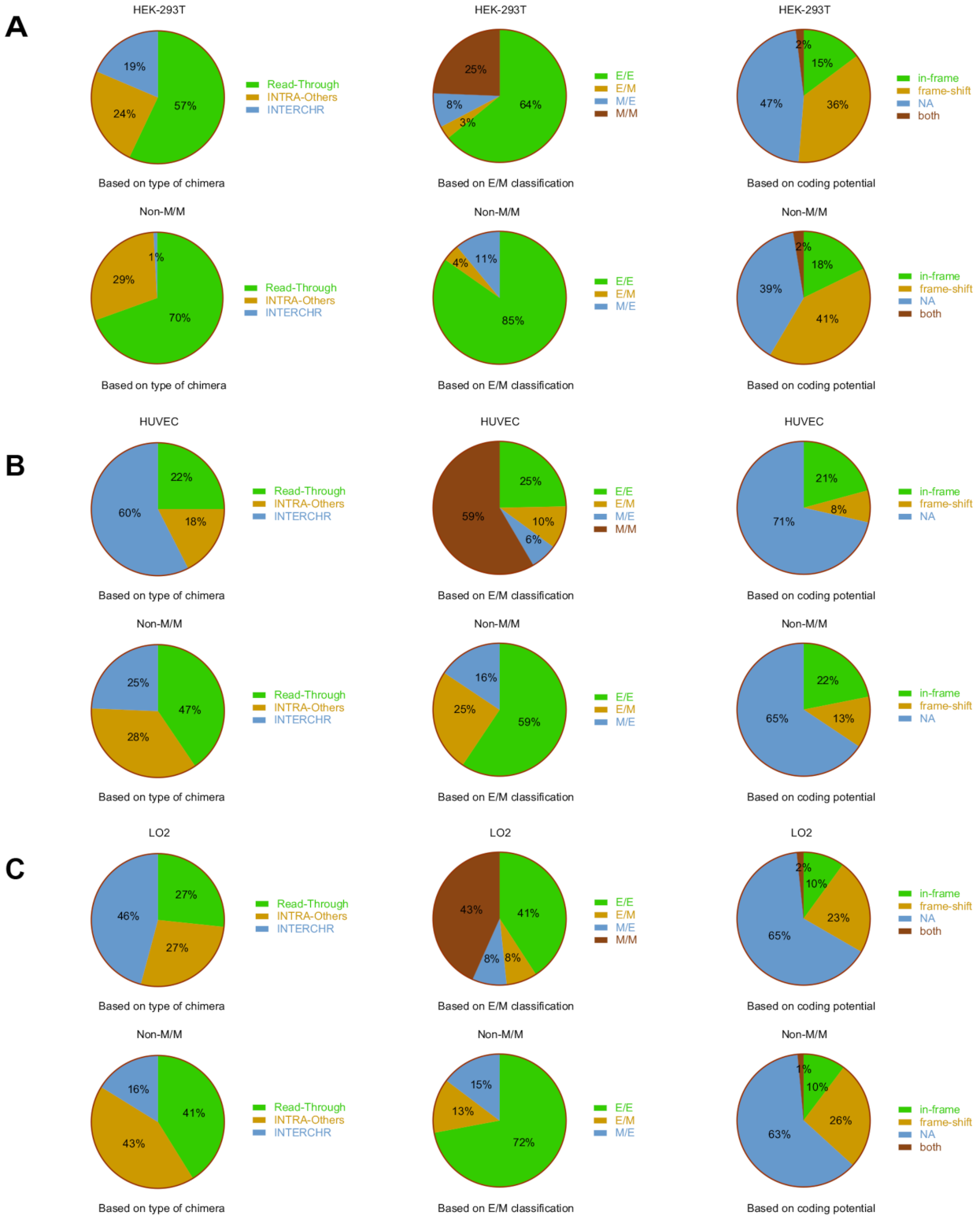
3.2. Classifications of Chimeric RNAs
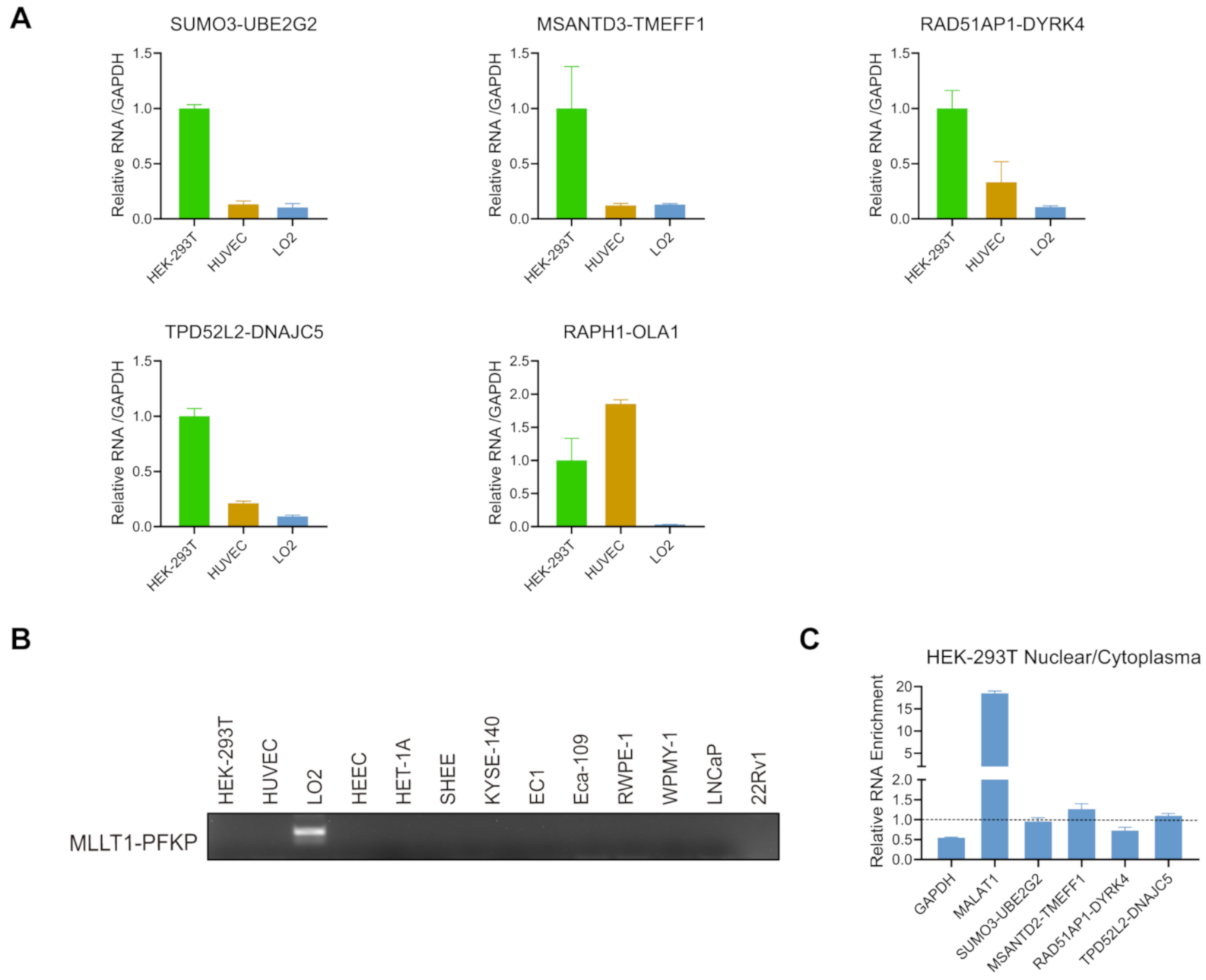
3.3. Validation of the Chimeric RNAs in HEK-293T, HUVEC, and LO2 Cell Lines
3.4. Expression of the Candidate Chimeric RNAs in HEK-293T, HUVEC, and LO2 Cell Lines
3.5. Expression of the Candidate Chimeric RNAs in Other Cells/Tissues
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asmann, Y.W.; Necela, B.M.; Kalari, K.R.; Hossain, A.; Baker, T.R.; Carr, J.M.; Davis, C.; Getz, J.E.; Hostetter, G.; Li, X.; et al. Detection of redundant fusion transcripts as biomarkers or disease-specific therapeutic targets in breast cancer. Cancer Res. 2012, 72, 1921–1928. [Google Scholar] [CrossRef] [PubMed]
- Cools, J.; DeAngelo, D.J.; Gotlib, J.; Stover, E.H.; Legare, R.D.; Cortes, J.; Kutok, J.; Clark, J.; Galinsky, I.; Griffin, J.D.; et al. A tyrosine kinase created by fusion of the PDGFRA and FIP1L1 genes as a therapeutic target of imatinib in idiopathic hypereosinophilic syndrome. N. Engl. J. Med. 2003, 348, 1201–1214. [Google Scholar] [CrossRef] [PubMed]
- Løvf, M.; Nome, T.; Bruun, J.; Eknaes, M.; Bakken, A.C.; Mpindi, J.P.; Kilpinen, S.; Rognum, T.O.; Nesbakken, A.; Kallioniemi, O.; et al. A novel transcript, VNN1-AB, as a biomarker for colorectal cancer. Int. J. Cancer 2014, 135, 2077–2084. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Yu, L.; Wang, Z. PCA3 and TMPRSS2-ERG gene fusions as diagnostic biomarkers for prostate cancer. Chin. J. Cancer Res. 2016, 28, 65–71. [Google Scholar] [PubMed]
- Rowley, J.D. A new consistent chromosomal abnormality in chronic myelogenous leukaemia identified by quinacrine fluorescence and Giemsa staining. Nature 1973, 243, 290–293. [Google Scholar] [CrossRef]
- Tomlins, S.A.; Laxman, B.; Varambally, S.; Cao, X.; Yu, J.; Helgeson, B.E.; Cao, Q.; Prensner, J.R.; Rubin, M.A.; Shah, R.B.; et al. Role of the TMPRSS2-ERG Gene Fusion in Prostate Cancer. Neoplasia 2008, 10, 177–188, IN1–IN9. [Google Scholar] [CrossRef]
- Tomlins, S.A.; Rhodes, D.R.; Perner, S.; Dhanasekaran, S.M.; Mehra, R.; Sun, X.W.; Varambally, S.; Cao, X.; Tchinda, J.; Kuefer, R.; et al. Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer. Science 2005, 310, 644–648. [Google Scholar] [CrossRef]
- Qin, F.; Song, Z.; Babiceanu, M.; Song, Y.; Facemire, L.; Singh, R.; Adli, M.; Li, H. Discovery of CTCF-sensitive Cis-spliced fusion RNAs between adjacent genes in human prostate cells. PLoS Genet. 2015, 11, e1005001. [Google Scholar]
- Zhang, Y.; Gong, M.; Yuan, H.; Park, H.G.; Frierson, H.F.; Li, H. Chimeric Transcript Generated by cis-Splicing of Adjacent Genes Regulates Prostate Cancer Cell Proliferation. Cancer Discov. 2012, 2, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Kumar-Sinha, C.; Kalyana-Sundaram, S.; Chinnaiyan, A.M. SLC45A3-ELK4 Chimera in Prostate Cancer: Spotlight on cis-Splicing: Figure 1. Cancer Discov. 2012, 2, 582–585. [Google Scholar] [CrossRef][Green Version]
- Qin, F.; Zhang, Y.; Liu, J.; Li, H. SLC45A3-ELK4 functions as a long non-coding chimeric RNA. Cancer Lett. 2017, 404, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Song, Z.; Chang, M.; Song, Y.; Frierson, H.; Li, H. Recurrent cis-SAGe chimeric RNA, D2HGDH-GAL3ST2, in prostate cancer. Cancer Lett. 2016, 380, 39–46. [Google Scholar] [CrossRef]
- Wu, P.; Yang, S.; Singh, S.; Qin, F.; Kumar, S.; Wang, L.; Ma, D.; Li, H. The Landscape and Implications of Chimeric RNAs in Cervical Cancer. EBioMedicine 2018, 37, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Singh, S.; Xie, Z.; Li, X.; Li, H. Landscape characterization of chimeric RNAs in colorectal cancer. Cancer Lett. 2020, 489, 56–65. [Google Scholar] [CrossRef]
- Wang, L.; Xiong, X.; Yao, Z.; Zhu, J.; Lin, Y.; Lin, W.; Li, K.; Xu, X.; Guo, Y.; Chen, Y.; et al. Chimeric RNA ASTN2-PAPPAas aggravates tumor progression and metastasis in human esophageal cancer. Cancer Lett. 2021, 501, 1–11. [Google Scholar] [CrossRef]
- Chase, A.; Ernst, T.; Fiebig, A.; Collins, A.; Grand, F.; Erben, P.; Reiter, A.; Schreiber, S.; Cross, N.C. TFG, a target of chromosome translocations in lymphoma and soft tissue tumors, fuses to GPR128 in healthy individuals. Haematologica 2009, 95, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-S.; Yu, C.-Y.; Chuang, C.-Y.; Hsiao, M.; Kao, C.-F.; Kuo, H.-C.; Chuang, T.-J. Integrative transcriptome sequencing identifies trans-splicing events with important roles in human embryonic stem cell pluripotency. Genome Res. 2014, 24, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Qin, F.; Movassagh, M.; Park, H.; Golden, W.L.; Xie, Z.; Zhang, P.; Sklar, J.; Li, H. A Chimeric RNA Characteristic of Rhabdomyosarcoma in Normal Myogenesis Process. Cancer Discov. 2013, 3, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Yang, S.; Zhao, W.; Tang, Z.; Zhang, T.; Li, K. Identification and analysis of pig chimeric mRNAs using RNA sequencing data. BMC Genom. 2012, 13, 429. [Google Scholar] [CrossRef]
- Babiceanu, M.; Qin, F.; Xie, Z.; Jia, Y.; Lopez, K.; Janus, N.; Facemire, L.; Kumar, S.; Pang, Y.; Qi, Y.; et al. Recurrent chimeric fusion RNAs in non-cancer tissues and cells. Nucleic Acids Res. 2016, 44, 2859–2872. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Zhang, Y.; Mao, X.; Liu, X.; Mercer, E.; Marzec, J.; Ding, D.; Jiao, Y.; Qiu, Q.; Sun, Y.; et al. Transcription-Mediated Chimeric RNAs in Prostate Cancer: Time to Revisit Old Hypothesis? OMICS J. Integr. Biol. 2014, 18, 615–624. [Google Scholar] [CrossRef]
- Yoshihara, K.; Wang, Q.; Torres-Garcia, W.; Zheng, S.; Vegesna, R.; Kim, H.; Verhaak, R.G.W. The landscape and therapeutic relevance of cancer-associated transcript fusions. Oncogene 2015, 34, 4845–4854. [Google Scholar] [CrossRef] [PubMed]
- Stransky, N.; Cerami, E.; Schalm, S.; Kim, J.L.; Lengauer, C. The landscape of kinase fusions in cancer. Nat. Commun. 2014, 5, 4846. [Google Scholar] [CrossRef]
- Finta, C.; Zaphiropoulos, P.G. Intergenic mRNA molecules resulting from trans-splicing. J. Biol. Chem. 2002, 277, 5882–5890. [Google Scholar] [CrossRef] [PubMed]
- Balamurali, D.; Gorohovski, A.; Detroja, R.; Palande, V.; Raviv-Shay, D.; Frenkel-Morgenstern, M. ChiTaRS 5.0: The compre-hensive database of chimeric transcripts matched with druggable fusions and 3D chromatin maps. Nucleic Acids Res. 2002, 48, D825–D834. [Google Scholar] [CrossRef]
- Levin, J.Z.; Berger, M.F.; Adiconis, X.; Rogov, P.; Melnikov, A.; Fennell, T.; Nusbaum, C.; Garraway, L.A.; Gnirke, A. Targeted next-generation sequencing of a cancer transcriptome enhances detection of sequence variants and novel fusion transcripts. Genome Biol. 2009, 10, R115. [Google Scholar] [CrossRef]
- Maher, C.A.; Kumar-Sinha, C.; Cao, X.; Kalyana-Sundaram, S.; Han, B.; Jing, X.; Sam, L.; Barrette, T.; Palanisamy, N.; Chinnaiyan, A.M. Transcriptome sequencing to detect gene fusions in cancer. Nat. Cell Biol. 2009, 458, 97–101. [Google Scholar] [CrossRef]
- Zhao, Q.; Caballero, O.L.; Levy, S.; Stevenson, B.J.; Iseli, C.; de Souza, S.J.; Galante, P.A.; Busam, D.; Leversha, M.A.; Chadalavada, K.; et al. Transcriptome-guided characterization of genomic re-arrangements in a breast cancer cell line. Proc. Natl. Acad. Sci. USA 2009, 106, 1886–1891. [Google Scholar] [CrossRef]
- The GTEx Consortium. The Genotype-Tissue Expression (GTEx) pilot analysis: Multitissue gene regulation in humans. Science 2015, 348, 648–660. [Google Scholar] [CrossRef] [PubMed]
- The GTEx Consortium. The Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Singh, S.; Qin, F.; Kumar, S.; Elfman, J.; Lin, E.; Pham, L.-P.; Yang, A.; Li, H. The landscape of chimeric RNAs in non-diseased tissues and cells. Nucleic Acids Res. 2020, 48, 1764–1778. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, J.; Mor, G.; Sklar, J. A Neoplastic Gene Fusion Mimics Trans-Splicing of RNAs in Normal Human Cells. Science 2008, 321, 1357–1361. [Google Scholar] [CrossRef]
- Jia, W.; Qiu, K.; He, M.; Song, P.; Zhou, Q.; Zhou, F.; Yu, Y.; Zhu, D.; Nickerson, M.L.; Wan, S.; et al. SOAPfuse: An algorithm for identifying fusion transcripts from paired-end RNA-Seq data. Genome Biol. 2013, 14, R12. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Singh, S.; Chen, X.; Zheng, Z.; Huang, J.; Lin, T.; Li, H. The landscape of chimeric RNAs in bladder urothelial carcinoma. Int. J. Biochem. Cell Biol. 2019, 110, 50–58. [Google Scholar] [CrossRef]
- Brown, C.J.; Hendrich, B.D.; Rupert, J.L.; Lafrenière, R.G.; Xing, Y.; Lawrence, J.; Willard, H.F. The human XIST gene: Analysis of a 17 kb inactive X-specific RNA that contains conserved repeats and is highly localized within the nucleus. Cell 1992, 71, 527–542. [Google Scholar] [CrossRef]
- Hutchinson, J.N.; Ensminger, A.W.; Clemson, C.M.; Lynch, C.R.; Lawrence, J.B.; Chess, A. A screen for nuclear transcripts identifies two linked noncoding RNAs associated with SC35 splicing domains. BMC Genom. 2007, 8, 39. [Google Scholar] [CrossRef]
- Sone, M.; Hayashi, T.; Tarui, H.; Agata, K.; Takeichi, M.; Nakagawa, S. The mRNA-like noncoding RNA Gomafu constitutes a novel nuclear domain in a subset of neurons. J. Cell Sci. 2007, 120, 2498–2506. [Google Scholar] [CrossRef] [PubMed]
- Teetson, W.; Cartwright, C.; Dreiling, B.J.; Steinberg, M.H. The Leukocyte Composition of Peripheral Blood Buffy Coat. Am. J. Clin. Pathol. 1983, 79, 500–501. [Google Scholar] [CrossRef]
- Druker, B.J. Current treatment approaches for chronic myelogenous leukemia. Cancer J. 2001, 7, 14–18. [Google Scholar]
- Shaw, A.T.; Ou, S.-H.I.; Bang, Y.-J.; Camidge, D.R.; Solomon, B.J.; Salgia, R.; Riely, G.J.; Varella-Garcia, M.; Shapiro, G.I.; Costa, D.B.; et al. Crizotinib in ROS1-Rearranged Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2014, 371, 1963–1971. [Google Scholar] [CrossRef]
- Jividen, K.; Li, H. Chimeric RNAs generated by intergenic splicing in normal and cancer cells. Genes Chromosom. Cancer 2014, 53, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Chwalenia, K.; Qin, F.; Singh, S.; Li, H. A cell-based splicing reporter system to identify regulators of cis-splicing between adjacent genes. Nucleic Acids Res. 2019, 47, e24. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Tsai, W.-H.; Ding, Y.; Chen, R.; Fang, Z.; Huo, Z.; Kim, S.; Ma, T.; Chang, T.-Y.; Priedigkeit, N.M.; et al. Comprehensive evaluation of fusion transcript detection algorithms and a meta-caller to combine top performing methods in paired-end RNA-seq data. Nucleic Acids Res. 2016, 44, e47. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fusion Genes | Sanger Sequencing | Neighboring Genes | Interchromosomal |
|---|---|---|---|
| ARL16-OXLD1 | N | Y | N |
| RAD51AP1-DYRK4-1 * | Y | Y | N |
| D2HGDH | N | Y | N |
| SUMO3-UBE2G2 * | Y | Y | N |
| DMKN-KRTDAP * | Y | Y | N |
| DPM2-PIP5KL1 | N | Y | N |
| MORF4L2-AS1-TMEM31 * | Y | Y | N |
| APEH-RNF123 | N | Y | N |
| MFGE8-HAPLN3 | N | Y | N |
| TBC1D23-NIT2 * | Y | Y | N |
| ZHX1-C8orf76 * | Y | Y | Y |
| TPD52L2-DNAJC5 * | Y | N | N |
| TBC1D24-ATP6VOC * | Y | Y | N |
| MSANTD3-TMEFF1 * | Y | Y | Y |
| SMOX-LINC012433 * | Y | Y | N |
| CLTC-VMP1 * | Y | N | N |
| RAPH1-OLA1-1 * | Y | N | N |
| RAPH1-OLA1-2 * | Y | N | N |
| RAD51AP1-DYRK4-2 * | Y | Y | Y |
| TLCD-RWDD3 | N | Y | N |
| FARSA-SUCE2 * | Y | Y | N |
| VAMP1-CD27-AS1 | N | Y | N |
| ZNF674-AS1-CHST7 | N | Y | N |
| SUMF1-BHLHE40-AS1 * | Y | N | N |
| TPCN2-SMIM38 | N | N | N |
| UBXN2A-MFSD2B | N | N | N |
| MLLT1-PFKP * | Y | N | Y |
| ASB16-AS1-PHF13 | N | N | Y |
| POLDIP2-TNRC18 | N | N | Y |
| CCDC32-CBX3 | N | N | Y |
| BMERB1-CHP1 | N | N | Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Haddox, S.; Tang, Y.; Qin, F.; Li, H. Landscape of Chimeric RNAs in Non-Cancerous Cells. Genes 2021, 12, 466. https://doi.org/10.3390/genes12040466
Chen C, Haddox S, Tang Y, Qin F, Li H. Landscape of Chimeric RNAs in Non-Cancerous Cells. Genes. 2021; 12(4):466. https://doi.org/10.3390/genes12040466
Chicago/Turabian StyleChen, Chen, Samuel Haddox, Yue Tang, Fujun Qin, and Hui Li. 2021. "Landscape of Chimeric RNAs in Non-Cancerous Cells" Genes 12, no. 4: 466. https://doi.org/10.3390/genes12040466
APA StyleChen, C., Haddox, S., Tang, Y., Qin, F., & Li, H. (2021). Landscape of Chimeric RNAs in Non-Cancerous Cells. Genes, 12(4), 466. https://doi.org/10.3390/genes12040466