Network Analysis of Gene Transcriptions of Arabidopsis thaliana in Spaceflight Microgravity

 , ,
, ,  ,
,
Abstract
1. Introduction
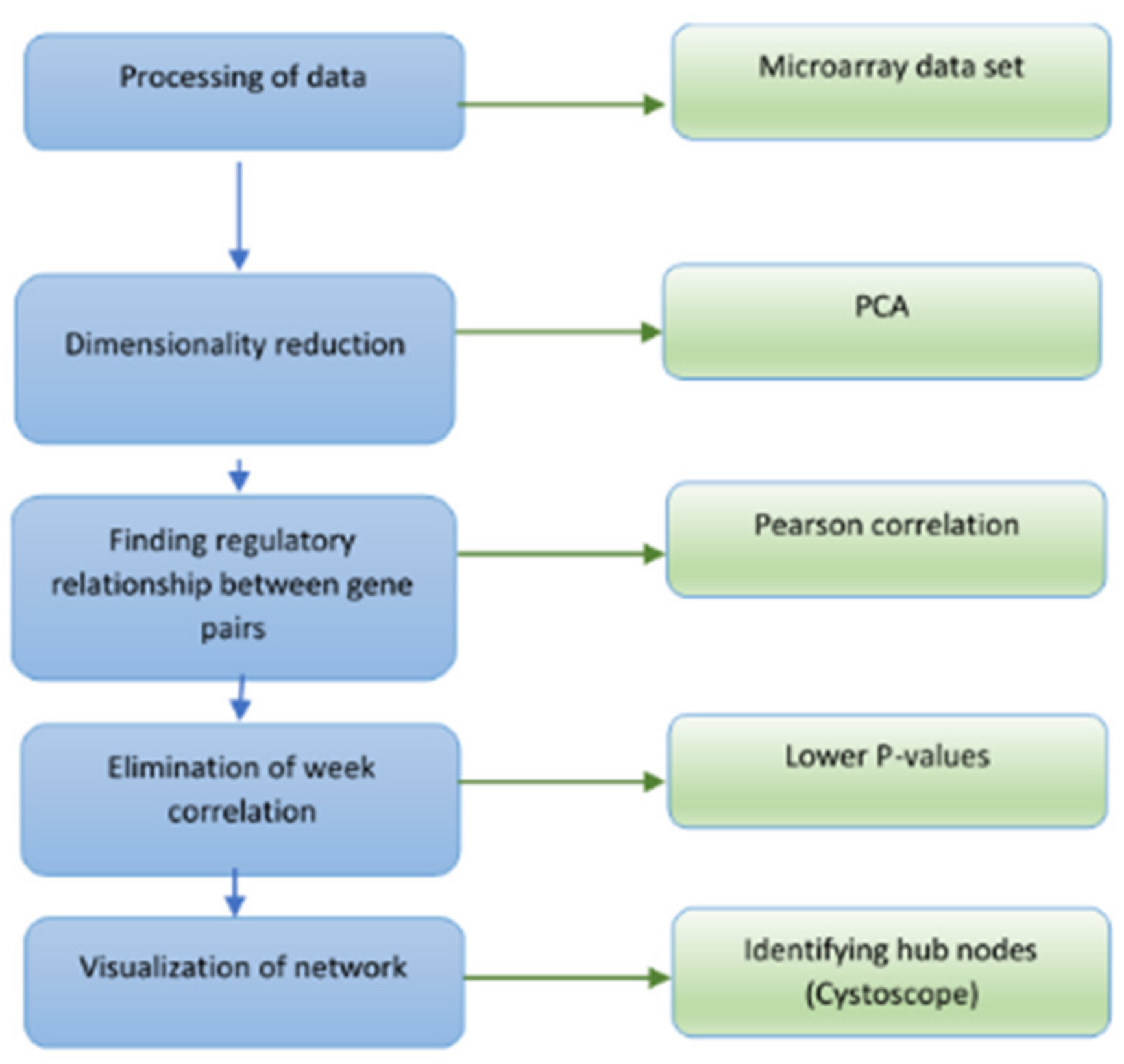
2. Materials and Methods
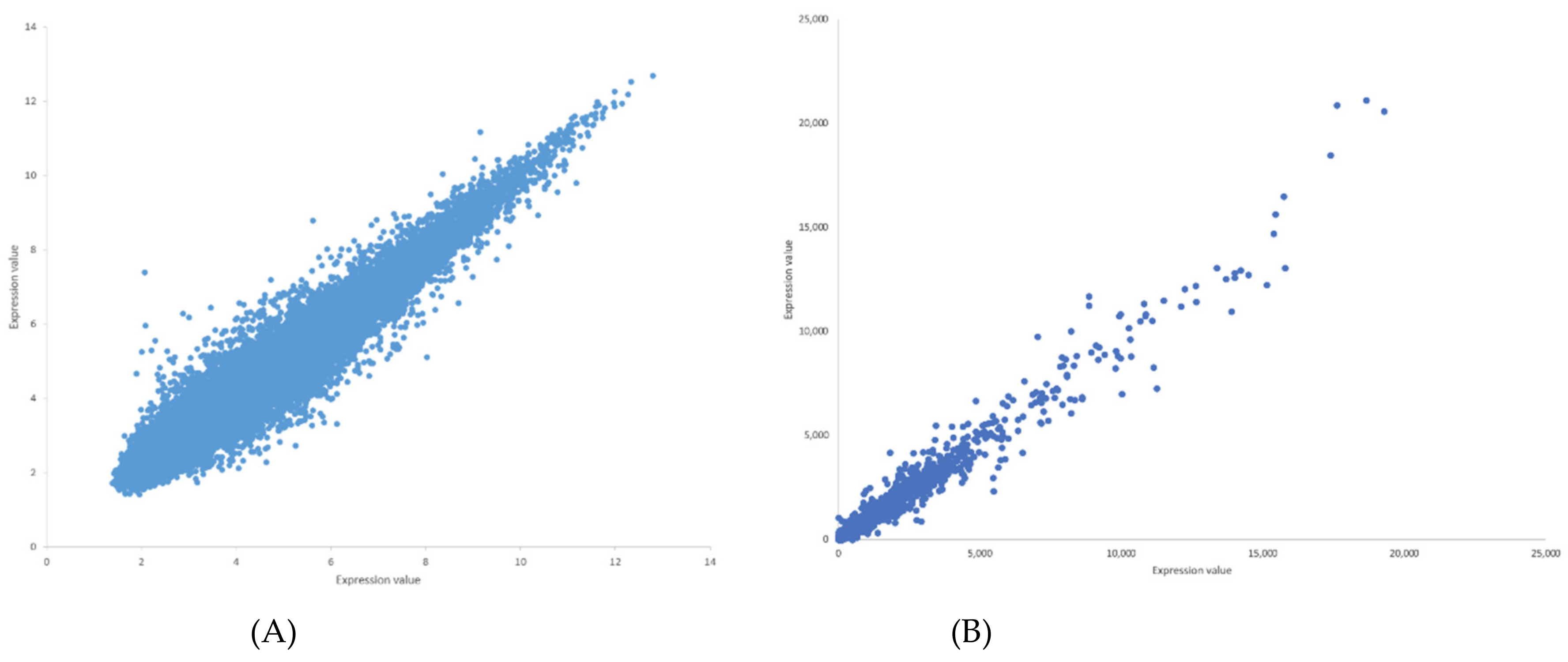
2.1. GeneLab Arabidopsis Datasets
2.2. Graph-Based GRN Inferencing
2.2.1. LASSO Regression
2.2.2. Pearson Correlation
2.2.3. Sparse Networks and the Weighted HITS Algorithm
3. Results and Discussion
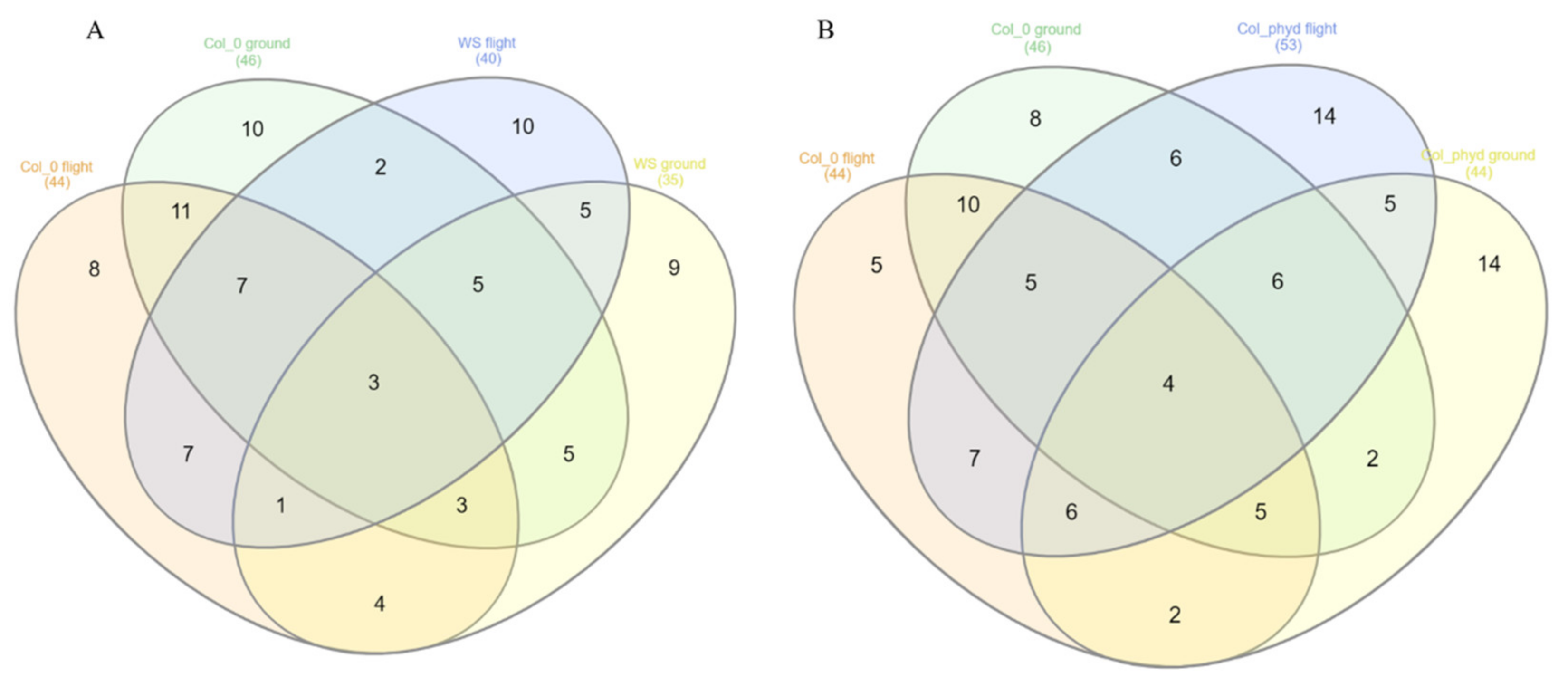
3.1. Gene Set Enrichment Analysis (GSEA)
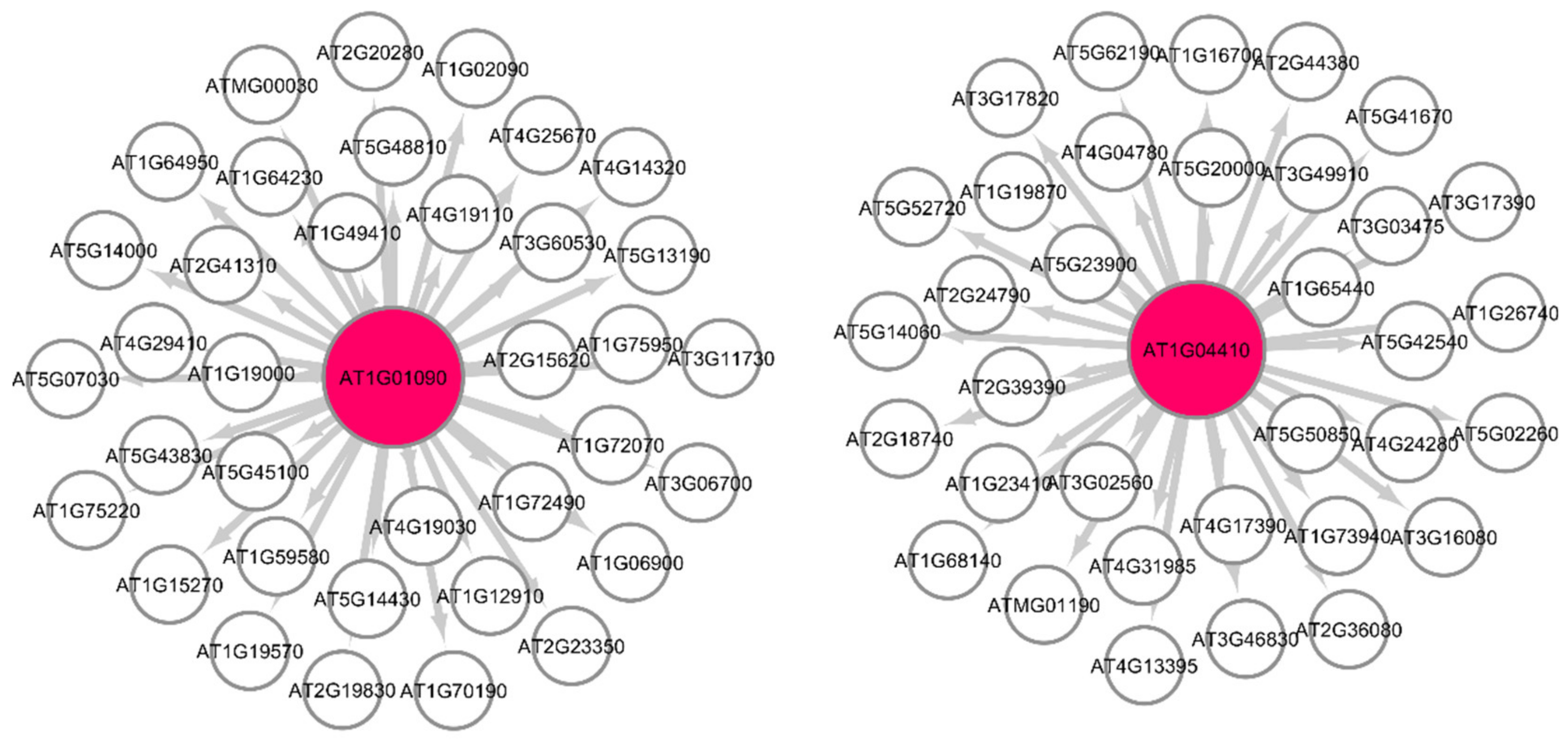
3.1.1. GSEA for Common Hub Genes
3.1.2. Gene Ontology of Transcription Factors (Genes) in Spaceflight
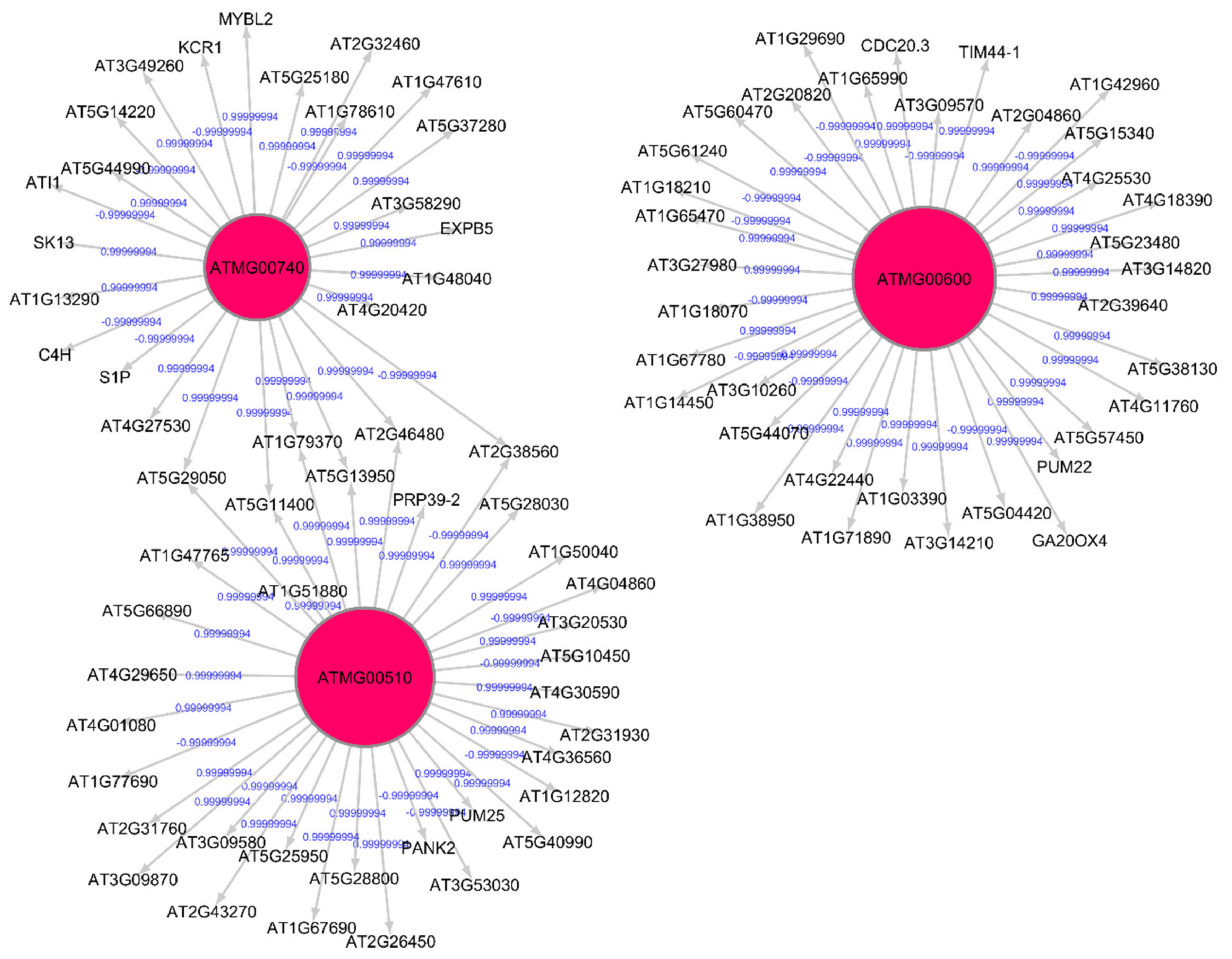
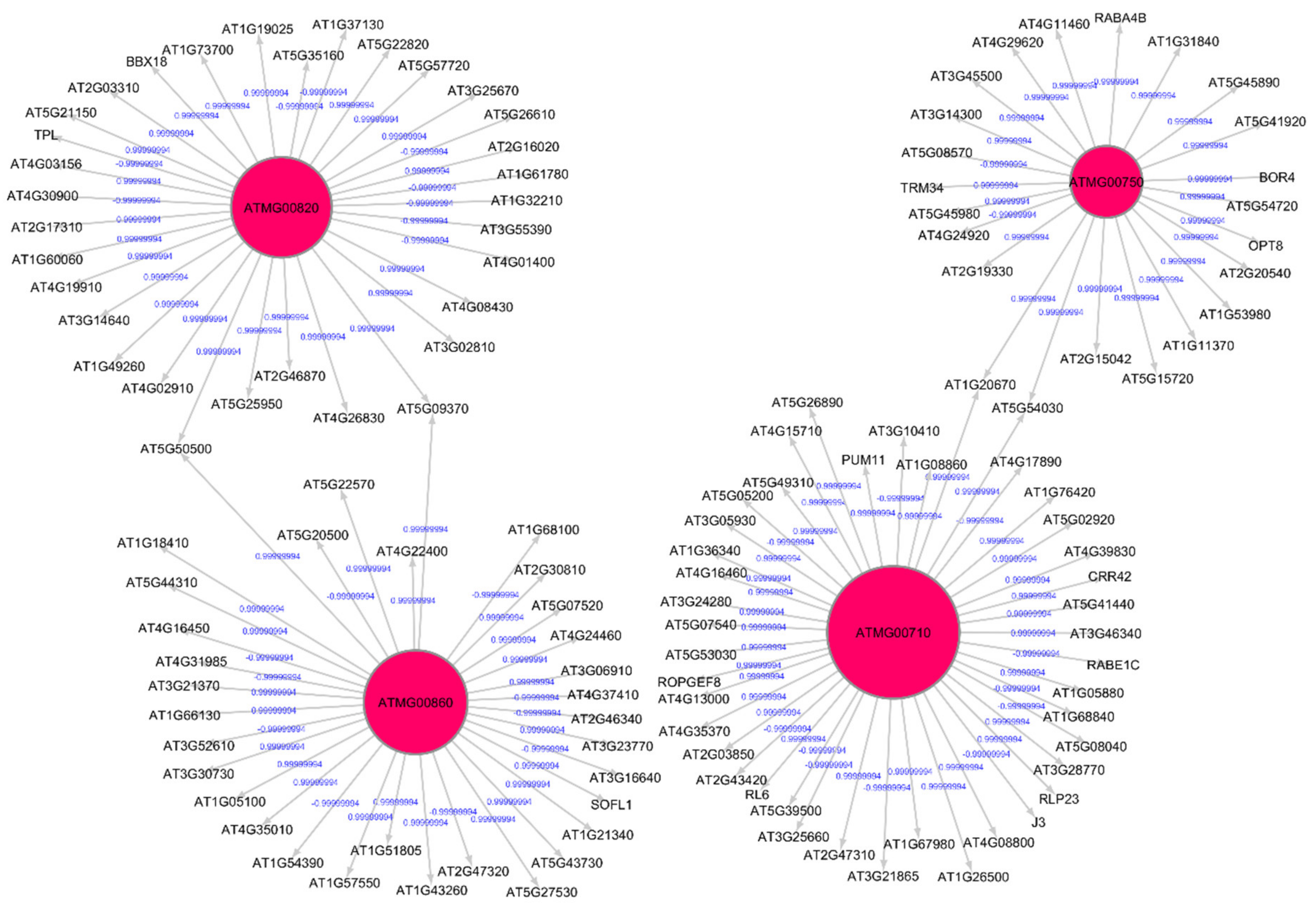
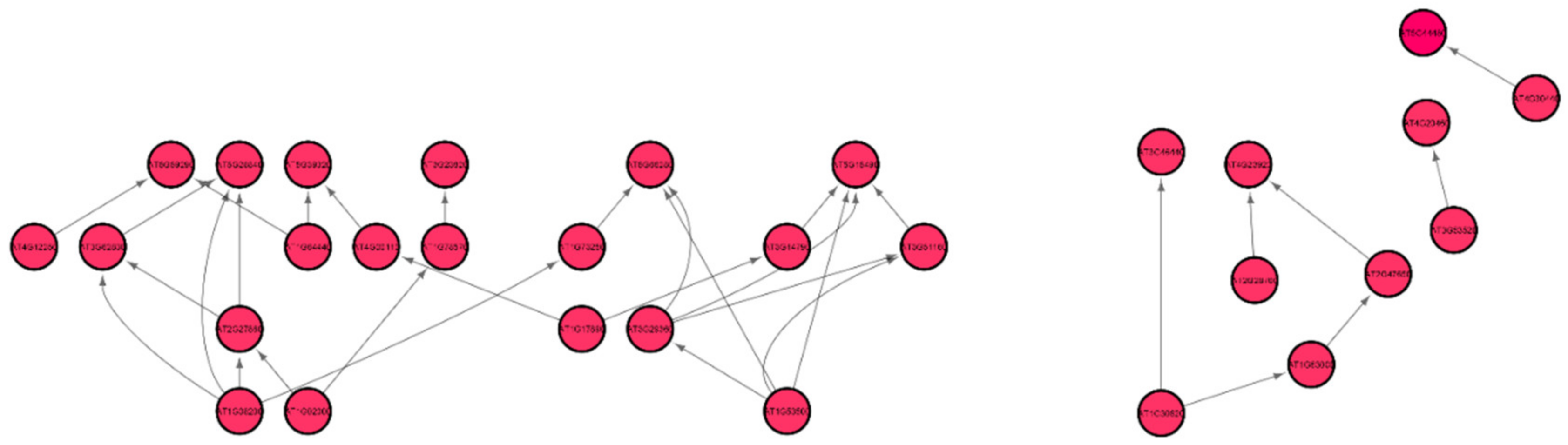
3.2. Topological and Spectral Analyses of Gene Regulatory Networks
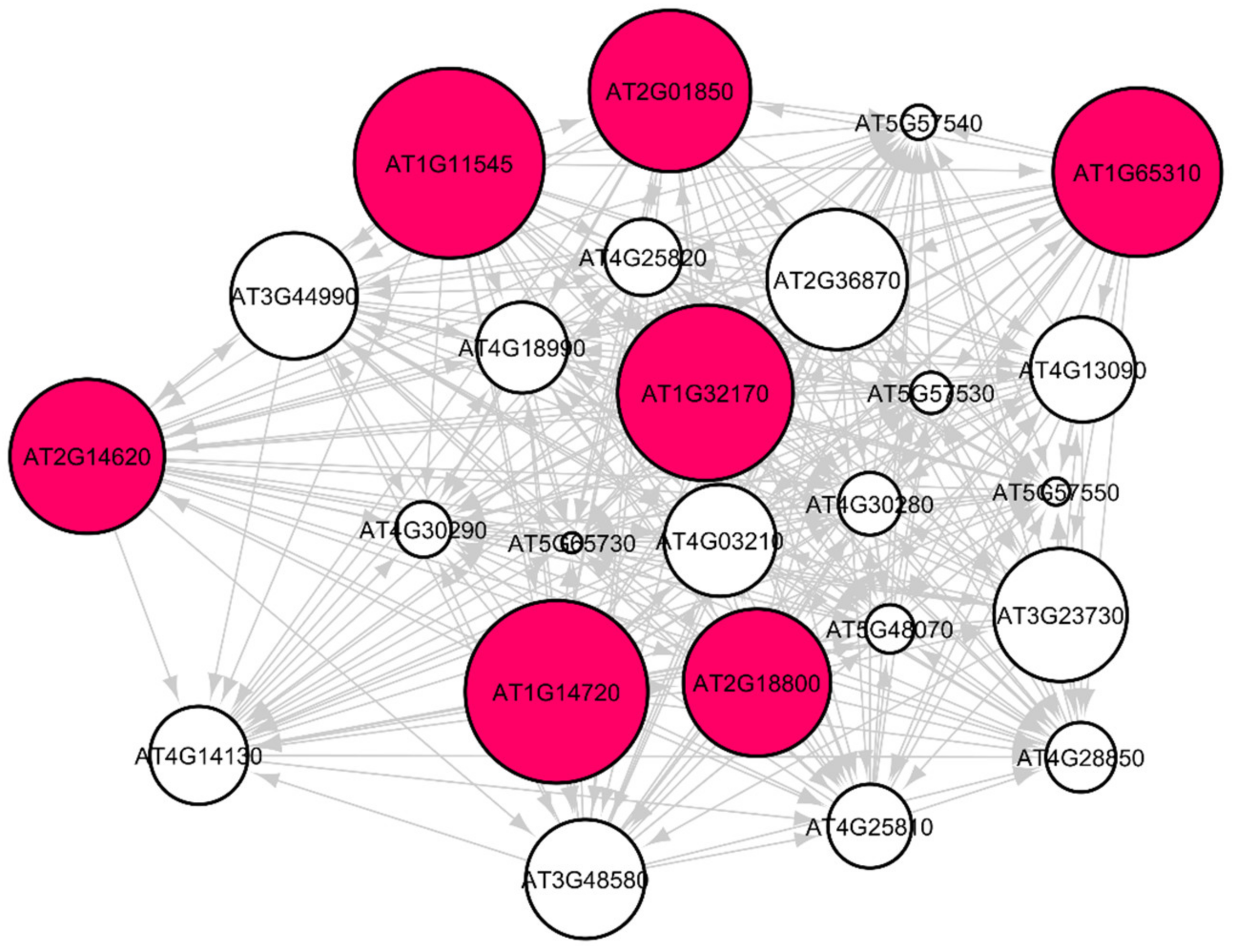
3.3. Arabidopsis Root Growth and Cell Wall Biosynthesis in Spaceflight Microgravity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoson, T. Plant growth and morphogenesis under different gravity conditions: Relevance to plant life in space. Life 2014, 4, 205–216. [Google Scholar] [CrossRef]
- Koornneef, M.; Meinke, D. The development of Arabidopsis as a model plant. Plant J. 2010, 61, 909–921. [Google Scholar] [CrossRef]
- Paul, A.L.; Zupanska, A.K.; Schultz, E.R.; Ferl, R.J. Organ-specific remodeling of the Arabidopsis transcriptome in response to spaceflight. BMC Plant Biol. 2013, 13, 112. [Google Scholar] [CrossRef]
- Kamal, K.Y.; van Loon, J.J.W.A.; Medina, F.J.; Herranz, R. Differential transcriptional profile through cell cycle progression in Arabidopsis cultures under simulated microgravity. Genomics 2019, 111, 1956–1965. [Google Scholar] [CrossRef] [PubMed]
- Kimbrough, J.M. The fast and transient transcriptional network of gravity and mechanical stimulation in the Arabidopsis root Apex. Plant Physiol. 2004, 136, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.Z.; Wolverton, C.; Wyatt, S.E.; Hasenstein, K.H.; Van Loon, J.J.W.A. Comparison of Microgravity Analogs to Spaceflight in Studies of Plant Growth and Development. Front. Plant Sci. 2019. [Google Scholar] [CrossRef]
- Meyer, R.C.; Steinfath, M.; Lisec, J.; Becher, M.; Witucka-Wall, H.; Torjek, O.; Fiehn, O.; Eckardt, A.; Willmitzer, L.; Selbig, J.; et al. The metabolic signature related to high plant growth rate in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2007, 104, 4759–4764. [Google Scholar] [CrossRef] [PubMed]
- Ferl, R.J.; Koh, J.; Denison, F.; Paul, A.L. Spaceflight induces specific alterations in the proteomes of Arabidopsis. Astrobiology 2015, 15, 32–56. [Google Scholar] [CrossRef]
- Johnson, C.M.; Subramanian, A.; Pattathil, S.; Correll, M.J.; Kiss, J.Z. Comparative transcriptomics indicate changes in cell wall organization and stress response in seedlings during spaceflight. Am. J. Bot. 2017, 104, 1219–1231. [Google Scholar] [CrossRef] [PubMed]
- Somssich, M.; Khan, G.A.; Staffan, S.P. Cell wall heterogeneity in root development of Arabidopsis. Front. Plant Sci. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Huang, J.; Gu, X.; Bar-Peled, M.; Xu, Y. Evolution of plant nucleotide-sugar interconversion enzymes. PLoS ONE 2011, 6, e27995. [Google Scholar] [CrossRef] [PubMed]
- Reiter, W.; Vanzin, G.F. Molecular genetics of nucleotide sugar interconversion pathways in plants. Plant Mol. Biol. 2001, 47, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Seifert, G.J. Nucleotide sugar interconversions and cell wall biosynthesis: How to bring the inside to the outside. Curr. Opin. Plant Biol. 2004, 7, 277–284. [Google Scholar] [CrossRef]
- Cai, B.; Li, C.H.; Huang, J. Systematic identification of cell-wall related genes in populus based on analysis of functional modules in co-expression network. PLoS ONE 2014, 9, e95176. [Google Scholar] [CrossRef]
- Wang, S.; Yin, Y.; Ma, Q.; Tang, X.; Hao, D.; Xu, Y. Genome-scale identification of cell-wall related genes in Arabidopsis based on co-expression network analysis. BMC Plant Biol. 2012, 12, 1. [Google Scholar] [CrossRef]
- Choi, W.G.; Barker, R.J.; Kim, S.H.; Swanson, S.J.; Gilroy, S. Variation in the transcriptome of different ecotypes of Arabidopsis thaliana reveals signatures of oxidative stress in plant responses to spaceflight. Am. J. Bot. 2019, 106, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Herranz, R.; Vanderbrink, J.P.; Villacampa, A.; Manzano, A.; Poehlman, W.L.; Feltus, F.A.; Kiss, J.Z.; Medina, F.J. RNAseq analysis of the response of Arabidopsis thaliana to fractional gravity under blue-light stimulation during spaceflight. Front. Plant Sci. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Paul, A.L.; Zupanska, A.K.; Ostrow, D.T.; Zhang, Y.; Sun, Y.; Li, J.L.; Shanker, S.; Farmerie, W.G.; Amalfitano, C.E.; Ferl, R.J. Spaceflight transcriptomes: Unique responses to a novel environment. Astrobiology 2012, 12, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.L.; Zhou, M.; Callaham, J.; Reyes, M.; Stasiak, M.; Riva, A.; Zupanska, A.K.; Dixon, M.; Ferl, R. Patterns of Arabidopsis gene expression in the face of hypobaric stress. AoB Plants 2017, 9, 1–19. [Google Scholar] [CrossRef][Green Version]
- Paul, A.L.; Sng, N.J.; Zupanska, A.K.; Krishnamurthy, A.; Schultz, E.R.; Ferl, R.J. Genetic dissection of the Arabidopsis spaceflight transcriptome: Are some responses dispensable for the physiological adaptation of plants to spaceflight? PLoS ONE 2017, 12, 1–24. [Google Scholar] [CrossRef]
- Dayanne, D. Multi-study inference of regulatory networks for more accurate models of gene regulation. PLoS Comput. Biol. 2019, 15, 1–22. [Google Scholar] [CrossRef]
- Needham, C.J.; Manfield, I.W.; Bulpitt, A.J.; Gilmartin, P.M.; Westhead, D.R. From gene expression to gene regulatory networks in Arabidopsis thaliana. BMC Syst. Biol. 2009, 3, 85. [Google Scholar] [CrossRef] [PubMed]
- Mochida, K.; Koda, S.; Inoue, K.; Nishii, R. Statistical and machine learning approaches to predict gene regulatory networks from transcriptome datasets. Front. Plant Sci. 2018, 871, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pratapa, A.; Jalihal, A.P.; Law, J.N.; Bharadwaj, A.; Murali, T.M. Benchmarking algorithms for gene regulatory network inference from single-cell transcriptomic data. Nat. Methods 2019, 17, 147–154. [Google Scholar] [CrossRef] [PubMed]
- De Lucas, M.; Brady, S.M. Gene regulatory networks in the Arabidopsis root. Curr. Opin. Plant Biol. 2013, 16, 50–55. [Google Scholar] [CrossRef] [PubMed]
- El-Showk, S.; Taylor-Teeples, M.; Helariutta, Y.; Brady, S.M. Gene regulatory networks during Arabidopsis root vascular development. Int. J. Plant Sci. 2013, 174, 1090–1097. [Google Scholar] [CrossRef]
- Taylor-Teeples, M.; Lin, L.; de Lucas, M.; Turco, G.; Toal, T.W.; Gaudinier, A.; Young, N.F.; Trabucco, G.M.; Veling, M.T.; Lamothe, R.; et al. An Arabidopsis gene regulatory network for secondary cell wall synthesis. Nature 2015, 517, 571–575. [Google Scholar] [CrossRef]
- Doroshkov, A.V.; Konstantinov, D.K.; Afonnikov, D.A.; Gunbin, K.V. The evolution of gene regulatory networks controlling Arabidopsis thaliana L. trichome development. BMC Plant Biol. 2019, 19, 53. [Google Scholar] [CrossRef]
- Hehl, R.; Bulow, L. AthaMap Web Tools for the Analysis of Transcriptional and Posttranscriptional Regulation of Gene Expression in Arabidopsis thaliana. Plant Circadian Networks; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2014; Volume 1158. [Google Scholar]
- Biggs, N. Algebraic Graph Theory, 2nd ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Du, Q.; Campbell, M.; Yu, H.; Liu, K.; Walia, H.; Zhang, Q.; Zhang, C. Network-based feature selection reveals substructures of gene modules responding to salt stress in rice. Plant Direct 2019, 3, e00154. [Google Scholar] [CrossRef]
- Zhang, R.; Ren, Z.; Chen, W. SILGGM: An extensive R package for efficient statistical inference in large-scale gene networks. PLoS Comput. Biol. 2018, 14, e1006369. [Google Scholar] [CrossRef]
- Serre, J.-P. Trees; Springer-Verlag: Berlin/Heidelberg, Germany, 1980. [Google Scholar]
- Kleinberg, J.M. Authoritative sources in a hyperlinked environment. J. ACM 1999, 46, 604–632. [Google Scholar] [CrossRef]
- Lei, X.; Wang, S.; Wu, F.X. Identification of essential proteins based on improved HITS algorithm. Genes 2019, 10, 177. [Google Scholar] [CrossRef] [PubMed]
- Easley, D.; Kleinberg, J. Networks, Crowds, and Markets; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Miller, J.C.; Rae, G.; Schaefer, F.; Ward, L.A.; LoFaro, T.; Farahat, A. Modifications of Kleinberg’s HITS algorithm using matrix exponentiation and web log records. In Proceedings of the 24th Annual International ACM SIGIR Conference on Research and Development in Information Retrieval, New Orleans, LA, USA, 9–13 September 2001; pp. 444–445. [Google Scholar]
- Langville, A.N.; Meyer, C.D. A survey of eigenvector methods for web information retrieval. SIAM Rev. 2005, 47, 135–161. [Google Scholar] [CrossRef]
- Emms, D.M.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef] [PubMed]
- Bian, T.; Hu, J.; Deng, Y. Identifying influential nodes in complex networks based on AHP. Phys. A Stat. Mech. Appl. 2017, 479, 422–436. [Google Scholar] [CrossRef]
- Urbinati, A.; Galimberti, E.; Ruffo, G. Hubs and authorities of the scientific migration network. arXiv 2019, arXiv:1907.07175. [Google Scholar]
- Eldén, L. Matrix Methods in Data Mining and Pattern Recognition, 2th ed.; SIAM: Philadelphia, PA, USA, 2019. [Google Scholar]
- Cowen, L.; Ideker, T.; Raphael, B.J.; Sharan, R. Network propagation: A universal amplifier of genetic associations. Nat. Rev. Genet. 2017, 18, 551. [Google Scholar] [CrossRef] [PubMed]
- Opsahl, T.; Agneessens, F.; Skvoretz, J. Node centrality in weighted networks: Generalizing degree and shortest paths. Soc. Netw. 2010, 32, 245–251. [Google Scholar] [CrossRef]
- Gazestani, V.H.; Lewis, N.E. From genotype to phenotype: Augmenting deep learning with networks and systems biology. Curr. Opin. Syst. Biol. 2019, 15, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Loscalzo, J.; Barabási, A.-L. Network Science, 1st ed.; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Janwa, H.; Massey, S.E.; Velev, J.; Mishra, B. On the origin of biomolecular networks. Front. Genet. 2019, 10, 240–257. [Google Scholar] [CrossRef]
- Aukerman, M.J.; Hirschfeld, M.; Wester, L.; Weaver, M.; Clack, T.; Amasino, R.M.; Sharrock, R.A. A deletion in the PHYD gene of the Arabidopsis wassilewskija ecotype defines a role for phytochrome D in red/far-red light sensing. Plant Cell 1997, 9, 1317–1326. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Interactor Gene A | Interactor Gene B | Correlation | t-Statistics | p-Value |
|---|---|---|---|---|
| AT1G03430 | AT1G11185 | −0.95787 | −4.716511 | 0.04213 |
| AT1G03430 | AT1G12390 | 0.984801 | 8.018483 | 0.0152 |
| AT1G03430 | AT1G17170 | −0.98068 | −7.090435 | 0.01932 |
| AT1G03430 | AT1G17180 | 0.937057 | 3.795227 | 0.06294 |
| AT1G03430 | AT1G21140 | 0.454929 | 0.722456 | 0.54507 |
| AT1G03430 | AT1G24240 | 0.993113 | 11.98728 | 0.00689 |
| AT1G03430 | AT1G26470 | 0.98765 | 8.914699 | 0.01235 |
| AT1G03430 | AT1G27130 AT1G27140 | 0.998482 | 25.63509 | 0.00152 |
| AT1G03430 | AT1G27570 | −0.95498 | −4.552277 | 0.04502 |
| AT1G03430 | AT1G30360 | 0.9732 | 5.985038 | 0.0268 |
| AT1G03430 | AT1G32460 | 0.4588 | 0.730233 | 0.5412 |
| AT1G03430 | AT1G34180 | −0.99271 | −11.64964 | 0.00729 |
| AT1G03430 | AT1G34844 | 0.947816 | 4.204318 | 0.05218 |
| Arabidopsis Root | ATG Number | Enriched Sene Set (Biological Process) |
|---|---|---|
| Root growth spaceflight hub genes | ATMG00880 ATMG00840 ATMG00660 | Part of mitochondrion; Enables molecular function; Involved in biological process |
| Root growth ground control hub genes | ATMG00720 ATMG01040 ATMG01020 ATMG00580 | Part of mitochondrion Enables molecular function; Involved in biological process; ATP synthesis coupled electron transport; Involved in oxidation-reduction process |
| Shared hub genes for root growth in spaceflight and ground control | ATMG00860 ATMG00510 ATMG00750 ATMG00740 ATMG00720 ATMG00600 ATMG00680 ATMG00820 | Part of mitochondrion Enables molecular function; Involved in biological process; Enables NADH dehydrogenase activity; Part of mitochondrial respiratory chain complex 1; Involved in oxidation-reduction process; Enables NAD binding; quinone binding |
| Dataset | Spectral Gap | Average Clustering Coefficient | Average Shortest Path Length (Distance) | Diameter (Maximum Eccentricity) | Authority Genes (Outdegree) |
|---|---|---|---|---|---|
| AT root spaceflight | 39.228 | 0.00897 | 1.452 | 3 | 95 |
| AT root ground | 59.96 | 0.00315 | 1.2017 | 3 | 166 |
| AT WS spaceflight light | 72.9 | 0.01272 | 1.89 | 6 | 1237.04 |
| AT WS ground light | 0.45 | 0.01788 | 1.11 | 2 | 45.63 |
| AT WS spaceflight dark | 3.99 | 0.01996 | 1.069 | 2 | 50.41 |
| AT WS ground dark | 2.32 | 0.02011 | 1.069 | 3 | 43.36 |
| AT phyD spaceflight light | 0.79 | 0.01000 | 1.036 | 2 | 35 |
| AT phyD ground light | 0.22 | 0.01996 | 1.069 | 2 | 50.41 |
| AT phyD spaceflight dark | 0.001 | 0.01219 | 1.146 | 2 | 38.24 |
| AT phyD ground dark | 3.15 | 0.0219 | 1.125 | 2 | 53.29 |
| AT Col-0 spaceflight light | 0.13 | 0.0212 | 1.107 | 2 | 40.26 |
| AT Col-0 ground light | 4.00 | 0.0146 | 1.113 | 2 | 44.28 |
| AT Col-0 spaceflight dark | 0.20 | 0.0224 | 1.066 | 1 | 29.09 |
| AT Col-0 ground dark | 2.67 | 0.0187 | 1.076 | 1 | 33.42 |
| ATG Number | Hub Gene Name/Description | Enriched Gene Set (Biological Process) |
|---|---|---|
| AT2G36870 | XTH32 | Involved in cellular glucan metabolic process |
| AT2G18800 | XTH21 | Involved in cell wall organization |
| AT3G44990 | XTH31 | Involved in glucanase activity |
| AT3G23730 | XTH16 | Involved in the metabolic process |
| AT2G14620 | XTH10 | Involved in cellular glucan metabolic process |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manian, V.; Orozco, J.; Gangapuram, H.; Janwa, H.; Agrinsoni, C. Network Analysis of Gene Transcriptions of Arabidopsis thaliana in Spaceflight Microgravity. Genes 2021, 12, 337. https://doi.org/10.3390/genes12030337
Manian V, Orozco J, Gangapuram H, Janwa H, Agrinsoni C. Network Analysis of Gene Transcriptions of Arabidopsis thaliana in Spaceflight Microgravity. Genes. 2021; 12(3):337. https://doi.org/10.3390/genes12030337
Chicago/Turabian StyleManian, Vidya, Jairo Orozco, Harshini Gangapuram, Heeralal Janwa, and Carlos Agrinsoni. 2021. "Network Analysis of Gene Transcriptions of Arabidopsis thaliana in Spaceflight Microgravity" Genes 12, no. 3: 337. https://doi.org/10.3390/genes12030337
APA StyleManian, V., Orozco, J., Gangapuram, H., Janwa, H., & Agrinsoni, C. (2021). Network Analysis of Gene Transcriptions of Arabidopsis thaliana in Spaceflight Microgravity. Genes, 12(3), 337. https://doi.org/10.3390/genes12030337