
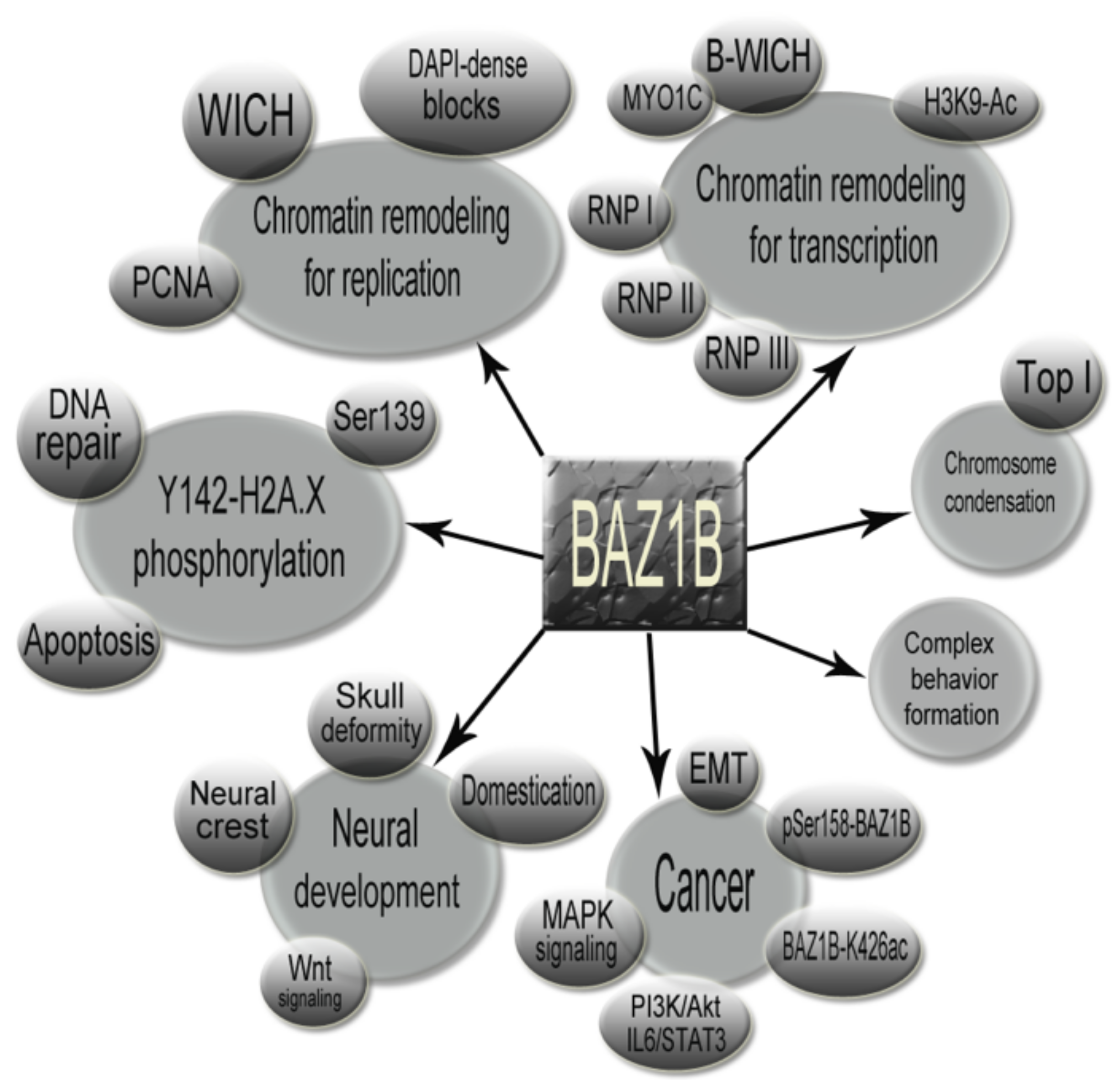
BAZ1B the Protean Protein


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Bromodomain

1.2. Plant Homeodomain Finger
1.3. WAKZ and WAC Motifs
1.4. WSTF or BAZ1B?
1.5. BAZ or WHIM Motifs?
1.6. DDT and LXXLL Motifs
1.7. Other Motifs and Conserved Regions
2. BAZ1B and Chromatin Remodeling
2.1. WICH: A Chromatin Remodeling Complex for Replication
2.2. B-WICH: A Transcription Regulator Complex
3. BAZ1B and DNA Repair
3.1. BAZ1B Atypical Kinase
3.2. Phosphorylated Tyr142
3.3. BAZ1B and Apoptosis
4. BAZ1B and Neural Development
5. BAZ1B and Other Outstanding Roles
5.1. BAZ1B and Chromosomes
5.2. BAZ1B and Cancer
5.3. BAZ1B and Behavior
5.4. BAZ1B and Noise-Induced Damage
6. Conclusions and Further Insights
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, X.; Meng, X.; Morris, C.A.; Keating, M.T. A Novel Human Gene, WSTF, Is Deleted in Williams Syndrome. Genomics 1998, 54, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Peoples, R.J.; Cisco, M.J.; Kaplan, P.; Francke, U. Identification of the WBSCR9 Gene, Encoding a Novel Transcriptional Regulator, in the Williams-Beuren Syndrome Deletion at 7q11.23. Cytogenet. Genome Res. 1998, 82, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.A.; Demsey, S.A.; Leonard, C.O.; Dilts, C.; Blackburn, B.L. Natural History of Williams Syndrome: Physical Characteristics. J. Pediatr. 1988, 113, 318–326. [Google Scholar] [CrossRef]
- Bellugi, U.; Bihrle, A.; Jernigan, T.; Trauner, D.; Doherty, S. Neuropsychological, Neurological, and Neuroanatomical Profile of Williams Syndrome. Am. J. Med. Genet. 2005, 37, 115–125. [Google Scholar] [CrossRef]
- Adams, G.N.; Schmaier, A.H. The Williams-Beuren Syndrome—A Window into Genetic Variants Leading to the Development of Cardiovascular Disease. PLoS Genet. 2012, 8, e1002479. [Google Scholar] [CrossRef]
- Ewart, A.K.; Morris, C.A.; Atkinson, D.; Jin, W.; Sternes, K.; Spallone, P.; Stock, A.D.; Leppert, M.; Keating, M.T. Hemizygosity at the Elastin Locus in a Developmental Disorder, Williams Syndrome. Nat. Genet. 1993, 5, 11–16. [Google Scholar] [CrossRef]
- Earhart, B.A.; Williams, M.E.; Zamora, I.; Randolph, L.M.; Votava-Smith, J.K.; Marcy, S.N. Phenotype of 7q11.23 Duplication: A Family Clinical Series. Am. J. Med. Genet. A 2017, 173, 114–119. [Google Scholar] [CrossRef]
- Samaras, P.; Schmidt, T.; Frejno, M.; Gessulat, S.; Reinecke, M.; Jarzab, A.; Zecha, J.; Mergner, J.; Giansanti, P.; Ehrlich, H.-C.; et al. ProteomicsDB: A Multi-Omics and Multi-Organism Resource for Life Science Research. Nucleic Acids Res. 2019, 48, D1153–D1163. [Google Scholar] [CrossRef]
- ProteomicsDB. Available online: https://www.proteomicsdb.org (accessed on 1 August 2021).
- STRING. Available online: https://string-db.org (accessed on 1 August 2021).
- Dhalluin, C.; Carlson, J.E.; Zeng, L.; He, C.; Aggarwal, A.K.; Zhou, M.-M.; Zhou, M.-M. Structure and Ligand of a Histone Acetyltransferase Bromodomain. Nature 1999, 399, 491–496. [Google Scholar] [CrossRef]
- Haynes, S.R.; Dollard, C.; Winston, F.; Beck, S.; Trowsdale, J.; Dawid, I.B. The Bromodomain: A Conserved Sequence Found in Human, Drosophila and Yeast Proteins. Nucleic Acids Res. 1992, 20, 2603. [Google Scholar] [CrossRef]
- Jones, M.H.; Hamana, N.; Nezu, J.; Shimane, M. A Novel Family of Bromodomain Genes. Genomics 2000, 63, 40–45. [Google Scholar] [CrossRef]
- Owen, D.J. The Structural Basis for the Recognition of Acetylated Histone H4 by the Bromodomain of Histone Acetyltransferase Gcn5p. EMBO J. 2000, 19, 6141–6149. [Google Scholar] [CrossRef]
- Zaware, N.; Zhou, M.-M. Bromodomain Biology and Drug Discovery. Nat. Struct. Mol. Biol. 2019, 26, 870–879. [Google Scholar] [CrossRef]
- Filippakopoulos, P.; Qi, J.; Picaud, S.; Shen, Y.; Smith, W.B.; Fedorov, O.; Morse, E.M.; Keates, T.; Hickman, T.T.; Felletar, I.; et al. Selective Inhibition of BET Bromodomains. Nature 2010, 468, 1067–1073. [Google Scholar] [CrossRef]
- Nicodeme, E.; Jeffrey, K.L.; Schaefer, U.; Beinke, S.; Dewell, S.; Chung, C.; Chandwani, R.; Marazzi, I.; Wilson, P.; Coste, H.; et al. Suppression of Inflammation by a Synthetic Histone Mimic. Nature 2010, 468, 1119–1123. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Levenstein, M.E.; Fyodorov, D.V.; Kutach, A.K.; Kobayashi, R.; Kadonaga, J.T. ACF Consists of Two Subunits, Acf1 and ISWI, That Function Cooperatively in the ATP-Dependent Catalysis of Chromatin Assembly. Genes Dev. 1999, 13, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Fyodorov, D.V.; Kadonaga, J.T. Binding of Acf1 to DNA Involves a WAC Motif and Is Important for ACF-Mediated Chromatin Assembly. Mol. Cell. Biol. 2002, 22, 6344–6353. [Google Scholar] [CrossRef] [PubMed]
- Doerks, T.; Copley, R.; Bork, P. DDT—A Novel Domain in Different Transcription and Chromosome Remodeling Factors. Trends Biochem. Sci. 2001, 26, 145–146. [Google Scholar] [CrossRef]
- Aravind, L.; Iyer, L.M. The HARE-HTH and Associated Domains: Novel Modules in the Coordination of Epigenetic DNA and Protein Modifications. Cell Cycle 2012, 11, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Frouws, T.D.; Angst, B.; Fitzgerald, D.J.; DeLuca, C.; Schimmele, K.; Sargent, D.F.; Richmond, T.J. Structure and Mechanism of the Chromatin Remodelling Factor ISW1a. Nature 2011, 472, 448–453. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Aasland, R. The PHD Finger: Implications for Chromatin-Mediated Transcriptional Regulation. Trends Biochem. Sci. 1995, 20, 56–59. [Google Scholar] [CrossRef]
- Sanchez, R.; Zhou, M.-M. The PHD Finger: A Versatile Epigenome Reader. Trends Biochem. Sci. 2011, 36, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Sehnal, D.; Rose, A.; Koca, J.; Burley, S.; Velankar, S. Mol*: Towards a common library and tools for web molecular graphics. In Workshop on Molecular Graphics and Visual Analysis of Molecular Data 2018; EuroVis: Brno, Czech Republic, 2018; p. 5. [Google Scholar] [CrossRef]
- Xiao, A.; Li, H.; Shechter, D.; Ahn, S.H.; Fabrizio, L.A.; Erdjument-Bromage, H.; Ishibe-Murakami, S.; Wang, B.; Tempst, P.; Hofmann, K.; et al. WSTF Regulates the H2A.X DNA Damage Response via a Novel Tyrosine Kinase Activity. Nature 2009, 457, 57–62. [Google Scholar] [CrossRef]
- Poot, R.A. HuCHRAC, a Human ISWI Chromatin Remodelling Complex Contains HACF1 and Two Novel Histone-Fold Proteins. EMBO J. 2000, 19, 3377–3387. [Google Scholar] [CrossRef]
- Heery, D.M.; Kalkhoven, E.; Hoare, S.; Parker, M.G. A Signature Motif in Transcriptional Co-Activators Mediates Binding to Nuclear Receptors. Nature 1997, 387, 733–736. [Google Scholar] [CrossRef]
- Le Douarin, B.; Nielsen, A.L.; Garnier, J.M.; Ichinose, H.; Jeanmougin, F.; Losson, R.; Chambon, P. A Possible Involvement of TIF1 α and TIF1 β in the Epigenetic Control of Transcription by Nuclear Receptors. EMBO J. 1996, 15, 6701–6715. [Google Scholar] [CrossRef]
- Bozhenok, L. WSTF-ISWI Chromatin Remodeling Complex Targets Heterochromatic Replication Foci. EMBO J. 2002, 21, 2231–2241. [Google Scholar] [CrossRef] [PubMed]
- Chishti, A.H.; Kim, A.C.; Marfatia, S.M.; Lutchman, M.; Hanspal, M.; Jindal, H.; Liu, S.-C.; Low, P.S.; Rouleau, G.A.; Mohandas, N.; et al. The FERM Domain: A Unique Module Involved in the Linkage of Cytoplasmic Proteins to the Membrane. Trends Biochem. Sci. 1998, 23, 281–282. [Google Scholar] [CrossRef]
- Conboy, J.; Kan, Y.W.; Shohet, S.B.; Mohandas, N. Molecular Cloning of Protein 4.1, a Major Structural Element of the Human Erythrocyte Membrane Skeleton. Proc. Natl. Acad. Sci. USA 1986, 83, 9512–9516. [Google Scholar] [CrossRef]
- Rogers, S.; Wells, R.; Rechsteiner, M. Amino Acid Sequences Common to Rapidly Degraded Proteins: The PEST Hypothesis. Science 1986, 234, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Rechsteiner, M.; Rogers, S.W. PEST Sequences and Regulation by Proteolysis. Trends Biochem. Sci. 1996, 21, 267–271. [Google Scholar] [CrossRef]
- Li, Y.; Jin, K.; Bunker, E.; Zhang, X.; Luo, X.; Liu, X.; Hao, B. Structural Basis of the Phosphorylation-Independent Recognition of Cyclin D1 by the SCF FBXO31 Ubiquitin Ligase. Proc. Natl. Acad. Sci. USA 2018, 115, 319–324. [Google Scholar] [CrossRef]
- Qile, M.; Ji, Y.; Houtman, M.J.C.; Veldhuis, M.; Romunde, F.; Kok, B.; van der Heyden, M.A.G. Identification of a PEST Sequence in Vertebrate KIR2.1 That Modifies Rectification. Front. Physiol. 2019, 10, 863. [Google Scholar] [CrossRef]
- Aydin, Ö.Z.; Vermeulen, W.; Lans, H. ISWI Chromatin Remodeling Complexes in the DNA Damage Response. Cell Cycle 2014, 13, 3016–3025. [Google Scholar] [CrossRef]
- Elfring, L.K.; Deuring, R.; McCallum, C.M.; Peterson, C.L.; Tamkun, J.W. Identification and Characterization of Drosophila Relatives of the Yeast Transcriptional Activator SNF2/SWI2. Mol. Cell. Biol. 1994, 14, 2225–2234. [Google Scholar] [CrossRef]
- Tyagi, M.; Imam, N.; Verma, K.; Patel, A.K. Chromatin Remodelers: We Are the Drivers!! Nucleus 2016, 7, 388–404. [Google Scholar] [CrossRef]
- Taddei, A.; Maison, C.; Roche, D.; Almouzni, G. Reversible Disruption of Pericentric Heterochromatin and Centromere Function by Inhibiting Deacetylases. Nat. Cell Biol. 2001, 3, 114–120. [Google Scholar] [CrossRef]
- Poot, R.A.; Bozhenok, L.; van den Berg, D.L.C.; Steffensen, S.; Ferreira, F.; Grimaldi, M.; Gilbert, N.; Ferreira, J.; Varga-Weisz, P.D. The Williams Syndrome Transcription Factor Interacts with PCNA to Target Chromatin Remodelling by ISWI to Replication Foci. Nat. Cell Biol. 2004, 6, 1236–1244. [Google Scholar] [CrossRef]
- Istomina, N.E.; Shushanov, S.S.; Springhetti, E.M.; Karpov, V.L.; Krasheninnikov, I.A.; Stevens, K.; Zaret, K.S.; Singh, P.B.; Grigoryev, S.A. Insulation of the Chicken β-Globin Chromosomal Domain from a Chromatin-Condensing Protein, MENT. Mol. Cell. Biol. 2003, 23, 6455–6468. [Google Scholar] [CrossRef] [PubMed]
- Culver-Cochran, A.E.; Chadwick, B.P. Loss of WSTF Results in Spontaneous Fluctuations of Heterochromatin Formation and Resolution, Combined with Substantial Changes to Gene Expression. BMC Genom. 2013, 14, 740. [Google Scholar] [CrossRef]
- Barr, M.L.; Bertram, E.G. A Morphological Distinction between Neurones of the Male and Female, and the Behaviour of the Nucleolar Satellite during Accelerated Nucleoprotein Synthesis. Nature 1949, 163, 676–677. [Google Scholar] [CrossRef]
- Culver-Cochran, A.E.; Chadwick, B.P. The WSTF-ISWI Chromatin Remodeling Complex Transiently Associates with the Human Inactive X Chromosome during Late S-Phase Prior to BRCA1 and γ-H2AX. PLoS ONE 2012, 7, e50023. [Google Scholar] [CrossRef]
- Willard, H.F.; Latt, S.A. Analysis of Deoxyribonucleic Acid Replication in Human X Chromosomes by Fluorescence Microscopy. Am. J. Hum. Genet. 1976, 28, 213–227. [Google Scholar]
- Tsurimoto, T. PCNA Binding Proteins. Front. Biosci. J. Virtual Libr. 1999, 4, D849–D858. [Google Scholar] [CrossRef]
- Poot, R.A.; Bozhenok, L.; van den Berg, D.L.C.; Hawkes, N.; Varga-Weisz, P.D. Chromatin Remodeling by WSTF-ISWI at the Replication Site: Opening a Window of Opportunity for Epigenetic Inheritance? Cell Cycle 2005, 4, 543–546. [Google Scholar] [CrossRef]
- Sugimoto, N.; Kitabayashi, I.; Osano, S.; Tatsumi, Y.; Yugawa, T.; Narisawa-Saito, M.; Matsukage, A.; Kiyono, T.; Fujita, M. Identification of Novel Human Cdt1-Binding Proteins by a Proteomics Approach: Proteolytic Regulation by APC/CCdh1. Mol. Biol. Cell 2008, 19, 1007–1021. [Google Scholar] [CrossRef]
- Arias, E.E.; Walter, J.C. PCNA Functions as a Molecular Platform to Trigger Cdt1 Destruction and Prevent Re-Replication. Nat. Cell Biol. 2006, 8, 84–90. [Google Scholar] [CrossRef]
- Sugimoto, N.; Yugawa, T.; Iizuka, M.; Kiyono, T.; Fujita, M. Chromatin Remodeler Sucrose Nonfermenting 2 Homolog (SNF2H) Is Recruited onto DNA Replication Origins through Interaction with Cdc10 Protein-Dependent Transcript 1 (Cdt1) and Promotes Pre-Replication Complex Formation. J. Biol. Chem. 2011, 286, 39200–39210. [Google Scholar] [CrossRef] [PubMed]
- Alexiadis, V.; Varga-Weisz, P.D.; Bonte, E.; Becker, P.B.; Gruss, C. In Vitro Chromatin Remodelling by Chromatin Accessibility Complex (CHRAC) at the SV40 Origin of DNA Replication. EMBO J. 1998, 17, 3428–3438. [Google Scholar] [CrossRef] [PubMed]
- Cavellán, E.; Asp, P.; Percipalle, P.; Farrants, A.-K.Ö. The WSTF-SNF2h Chromatin Remodeling Complex Interacts with Several Nuclear Proteins in Transcription. J. Biol. Chem. 2006, 281, 16264–16271. [Google Scholar] [CrossRef]
- Percipalle, P.; Fomproix, N.; Cavellán, E.; Voit, R.; Reimer, G.; Krüger, T.; Thyberg, J.; Scheer, U.; Grummt, I.; Östlund Farrants, A. The Chromatin Remodelling Complex WSTF–SNF2h Interacts with Nuclear Myosin 1 and Has a Role in RNA Polymerase I Transcription. EMBO Rep. 2006, 7, 525–530. [Google Scholar] [CrossRef]
- Bradsher, J.; Auriol, J.; Proietti de Santis, L.; Iben, S.; Vonesch, J.L.; Grummt, I.; Egly, J.M. CSB Is a Component of RNA Pol I Transcription. Mol. Cell 2002, 10, 819–829. [Google Scholar] [CrossRef]
- Selby, C.P.; Sancar, A. Cockayne Syndrome Group B Protein Enhances Elongation by RNA Polymerase II. Proc. Natl. Acad. Sci. USA 1997, 94, 11205–11209. [Google Scholar] [CrossRef]
- Ciaffardini, F.; Nicolai, S.; Caputo, M.; Canu, G.; Paccosi, E.; Costantino, M.; Frontini, M.; Balajee, A.S.; Proietti-De-Santis, L. The Cockayne Syndrome B Protein Is Essential for Neuronal Differentiation and Neuritogenesis. Cell Death Dis. 2014, 5, e1268. [Google Scholar] [CrossRef] [PubMed]
- Henning, D.; So, R.B.; Jin, R.; Lau, L.F.; Valdez, B.C. Silencing of RNA Helicase II/Guα Inhibits Mammalian Ribosomal RNA Production. J. Biol. Chem. 2003, 278, 52307–52314. [Google Scholar] [CrossRef] [PubMed]
- Calo, E.; Flynn, R.A.; Martin, L.; Spitale, R.C.; Chang, H.Y.; Wysocka, J. RNA Helicase DDX21 Coordinates Transcription and Ribosomal RNA Processing. Nature 2015, 518, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Cretu, C.; Schmitzová, J.; Ponce-Salvatierra, A.; Dybkov, O.; De Laurentiis, E.I.; Sharma, K.; Will, C.L.; Urlaub, H.; Lührmann, R.; Pena, V. Molecular Architecture of SF3b and Structural Consequences of Its Cancer-Related Mutations. Mol. Cell 2016, 64, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Tavner, F.J.; Simpson, R.; Tashiro, S.; Favier, D.; Jenkins, N.A.; Gilbert, D.J.; Copeland, N.G.; Macmillan, E.M.; Lutwyche, J.; Keough, R.A.; et al. Molecular Cloning Reveals That the P160 Myb-Binding Protein Is a Novel, Predominantly Nucleolar Protein Which May Play a Role in Transactivation by Myb. Mol. Cell. Biol. 1998, 18, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Keough, R.A.; Gonda, T.J.; Ishii, S. Ribosomal Stress Induces Processing of Mybbp1a and Its Translocation from the Nucleolus to the Nucleoplasm: Processing of Mybbp1a. Genes Cells 2007, 13, 27–39. [Google Scholar] [CrossRef]
- Waldmann, T.; Scholten, I.; Kappes, F.; Hu, H.G.; Knippers, R. The DEK Protein—An Abundant and Ubiquitous Constituent of Mammalian Chromatin. Gene 2004, 343, 1–9. [Google Scholar] [CrossRef]
- Von Lindern, M.; Fornerod, M.; Soekarman, N.; van Baal, S.; Jaegle, M.; Hagemeijer, A.; Bootsma, D.; Grosveld, G. Translocation t(6;9) in Acute Non-Lymphocytic Leukaemia Results in the Formation of a DEK-CAN Fusion Gene. Baillieres Clin. Haematol. 1992, 5, 857–879. [Google Scholar] [CrossRef]
- Wise-Draper, T.M.; Allen, H.V.; Thobe, M.N.; Jones, E.E.; Habash, K.B.; Münger, K.; Wells, S.I. The Human DEK Proto-Oncogene Is a Senescence Inhibitor and an Upregulated Target of High-Risk Human Papillomavirus E7. J. Virol. 2005, 79, 14309–14317. [Google Scholar] [CrossRef][Green Version]
- Pestic-Dragovich, L.; Stojiljkovic, L.; Philimonenko, A.A.; Nowak, G.; Ke, Y.; Settlage, R.E.; Shabanowitz, J.; Hunt, D.F.; Hozak, P.; de Lanerolle, P. A Myosin I Isoform in the Nucleus. Science 2000, 290, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Venit, T.; Kalendová, A.; Petr, M.; Dzijak, R.; Pastorek, L.; Rohožková, J.; Malohlava, J.; Hozák, P. Nuclear Myosin I Regulates Cell Membrane Tension. Sci. Rep. 2016, 6, 30864. [Google Scholar] [CrossRef] [PubMed]
- Almuzzaini, B.; Sarshad, A.A.; Farrants, A.-K.Ö.; Percipalle, P. Nuclear Myosin 1 Contributes to a Chromatin Landscape Compatible with RNA Polymerase II Transcription Activation. BMC Biol. 2015, 13, 35. [Google Scholar] [CrossRef]
- Sarshad, A.; Sadeghifar, F.; Louvet, E.; Mori, R.; Böhm, S.; Al-Muzzaini, B.; Vintermist, A.; Fomproix, N.; Östlund, A.-K.; Percipalle, P. Nuclear Myosin 1c Facilitates the Chromatin Modifications Required to Activate rRNA Gene Transcription and Cell Cycle Progression. PLoS Genet. 2013, 9, e1003397. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Chittiboyina, S.; Bai, Y.; Abad, P.C.; Vidi, P.-A.; Stauffacher, C.V.; Lelièvre, S.A. The Nuclear Mitotic Apparatus Protein NuMA Controls RDNA Transcription and Mediates the Nucleolar Stress Response in a P53-Independent Manner. Nucleic Acids Res. 2017, 45, 11725–11742. [Google Scholar] [CrossRef] [PubMed]
- Vintermist, A.; Böhm, S.; Sadeghifar, F.; Louvet, E.; Mansén, A.; Percipalle, P.; Östlund Farrants, A.-K. The Chromatin Remodelling Complex B-WICH Changes the Chromatin Structure and Recruits Histone Acetyl-Transferases to Active rRNA Genes. PLoS ONE 2011, 6, e19184. [Google Scholar] [CrossRef]
- Sadeghifar, F.; Böhm, S.; Vintermist, A.; Östlund Farrants, A.-K. The B-WICH Chromatin-Remodelling Complex Regulates RNA Polymerase III Transcription by Promoting Max-Dependent c-Myc Binding. Nucleic Acids Res. 2015, 43, 4477–4490. [Google Scholar] [CrossRef]
- Lundqvist, J.; Wikvall, K.; Norlin, M. Vitamin D-Mediated Regulation of CYP21A2 Transcriptio—A Novel Mechanism for Vitamin D Action. Biochim. Biophys. Acta BBA Gen. Subj. 2012, 1820, 1553–1559. [Google Scholar] [CrossRef] [PubMed]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA Double-Stranded Breaks Induce Histone H2AX Phosphorylation on Serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [PubMed]
- Bonner, W.M.; Redon, C.E.; Dickey, J.S.; Nakamura, A.J.; Sedelnikova, O.A.; Solier, S.; Pommier, Y. ΓH2AX and Cancer. Nat. Rev. Cancer 2008, 8, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Burma, S.; Chen, B.P.; Murphy, M.; Kurimasa, A.; Chen, D.J. ATM Phosphorylates Histone H2AX in Response to DNA Double-Strand Breaks. J. Biol. Chem. 2001, 276, 42462–42467. [Google Scholar] [CrossRef] [PubMed]
- Redon, C.; Pilch, D.; Rogakou, E.; Sedelnikova, O.; Newrock, K.; Bonner, W. Histone H2A Variants H2AX and H2AZ. Curr. Opin. Genet. Dev. 2002, 12, 162–169. [Google Scholar] [CrossRef]
- Cook, P.J.; Ju, B.G.; Telese, F.; Wang, X.; Glass, C.K.; Rosenfeld, M.G. Tyrosine Dephosphorylation of H2AX Modulates Apoptosis and Survival Decisions. Nature 2009, 458, 591–596. [Google Scholar] [CrossRef]
- Soldi, M.; Bonaldi, T. The Proteomic Investigation of Chromatin Functional Domains Reveals Novel Synergisms among Distinct Heterochromatin Components. Mol. Cell. Proteomics 2013, 12, 764–780. [Google Scholar] [CrossRef] [PubMed]
- Stucki, M.; Clapperton, J.A.; Mohammad, D.; Yaffe, M.B.; Smerdon, S.J.; Jackson, S.P. MDC1 Directly Binds Phosphorylated Histone H2AX to Regulate Cellular Responses to DNA Double-Strand Breaks. Cell 2005, 123, 1213–1226. [Google Scholar] [CrossRef]
- Singh, N.; Basnet, H.; Wiltshire, T.D.; Mohammad, D.H.; Thompson, J.R.; Héroux, A.; Botuyan, M.V.; Yaffe, M.B.; Couch, F.J.; Rosenfeld, M.G.; et al. Dual Recognition of Phosphoserine and Phosphotyrosine in Histone Variant H2A.X by DNA Damage Response Protein MCPH1. Proc. Natl. Acad. Sci. USA 2012, 109, 14381–14386. [Google Scholar] [CrossRef]
- Rebay, I. Multiple Functions of the Eya Phosphotyrosine Phosphatase. Mol. Cell. Biol. 2016, 36, 668–677. [Google Scholar] [CrossRef]
- Jeffery, N.N.; Davidson, C.; Peslak, S.A.; Kingsley, P.D.; Nakamura, Y.; Palis, J.; Bulger, M. Histone H2A.X Phosphorylation and Caspase-Initiated Chromatin Condensation in Late-Stage Erythropoiesis. Epigenet. Chromatin 2021, 14, 37. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, C.; Cao, Y.; Luo, Y.; Bao, R.; Yan, S.; Xue, M.; Zhu, F.; Wang, Z.; Duan, L. Imatinib Induces H2AX Phosphorylation and Apoptosis in Chronic Myelogenous Leukemia Cells in Vitro via Caspase-3/Mst1 Pathway. Acta Pharmacol. Sin. 2012, 33, 551–557. [Google Scholar] [CrossRef]
- Brestovitsky, A.; Sharf, R.; Mittelman, K.; Kleinberger, T. The Adenovirus E4orf4 Protein Targets PP2A to the ACF Chromatin-Remodeling Factor and Induces Cell Death through Regulation of SNF2h-Containing Complexes. Nucleic Acids Res. 2011, 39, 6414–6427. [Google Scholar] [CrossRef] [PubMed]
- Barnett, C.; Yazgan, O.; Kuo, H.-C.; Malakar, S.; Thomas, T.; Fitzgerald, A.; Harbour, W.; Henry, J.J.; Krebs, J.E. Williams Syndrome Transcription Factor Is Critical for Neural Crest Cell Function in Xenopus Laevis. Mech. Dev. 2012, 129, 324–338. [Google Scholar] [CrossRef] [PubMed]
- Knöchel, S.; Dillinger, K.; Köster, M.; Knöchel, W. Structure and Expression of Xenopus Tropicalis BMP-2 and BMP-4 Genes. Mech. Dev. 2001, 109, 79–82. [Google Scholar] [CrossRef]
- Van Acker, H.H.; Capsomidis, A.; Smits, E.L.; Van Tendeloo, V.F. CD56 in the Immune System: More Than a Marker for Cytotoxicity? Front. Immunol. 2017, 8, 892. [Google Scholar] [CrossRef] [PubMed]
- Marigo, V.; Roberts, D.J.; Lee, S.M.; Tsukurov, O.; Levi, T.; Gastier, J.M.; Epstein, D.J.; Gilbert, D.J.; Copeland, N.G.; Seidman, C.E. Cloning, Expression, and Chromosomal Location of SHH and IHH: Two Human Homologues of the Drosophila Segment Polarity Gene Hedgehog. Genomics 1995, 28, 44–51. [Google Scholar] [CrossRef]
- Moncaut, N.; Rigby, P.W.J.; Carvajal, J.J. Dial M(RF) for Myogenesis. FEBS J. 2013, 280, 3980–3990. [Google Scholar] [CrossRef]
- Barua, M.; Stellacci, E.; Stella, L.; Weins, A.; Genovese, G.; Muto, V.; Caputo, V.; Toka, H.R.; Charoonratana, V.T.; Tartaglia, M.; et al. Mutations in PAX2 Associate with Adult-Onset FSGS. J. Am. Soc. Nephrol. 2014, 25, 1942–1953. [Google Scholar] [CrossRef]
- Fu, W.-Y.; Chen, Y.; Sahin, M.; Zhao, X.-S.; Shi, L.; Bikoff, J.B.; Lai, K.-O.; Yung, W.-H.; Fu, A.K.Y.; Greenberg, M.E.; et al. Cdk5 Regulates EphA4-Mediated Dendritic Spine Retraction through an Ephexin1-Dependent Mechanism. Nat. Neurosci. 2007, 10, 67–76. [Google Scholar] [CrossRef]
- Feng, R.; Wen, J. Overview of the Roles of Sox2 in Stem Cell and Development. Biol. Chem. 2015, 396, 883–891. [Google Scholar] [CrossRef]
- Lalli, M.A.; Jang, J.; Park, J.-H.C.; Wang, Y.; Guzman, E.; Zhou, H.; Audouard, M.; Bridges, D.; Tovar, K.R.; Papuc, S.M.; et al. Haploinsufficiency of BAZ1B Contributes to Williams Syndrome through Transcriptional Dysregulation of Neurodevelopmental Pathways. Hum. Mol. Genet. 2016, 25, 1294–1306. [Google Scholar] [CrossRef]
- Bakrania, P.; Efthymiou, M.; Klein, J.C.; Salt, A.; Bunyan, D.J.; Wyatt, A.; Ponting, C.P.; Martin, A.; Williams, S.; Lindley, V.; et al. Mutations in BMP4 Cause Eye, Brain, and Digit Developmental Anomalies: Overlap between the BMP4 and Hedgehog Signaling Pathways. Am. J. Hum. Genet. 2008, 82, 304–319. [Google Scholar] [CrossRef]
- Ashe, A.; Morgan, D.K.; Whitelaw, N.C.; Bruxner, T.J.; Vickaryous, N.K.; Cox, L.L.; Butterfield, N.C.; Wicking, C.; Blewitt, M.E.; Wilkins, S.J.; et al. A Genome-Wide Screen for Modifiers of Transgene Variegation Identifies Genes with Critical Roles in Development. Genome Biol. 2008, 9, R182. [Google Scholar] [CrossRef]
- Shilton, D.; Breski, M.; Dor, D.; Jablonka, E. Human Social Evolution: Self-Domestication or Self-Control? Front. Psychol. 2020, 11, 134. [Google Scholar] [CrossRef]
- Kuhlwilm, M.; Boeckx, C. A Catalog of Single Nucleotide Changes Distinguishing Modern Humans from Archaic Hominins. Sci. Rep. 2019, 9, 8463. [Google Scholar] [CrossRef] [PubMed]
- Zanella, M.; Vitriolo, A.; Andirko, A.; Martins, P.T.; Sturm, S.; O’Rourke, T.; Laugsch, M.; Malerba, N.; Skaros, A.; Trattaro, S.; et al. Dosage Analysis of the 7q11.23 Williams Region Identifies BAZ1B as a Major Human Gene Patterning the Modern Human Face and Underlying Self-Domestication. Sci. Adv. 2019, 5, eaaw7908. [Google Scholar] [CrossRef]
- Von Holdt, B.M.; Ji, S.S.; Aardema, M.L.; Stahler, D.R.; Udell, M.A.R.; Sinsheimer, J.S. Activity of Genes with Functions in Human Williams–Beuren Syndrome Is Impacted by Mobile Element Insertions in the Gray Wolf Genome. Genome Biol. Evol. 2018, 10, 1546–1553. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.S.; Wrangham, R.W.; Fitch, W.T. The “Domestication Syndrome” in Mammals: A Unified Explanation Based on Neural Crest Cell Behavior and Genetics. Genetics 2014, 197, 795–808. [Google Scholar] [CrossRef] [PubMed]
- MacCallum, D.E.; Losada, A.; Kobayashi, R.; Hirano, T. ISWI Remodeling Complexes in Xenopus Egg Extracts: Identification as Major Chromosomal Components That Are Regulated by INCENP-Aurora B. Mol. Biol. Cell 2002, 13, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Cuvier, O.; Hirano, T. Chromosome Condensation by a Human Condensin Complex InXenopus Egg Extracts. J. Biol. Chem. 2001, 276, 5417–5420. [Google Scholar] [CrossRef]
- Ribeyre, C.; Zellweger, R.; Chauvin, M.; Bec, N.; Larroque, C.; Lopes, M.; Constantinou, A. Nascent DNA Proteomics Reveals a Chromatin Remodeler Required for Topoisomerase I Loading at Replication Forks. Cell Rep. 2016, 15, 300–309. [Google Scholar] [CrossRef]
- Ohta, S.; Taniguchi, T.; Sato, N.; Hamada, M.; Taniguchi, H.; Rappsilber, J. Quantitative Proteomics of the Mitotic Chromosome Scaffold Reveals the Association of BAZ1B with Chromosomal Axes*. Mol. Cell. Proteomics 2019, 18, 169–181. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Y.; Li, Y.; Zhang, J.; Liu, Y.; Li, Y.; Deng, Y.; Wang, S. Williams Syndrome Transcription Factor Is a Target of Pro-Oncogenic Ser158 Phosphorylation Mediated by Ras-MAPK Pathway in Human Breast Cancer. Int. J. Clin. Exp. Pathol. 2016, 9, 1668–1675. [Google Scholar]
- Dunn, K.L.; Espino, P.S.; Drobic, B.; He, S.; Davie, J.R. The Ras-MAPK Signal Transduction Pathway, Cancer and Chromatin Remodeling. Biochem. Cell Biol. 2005, 83, 1–14. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.-Y.; Wang, S.-Q.; Li, M.; Long, Y.-H.; Li, Y.-F.; Liu, Y.-K.; Li, Y.-H.; Wang, Y.-Q.; Mi, J.-S.; et al. WSTF Acetylation by MOF Promotes WSTF Activities and Oncogenic Functions. Oncogene 2020, 39, 5056–5067. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Costa, M.; Sun, H. Structure and Function of Histone Acetyltransferase MOF. AIMS Biophys. 2015, 2, 555–569. [Google Scholar] [CrossRef]
- McBurney, M.W.; Clark-Knowles, K.V.; Caron, A.Z.; Gray, D.A. SIRT1 Is a Highly Networked Protein That Mediates the Adaptation to Chronic Physiological Stress. Genes Cancer 2013, 4, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Ocaña, A.; Díez-González, L.; Esparís-Ogando, A.; Montero, J.C.; Amir, E.; Pandiella, A. Neuregulin Expression in Solid Tumors: Prognostic Value and Predictive Role to Anti-HER3 Therapies. Oncotarget 2016, 7, 45042–45051. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, S.-Q.; Long, Y.-H.; Chen, S.; Li, Y.-F.; Zhang, J.-H. KRASG12 Mutant Induces the Release of the WSTF/NRG3 Complex, and Contributes to an Oncogenic Paracrine Signaling Pathway. Oncotarget 2016, 7, 53153–53164. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Zhang, X.-T.; Liu, X.-L.; Fan, L.; Li, C.; Sun, Y.; Liang, X.-H.; Wang, J.-B.; Mei, Q.-B.; Zhang, F.; et al. WSTF Promotes Proliferation and Invasion of Lung Cancer Cells by Inducing EMT via PI3K/Akt and IL-6/STAT3 Signaling Pathways. Cell. Signal. 2016, 28, 1673–1682. [Google Scholar] [CrossRef]
- Jamesdaniel, S.; Hu, B.; Kermany, M.H.; Jiang, H.; Ding, D.; Coling, D.; Salvi, R. Noise Induced Changes in the Expression of P38/MAPK Signaling Proteins in the Sensory Epithelium of the Inner Ear. J. Proteomics 2011, 75, 410–424. [Google Scholar] [CrossRef]
- Smith, C.L.; Peterson, C.L. ATP-Dependent Chromatin Remodeling. Curr. Top. Dev. Biol. 2005, 65, 115–148. [Google Scholar] [CrossRef]
- Clapier, C.R.; Cairns, B.R. The Biology of Chromatin Remodeling Complexes. Annu. Rev. Biochem. 2009, 78, 273–304. [Google Scholar] [CrossRef]
- Segal, E.; Fondufe-Mittendorf, Y.; Chen, L.; Thåström, A.; Field, Y.; Moore, I.K.; Wang, J.-P.Z.; Widom, J. A Genomic Code for Nucleosome Positioning. Nature 2006, 442, 772–778. [Google Scholar] [CrossRef]
- Blossey, R.; Schiessel, H. The Latest Twists in Chromatin Remodeling. Biophys. J. 2018, 114, 2255–2261. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharif, S.B.; Zamani, N.; Chadwick, B.P. BAZ1B the Protean Protein. Genes 2021, 12, 1541. https://doi.org/10.3390/genes12101541
Sharif SB, Zamani N, Chadwick BP. BAZ1B the Protean Protein. Genes. 2021; 12(10):1541. https://doi.org/10.3390/genes12101541
Chicago/Turabian StyleSharif, Shahin Behrouz, Nina Zamani, and Brian P. Chadwick. 2021. "BAZ1B the Protean Protein" Genes 12, no. 10: 1541. https://doi.org/10.3390/genes12101541
APA StyleSharif, S. B., Zamani, N., & Chadwick, B. P. (2021). BAZ1B the Protean Protein. Genes, 12(10), 1541. https://doi.org/10.3390/genes12101541