
An Integrative Transcriptomic and Metabolomic Analysis of Red Pitaya (Hylocereus polyrhizus) Seedlings in Response to Heat Stress

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
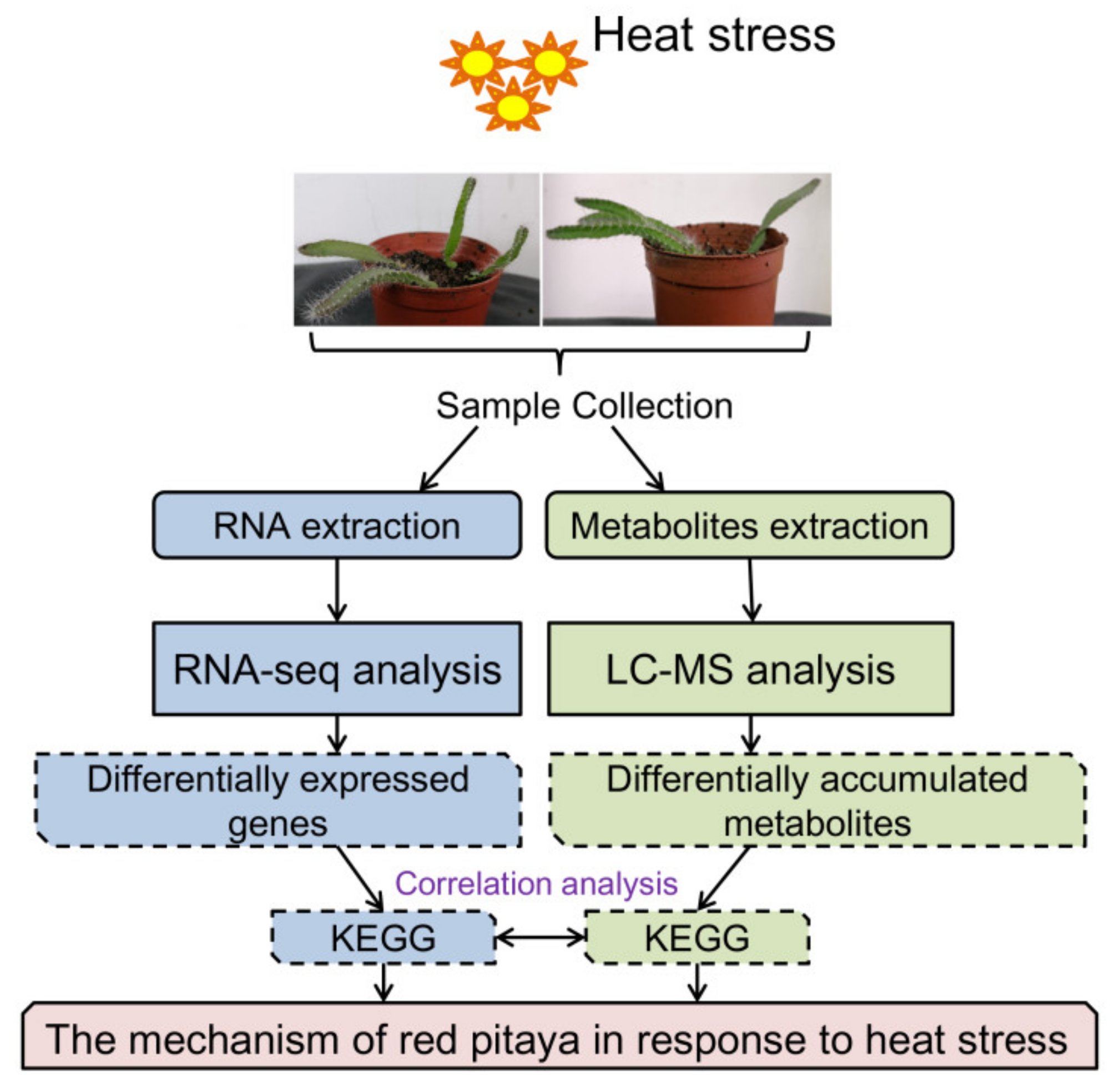
2.1. Plant Materials and Heat Treatments
2.2. RNA Isolation and Library Preparation for Transcriptomic Analysis
2.3. Sequencing, De Novo Assembly, and Annotation
2.4. Identification of DEGs
2.5. Transcriptome Assembly and DEG Validation
2.6. Extraction of Metabolites
2.7. LC–MS/MS Analysis
2.8. Data Preprocessing and OPLS-DA
2.9. DNA Constructs and Plant Transformation
2.10. HS Tolerance Assays in Transgenic Arabidopsis
2.11. Statistical Analysis
3. Results
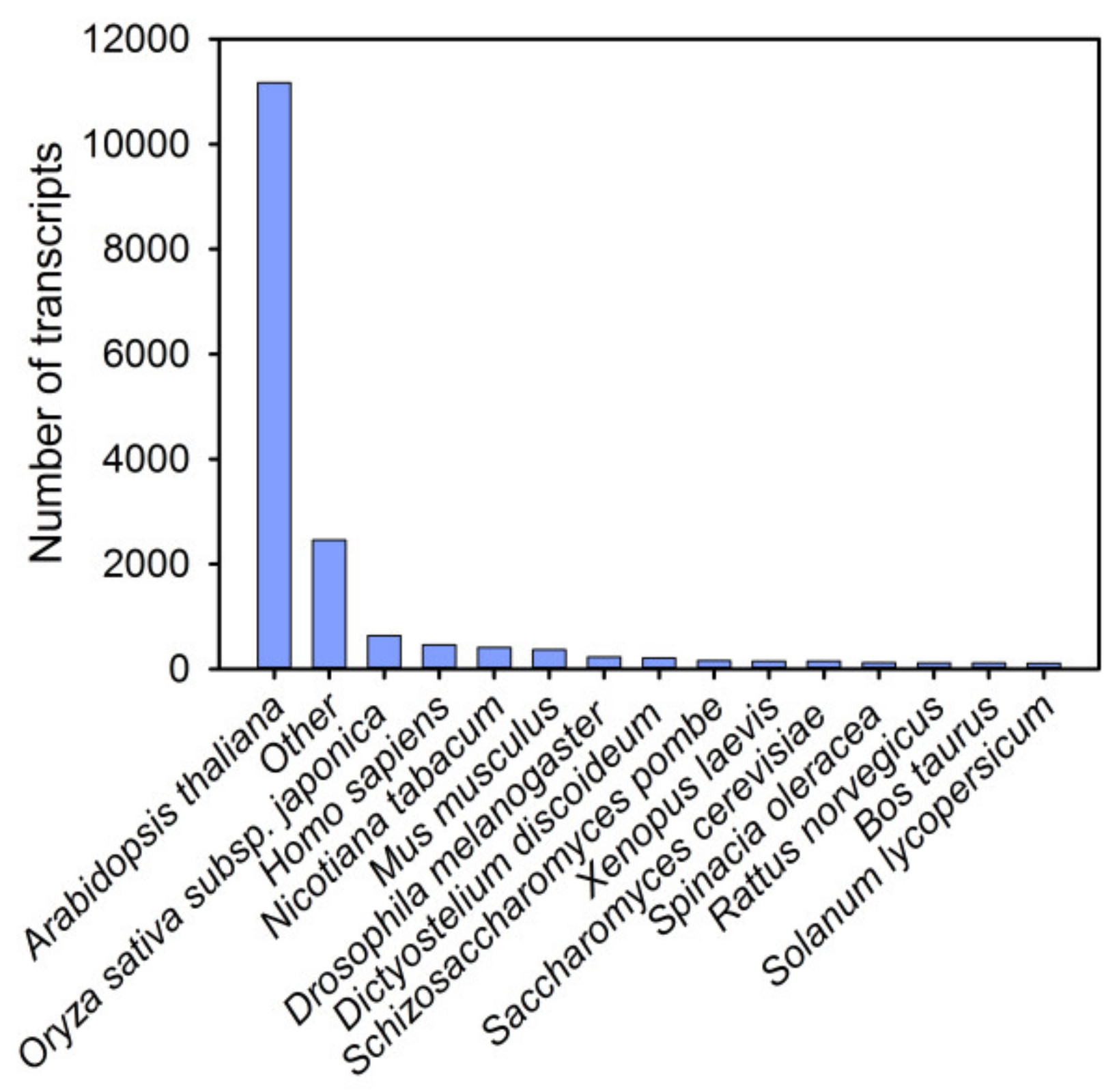
3.1. Sequencing and De Novo Assembly of the Pitaya Transcriptome
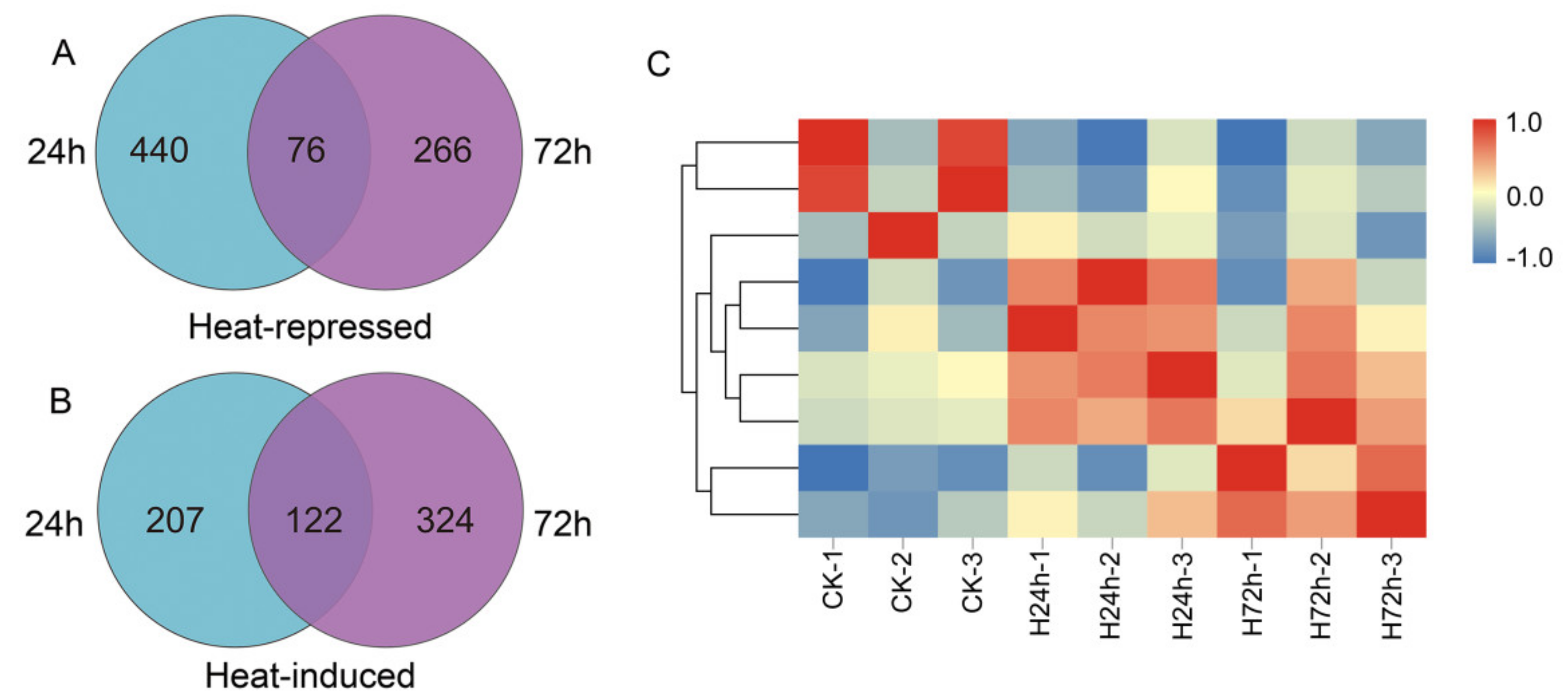
3.2. Identification of Differentially Expressed Genes (DEGs) under HS
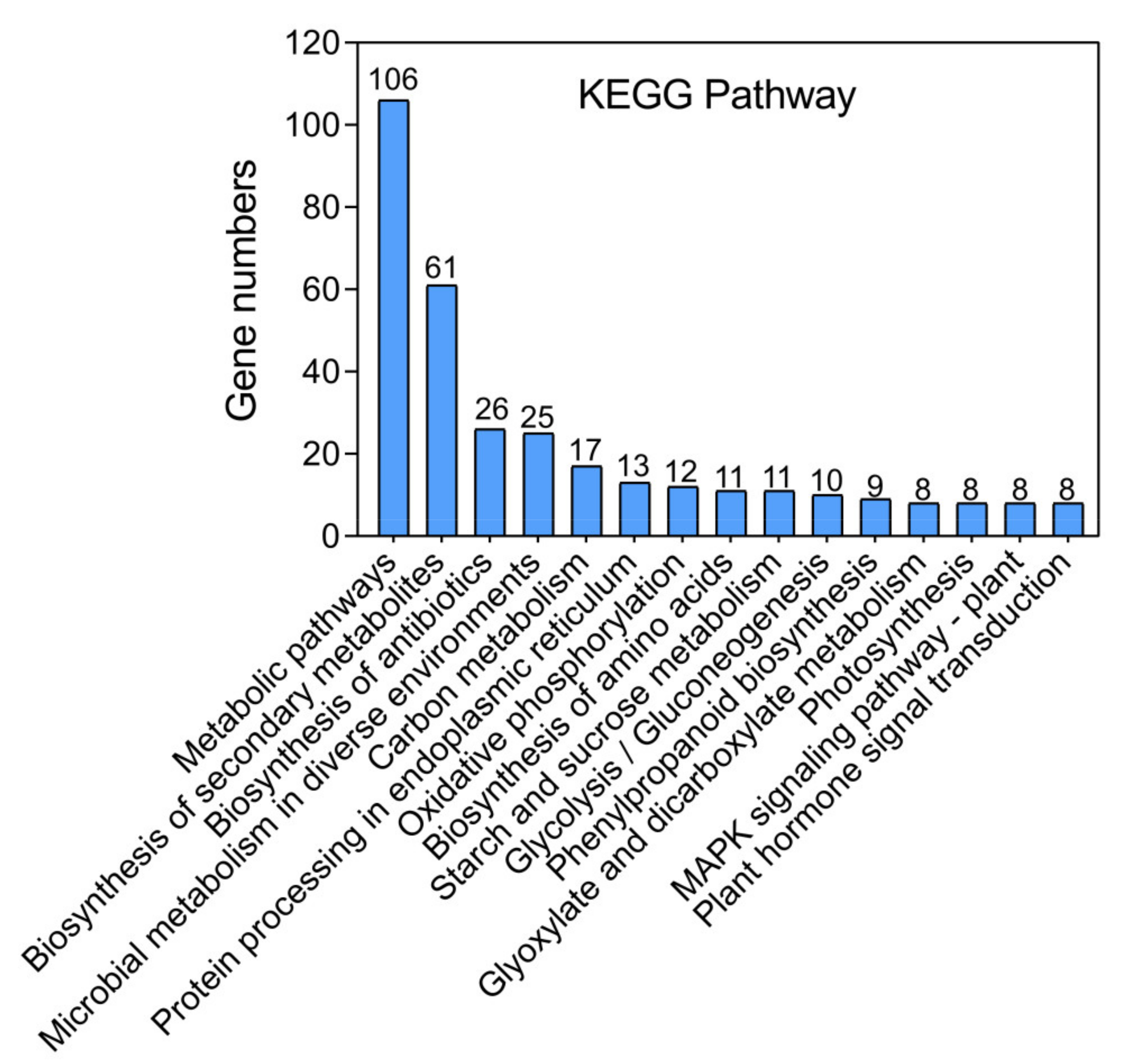
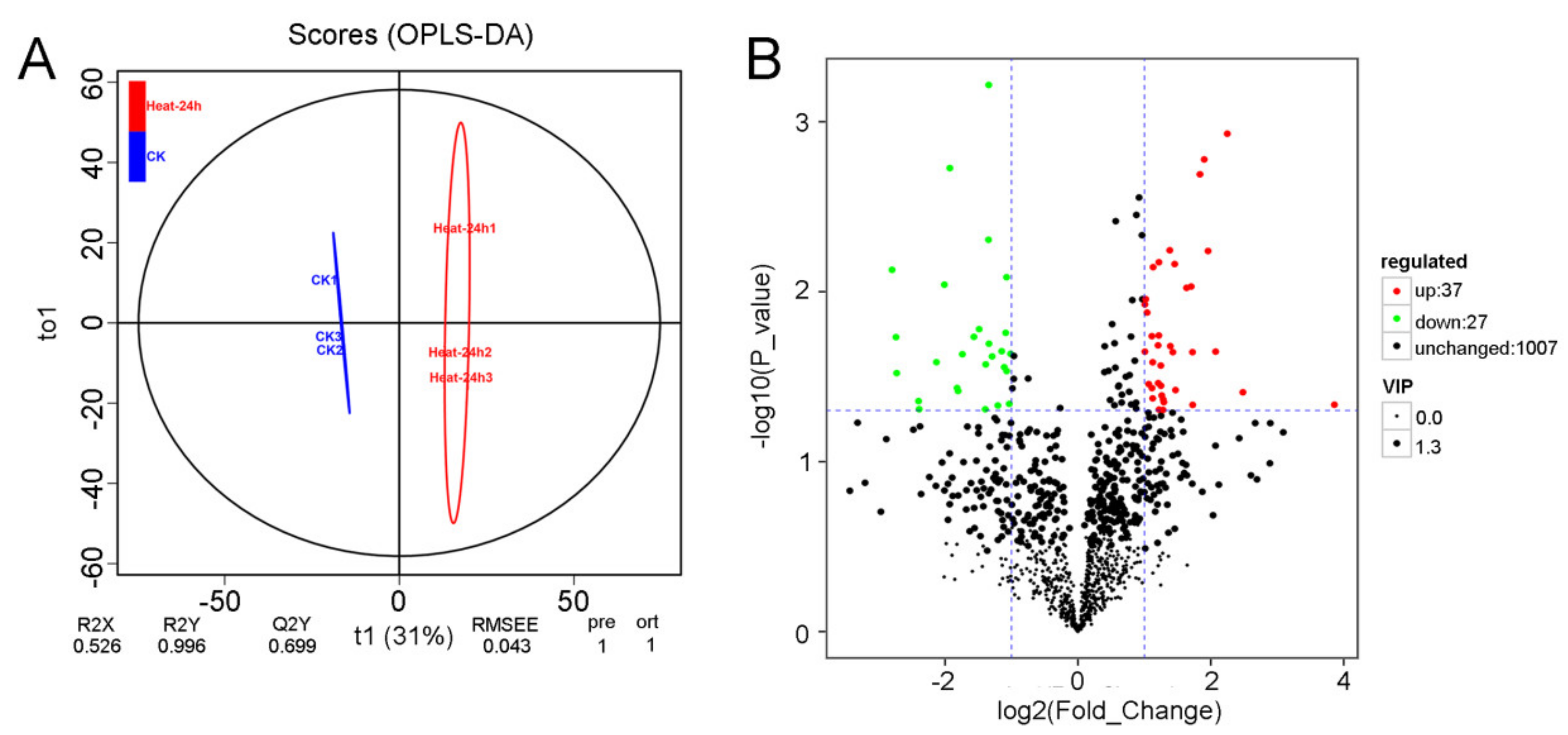
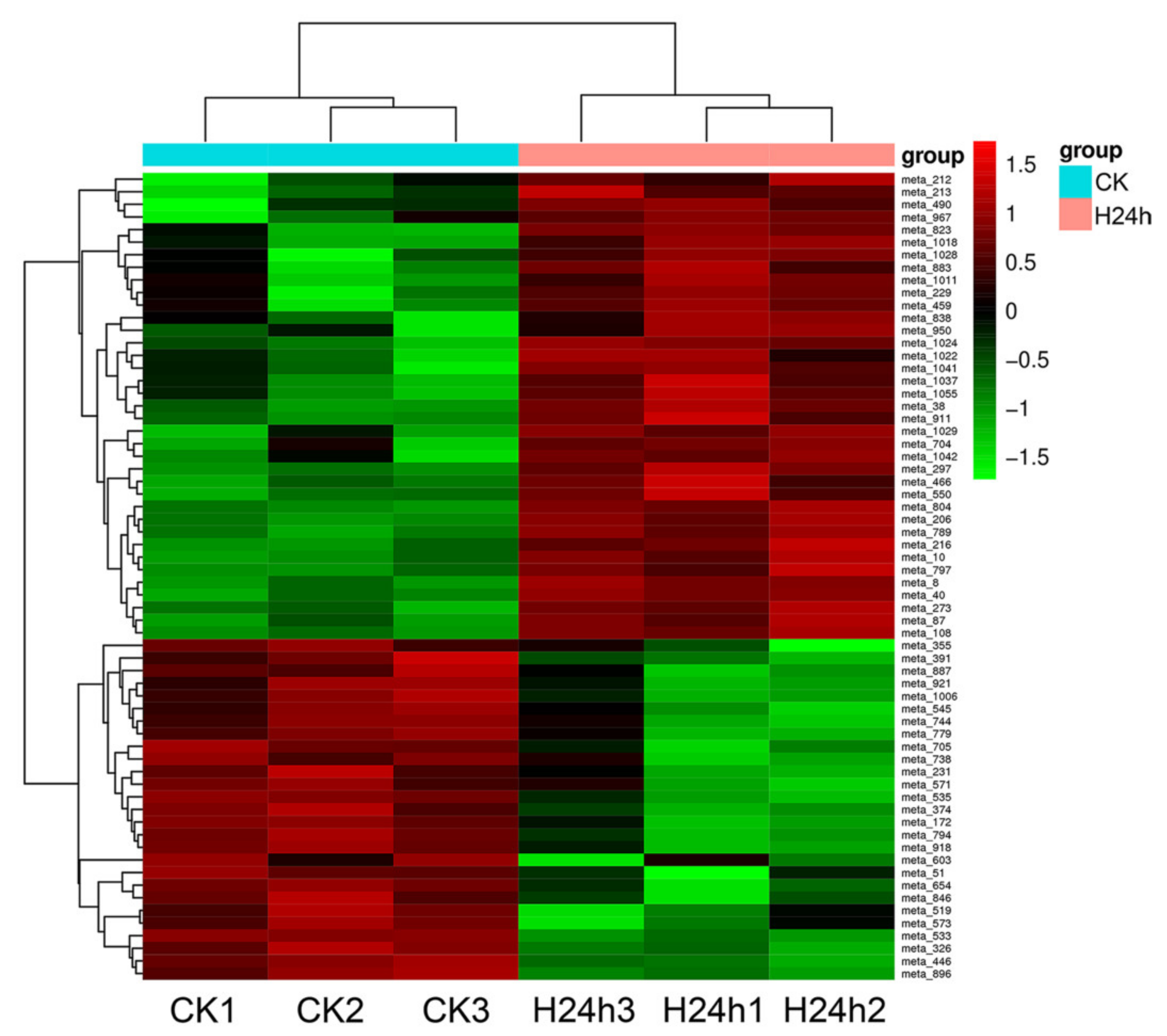
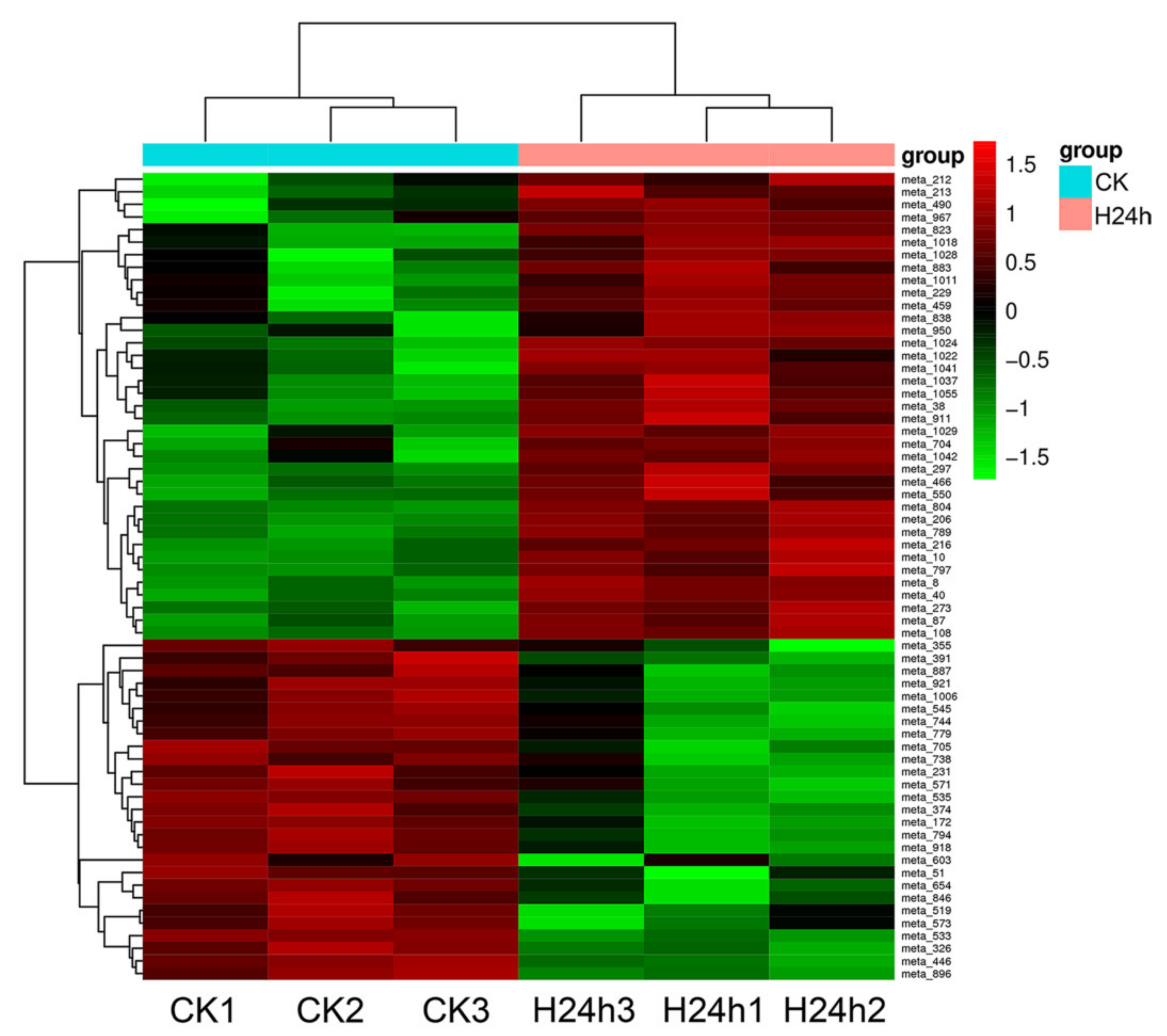
3.3. Analysis of the Metabolites in Red Pitaya Seedlings under HS
3.4. Interaction Network Analysis between Heat-Regulated Genes and Metabolites
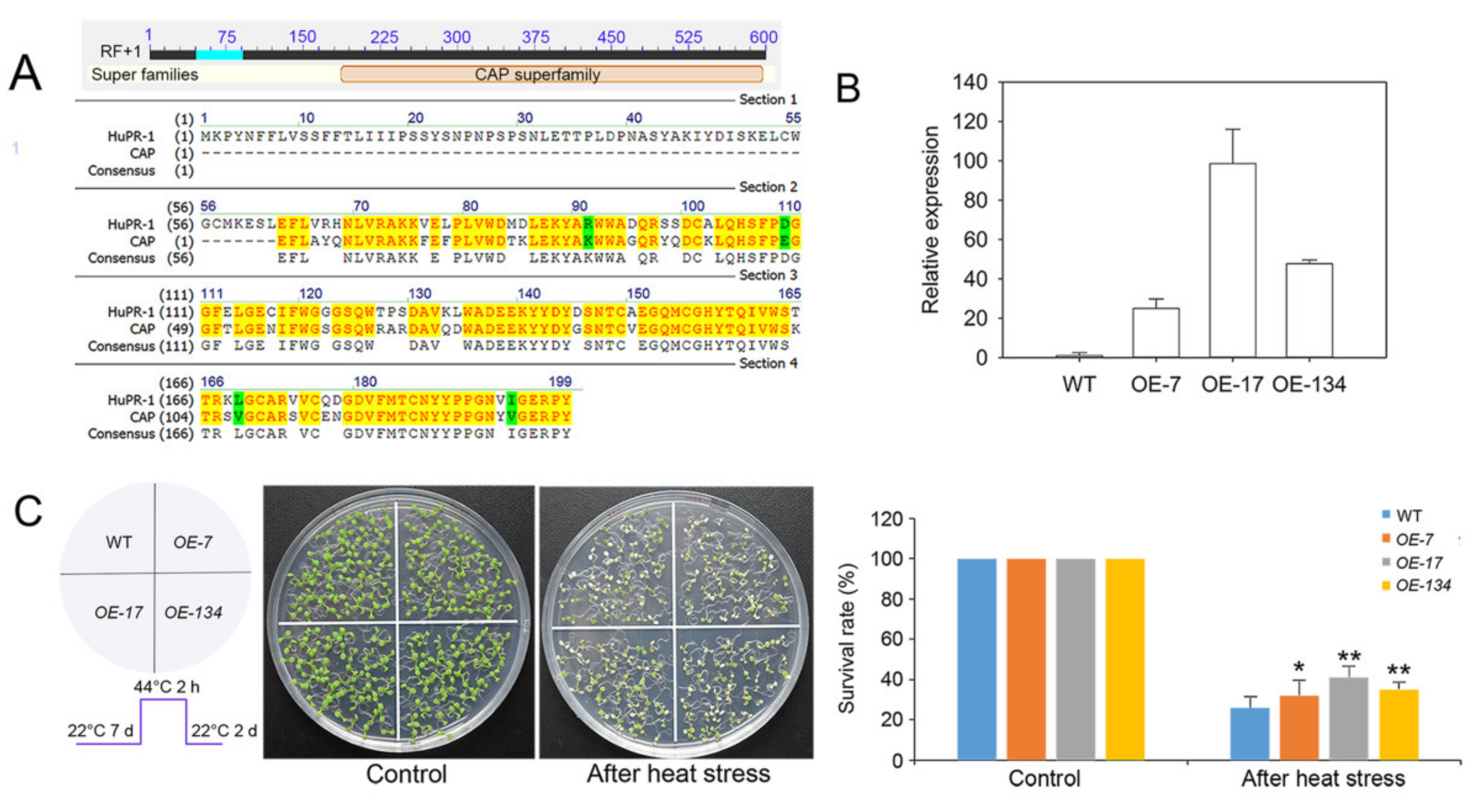
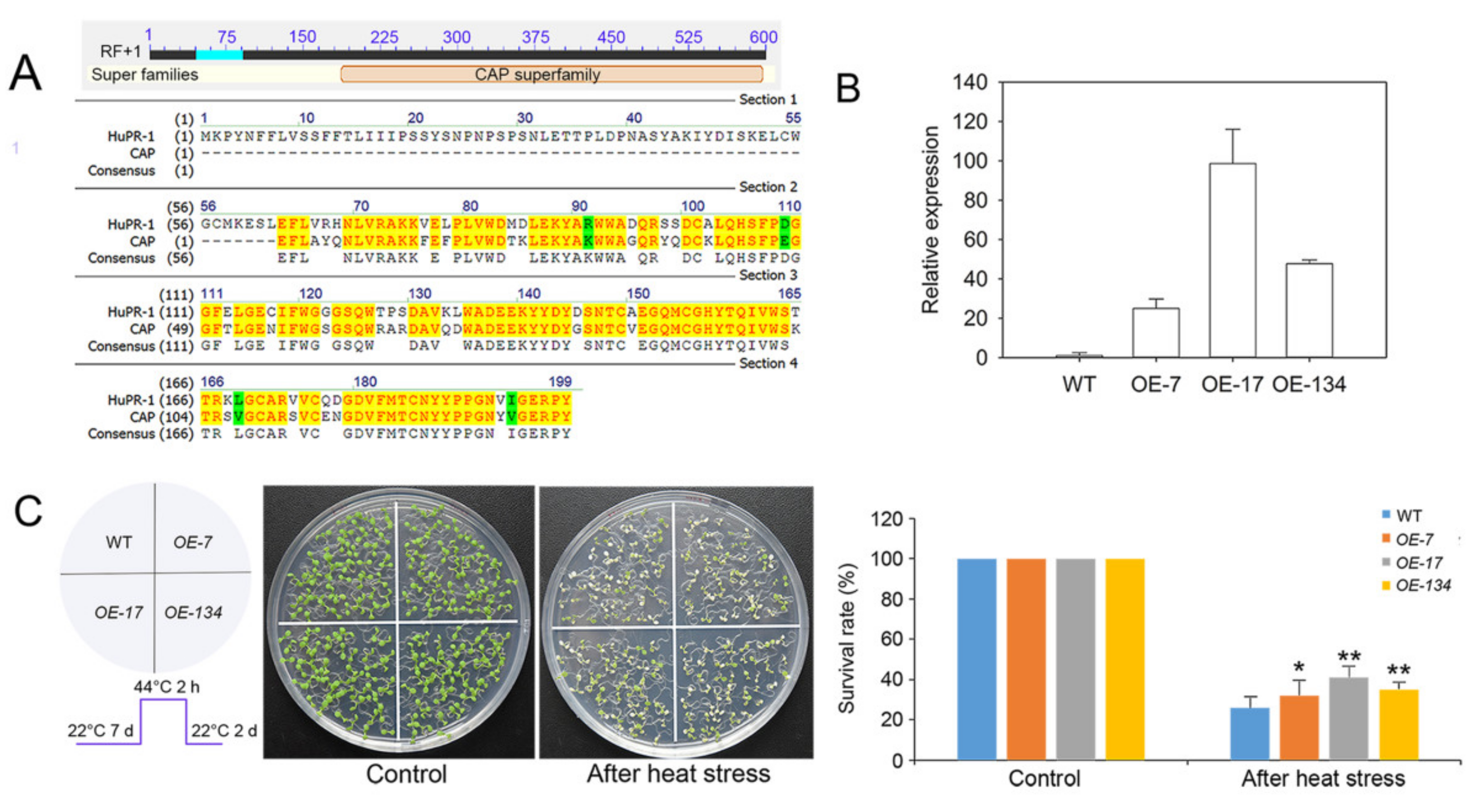
3.5. Overexpression of HuPR-1 in Arabidopsis Increased Heat Tolerance
4. Discussion
4.1. Roles of Heat-Responsive TFs
4.2. Mechanism Underlying the Response of Red Pitaya to HS
4.3. HuPR-1 Plays an Active Role in the Response to HS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.M.; Fischhoff, D.A.; Hodges, C.N.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D.; et al. Radically rethinking agriculture for the 21st century. Science 2010, 327, 833–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tubiello, F.N.; Soussana, J.F.; Howden, S.M. Crop and pasture response to climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19686–19690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vollenweider, P.; Gunthardt-Goerg, M.S. Diagnosis of abiotic and biotic stress factors using the visible symptoms in foliage. Environ. Pollut. 2005, 137, 455–465. [Google Scholar] [CrossRef]
- Guy, C. Molecular responses of plants to cold shock and cold acclimation. J. Mol. Microbiol. Biotechnol. 1999, 1, 231–242. [Google Scholar]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. Biochim. Biophys. Acta 2012, 1819, 104–119. [Google Scholar] [CrossRef]
- Maestri, E.; Klueva, N.; Perrotta, C.; Gulli, M.; Nguyen, H.T.; Marmiroli, N. Molecular genetics of heat tolerance and heat shock proteins in cereals. Plant Mol. Biol. 2002, 48, 667–681. [Google Scholar] [CrossRef]
- Sangwan, V.; Orvar, B.L.; Beyerly, J.; Hirt, H.; Dhindsa, R.S. Opposite changes in membrane fluidity mimic cold and heat stress activation of distinct plant MAP kinase pathways. Plant J. 2002, 31, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Barnabas, B.; Jager, K.; Feher, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef]
- Ruan, Y.L.; Jin, Y.; Yang, Y.J.; Li, G.J.; Boyer, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant 2010, 3, 942–955. [Google Scholar] [CrossRef]
- Zhuang, Y.L.; Zhang, Y.F.; Sun, L.P. Characteristics of fibre-rich powder and antioxidant activity of pitaya (Hylocereus undatus) peels. Int. J. Food Sci. Tech. 2012, 47, 1279–1285. [Google Scholar] [CrossRef]
- Adnan, L.; Osman, A.; Hamid, A.A. Antioxidant activity of different extracts of red pitaya (Hylocereus polyrhizus) seed. Int. J. Food Prop. 2011, 14, 1171–1181. [Google Scholar] [CrossRef] [Green Version]
- Lim, T.K. Edible Medicinal and Non-Medicinal Plants; Springer: Berlin/Heidelberg, Germany, 2012; Volume 1, pp. 643–655. [Google Scholar]
- Wybraniec, S.; Stalica, P.; Jerz, G.; Klose, B.; Gebers, N.; Winterhalter, P.; Spórna-Kucab, A.; Szaleniec, M.; Mizrahi, Y. Separation of polar betalain pigments from cacti fruits of Hylocereus polyrhizus by ion-pair high-speed countercurrent chromatography. J. Chromatogr. A 2009, 1216, 6890–6899. [Google Scholar] [CrossRef]
- Sogi, D.S.; Sharma, S.; Oberoi, D.P.S.; Wani, I.A. Effect of extraction parameters on curcumin yield from turmeric. J. Food Sci. Tech. 2010, 47, 300–304. [Google Scholar] [CrossRef] [Green Version]
- Woo, K.K.; Wong, F.N.F.; Chua, H.S.C.; Tang, P.Y. Stability of the spray-dried pigment of red dragon fruit [Hylocereus polyrhizus (Weber) Britton and Rose] as a function of organic acid additives and storage conditions. Philipp. Agric. Sci. 2011, 94, 264–269. [Google Scholar]
- Garcia-Cruz, L.; Valle-Guadarrama, S.; Salinas-Moreno, Y.; Joaquin-Cruz, E. Physical, chemical, and antioxidant activity characterization of Pitaya (Stenocereus pruinosus) fruits. Plant Food Hum. Nutr. 2013, 68, 403–410. [Google Scholar] [CrossRef]
- Suh, D.H.; Lee, S.; Heo, D.Y.; Kim, Y.S.; Cho, S.K.; Lee, S. Metabolite profiling of red and white pitayas (Hylocereus polyrhizus and Hylocereus undatus) for comparing betalain biosynthesis and antioxidant activity. J. Agr. Food Chem. 2014, 62, 8764–8771. [Google Scholar] [CrossRef]
- Hua, Q.Z.; Chen, C.J.; Chen, Z.; Chen, P.K.; Ma, Y.W.; Wu, J.Y.; Zheng, J.; Hu, G.; Zhao, J.; Qin, Y. Transcriptomic analysis reveals key genes related to betalain biosynthesis in pulp coloration of Hylocereus polyrhizus. Front. Plant Sci. 2016, 6, 1179. [Google Scholar]
- Zhao, Q.; Zhang, H.; Wang, T.; Chen, S.X.; Dai, S.J. Proteomics-based investigation of salt-responsive mechanisms in plant roots. J. Proteom. 2013, 82, 230–253. [Google Scholar] [CrossRef]
- Fan, Q.J.; Yan, F.X.; Qiao, G.; Zhang, B.X.; Wen, X.P. Identification of differentially-expressed genes potentially implicated in drought response in pitaya (Hylocereus undatus) by suppression subtractive hybridization and cDNA microarray analysis. Gene 2014, 533, 322–331. [Google Scholar] [CrossRef]
- Zhou, J.L.; Wang, Z.; Mao, Y.Y.; Wang, L.J.; Xiao, T.J.; Hu, Y.; Zhang, Y.; Ma, Y. Proteogenomic analysis of pitaya reveals cold stress-related molecular signature. PeerJ 2020, 8, e8540. [Google Scholar] [CrossRef] [PubMed]
- Li, A.L.; Wen, Z.; Yang, K.; Wen, X.P. Conserved miR396b-GRF regulation is involved in abiotic stress responses in pitaya (Hylocereus polyrhizus). Int. J. Mol. Sci. 2019, 20, 2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, Q.; Gao, G.-L.; Fan, Q.-J.; Qiao, G.; Wen, X.-P.; Liu, T.; Peng, Z.-J.; Cai, Y.-Q. Isolation and characterization of a catalase gene "HuCAT3" from pitaya (Hylocereus undatus) and its expression under abiotic stress. Gene 2015, 563, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.J.; Nong, Q.D.; Jian, S.G.; Lu, H.F.; Zhang, M.Y.; Xia, K.F. An AP2/ERF gene, HuERF1, from Pitaya (Hylocereus undatus) positively regulates salt tolerance. Int. J. Mol. Sci. 2020, 21, 4586. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R.; Finka, A.; Goloubinoff, P. How do plants feel the heat? Trends Biochem. Sci. 2012, 37, 118–125. [Google Scholar] [CrossRef]
- Xin, C.; Wang, X.; Cai, J.; Zhou, Q.; Liu, F.; Dai, T.; Cao, W.; Jiang, D. Changes of transcriptome and proteome are associated with the enhanced post-anthesis high temperature tolerance induced by pre-anthesis heat priming in wheat. Plant Growth Regul. 2016, 79, 135–145. [Google Scholar] [CrossRef]
- Mangelsen, E.; Kilian, J.; Harter, K.; Jansson, C.; Wanke, D.; Sundberg, E. Transcriptome analysis of high-temperature stress in developing barley caryopses: Early stress responses and effects on storage compound biosynthesis. Mol. Plant. 2011, 4, 97–115. [Google Scholar] [CrossRef]
- Qin, D.D.; Wu, H.Y.; Peng, H.R.; Yao, Y.Y.; Ni, Z.F.; Li, Z.X.; Zhou, C.; Sun, Q. Heat stress-responsive transcriptome analysis in heat susceptible and tolerant wheat (Triticum aestivum L.) by using wheat genome array. BMC Genom. 2008, 9, 432. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Liao, K.; Du, H.; Xu, Y.; Song, H.; Li, X.; Xiong, L. A stress-responsive NAC transcription factor SNAC3 confers heat and drought tolerance through modulation of reactive oxygen species in rice. J. Exp. Bot. 2015, 66, 6803–6817. [Google Scholar] [CrossRef] [Green Version]
- Akhter, D.; Qin, R.; Nath, U.K.; Eshag, J.; Jin, X.L.; Shi, C.H. A rice gene, OsPL, encoding a MYB family transcription factor confers anthocyanin synthesis, heat stress response and hormonal signaling. Gene 2019, 699, 62–72. [Google Scholar] [CrossRef]
- Li, S.J.; Fu, Q.T.; Huang, W.D.; Yu, D.Q. Functional analysis of an Arabidopsis transcription factor WRKY25 in heat stress. Plant Cell Rep. 2009, 28, 683–693. [Google Scholar] [CrossRef]
- Angelcheva, L.; Mishra, Y.; Antti, H.; Kjellsen, T.D.; Funk, C.; Strimbeck, R.G.; Schröder, W.P. Metabolomic analysis of extreme freezing tolerance in Siberian spruce (Picea obovata). New Phytol. 2014, 204, 545–555. [Google Scholar] [CrossRef]
- Yang, D.S.; Zhang, J.; Li, M.X.; Shi, L.X. Metabolomics analysis reveals the salt-tolerant mechanism in glycine soja. J. Plant Growth Regul. 2017, 36, 460–471. [Google Scholar] [CrossRef]
- Shulaev, V.; Cortes, D.; Miller, G.; Mittler, R. Metabolomics for plant stress response. Physiol. Plant 2008, 13, 199–208. [Google Scholar] [CrossRef]
- Nong, Q.D.; Zhang, M.Y.; Chen, J.T.; Zhang, M.; Cheng, H.P.; Jian, S.G.; Lu, H.; Xia, K. RNA-Seq de novo assembly of red pitaya (Hylocereus polyrhizus) roots and differential transcriptome analysis in response to salt stress. Trop. Plant Biol. 2019, 12, 55–66. [Google Scholar] [CrossRef]
- Fumagalli, M.; Vieira, F.G.; Linderoth, T.; Nielsen, R. ngsTools: Methods for population genetics analyses from next-generation sequencing data. Bioinformatics 2014, 30, 1486–1487. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Xia, K.F.; Ou, X.J.; Tang, H.D.; Wang, R.; Wu, P.; Jia, Y.X.; Wei, X.; Xu, X.; Kang, S.H.; Kim, S.K.; et al. Rice microRNA osa-miR1848 targets the obtusifoliol 14α-demethylase gene OsCYP51G3 and mediates the biosynthesis of phytosterols and brassinosteroids during development and in response to stress. New Phytol. 2015, 208, 790–802. [Google Scholar] [CrossRef] [Green Version]
- Nong, Q.D.; Yang, Y.C.; Zhang, M.Y.; Zhang, M.; Chen, J.T.; Jian, S.G.; Lu, H.; Xia, K. RNA-seq-based selection of reference genes for RT-qPCR analysis of pitaya. FEBS Open Bio 2019, 9, 1403–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.S.; Ding, Y.F.; Zhu, C. Sensitivity and responses of chloroplasts to heat stress in plants. Front. Plant Sci. 2020, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.G.; Lu, Z.G.; Wang, L.; Jin, B. Plant responses to heat stress: Physiology, transcription, noncoding RNAs, and epigenetics. Int. J. Mol. Sci. 2021, 22, 117. [Google Scholar]
- Wu, Z.G.; Jiang, W.; Chen, S.L.; Mantri, N.; Tao, Z.M.; Jiang, C.X. Insights from the cold transcriptome and metabolome of dendrobium officinale: Global reprogramming of metabolic and gene regulation networks during cold acclimation. Front. Plant Sci. 2016, 7, e0121658. [Google Scholar] [CrossRef] [Green Version]
- Zou, M.Q.; Yuan, L.Y.; Zhu, S.D.; Liu, S.; Ge, J.T.; Wang, C.G. Response of osmotic adjustment and ascorbate-glutathione cycle to heat stress in a heat-sensitive and a heat-tolerant genotype of wucai (Brassica campestris L.). Sci. Hortic. 2016, 211, 87–94. [Google Scholar] [CrossRef]
- Matsukura, S.; Mizoi, J.; Yoshida, T.; Todaka, D.; Ito, Y.; Maruyama, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes. Mol. Genet. Genom. 2010, 283, 185–196. [Google Scholar] [CrossRef]
- Qin, F.; Kakimoto, M.; Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Tran, L.S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Regulation and functional analysis of ZmDREB2A in response to drought and heat stresses in Zea mays L. Plant J. 2010, 50, 54–69. [Google Scholar] [CrossRef]
- Cai, H.Y.; Yang, S.; Yan, Y.; Xiao, Z.L.; Cheng, J.B.; Wu, J.; Qiu, A.L.; Lai, Y.; Mou, S.L.; Guan, D.Y. CaWRKY6 transcriptionally activates CaWRKY40, regulates Ralstonia solanacearum resistance, and confers high-temperature and high-humidity tolerance in pepper. J. Exp. Bot. 2015, 66, 3163–3174. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.L.; Shiroto, Y.; Kishitani, S.; Ito, Y.; Toriyama, K. Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Rep. 2009, 28, 21–30. [Google Scholar] [CrossRef]
- Guo, W.W.; Zhang, J.X.; Zhang, N.; Xin, M.M.; Peng, H.R.; Hu, Z.R.; Ni, Z.F.; Du, J.K. The wheat NAC transcription factor TaNAC2L is regulated at the transcriptional and post-translational levels and promotes heat stress tolerance in transgenic Arabidopsis. PLoS ONE 2015, 10, e0135667. [Google Scholar] [CrossRef]
- Suzuki, N.; Sejima, H.; Tam, R.; Schlauch, K.; Mittler, R. Identification of the MBF1 heat-response regulon of Arabidopsis thaliana. Plant J. 2011, 66, 844–851. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.P.; Andreasson, E.; Maslak, A.; Mock, H.P.; Mattsson, O.; Mundy, J. Arabidopsis MYB68 in development and responses to environmental cues. Plant Sci. 2004, 167, 1099–1107. [Google Scholar] [CrossRef]
- Meng, X.; Wang, J.R.; Wang, G.D.; Liang, X.Q.; Li, X.D.; Meng, Q.W. An R2R3-MYB gene, LeAN2, positively regulated the thermo-tolerance in transgenic tomato. J. Plant Physiol. 2015, 175, 1–8. [Google Scholar] [CrossRef]
- El-kereamy, A.; Bi, Y.M.; Ranathunge, K.; Beatty, P.H.; Good, A.G.; Rothstein, S.J. The rice R2R3-MYB transcription factor OsMYB55 is involved in the tolerance to high temperature and modulates amino acid metabolism. PLoS ONE 2012, 7, e52030. [Google Scholar] [CrossRef] [Green Version]
- Zhaorong, H.; Tian, X.; Wang, F.; Zhang, L.; Xin, M.; Hu, Z.; Yao, Y.; Ni, Z.; Sun, Q.; Peng, H. Characterization of wheat MYB genes responsive to high temperatures. BMC Plant Biol. 2017, 17, 208. [Google Scholar]
- Liao, C.C.; Zheng, Y.; Guo, Y. MYB30 transcription factor regulates oxidative and heat stress responses through ANNEXIN-mediated cytosolic calcium signaling in Arabidopsis. New Phytol. 2017, 216, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Montero-Barrientos, M.; Hermosa, R.; Cardoza, R.E.; Gutierrez, S.; Nicolas, C.; Monte, E. Transgenic expression of the Trichoderma harzianum hsp70 gene increases Arabidopsis resistance to heat and other abiotic stresses. J. Plant Physiol. 2010, 167, 659–665. [Google Scholar] [CrossRef]
- Qi, Y.C.; Wang, H.J.; Zou, Y.; Liu, C.; Liu, Y.Q.; Wang, Y.; Zhang, W. Over-expression of mitochondrial heat shock protein 70 suppresses programmed cell death in rice. FEBS Lett. 2011, 585, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Kotak, S.; Larkindale, J.; Lee, U.; von Koskull-Doring, P.; Vierling, E.; Scharf, K.D. Complexity of the heat stress response in plants. Curr. Opin. Plant Biol. 2007, 10, 310–316. [Google Scholar] [CrossRef]
- Penfield, S. Temperature perception and signal transduction in plants. New Phytol. 2008, 179, 615–628. [Google Scholar] [CrossRef]
- Zhang, L.R.; Li, Y.S.; Xing, D.; Gao, C.J. Characterization of mitochondrial dynamics and subcellular localization of ROS reveal that HsfA2 alleviates oxidative damage caused by heat stress in Arabidopsis. J. Exp. Bot. 2009, 60, 2073–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banti, V.; Mafessoni, F.; Loreti, E.; Alpi, A.; Perata, P. The heat-inducible transcription factor HsfA2 enhances anoxia tolerance in Arabidopsis. Plant Physiol. 2010, 152, 1471–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaya, H.; Shibahara, K.; Taoka, K.; Iwabuchi, M.; Stillman, B.; Araki, T. FASCIATA genes for chromatin assembly factor-1 in Arabidopsis maintain the cellular organization of apical meristems. Cell 2001, 104, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Rivero, R.M.; Ruiz, J.M.; Garcia, P.C.; Lopez-Lefebre, L.R.; Sanchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef]
- Ogweno, J.O.; Song, X.S.; Shi, K.; Hu, W.H.; Mao, W.H.; Zhou, Y.H.; Yu, J.Q.; Nogués, S. Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems in Lycopersicon esculentum. J. Plant Growth Regul. 2008, 27, 49–57. [Google Scholar] [CrossRef]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiol. Biochem. 2009, 47, 1–8. [Google Scholar] [CrossRef]
- Akbudak, M.A.; Yildiz, S.; Filiz, E. Pathogenesis related protein-1 (PR-1) genes in tomato (Solanum lycopersicum L.): Bioinformatics analyses and expression profiles in response to drought stress. Genomics 2020, 112, 4089–4099. [Google Scholar] [CrossRef]
- Wang, J.; Mao, X.; Wang, R.; Li, A.; Zhao, G.; Zhao, J.; Jing, R. Identification of wheat stress-responding genes and TaPR-1-1 function by screening a cDNA yeast library prepared following abiotic stress. Sci. Rep. 2019, 9, 141. [Google Scholar] [CrossRef]
- Liu, W.X.; Zhang, F.C.; Zhang, W.Z.; Song, L.F.; Wu, W.H.; Chen, Y.F. Arabidopsis Di19 functions as a transcription factor and modulates PR1, PR2, and PR5 expression in response to drought stress. Mol. Plant 2013, 6, 1487–1502. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Statistic Value |
|---|---|
| Total sequences | 36,842 |
| Total bases | 78,871,914 |
| Min sequence length | 206 |
| Max sequence length | 40,937 |
| Average sequence length | 2141.82 |
| Median sequence length | 1749.00 |
| N25 length | 4210 |
| N50 length | 2848 |
| N75 length | 1799 |
| N90 length | 1107 |
| N95 length | 827 |
| As | 29.40% |
| Ts | 29.33% |
| Gs | 20.79% |
| Cs | 20.48% |
| (A + T)s | 58.72% |
| (G + C)s | 41.28% |
| Ns | 0.00% |
| Gene ID | Log2 FC | p-Value | Annotation | |
|---|---|---|---|---|
| Contig19129 | 7.63 | 3.08 × 10−6 | GRXC11; glutathione-disulfide oxidoreductase activity in the presence of NADPH and glutathione reductase. | up |
| Contig20825 | 7.26 | 2.29 × 10−5 | NA | up |
| Contig32725 | 5.01 | 3.76 × 10−10 | FER2, chloroplastic; stores iron in a soluble, non-toxic, readily available form. Important for iron homeostasis. Has ferroxidase activity. | up |
| Contig32724 | 4.98 | 1.03 × 10−9 | N/A, chloroplastic; stores iron in a soluble, non-toxic, readily available form. Important for iron homeostasis. Has ferroxidase activity. | up |
| Contig21847 | 4.74 | 1.61 × 10−8 | PFE, chloroplastic; stores iron in a soluble, non-toxic, readily available form. Important for iron homeostasis. | up |
| Contig2380 | 4.67 | 6.45 × 10−14 | ELIP1, chloroplastic; early light-induced protein 1; prevents excess accumulation of free chlorophyll by inhibiting the entire chlorophyll biosynthesis pathway, and then prevents photooxidative stress; involved in seed germination | up |
| Contig785 | 4.63 | 1.25 × 10−10 | PFK3; catalyzes the phosphorylation of D-fructose 6-phosphate to fructose 1,6-bisphosphate by ATP, the first committing step of glycolysis. | up |
| Contig9300 | 4.41 | 9.1 × 10−4 | NA | up |
| Contig3696 | 4.39 | 1.63 × 10−21 | PGM1, 2,3-bisphosphoglycerate-independent phosphoglycerate mutase; catalyzes the interconversion of 2-phosphoglycerate and 3-phosphoglycerate. | up |
| Contig9996 | 4.28 | 9.30 × 10−6 | NA | up |
| Contig9364 | −5.29 | 4.08 × 10−5 | ART2; encoded on the antisense strand of the nuclear 25S rDNA. | down |
| Contig17684 | −5.49 | 6.21 × 10−5 | PER72, peroxidase 72; removal of H2O2; oxidation of toxic reductants; biosynthesis and degradation of lignin; suberization; auxin catabolism; response to environmental stresses such as wounding, pathogen attack, and oxidative stress. | down |
| Contig7023 | −5.51 | 1.27 × 10−8 | NA | down |
| Contig2725 | −6.29 | 3.73 × 10−18 | rbgA, ribosome biogenesis; GTPase A; essential protein that is required for a late step of 50S ribosomal subunit assembly. | down |
| Contig16950 | −6.58 | 1.09 × 10−7 | NA | down |
| Contig18702 | −6.91 | 6.00 × 10−4 | ALMT2, aluminum-activated malate transporter 2. | down |
| Contig31875 | −7.22 | 1.92 × 10−31 | LBD41, LOB domain-containing protein 41; cellular response to hypoxia; regulation of transcription. | down |
| Contig29974 | −8.03 | 2.66 × 10−5 | ALMT10, aluminum-activated malate transporter 10. | down |
| Contig20699 | −8.98 | 1.78 × 10−12 | RAB15, glycine-rich RNA-binding, abscisic acid-inducible protein. | down |
| Contig352 | −10.55 | 4.34 × 10−18 | NA | down |
| Gene ID | Log2 FC | Functional Description | Homology | Species |
|---|---|---|---|---|
| Contig9674 | −1.66 | MYB domain-containing protein | MYBB | Xenopus laevis |
| Contig26304 | −1.58 | MYB domain-containing protein | MYBB | Xenopus laevis |
| Contig25078 | −2.38 | MYB-related protein 306 | MYB06 | Antirrhinum majus |
| Contig25077 | −2.70 | MYB-related protein 306 | MYB06 | Antirrhinum majus |
| Contig25252 | 1.66 | MYB domain protein 30 | MYB30 | Antirrhinum majus |
| Contig28175 | −1.79 | MYB domain protein 44 | MYB44 | Arabidopsis thaliana |
| Contig28176 | −1.77 | MYB domain protein 44 | MYB44 | Arabidopsis thaliana |
| Contig21877 | −1.37 | MYB domain protein 86 | MYB86 | Arabidopsis thaliana |
| Gene ID | Log2 FC | Functional Description | Homology | Species |
|---|---|---|---|---|
| Contig26416 | 1.86 | Heat shock factor A2 | HsfA2 | Arabidopsis thaliana |
| Contig26417 | 2.54 | Heat shock factor A2 | HsfA2 | Arabidopsis thaliana |
| Contig27952 | 1.67 | Heat shock factor R2 | HspR2 | Arabidopsis thaliana |
| Contig20486 | 1.13 | Heat shock protein 83 | Hsp83 | D. melanogaster |
| Contig1864 | 3.72 | Heat shock protein 16 | Hsp16 | Glycine max |
| Contig1898 | 4.91 | Heat-shock cognate 71 kDa protein | Hsp7C | Petunia hybrida |
| Contig1899 | 1.43 | Heat-shock cognate 71 kDa protein | Hsp7C | Petunia hybrida |
| Contig3506 | 2.20 | Heat-shock cognate 71 kDa protein | Hsp7C | Petunia hybrida |
| Contig13687 | 3.89 | Heat shock protein 41 | Hsp41 | Glycine max |
| Contig1021 | 4.67 | Heat shock protein 83 | Hsp83 | Ipomoea nil |
| Contig1022 | 2.34 | Heat shock protein 83 | Hsp83 | Ipomoea nil |
| Contig1023 | 2.27 | Heat shock protein 83 | Hsp83 | Ipomoea nil |
| Contig27132 | 6.29 | Heat-shock cognate 71 kDa protein | Hsp7C | Rattus norvegicus |
| Contig22611 | 1.37 | 70 kDa heat shock-related protein | Hsp7S | Pisum sativum |
| Contig20223 | −1.94 | heat shock 70 kDa protein cognate 5 | Hsp7E | Spinacia oleracea |
| Contig26216 | −1.31 | Heat shock protein 72 | Hsp72 | Solanum lycopersicum |
| Contig14935 | 4.25 | 22.0 kDa class IV heat shock protein | Hsp22 | Ipomoea nil |
| Contig13847 | 1.59 | Heat shock protein 70 | Hsp70 | Arabidopsis thaliana |
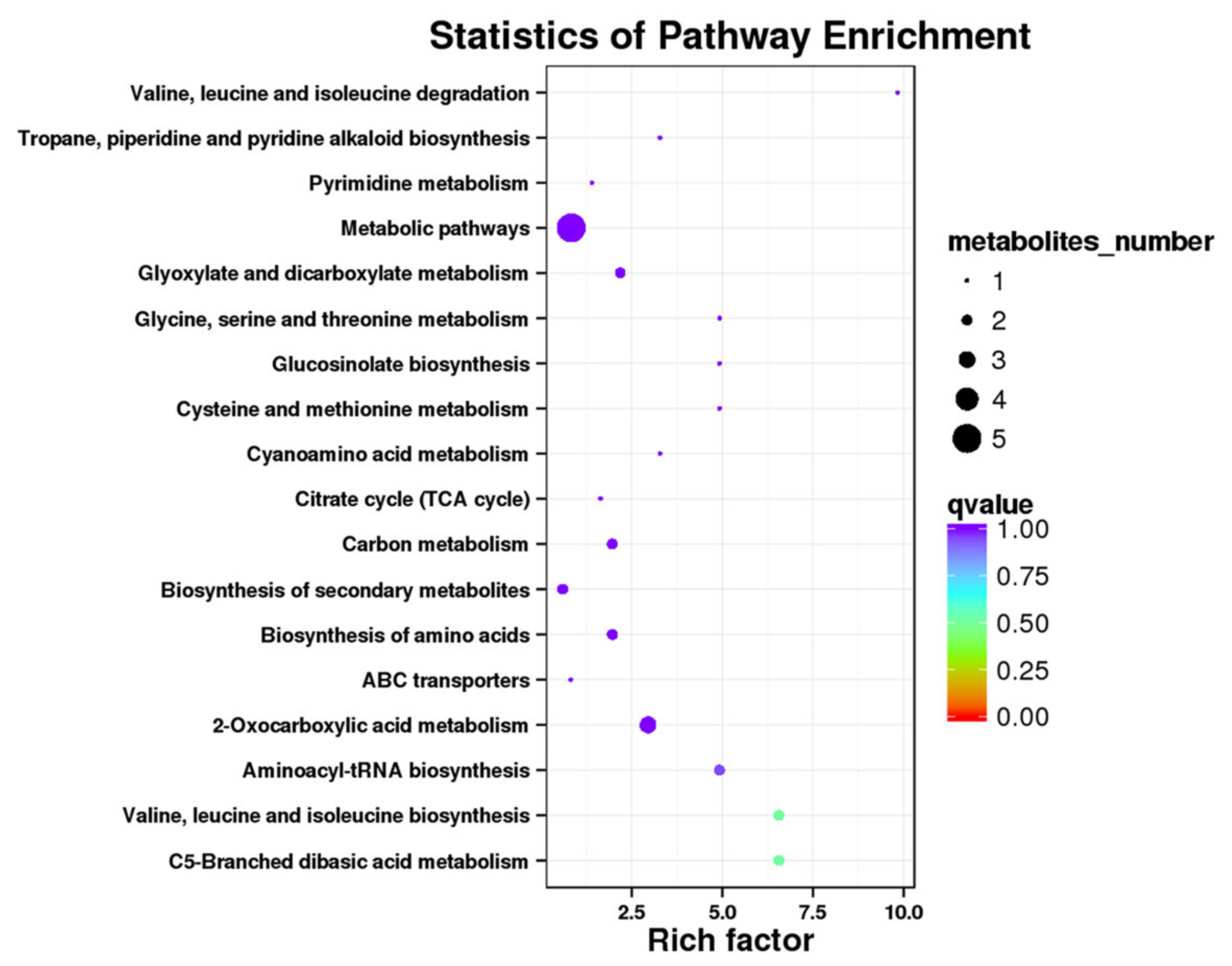
| ID a | Term | Diff_Metabolites | Metabolite_id |
|---|---|---|---|
| ko00970 | Aminoacyl-tRNA biosynthesis | 2 | meta_51; meta_10 |
| ko00630 | Glyoxylate and dicarboxylate metabolism | 2 | meta_40; meta_8 |
| ko00960 | Tropane, piperidine, and pyridine alkaloid biosynthesis | 1 | meta_10 |
| ko00460 | Cyanoamino acid metabolism | 1 | meta_10 |
| ko00020 | Citrate cycle (TCA cycle) | 1 | meta_40 |
| ko00240 | Pyrimidine metabolism | 1 | meta_212 |
| ko00966 | Glucosinolate biosynthesis | 1 | meta_10 |
| ko01200 | Carbon metabolism | 2 | meta_51; meta_8 |
| ko00660 | C5-Branched dibasic acid metabolism | 2 | meta_8; meta_40 |
| ko01110 | Biosynthesis of secondary metabolites | 2 | meta_10; meta_40 |
| ko00260 | Glycine, serine, and threonine metabolism | 1 | meta_51 |
| ko02010 | ABC transporters | 1 | meta_10 |
| ko00280 | Valine, leucine, and isoleucine degradation | 1 | meta_10 |
| ko01210 | 2-Oxocarboxylic acid metabolism | 3 | meta_8; meta_10; meta_40 |
| ko00290 | Valine, leucine, and isoleucine biosynthesis | 2 | meta_10; meta_8 |
| ko01230 | Biosynthesis of amino acids | 2 | meta_51; meta_10 |
| ko00270 | Cysteine and methionine metabolism | 1 | meta_51 |
| ko01100 | Metabolic pathways | 5 | meta_212; meta_8; meta_51; meta_40; meta_10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, Z.; Xu, W.; Nong, Q.; Zhang, M.; Jian, S.; Lu, H.; Chen, J.; Zhang, M.; Xia, K. An Integrative Transcriptomic and Metabolomic Analysis of Red Pitaya (Hylocereus polyrhizus) Seedlings in Response to Heat Stress. Genes 2021, 12, 1714. https://doi.org/10.3390/genes12111714
Jiao Z, Xu W, Nong Q, Zhang M, Jian S, Lu H, Chen J, Zhang M, Xia K. An Integrative Transcriptomic and Metabolomic Analysis of Red Pitaya (Hylocereus polyrhizus) Seedlings in Response to Heat Stress. Genes. 2021; 12(11):1714. https://doi.org/10.3390/genes12111714
Chicago/Turabian StyleJiao, Zhengli, Weijuan Xu, Quandong Nong, Mei Zhang, Shuguang Jian, Hongfang Lu, Jiantong Chen, Mingyong Zhang, and Kuaifei Xia. 2021. "An Integrative Transcriptomic and Metabolomic Analysis of Red Pitaya (Hylocereus polyrhizus) Seedlings in Response to Heat Stress" Genes 12, no. 11: 1714. https://doi.org/10.3390/genes12111714
APA StyleJiao, Z., Xu, W., Nong, Q., Zhang, M., Jian, S., Lu, H., Chen, J., Zhang, M., & Xia, K. (2021). An Integrative Transcriptomic and Metabolomic Analysis of Red Pitaya (Hylocereus polyrhizus) Seedlings in Response to Heat Stress. Genes, 12(11), 1714. https://doi.org/10.3390/genes12111714