
Population Genetic Structures of Puccinia striiformis f. sp. tritici in the Gansu-Ningxia Region and Hubei Province, China

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
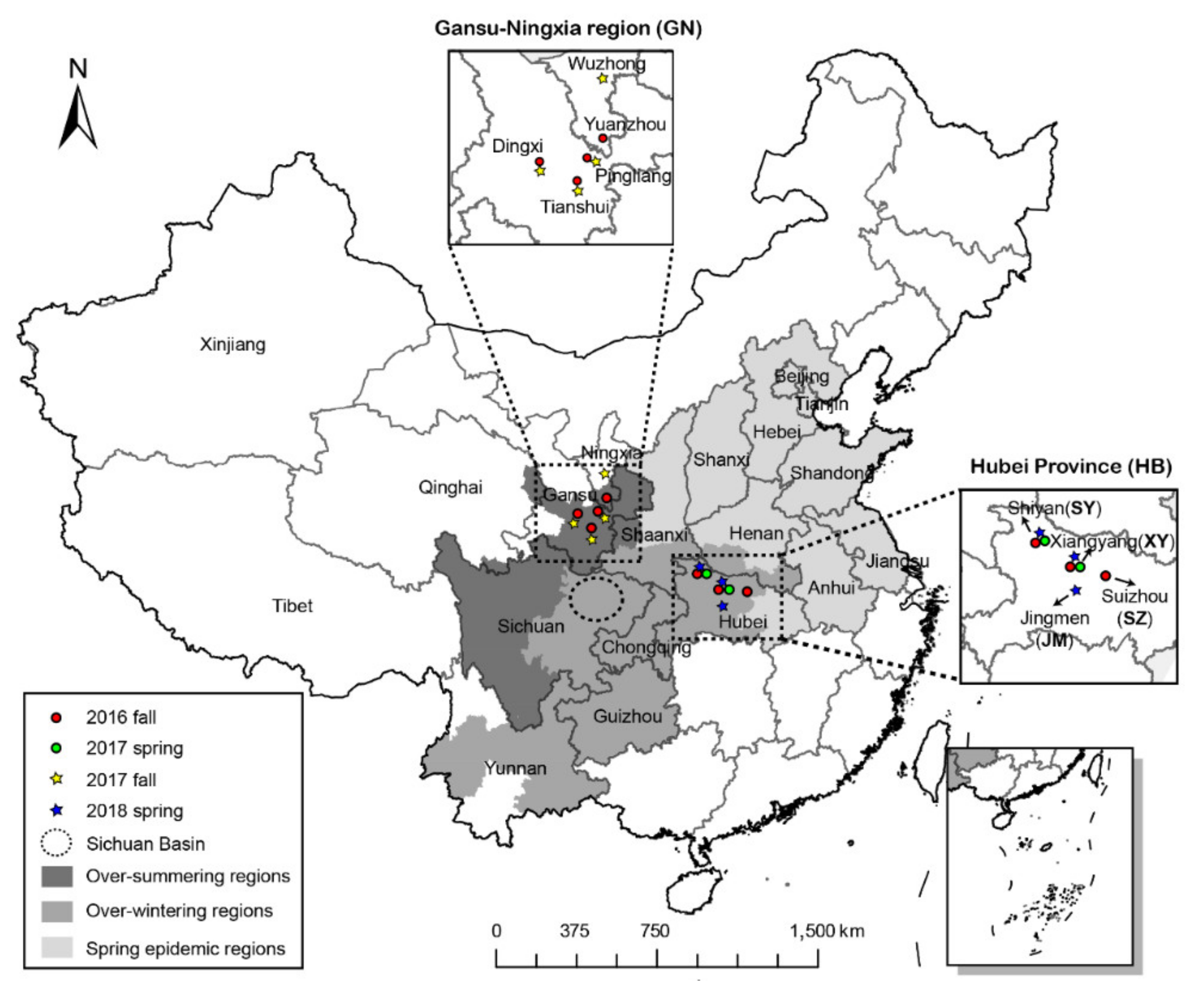
2.1. Sampling and Reproduction of the Isolates
2.2. DNA Extraction and Simple Sequence Repeat (SSR) Genotyping
2.3. Data Analysis
3. Results
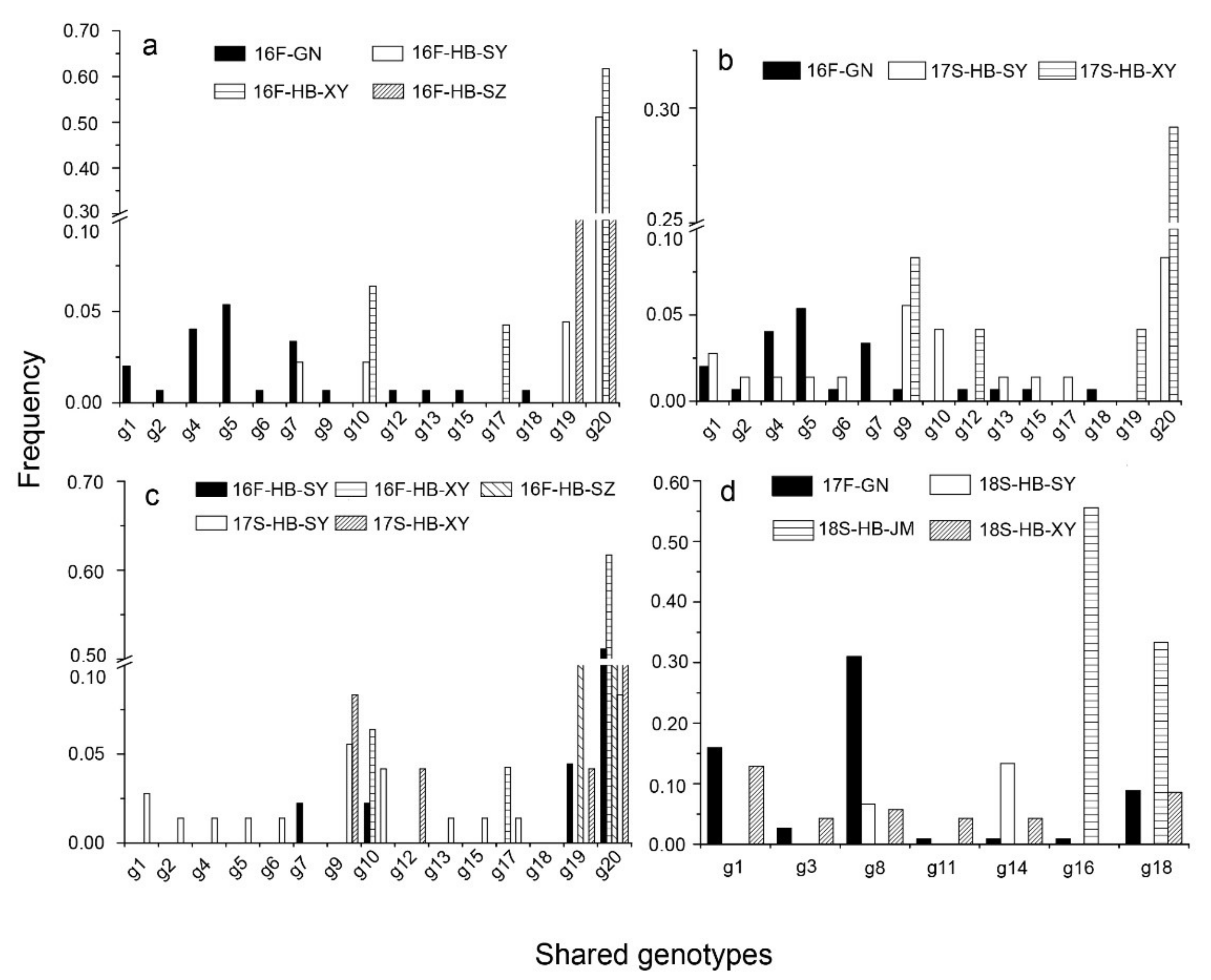
3.1. Genotypic Diversity
3.2. Genetic Diversity
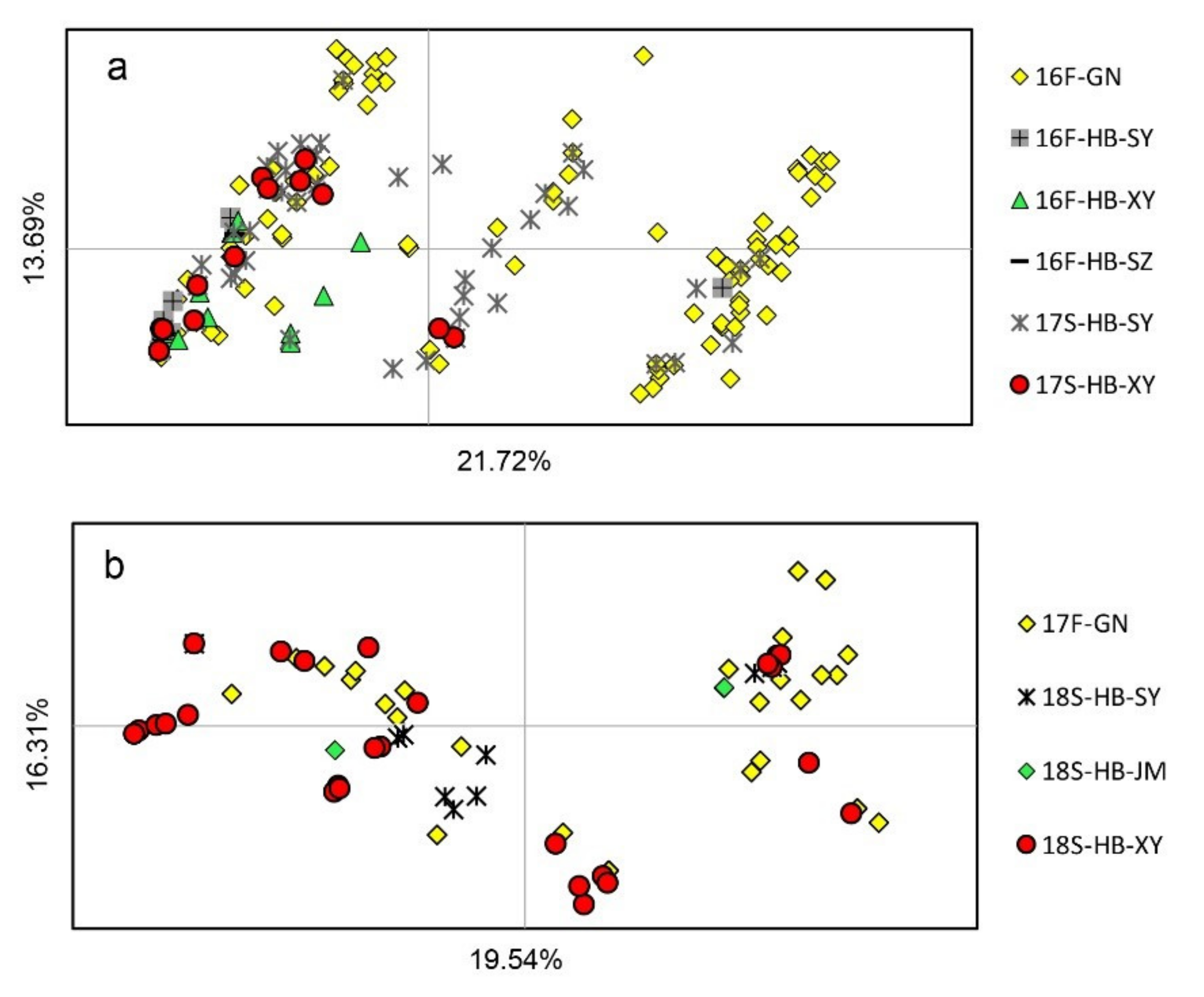
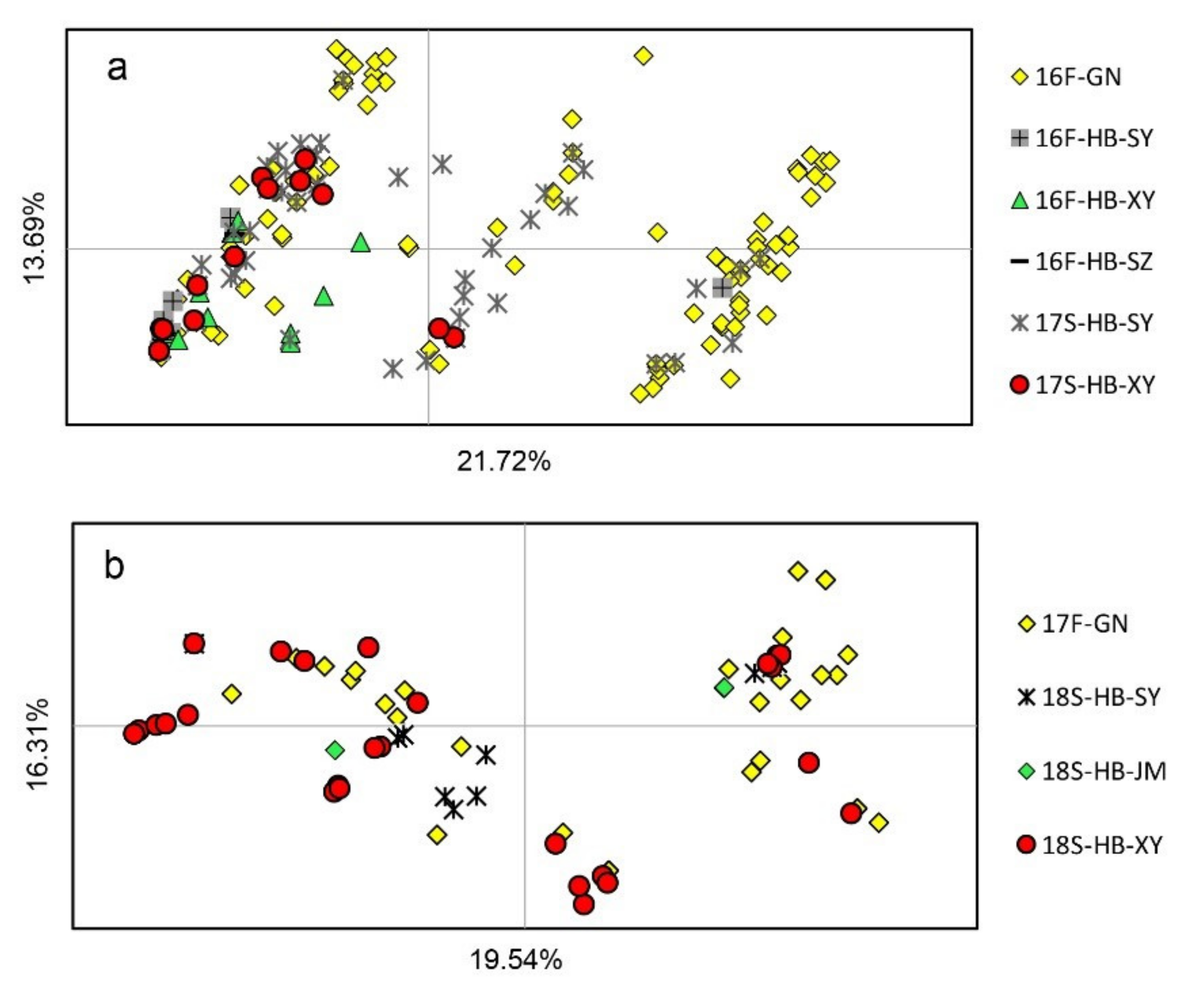
3.3. Population Subdivision on Spatial and Temporal Scales
3.4. LD Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, X. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Hovmøller, M.S.; Walter, S.; Justesen, A.F. Escalating threat of wheat rusts. Science 2010, 329, 369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Gladieux, P.; Leconte, M.; Gautier, A.; Justesen, A.F.; Hovmøller, M.S.; Enjalbert, J.; de Vallavieille-Pope, C. Origin, migration routes and worldwide population genetic structure of the wheat yellow rust pathogen Puccinia striiformis f. sp. tritici. PLoS Pathog. 2014, 10, e1003903. [Google Scholar] [CrossRef] [Green Version]
- Wan, A.; Zhao, Z.; Chen, X.; He, Z.; Jin, S.; Jia, Q.; Yao, G.; Yang, J.; Wang, B.; Li, G.; et al. Wheat stripe rust epidemic and virulence of Puccinia striiformis f. sp. tritici in China in 2002. Plant Dis. 2004, 88, 896–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, S.; Luo, Y. Long-distance spread and interregional epidemics of wheat stripe rust in China. Plant Dis. 2006, 90, 980–988. [Google Scholar] [CrossRef] [Green Version]
- Wan, A.; Chen, X.; He, Z. Wheat stripe rust in China. Aust. J. Agric. Res. 2007, 58, 605–619. [Google Scholar] [CrossRef]
- Huang, C.; Jiang, Y.; Ji, G.; Zhang, G.; Li, H.; Li, Y. Spatiotemporal dynamics of wheat stripe rust epidemics at regional level in China in 2017 (in Chinese with English abstract). J. Plant Prot. 2018, 45, 20–26. [Google Scholar] [CrossRef]
- Chen, W.; Kang, Z.; Ma, Z.; Xu, S.; Jin, S.; Jiang, Y. Integrated management of wheat stripe rust caused by Puccinia striiformis f. sp. tritici in China (in Chinese with English abstract). Sci. Agric. Sin. 2013, 46, 4254–4262. [Google Scholar] [CrossRef]
- Li, Z.; Zeng, S. Wheat Rusts in China; China Agricultural Press: Beijing, China, 2002; p. 379. [Google Scholar]
- Liang, J.; Wan, Q.; Luo, Y.; Ma, Z. Population genetic structures of Puccinia striiformis in Ningxia and Gansu Provinces of China. Plant Dis. 2013, 97, 501–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Yang, Q.; Gong, G.; Zhang, L.; Wang, C.; Li, X. Study on the occurrence of wheat stripe rust in the northwest of Hubei Province. Hubei Plant Prot. 2003, 3, 4–6. [Google Scholar]
- Ye, Q.; Xiao, N.; Zhang, F.; Cai, G.; Yang, L.; Xiang, L. Study on oversummering and overwintering of Puccinia striiformis f. sp. tritici in Shiyan, Hubei. (in Chinese with English abstract). J. Trit. Crop. 2015, 35, 1306–1313. [Google Scholar] [CrossRef]
- Liu, B.; Liu, T.; Zhang, Z.; Jia, Q.; Wang, B.; Gao, L.; Peng, Y.; Jin, S.; Chen, W. Discovery and pathogenicity of CYR34,a new race of Puccinia striiformis f. sp. tritici in China. Acta Phytopathol. Sinia 2017, 47, 681–687. [Google Scholar] [CrossRef]
- Meng, Y.; Yang, C.; Jiang, S.; Huang, L.; Kang, Z.; Zhan, G. Development and evaluation of SNP molecular markers of wheat stripe rust based on KASP technology. J. Plant Prot. 2020, 47, 65–73. [Google Scholar] [CrossRef]
- Liang, J.; Liu, X.; Li, Y.; Wan, Q.; Ma, Z.; Luo, Y. Population genetic structure and the migration of Puccinia striiformis f. sp. tritici between the Gansu and Sichuan Basin populations of China. Phytopathology 2016, 106, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Q.; Liang, J.; Luo, Y.; Ma, Z. Population genetic structure of Puccinia striiformis in northwestern China. Plant Dis. 2015, 99, 1764–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Zheng, W.; Buchenauer, H.; Huang, L.; Lu, N.; Kang, Z. Isolation of microsatellite loci from expressed sequence tag library of Puccinia striiformis f. sp. tritici. Mol. Ecol. Notes 2009, 9, 236–238. [Google Scholar] [CrossRef]
- Enjalbert, J.; Duan, X.; Giraud, T.; Vautrin, D.; de Vallavieille-Pope, C.; Solignac, M. Isolation of twelve microsatellite loci, using an enrichment protocol, in the phytopathogenic fungus Puccinia. striiformis f. sp. tritici. Mol. Ecol. Notes 2002, 2, 563–565. [Google Scholar] [CrossRef]
- Bahri, B.; Leconte, M.; de Vallavieille-Pope, C.; Enjalbert, J. Isolation of ten microsatellite loci in an EST library of the phytopathogenic fungus Puccinia striiformis f. sp. tritici. Conserv. Genet. 2009, 10, 1425–1428. [Google Scholar] [CrossRef]
- Zhan, G.; Wang, F.; Luo, H.; Jiang, S.; Zheng, W.; Huang, L.; Kang, Z. Screening for simple sequence repeat markers in Puccinia striiformis tritici based on genomic sequence. J. Zhejiang Univ. Sci. B. 2015, 16, 727–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulce, D.; Li, X.; Snyder-Leiby, T.; Liu, C.J. GeneMarker® genotyping software: Tools to increase the statistical power of DNA fragment analysis. J. Biomol. Tech. 2011, 22, 35. [Google Scholar]
- Kamvar, Z.N.; Tabima, J.F.; Grunwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. Peer J. 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Arnaud-Haond, S.; Belkhir, K. GenClone: A computer program to analyze genotypic data, test for clonality and describe spatial clonal organization. Mol. Ecol. Notes 2007, 7, 15–17. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.; Boyle, T.; Ye, Z.; Mao, J. POPGENE, the User-Friendly Shareware for Population Genetic Analysis. Mol. Biol. Biotechnol. Cent. Univ. Alta. Can. 1997, 10, 295–301. [Google Scholar]
- Nei, M. Genetic distance between populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals 439 using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, N.A. Distruct: A program for the graphical display of population structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Agapow, P.M.; Burt, A. Indices of multilocus linkage disequilibrium. Mol. Ecol. Notes 2001, 1, 101–102. [Google Scholar] [CrossRef]
- Liu, W.; Han, L. Effects of 2016/2017 winter climate on agricultural production. (in Chinese with English abstract). Chin. J. Agrom. 2017, 38, 330–331. [Google Scholar] [CrossRef]
- Wan, A.; Zhang, Z.; Jin, S.; Yu, D.; Zhao, Z.; Shi, S. Investigation on the over-summering of Pccinia stiriiformis f. sp. tritici in the north-western mountain area of Hubei Province of China. Acta Phytopathol. Sin. 2004, 34, 90–92. [Google Scholar] [CrossRef]
- Huang, C.; Yang, L.; Zhang, J.; Yu, Q.; Chen, F. Analysis on the new features of wheat stripe rust and its causes in Xiangyang wheat area (in Chinese with English abstract). J. Hubei Agric. Sci. 2011, 50, 5098–5100. [Google Scholar] [CrossRef]
- Wang, C.; Li, L.; Jiang, B.; Zhang, K.; Chu, B.; Luo, Y.; Ma, Z. Genetic diversity and population structure of Puccinia striiformis f. sp. tritici reveal its migration from central to eastern China. Crop Prot. 2020, 128, 104974. [Google Scholar] [CrossRef]
- Xie, S.; Wang, K.; Chen, Y.; Chen, W. Preliminary studies on the relationship between transport of wheat stripe rust and the upper air current in China (in Chinese with English abstract). Acta Phytopathol. Sin. 1993, 23, 203–209. [Google Scholar]
- Hu, X.; Ma, L.; Liu, T.; Wang, C.; Peng, Y.; Pu, Q.; Xu, X. Population genetic analysis of Puccinia striiformis f. sp. tritici suggests two distinct populations in Tibet and the other regions of China. Plant Dis. 2017, 101, 288–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, M.; Jia, M.; Lu, H.; Shang, H.; Kang, Z.; Yang, S.; Sun, S.; Shi, Y. Study on the epidemiology and control strategy of wheat stripe rust in Longnan of Gansu. Acta Univ. Agric. Boreali-Occident. 1997, 25, 1–5. [Google Scholar]
- Blanco-bercial, L.; Buckl, A. New view of population genetics of zooplankton: RAD seq analysis reveals population structure of the North Atlantic planktonic copepod Centropages typicus. Mol. Ecol. 2016, 25, 1566–1580. [Google Scholar] [CrossRef] [PubMed]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
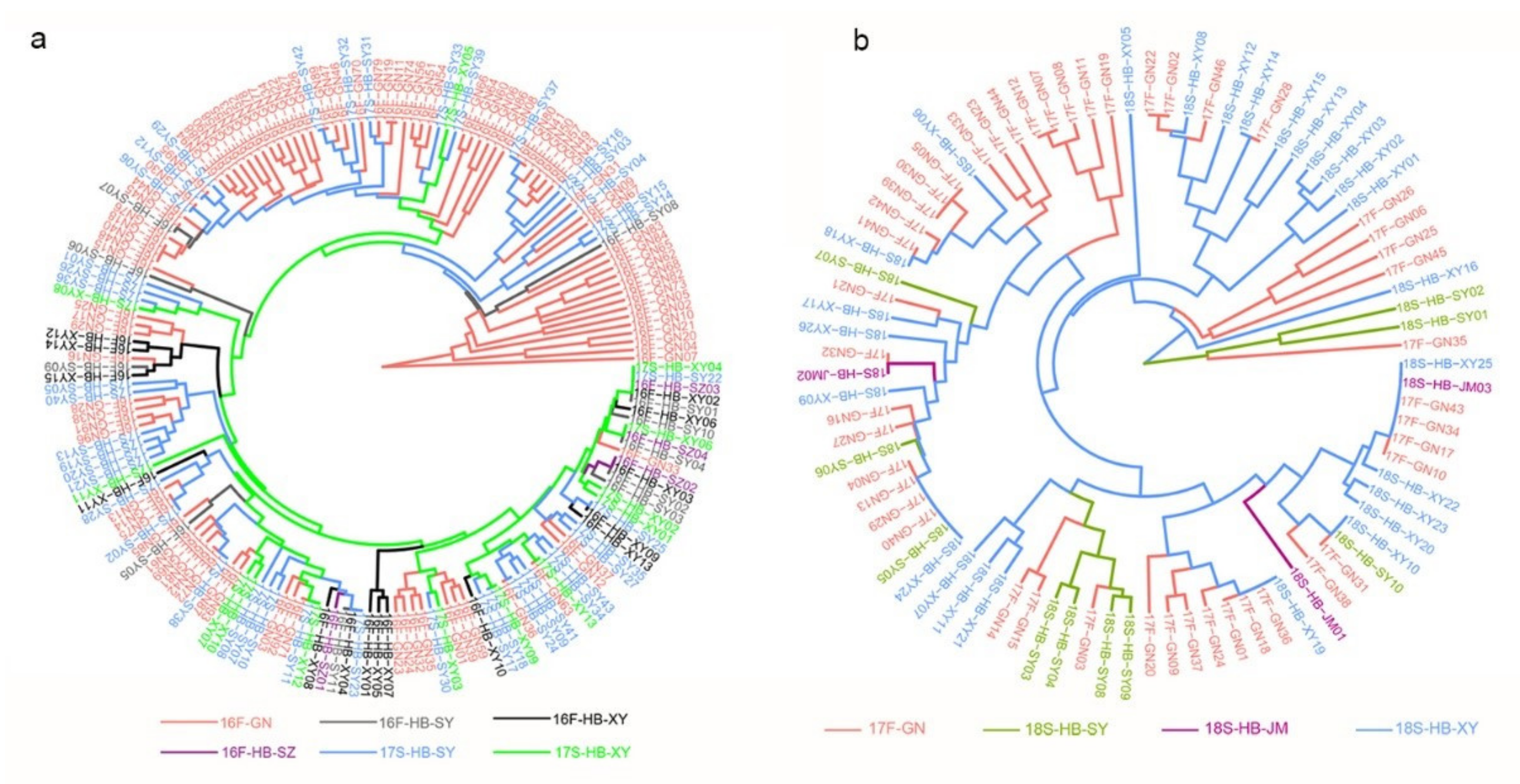{kind=link}
| Subpopulation a | Sizesub b | Sampling Time (Date/Month) | City | County | Sizecou c | Elevation (m) | Cultivar |
|---|---|---|---|---|---|---|---|
| 16F-GN | 149 | 23/11 | Tianshui | Gangu | 14 | 1810 | Unknown |
| 22/11 | Qingshui | 1 | 1560 | Unknown | |||
| 23/11 | Qinzhou | 3 | 1710 | Unknown | |||
| 24/11 | Dingxi | Longxi | 4 | 2090–2120 | Unknown | ||
| 23/11 | Lintao | 20 | Unknown | Unknown | |||
| 22/11 | Pingliang | Zhuanglang | 11 | 1640 | Unknown | ||
| 23/11 | Guyuan | Longde | 37 | 1479–2082 | Zhuang 88, Zhongyin 16, Laoxuan 1, | ||
| 21/11 | Jingyuan | 20 | 1704–1919 | Zhongyin 3, Longjian 127, Qinmai 4, Ningdong 3, Ningdong 5 | |||
| 23/11 | Pengyang | 14 | 1479–1633 | Unknown | |||
| 23/11 | Yuanzhou | 25 | 1595–1832 | Ningdong 1, Longzhong 6 | |||
| 16F-HB-SY | 45 | 22/12 | Shiyan | Yunyang | 15 | 190 | Unknown |
| 23/12 | Yunxi | 30 | 170–400 | Mianyang 26 | |||
| 16F-HB-XY | 47 | 21/1 | Xiangyang | Laohekou | 20 | Unknown | Unknown |
| 19/1 | Xiangcheng | 20 | Unknown | Unknown | |||
| 17/1 | Zaoyang | 7 | 94 | Unknown | |||
| 16F-HB-SZ | 8 | 17/1 | Suizhou | Suizhou | 8 | Unknown | Unknown |
| 17S-HB-SY | 72 | 29/4 | Shiyan | Yunyang | 44 | 205-680 | Echun series |
| 1/5 | Yunxi | 28 | Unknown | Mianyang 31 | |||
| 17S-HB-XY | 24 | 20/4 | Xiangyang | Fancheng | 24 | Unknown | Unknown |
| 17F-GN | 113 | 23/11 | Tianshui | Qinzhou | 4 | 1630–1690 | Zhongliang |
| 23/11 | Maiji | 15 | 1700 | Zhongliang | |||
| 24/11 | Dingxi | Longxi | 13 | 2190 | Unknown | ||
| 24/11 | Tongwei | 30 | 1850–1990 | Lantian | |||
| 23/11 | Pingliang | Kongtong | 9 | 1490 | Pingliang 42–44 | ||
| 22/11 | Zhuanglang | 19 | 1650–1740 | 1028, 1038, Lantian 26 | |||
| 16/11 | Wuzhong | Tongxin | 23 | Unknown | Unknown | ||
| 18S-HB-SY | 30 | 23/3 | Shiyan | Yunyang | 30 | 167–374 | Xiangmai75, Emai596, Zhengmai005 |
| 18S-HB-JM | 9 | 19/3 | Jingmen | Zhongxiang | 9 | 40 | Huayu911, |
| 18S-HB-XY | 70 | 2/4 | Xiangyang | Nanzhang | 14 | Unknown | Unknown |
| 29/3 | Yicheng | 11 | Unknown | Xiangmai55, Zhengmai9023 | |||
| 1/4 | Laohekou | 24 | Unknown | Unknown | |||
| 1/4 | Fancheng | 11 | Unknown | Unknown | |||
| 1/4 | Xiangcheng | 10 | 70 | Zhengmai101, Zhouheimai1 | |||
| Total | 567 |
| Locus | Repeat Motif | Primer Sequence (5′–3′) | Ta (°C) | No. of Alleles (Size) | Reference |
|---|---|---|---|---|---|
| CPS8 | (CAG)14 | F: FAM-GATAAGAAACAAGGGACAGC R: CAGTGAACCCAATTACTCAG | 55 | 5 (200–212) | [17] |
| CPS13 | (GAC)6 | F: FAM-TCCAGGCAGTAAATCAGACGC R: ATCAGCAGGTGTAGCCCCATC | 58 | 2 (125–128) | [17] |
| CPS27 | (TTC)4 | F: TAMRA-GATGGGGAAAAGTAAGAAGT R: GGTGGGGGATGTAAGTATGTA | 57 | 2 (225–228) | [17] |
| CPS34 | (TC)9 | F: TAMRA-GTTGGCTACGAGTGGTCATC R: TAACACTACAAAAGGGGTC | 55 | 5 (104–114) | [17] |
| RJO3 | (TGG)8 | F: FAM- GCAGCACTGGCAGGTGG R: GATGAATCAGGATGGCTCC | 52 | 4 (201–212) | [18] |
| RJO20 | (CAG)4 | F: HEX-AGAAGATCGACGCACCCG R: CCTCCGATTGGCTTAGGC | 52 | 3 (283–289) | [18] |
| RJ3N | (CT)9 | F: ROX-TGGTGGTGCTCCTCTAGTC R: AGGGGTCTTGTAAGATGCTC | 52 | 4 (335–343) | [19] |
| RJ5N | (CT)8 | F: ROX-AACGGTCAACAGCACTCAC R: AGTTGGTCGCGTTTTGCTC | 52 | 3 (223–229) | [19] |
| RJ6N | (AAC)9 | F: TAMRA-CAATCTGGCGGACAGCAAC R: CACCTAGGATACCACCGCC | 52 | 4 (309–318) | [19] |
| RJ8N | (GAT)8 | F: FAM-ACTGGGCAGACTGGTCAAC R: TCGTTTCCCTCCAGATGGC | 52 | 6 (301–330) | [19] |
| RJ13N | (ACG)6 | F: HEX-TTAGCTCAGCCGGTTCCTC R: CAGGTGTAGCCCCATCTCC | 52 | 2 (149–152) | [19] |
| WSR44 | (GT)6 | F: HEX-AGGCCCCAGGAACACAAAAA R: TCACACACGCTCCACAGTAC | 56 | 2 (188–190) | [20] |
| Subpopulation a | n | MLG b | eMLG c | Lambda | Corrected Lambda d | No. of Clones | e Standardized Index of Association (rBarD) | Nei’s Unbiased Gene Diversity |
|---|---|---|---|---|---|---|---|---|
| 16F-GN | 149 | 90 | 9.01 | 0.97 | 0.97 | 96 | 0.0065 | 0.27 |
| 17F-GN | 113 | 34 | 6.59 | 0.86 | 0.87 | 46 | 0.0736 ** | 0.29 |
| 16F-HB-SY | 45 | 11 | 4.44 | 0.68 | 0.69 | 11 | −0.034 | 0.16 |
| 17S-HB-SY | 72 | 43 | 9.00 | 0.96 | 0.98 | 43 | 0.055 | 0.25 |
| 18S-HB-SY | 30 | 10 | 6.03 | 0.84 | 0.87 | 10 | 0.1765 ** | 0.30 |
| 16F-HB-XY | 47 | 15 | 4.63 | 0.61 | 0.62 | 15 | 0.040 | 0.17 |
| 17S-HB-XY | 24 | 13 | 7.15 | 0.86 | 0.90 | 13 | 0.024 | 0.19 |
| 18S-HB-XY | 70 | 26 | 7.47 | 0.91 | 0.93 | 26 | 0.0579 ** | 0.31 |
| 16F-HB-SZ | 8 | 4 | 4 | 0.66 | 0.75 | 4 | −0.115 | 0.18 |
| 18S-HB-JM | 9 | 3 | 3 | 0.57 | 0.64 | 3 | NA | 0.26 |
| Total | 567 | 208 | 8.79 | 0.96 | 0.97 | 267 | 0.2320 ** | 0.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Jiang, B.; Liang, J.; Li, L.; Gu, Y.; Li, J.; Luo, Y.; Ma, Z. Population Genetic Structures of Puccinia striiformis f. sp. tritici in the Gansu-Ningxia Region and Hubei Province, China. Genes 2021, 12, 1712. https://doi.org/10.3390/genes12111712
Wang C, Jiang B, Liang J, Li L, Gu Y, Li J, Luo Y, Ma Z. Population Genetic Structures of Puccinia striiformis f. sp. tritici in the Gansu-Ningxia Region and Hubei Province, China. Genes. 2021; 12(11):1712. https://doi.org/10.3390/genes12111712
Chicago/Turabian StyleWang, Cuicui, Bingbing Jiang, Junmin Liang, Leifu Li, Yilin Gu, Jintang Li, Yong Luo, and Zhanhong Ma. 2021. "Population Genetic Structures of Puccinia striiformis f. sp. tritici in the Gansu-Ningxia Region and Hubei Province, China" Genes 12, no. 11: 1712. https://doi.org/10.3390/genes12111712
APA StyleWang, C., Jiang, B., Liang, J., Li, L., Gu, Y., Li, J., Luo, Y., & Ma, Z. (2021). Population Genetic Structures of Puccinia striiformis f. sp. tritici in the Gansu-Ningxia Region and Hubei Province, China. Genes, 12(11), 1712. https://doi.org/10.3390/genes12111712