
Identification and Characterization of New Alu Element Insertion in the BRCA1 Exon 14 Associated with Hereditary Breast and Ovarian Cancer

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
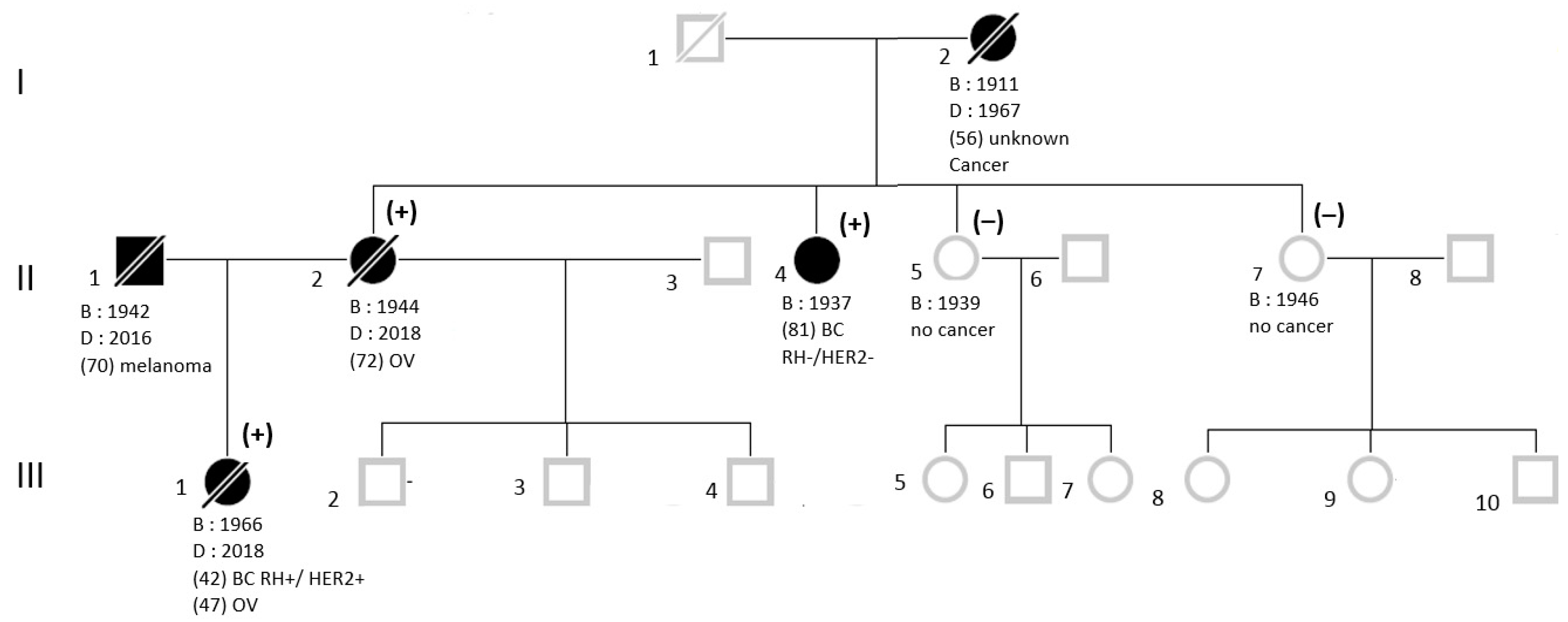
2.1. Subjects
2.2. NGS Analysis
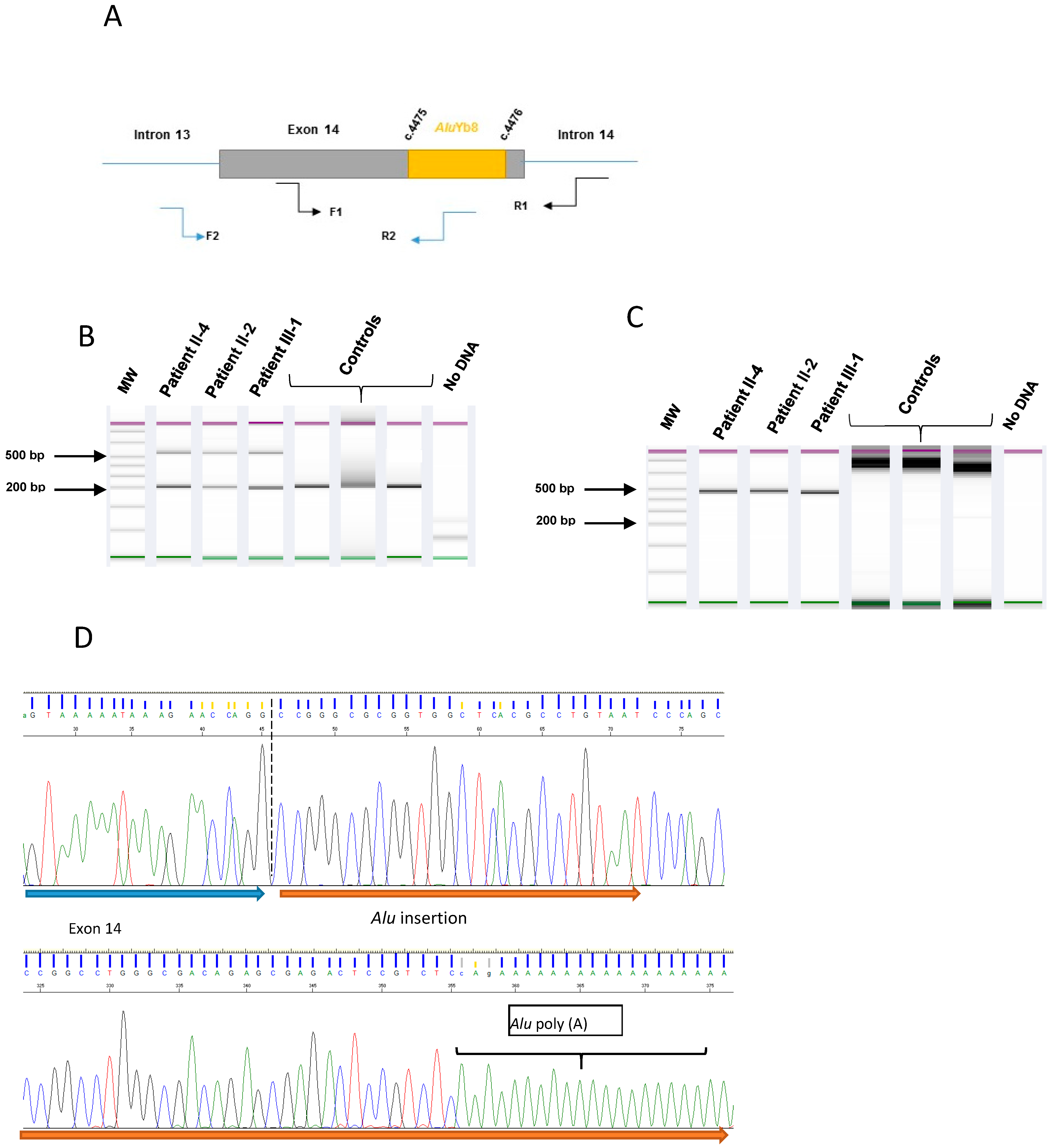
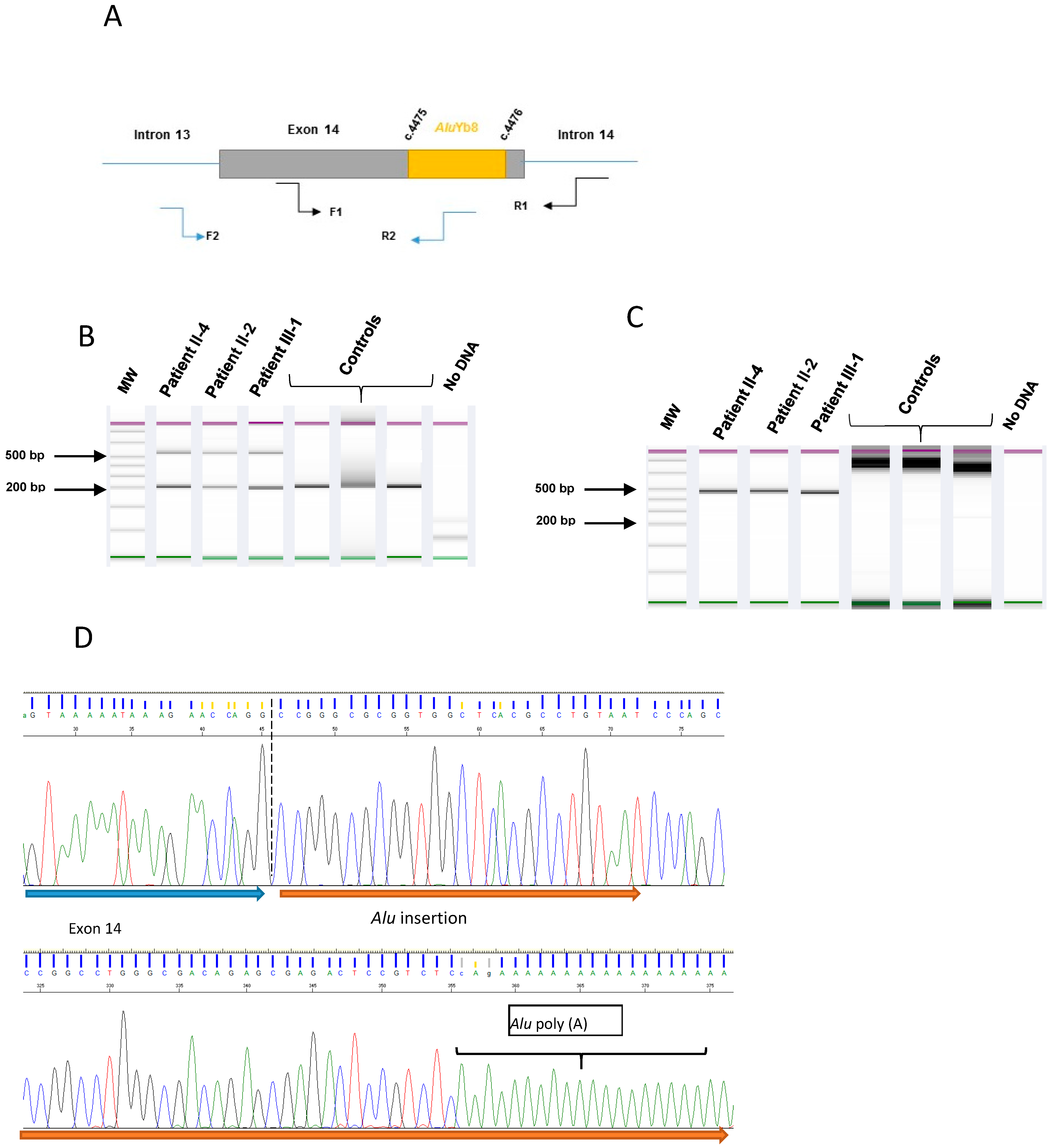
2.3. Characterization for the Alu Insertion
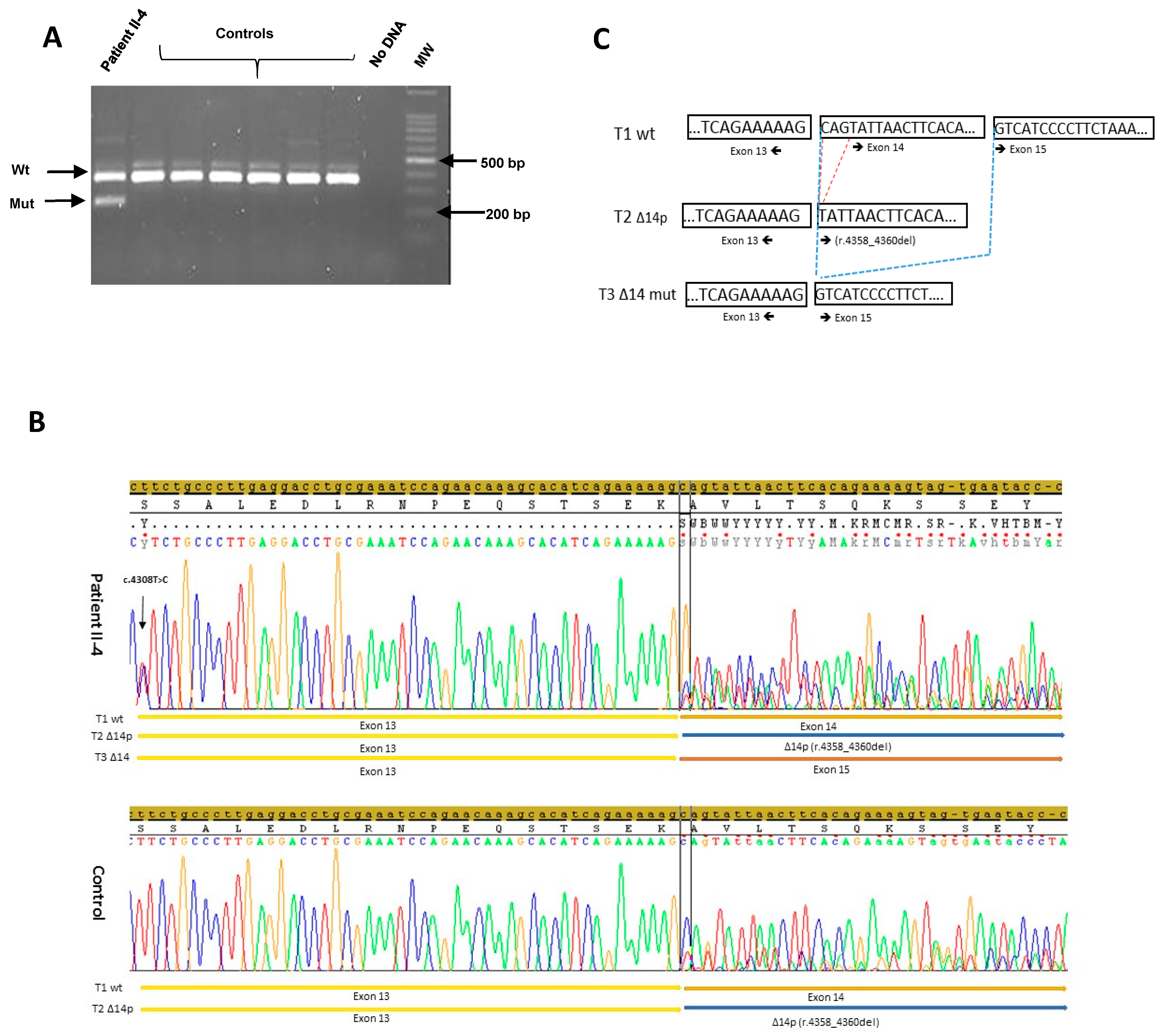
2.4. RT-PCR Analysis
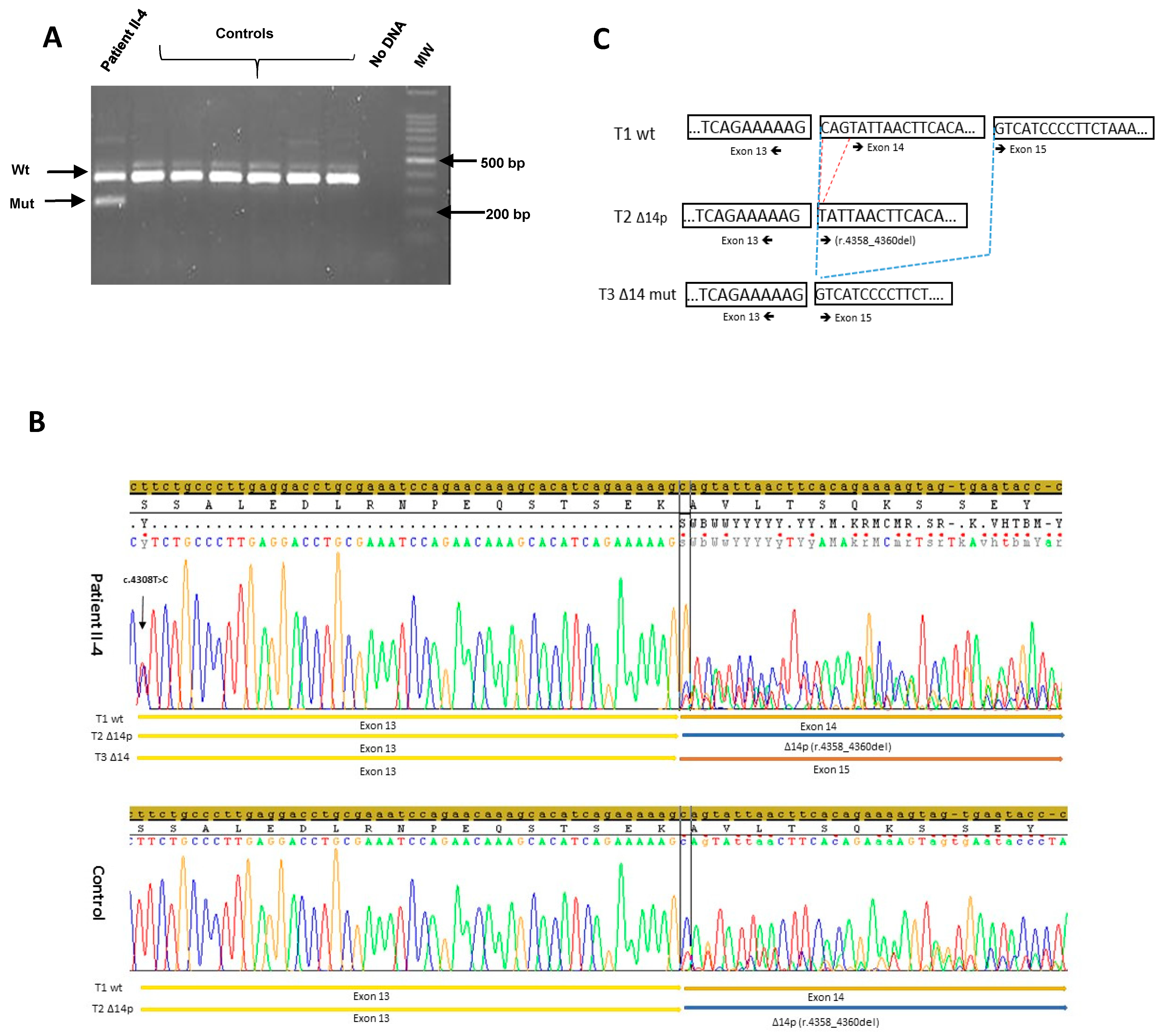
3. Results
3.1. NGS Sequencing
3.2. Alu Sequence Characterization
3.3. RNA Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoshida, R. Hereditary breast and ovarian cancer (HBOC): Review of its molecular characteristics, screening, treatment, and prognosis. Breast Cancer 2021, 28, 1167–1180. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Ohno, S.; Sasaki, Y.; Matsuura, M. Hereditary breast and ovarian cancer susceptibility genes (Review). Oncol. Rep. 2013, 30, 1019–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalloo, F.; Evans, D.G. Familial Breast Cancer. Clin. Genet. 2012, 82, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.; Casadei, S.; Lee, M.K.; Pennil, C.C.; Nord, A.; Thornton, A.M.; Roeb, W.; Agnew, K.J.; Stray, S.M.; Wickramanayake, A.; et al. Mutations in 12 genes for inherited ovarian, fallopian tube, and peritoneal carcinoma identified by massively parallel sequencing. Proc. Natl. Acad. Sci. USA 2011, 108, 18032–18037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordaux, R.; Batzer, M.A. The impact of retrotransposons on human genome evolution. Nat. Rev. Genet. 2009, 10, 691–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peixoto, A.; Santos, C.; Pinheiro, M.; Pinto, P.; Soares, M.J.; Rocha, P.; Gusmão, L.; Amorim, A.; Van Der Hout, A.; Gerdes, A.-M.; et al. International distribution and age estimation of the Portuguese BRCA2 c.156_157insAlu founder mutation. Breast Cancer Res. Treat. 2010, 127, 671–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Salo-Mullen, E.; Varghese, A.; Trottier, M.; Stadler, Z.K.; Zhang, L. Insertion of an Alu-like element in MLH1 intron 7 as a novel cause of Lynch syndrome. Mol. Genet Genom. Med. 2020, 8, e1523. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Mancini-DiNardo, D.; Judkins, T.; Cox, H.C.; Brown, K.; Elias, M.; Singh, N.; Daniels, C.; Holladay, J.; Coffee, B.; et al. Identification of pathogenic retrotransposon in-sertions in cancer predisposition genes. Cancer Genet. 2017, 216–217, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peixoto, A.; Santos, C.; Rocha, P.; Pinheiro, M.; Príncipe, S.; Pereira, D.; Rodrigues, H.; Castro, F.; Abreu, J.; Gusmão, L.; et al. The c.156_157insAlu BRCA2 rearrangement accounts for more than one-fourth of deleterious BRCA mutations in northern/central Portugal. Breast Cancer Res. Treat. 2008, 114, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Solassol, J.; Larrieux, M.; Leclerc, J.; Ducros, V.; Corsini, C.; Chiésa, J.; Pujol, P.; Rey, J.-M. Alu element insertion in the MLH1 exon 6 coding se-quence as a mutation predisposing to Lynch syndrome. Hum. Mutat. 2019, 40, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Baert-Desurmont, S.; Coutant, S.; Charbonnier, F.; Macquere, P.; Lecoquierre, F.; Schwartz, M.; Blanluet, M.; Vezain, M.; Lanos, R.; Quenez, O.; et al. Optimization of the diagno-sis of inherited colorectal cancer using NGS and capture of exonic and intronic sequences of panel genes. Eur. J. Hum. Genet. 2018, 26, 1597–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parenti, S.; Rabacchi, C.; Marino, M.; Tenedini, E.; Artuso, L.; Castellano, S.; Carretta, C.; Mallia, S.; Cortesi, L.; Toss, A.; et al. Characterization of New ATM Deletion Associ-ated with Hereditary Breast Cancer. Genes 2021, 12, 136. [Google Scholar] [CrossRef] [PubMed]
- Houdayer, C.; Caux-Moncoutier, V.; Krieger, S.; Barrois, M.; Bonnet, F.; Bourdon, V.; Bronner, M.; Buisson, M.; Coulet, F.; Gaildrat, P.; et al. Guidelines for splicing analysis in mo-lecular diagnosis derived from a set of 327 combined in silico/in vitro studies on BRCA1 and BRCA2 variants. Hum. Mutat. 2012, 33, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Hubley, R.; Finn, R.; Clements, J.; Eddy, S.R.; Jones, T.A.; Bao, W.; Smit, A.F.A.; Wheeler, T.J. The Dfam database of repetitive DNA families. Nucleic Acids Res. 2016, 44, D81–D89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davy, G.; Rousselin, A.; Goardon, N.; Castéra, L.; Harter, V.; Legros, A.; Muller, E.; Fouillet, R.; Brault, B.; Smirnova, A.S.; et al. Detecting splicing patterns in genes involved in hereditary breast and ovarian cancer. Eur. J. Hum. Genet. 2017, 25, 1147–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, G.K.; Collier, A.L.; Lee, D.; Hoefer, R.A.; Zheleva, V.; van Reesema, L.L.S.; Tang-Tan, A.M.; Guye, M.L.; Chang, D.Z.; Winston, J.S.; et al. Perspectives on Triple-Negative Breast Cancer: Current Treatment Strategies, Unmet Needs, and Potential Targets for Future Therapies. Cancers 2020, 12, 2392. [Google Scholar] [CrossRef] [PubMed]
- Disdero, E.; Filée, J. LoRTE: Detecting transposon-induced genomic variants using low coverage PacBio long read sequences. Mob. DNA 2017, 8, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouras, A.; Leone, M.; Bonadona, V.; Lebrun, M.; Calender, A.; Boutry-Kryza, N. Identification and Characterization of New Alu Element Insertion in the BRCA1 Exon 14 Associated with Hereditary Breast and Ovarian Cancer. Genes 2021, 12, 1736. https://doi.org/10.3390/genes12111736
Bouras A, Leone M, Bonadona V, Lebrun M, Calender A, Boutry-Kryza N. Identification and Characterization of New Alu Element Insertion in the BRCA1 Exon 14 Associated with Hereditary Breast and Ovarian Cancer. Genes. 2021; 12(11):1736. https://doi.org/10.3390/genes12111736
Chicago/Turabian StyleBouras, Ahmed, Melanie Leone, Valerie Bonadona, Marine Lebrun, Alain Calender, and Nadia Boutry-Kryza. 2021. "Identification and Characterization of New Alu Element Insertion in the BRCA1 Exon 14 Associated with Hereditary Breast and Ovarian Cancer" Genes 12, no. 11: 1736. https://doi.org/10.3390/genes12111736
APA StyleBouras, A., Leone, M., Bonadona, V., Lebrun, M., Calender, A., & Boutry-Kryza, N. (2021). Identification and Characterization of New Alu Element Insertion in the BRCA1 Exon 14 Associated with Hereditary Breast and Ovarian Cancer. Genes, 12(11), 1736. https://doi.org/10.3390/genes12111736