
How to Choose a Good Marker to Analyze the Olive Germplasm (Olea europaea L.) and Derived Products


 , , ,
, , ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Morphological Markers
3. Molecular Markers
3.1. RFLP Markers
3.2. RAPD Markers
3.3. AFLP Markers
3.4. SCAR and CAPS Markers
3.5. ISSRs
3.6. SSR Marker
3.7. EST-SSR
3.8. SNP Markers
3.9. Molecular Markers Based on Transcriptome Analysis
3.10. Organelle Based and Ribosomal Markers
3.11. The Molecular Markers Used for Olive Oil Traceability Purposes
3.12. Tips for Choosing the Best Molecular Marker to Dissect the Olive Diversity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Besnard, G. Origin and Domestication. In The Olive Tree Genome; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–12. ISBN 978-3-319-48887-5. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAOSTAT) Database. Available online: http://www.fao.org/faostat/en/#data/ (accessed on 9 July 2021).
- Lanza, B.; Ninfali, P. Antioxidants in Extra Virgin Olive Oil and Table Olives: Connections between Agriculture and Processing for Health Choices. Antioxidants 2020, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Green, P.S.; Wickens, G.E. The Olea europaea complex. In The Davis & Hedge Festschrift; Tan, K., Ed.; Edinburgh University Press: Edinburgh, UK, 1989; pp. 287–299. [Google Scholar]
- Besnard, G.; Baali-Cherif, D. Coexistence of diploids and triploids in a Saharan relict olive: Evidence from nuclear microsatellite and flow cytometry analyses. C. R. Biol. 2009, 332, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Green, P.S. A revision of Olea L. (Oleaceae). Kew Bull. 2002, 57, 91–140. [Google Scholar] [CrossRef]
- Besnard, G.; Bervillé, A. Multiple origins for Mediterranean olive (Olea europaea L. ssp. europaea) based upon mitochondrial DNA polymorphisms. C. R. Acad. Sci. Ser. III Sci. Vie 2000, 323, 173–181. [Google Scholar] [CrossRef]
- Vossen, P. Olive oil: History, production, and characteristics of the world’s classic oils. HortScience 2007, 42, 1093–1100. [Google Scholar] [CrossRef] [Green Version]
- Breton, C.; Pinatel, C.; Medail, F.; Bonhommea, F.; Bervillé, A. Comparison between classical and Bayesian methods to investigate the history of olive cultivars using SSR-polymorphisms. Plant Sci. 2008, 175, 524–532. [Google Scholar] [CrossRef]
- Besnard, G.; El Bakkali, A.; Haouane, H.; Baali-Cherif, D.; Moukhli, A.; Khadari, B. Population genetics of Mediterranean and Saharan olives: Geographic patterns of differentiation and evidence for early-generations of admixture. Ann. Bot. 2013, 112, 1293–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dίez, C.M.; Trujillo, I.; Martinez-Uriroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S. Olive domestication and diversification in the Mediterranean basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef]
- D’Agostino, N.; Taranto, F.; Camposeo, S.; Mangini, G.; Fanelli, V.; Gadaleta, S.; Miazzi, M.M.; Pavan, S.; di Rienzo, V.; Sabetta, W.; et al. GBS-derived SNP catalogue unveiled wide genetic variability and geographical relationships of Italian olive cultivars. Sci. Rep. 2018, 8, 15877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muzzalupo, I. Olive Germplasm: Italian Catalogue of Olive Varieties; InTech: Rijeka, Croatia, 2012; ISBN 978-953-51-0884-9. [Google Scholar] [CrossRef]
- Muzzalupo, I.; Vendramin, G.G.; Chiappetta, A. Genetic Biodiversity of Italian Olives (Olea europaea) Germplasm Analyzed by SSR Markers. Sci. World J. 2014, 2014, 296590. [Google Scholar] [CrossRef] [Green Version]
- Miazzi, M.M.; di Rienzo, V.; Mascio, I.; Montemurro, C.; Sion, S.; Sabetta, W.; Vivaldi, G.A.; Camposeo, S.; Caponio, F.; Squeo, G.; et al. Re.Ger.O.P.: An Integrated Project for the Recovery of Ancient and Rare Olive Germplasm. Front. Plant Sci. 2020, 11, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saddoud Debbabi, O.; Miazzi, M.M.; Elloumi, O.; Fendri, M.; Ben Amar, F.; Savoia, M.; Sion, S.; Souabni, H.; Mnasri, S.R.; Ben Abdelaali, S.; et al. Recovery, Assessment, and Molecular Characterization of Minor Olive Genotypes in Tunisia. Plants 2020, 9, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falek, W.; Sion, S.; Montemurro, C.; Mascio, I.; Gadaleta, S.; Fanelli, V.; Savoia, M.A.; Piarulli, L.; Bechkri, S.; Khelifi, D.; et al. Molecular diversity and ecogeographic distribution of Algerian wild olives (Olea europaea subsp. europaea var. sylvestris). Sci. Agric. 2021, 79, e20200308. [Google Scholar] [CrossRef]
- Belaj, A.; Caballero, J.M.; Barranco, D.; Rallo, L.; Trujillo, I. Genetic characterization and identification of new accessions from Syria in an olive germplasm Bank by means of RAPD markers. Euphytica 2003, 134, 261–268. [Google Scholar] [CrossRef]
- Noormohammadi, Z.; Hosseini-Mazinani, M.; Trujillo, I.; Rallo, L.; Belaj, A.; Sadeghizadeh, M. Identification and classification of main Iranian olive cultivars using microsatellite markers. HortScience 2007, 42, 1545–1550. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, V.; Miazzi, M.M.; Fanelli, V.; Sabetta, W.; Montemurro, C. The preservation and characterization of Apulian olive germplasm biodiversity. Acta Hortic. 2018, 1199, 1–6. [Google Scholar] [CrossRef]
- Hosseini-Mazinani, M.; Mariotti, R.; Torkzaban, B.; Sheikh-Hassani, M.; Ataei, S.; Cultrera, N.G.M.; Pandolfi, S.; Baldoni, L. High genetic diversity detected in olives beyond the boundaries of the Mediterranean sea. PLoS ONE 2014, 9, e93146. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.; Mariotti, R.; Bagnoli, F.; Costantini, L.; Cultrera, N.G.M.; Arzani, K.; Pandolfi, S.; Vendramin, G.G.; Torkzaban, B.; Hosseini-Mazinani, M.; et al. The eastern part of the Fertile Crescent concealed an unexpected route of olive (Olea europaea L.) differentiation. Ann. Bot. 2017, 119, 1305–1318. [Google Scholar] [CrossRef]
- Marra, F.P.; Caruso, T.; Costa, F.; Di Vaio, C.; Mafrica, R.; Marchese, A. Genetic relationships, structure and parentage simulation among the olive tree (Olea europaea L. subsp. europaea) cultivated in Southern Italy revealed by SSR markers. Tree Genet. Genomes 2013, 9, 961–973. [Google Scholar] [CrossRef]
- Lazović, B.; Adakalić, M.; Pucci, C.; Perović, T.; Bandelj, D.; Belaj, A.; Mariotti, R.; Baldoni, L. Characterizing ancient and local olive germplasm from Montenegro. Sci. Hortic. 2016, 209, 117–123. [Google Scholar] [CrossRef]
- Caruso, T.; Marra, F.P.; Costa, F.; Campisi, G.; Macaluso, L.; Marchese, A. Genetic diversity and clonal variation within the main Sicilian olive cultivars based on morphological traits and microsatellite markers. Sci. Hortic. 2014, 180, 130–138. [Google Scholar] [CrossRef]
- Ipek, A.; Barut, E.; Gulen, H.; Ipek, M. Assessment of inter- and intra-cultivar variations in olive using SSR markers. Sci. Agric. 2012, 69, 327–335. [Google Scholar] [CrossRef]
- Ganino, T.; Bartolini, G.; Fabbri, A. The classification of olive germplasm—A review. J. Hortic. Sci. Biotechnol. 2006, 81, 319–334. [Google Scholar] [CrossRef]
- Garcia-Muñoz, S.; Muñoz-Organero, G.; de Andres, M.T.; Cabello, F. Ampelography-an old technique with future uses: The case of minor varieties of Vitis vinifera L. from The Balearic Islands. OENO One 2011, 45, 125–137. [Google Scholar] [CrossRef]
- Ciferri, R.; Breviglieri, N. Introduzione ad una classificazione morfo—Ecologica dell’olivo coltivato in Italia. L’Olivicoltore 1942, 1, 1–2. [Google Scholar]
- Baldini, E.; Scaramuzzi, F. Ulteriori indagini sulla validità del metodo bio-statistico nella descrizione e classificazione delle cultivar di olivo. Ann. Ist. Sper. Agron. 1955, 9, 171–186. [Google Scholar]
- Damigella, P. Variabilità dei caratteri biometrici dell’olivo e impiego delle funzioni discriminanti. Ric. Sci. 1960, 4, 522–530. [Google Scholar]
- Rallo, L.; Barranco, D. Autochthonous olive cultivars in Andalusia. Acta Hortic. 1983, 140, 169–179. [Google Scholar] [CrossRef]
- UPOV. Guidelines for the Conduct of Tests for Distinctness, Homogeneity and Stability: Olive; International Union for the Protection of New Varieties of Plants: Genéve, Switzerland, 1985. [Google Scholar]
- IOOC Encyclopédie Mondiale de L’Olivier; Conseil Oléicole International: Madrid, Spain, 1997.
- Barranco, D.; Cimato, A.; Fiorino, P.; Rallo, L.; Touzani, A.; Castaneda, C.; Serafin, F.; Trujillo, I. World Catalogue of Olive Varieties; Consejo Oleìcola Internacional: Madrid, Spain, 2000; 360p. [Google Scholar]
- Terral, J.; Alonso, N.; Buxo i Capdevila, R.; Chatti, N.; Fabre, L.; Fiorentino, G.; Marinval, P.; Perez Jorda, G.; Pradat, B.; Rovira, N.; et al. Historical biogeography of olive domestication (Olea europaea L.) as revealed by geometrical morphometry applied to biological and archaeological material. J. Biogeogr. 2004, 31, 63–77. [Google Scholar] [CrossRef]
- Blazakis, K.N.; Kosma, M.; Kostelenos, G.; Baldoni, L.; Bufacchi, M.; Kalaitzis, P. Description of olive morphological parameters by using open access software. Plant Methods 2017, 13, 111. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Gálvez, F.J.; Pérez-Mohedano, D.; de la Rosa-Navarro, R.; Belaj, A. High-throughput analysis of the canopy traits in the worldwide olive germplasm bank of Córdoba using very high-resolution imagery acquired from unmanned aerial vehicle (UAV). Sci. Hortic. 2021, 278, 109851. [Google Scholar] [CrossRef]
- Cimato, A.; Cantini, C.; Sani, G.; Marranci, M. Il Germoplasma dell’Olivo in Toscana; Regione Toscana: Florence, Italy, 1993. [Google Scholar]
- Hairi, I. Etude des caracteristiques pomologiques des cultivars plus importants de l’olivier en Albanie. In Proceedings of the Atti del Convegno L’Olivicoltura Mediterranea, Rende, Italy, 26–28 January 1995. [Google Scholar]
- Pannelli, G.; Alfei, B.; Santinelli, A. Varietà d’Olivo nelle Marche; ASSAM: Ancona, Italy, 1998. [Google Scholar]
- Pannelli, G.; Alfei, B.; D’Ambrosio, A.; Rosati, S.; Famiani, F. Varietà di Olivo in Umbria; Pliniana: Perugia, Italy, 2000. [Google Scholar]
- De Castro, A.I.; Rallo, P.; Suárez, M.P.; Torres-Sánchez, J.; Casanova, L.; Jiménez-Brenes, F.M.; Morales-Sillero, A.; Jiménez, M.R.; López-Granados, F. High-Throughput System for the Early Quantification of Major Architectural Traits in Olive Breeding Trials Using UAV Images and OBIA Techniques. Front. Plant. Sci. 2019, 10, 1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, R.; Leolini, C.; Costafreda-Aumedes, S.; Leolini, L.; Bindi, M.; Zaldei, A.; Moriondo, M. Performances Evaluation of a Low-Cost Platform for High-Resolution Plant Phenotyping. Sensors 2020, 20, 3150. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2017, 32, 261–285. [Google Scholar] [CrossRef] [Green Version]
- Grati-Kamoun, N.; Mahmoud, F.L.; Rebaï, A.; Gargouri, A.; Panaud, O.; Saar, A. Genetic Diversity of Tunisian Olive Tree (Olea europaea L.) Cultivars Assessed by AFLP Markers. Genet. Resour. Crop Evol. 2006, 53, 265–275. [Google Scholar] [CrossRef]
- di Rienzo, V.; Sion, S.; Taranto, F.; D’Agostino, N.; Montemurro, C.; Fanelli, V.; Sabetta, W.; Boucheffa, S.; Tamendjari, A.; Pasqualone, A.; et al. Genetic flow among olive populations within the Mediterranean basin. Peer J. 2018, 6, e5260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piarulli, L.; Savoia, M.A.; Taranto, F.; D’Agostino, N.; Sardaro, R.; Girone, S.; Gadaleta, S.; Fucili, V.; De Giovanni, C.; Montemurro, C.; et al. A robust DNA isolation protocol from filtered commercial olive oil for PCR-based fingerprinting. Foods 2019, 8, 462. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.C. Restriction fragment length polymorphism (RFLP). Am. J. Phys. Anthropol. 1989, 32, 159–184. [Google Scholar] [CrossRef]
- Amane, M.; Ouazzani, N.; Lumaret, R.; Debain, C. Chloroplast-DNA variation in the wild and cultivated olives (Olea europaea L.) of Morocco. Euphytica 2000, 116, 59–64. [Google Scholar] [CrossRef]
- Lumaret, R.; Amane, M.; Ouazzani, N.; Baldoni, L.; Debain, C. Chloroplast DNA variation in the cultivated and wild olive taxa of the genus Olea L. Theor. Appl. Genet. 2000, 101, 547–553. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P.; Besnard, G.; Khadari, B. Systematics, ecology and phylogeographic significance of Olea europaea L. ssp. maroccana (Greuter & Burdet) P. Vargas et al., a relictual olive tree in south-west Morocco. Bot. J. Linn. Soc. 2001, 137, 249–266. [Google Scholar]
- Besnard, G.; Bervillè, A. On chloroplast DNA variations in the olive (Olea europaea L.) complex: Comparison of RFLP and PCR polymorphisms. Theor. Appl. Genet. 2002, 104, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Khadari, B.; Breton, C.; Moutier, N.; Roger, J.; Besnard, G.; Bervillé, A.; Dosba, F. The use of molecular markers for germplasm management in a French olive collection. Theor. Appl. Genet. 2003, 106, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Besnard, G.; Baradat, P.H.; Chevalier, D.; Tagmount, A.; Bervillé, A. Genetic differentiation in the olive complex (Olea europaea) revealed by RAPDs and RFLPs in the rRNA genes. Genet. Resour. Crop Evol. 2001, 48, 165–182. [Google Scholar] [CrossRef]
- Cavallotti, A.; Regina, T.M.; Quagliariello, C. New sources of cytoplasmic diversity in the Italian population of Olea europaea L. as revealed by RFLP analysis of mitochondrial DNA: Characterization of the cox3 locus and possible relationship with cytoplasmic male sterility. Plant Sci. 2003, 164, 241–252. [Google Scholar] [CrossRef]
- De la Rosa, R.; Angiolillo, A.; Guerrero, C.; Pellegrini, M.; Rallo, L.; Besnard, G.; Bervillé, A.; Martín, A.; Baldoni, L. A first linkage map of olive (Olea europaea L.) cultivars using RAPD, AFLP, RFLP and SSR markers. Theor. Appl. Genet. 2003, 106, 1273–1282. [Google Scholar] [CrossRef]
- Wu, S.B.; Collins, G.; Sedgley, M. A molecular linkage map of olive (Olea europaea L.) based on RAPD, microsatellite, and SCAR markers. Genome 2004, 47, 26–35. [Google Scholar] [CrossRef]
- Williams, J.G.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belaj, A.; Trujillo, I.; De la Rosa, R.; Rallo, L.; Gimenez, M.J. Polymorphism and discrimination capacity of randomly amplified polymorphic markers in an olive germplasm bank. J. Am. Soc. Hortic. Sci. 2001, 126, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Durgac, C.; Kiyga, Y.; Ulas, M. Comparative molecular analysis of old olive (Olea europaea L.) genotypes from Eastern Mediterranean Region of Turkey. Afr. J. Biotechnol. 2010, 9, 428–433. [Google Scholar]
- Iqbal, M.Z.; Jamil, S.; Mehmood, A.; Shahzad, R. Identification of seven olive varieties using rapd molecular markers. J. Agric. Res. 2019, 57, 7–14. [Google Scholar]
- Hess, J.; Kadereit, J.W.; Vargas, P. The colonization history of Olea europea L. in Macaronesia based on internal transcribed spacer 1 (ITS-1) sequences, randomly amplified polymorphic DNAs (RAPD) and intersimple sequence repeats (ISSR). Mol. Ecol. 2000, 9, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van De Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [Green Version]
- Angiolillo, A.; Mencuccini, M.; Baldoni, L. Olive genetic diversity assessed using amplified fragment length polymorphisms. Theor. Appl. Genet. 1999, 98, 411–421. [Google Scholar] [CrossRef]
- Montemurro, C.; Simeone, R.; Pasqualone, A.; Ferrara, E.; Blanco, A. Genetic relationships and cultivar identification among 112 olive accessions using AFLP and SSR markers. J. Hortic. Sci. Biotechnol. 2005, 80, 105–110. [Google Scholar] [CrossRef]
- Owen, C.A.; Bita, E.; Banilas, G.; Hajjar, S.E.; Sellianakis, V.; Aksoy, U.; Hepaksoy, S.; Chamoun, R.; Talhook, S.N.; Metzidakis, I.; et al. AFLP reveals structural details of genetic diversity within cultivated olive germplasm from eastern Mediterranean. Theor. Appl. 2005, 110, 1169–1176. [Google Scholar] [CrossRef]
- Albertini, E.; Torricelli, R.; Bitocchi, E.; Raggi, L.; Marconi, G.; Pollastri, L.; Di Minco, G.; Battistini, A.; Papa, R.; Veronesi, F. Structure of genetic diversity in Olea europaea L. cultivars from central Italy. Mol. Breed. 2011, 27, 533–547. [Google Scholar] [CrossRef]
- El Aabidine, A.Z.; Charafi, J.; Grout, C.; Doligez, A.; Santoni, S.; Moukhli, A.; Jay-Allemand, C.; El Modafar, C.; Khadari, B. Construction of a Genetic Linkage Map for the Olive Based on AFLP and SSR Markers. Crop Sci. 2010, 50, 2291–2302. [Google Scholar] [CrossRef]
- Khadari, B.; El Aabidine, A.Z.; Grout, C.; Ben Sadok, I.; Doligez, A.; Moutier, N.; Santoni, S.; Sadok, E.C. A Genetic Linkage Map of Olive Based on AFLP, ISSR and SSR Markers. J. Am. Soc. Hortic. Sci. 2013, 135, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Sadok, I.B.; Celton, J.M.; Essalouh, L.; El Aabidine, A.Z.; Garcia, G.; Martinez, S.; Grati-Kamoun, N.; Rebai, A.; Costes, E.; Khadari, B. QTL Mapping of Flowering and Fruiting Traits in Olive. PLoS ONE 2014, 9, e62831. [Google Scholar] [CrossRef] [Green Version]
- Atienza, S.G.; de la Rosa, R.; León, L.; Martín, A.; Belaj, A. Identification of QTL for agronomic traits of importance for olive breeding. Mol. Breed. 2014, 34, 725–737. [Google Scholar] [CrossRef]
- Bautista, R.; Crespillo, R.; Cánovas, F.M.; Claros, M.G. Identification of olive-tree cultivars with SCAR markers. Euphytica 2003, 129, 33–41. [Google Scholar] [CrossRef]
- Busconi, M.; Sebastiani, L.; Fogher, C. Development of SCAR Markers for Germplasm Characterisation in Olive Tree (Olea europea L.). Mol. Breed. 2006, 17, 59–68. [Google Scholar] [CrossRef]
- De la Torre, F.; Canovas, F.M.; Claros, M.G. Isolation of DNA from olive oil and oil sediments: Application in oil fingerprinting. J. Food Agric. Environ. 2004, 2, 84–89. [Google Scholar]
- Pafundo, S.M.; Agrimonti, C.; Maestri, E.; Marmiroli, N. Applicability of SCAR markers to food genomics: Olive oil traceability. J. Agric. Food Chem. 2007, 55, 6052–6059. [Google Scholar] [CrossRef] [PubMed]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef]
- Pasqualone, A.; Caponio, F.; Blanco, A. Inter-simple sequence repeat DNA markers for identification of drupes from different Olea europaea L. cultivars. Eur. Food Res. Technol. 2001, 213, 240–243. [Google Scholar] [CrossRef]
- Vargas, P.; Garmendia, F.; Hess, J.; Kadereit, J.W. Olea europaea subsp. guanchica and subsp. maroccana (Oleaceae), two new names for olive tree relatives. Ann. Jard. Bot. Madr. 2001, 58, 360–361. [Google Scholar]
- Essadki, M.; Ouazzani, N.; Lumaret, R.; Moumni, M. ISSR Variation in Olive-tree Cultivars from Morocco and other Western Countries of the Mediterranean Basin. Genet. Resour. Crop Evol. 2006, 53, 475–482. [Google Scholar] [CrossRef]
- Martins-Lopes, P.; Lima-Brito, J.; Gomes, S.; Meirinhos, J.; Santos, L.; Guedes-Pinto, H. RAPD and ISSR molecular markers in Olea europaea L.: Genetic variability and molecular cultivar identification. Genet. Resour. Crop Evol. 2007, 54, 117–128. [Google Scholar] [CrossRef]
- Ergun, K. ISSR analysis for determination of genetic diversity and relationship in eight Turkish olive (Olea europaea L.) cultivars. Not. Bot. Horti Agrobot. Cluj-Napoca 2015, 43, 96–99. [Google Scholar]
- Abood, A.A.; Al-Ansari, A.M.; Migdadi, H.M.; Okla, M.K.; Assaeed, A.M.; Hegazy, A.K.; Alshameri, A.M.; Khan, M.A. Molecular and phytochemical analysis of wild type and olive cultivars grown under Saudi Arabian environment. 3 Biotech. 2017, 7, 289:1–289:14. [Google Scholar] [CrossRef] [PubMed]
- Kaya, E.; Yilmaz-gokdogan, E. Using Two Retrotransposon Based Marker Systems (IRAP and REMAP) for Molecular Characterization of Olive (Olea europaea L.) Cultivars. Not. Bot Horti Agrobot. Cluj-Napoca 2016, 44, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–222. [Google Scholar] [CrossRef]
- Condit, R.; Hubbell, S.P. Abundance and DNA sequence of two-base repeat regions in tropical tree genomes. Genome 1991, 34, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.A.F.; Benchimol, L.L.; Barbosa, A.M.M.; Geraldi, I.O.; Souza, C.L., Jr.; de Souza, A.P. Comparison of RAPD, RFLP, AFLP and SSR markers for diversity studies in tropical maize inbred lines. Genet. Mol. Biol. 2004, 27, 579–588. [Google Scholar] [CrossRef]
- Sefc, K.M.; Lopes, M.S.; Mendoca, D.; Rodrigues Dos Santos, M.; Laimer Da Camara Machado, M.; Da Camara Machado, A. Identification of microsatellite loci in olive (Olea europaea L) and their characterization in Italian and Iberian olive trees. Mol. Ecol. 2000, 9, 1171–1173. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, G.; Marrazzo, M.T.; Marconi, R.; Cimato, A.; Testolin, R. Microsatellite markers isolated in olive (Olea europea L.) are suitable for individual fingerprinting and reveal polymorphism within ancient cultivars. Theor. Appl. Genet. 2002, 104, 223–228. [Google Scholar] [CrossRef]
- Sebastiani, L.; Busconi, M. Recent developments in olive (Olea europaea L.) genetics and genomics: Applications in taxonomy, varietal identification, traceability and breeding. Plant Cell Rep. 2017, 36, 1345–1360. [Google Scholar] [CrossRef]
- Belaj, A.; Satovic, Z.; Cipriani, G.; Baldoni, L.; Testolin, R.; Rallo, L.; Trujillo, I. Comparative study of the discriminating capacity of RAPD, AFLP and SSR markers and of their effectiveness in establishing genetic relationships in olive. Theor. Appl. Genet. 2003, 107, 736–744. [Google Scholar] [CrossRef]
- Rotondi, A.; Magli, M.; Ricciolini, C.; Baldoni, L. Morphological and molecular analyses for the characterization of a group of Italian olive cultivars. Euphytica 2003, 132, 129–137. [Google Scholar] [CrossRef]
- Bandelj, D.; Jakše, J.; Javornik, B. Assessment of genetic variability of olive varieties by microsatellite and AFLP markers. Euphytica 2004, 136, 93–102. [Google Scholar] [CrossRef]
- Montemurro, C.; Simeone, R.; Blanco, A.; Saponari, M.; Bottalico, G.; Savino, V.; Martelli, G.P.; Pasqualone, A. Sanitary selection and molecular characterization of olive cultivars grown in Apulia. Acta Hortic. 2008, 791, 603–609. [Google Scholar] [CrossRef]
- Besnard, G.; Rubio de Casas, R.; Vargas, P. Plastid and nuclear DNA polymorphism reveals historical processes of isolation and reticulation in the olive tree complex (Olea europaea). J. Biogeogr. 2007, 34, 736–752. [Google Scholar] [CrossRef]
- Belaj, A.; Muñoz-Diez, C.; Baldoni, L.; Porceddu, A.; Barranco, D.; Satovic, Z. Genetic diversity and population structure of wild olives from the north-western Mediterranean assessed by SSR markers. Ann. Bot. 2007, 100, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erre, P.; Chessa, I.; Muñoz-Diez, C.; Belaj, A.; Rallo, L.; Trujillo, I. Genetic diversity and relationships between wild and cultivated olives (Olea europaea L.) in Sardinia as assessed by SSR markers. Genet. Resour. Crop Evol. 2010, 57, 41–54. [Google Scholar] [CrossRef]
- Belaj, A.; Muñoz-Diez, C.; Baldoni, L.; Satovic, Z.; Barranco, D. Genetic diversity and relationships of wild and cultivated olives at regional level in Spain. Sci. Hortic. 2010, 124, 323–330. [Google Scholar] [CrossRef]
- Díez, C.M.; Trujillo, I.; Barrio, E.; Belaj, A.; Barranco, D.; Rallo, L. Centennial olive trees as a reservoir of genetic diversity. Ann. Bot. 2011, 108, 797–807. [Google Scholar] [CrossRef] [Green Version]
- Hannachi, H.; Breton, C.; Msallem, M.; El Hadj, S.B.; El Gazzah, M.; Bervillé, A. Genetic relationships between cultivated and wild olive trees (Olea europaea L. var. europaea and var. sylvestris) based on nuclear and chloroplast SSR markers. Nat. Resour. 2010, 1, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Boucheffa, S.; Miazzi, M.M.; di Rienzo, V.; Mangini, G.; Fanelli, V.; Tamendjari, A.; Pignone, D.; Montemurro, C. The coexistence of oleaster and traditional varieties genetic diversity and population structure in Algerian olive (Olea europaea) germplasm. Genet. Resour. Crop Evol. 2017, 64, 379–390. [Google Scholar] [CrossRef]
- Boucheffa, S.; Tamendjari, A.; Sanchez-Gimeno, A.C.; Rovellini, P.; Venturini, S.; di Rienzo, V.; Miazzi, M.M.; Montemurro, C. Diversity Assessment of Algerian Wild and Cultivated Olives (Olea europaea L.) by Molecular, Morphological, and Chemical Traits. Eur. J. Lipid Sci. Technol. 2019, 121, 1800302. [Google Scholar] [CrossRef] [Green Version]
- Barazani, O.; Keren-Keiserman, A.; Westberg, E.; Hanin, N.; Dag, A.; Ben-Ari, G.; Fragman-Sapir, O.; Tugendhaft, Y.; Kerem, Z.; Kadereit, J.W. Genetic variation of naturally growing olive trees in Israel: From abandoned groves to feral and wild? BMC Plant Biol. 2016, 16, 261. [Google Scholar] [CrossRef] [Green Version]
- Aumeeruddy-Thomas, Y.; Moukhli, A.; Haouane, H.; Khadari, B. Ongoing domestication and diversification in grafted olive–oleaster agroecosystems in Northern Morocco. Reg. Environ. Chang. 2017, 17, 1315–1328. [Google Scholar] [CrossRef]
- Rallo, P.; Tenzer, I.; Gessler, C.; Baldoni, L.; Dorado, G.; Martin, A. Transferability of olive microsatellite loci across the genus Olea. Theor. Appl. Genet. 2003, 107, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Hannachi, H.; Sommerlatte, H.; Breton, C.; Msallem, M.; El Gazzah, M.; El Hadj, S.B.; Bervillé, A. Oleaster (var. sylvestris) and subsp. cuspidata are suitable genetic resources for improvement of the olive (Olea europaea subsp. europaea var. europaea). Genet. Resour. Crop Evol. 2009, 56, 393–403. [Google Scholar] [CrossRef]
- Kaya, H.B.; Cetin, O.; Kaya, H.; Sahin, M.; Sefer, F.; Kahraman, A.; Tanyolac, B. SNP discovery by Illumina-based transcriptome sequencing of the olive and the genetic characterization of Turkish olive genotypes revealed by AFLP, SSR and SNP markers. PLoS ONE 2013, 8, e73674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xanthopoulou, A.; Ganopoulos, I.; Koubouris, G.; Tsaftaris, A.; Sergendani, C.; Kalivas, A.; Madesis, P. Microsatellite high-resolution melting (SSR-HRM) analysis for genotyping and molecular characterization of an Olea europaea germplasm collection. Plant. Genet. Resour. 2014, 12, 273–277. [Google Scholar] [CrossRef]
- Díaz-Rueda, P.; Franco-Navarro, J.D.; Messora, R.; Espartero, J.; Rivero-Núñez, C.M.; Aleza, P.; Capote, N.; Cantos, M.; García-Fernández, J.L.; de Cires, A.; et al. SILVOLIVE, a germplasm collection of wild subspecies with high genetic variability as a source of rootstocks and resistance genes for olive breeding. Front. Plant Sci. 2020, 11, 629. [Google Scholar] [CrossRef] [PubMed]
- Koehmstedt, A.M.; Aradhya, M.K.; Soleri, D.; Smith, J.L.; Polito, V.S. Molecular characterization of genetic diversity, structure, and differentiation in the olive (Olea europaea L.) germplasm collection of the United States Department of Agriculture. Genet. Resour. Crop Evol. 2011, 58, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Zelasco, S.; Salimonti, A.; Baldoni, L.; Mariotti, R.; Preece, J.E.; Aradhya, M.; Koehmstedt, A.M. Efficiency Of SSR Markers for Exploring Olive Germplasm Diversity through a Genetic Comparison between The USDA-NCGR and the CRA-OLI Olive Collections. Acta Hortic. 2014, 1057, 585–592. [Google Scholar] [CrossRef]
- Trentacoste, E.R.; Puertas, C.M. Preliminary characterization and morpho-agronomic evaluation of the olive germplasm collection of the Mendoza province (Argentina). Euphytica 2011, 177, 99–109. [Google Scholar] [CrossRef]
- Guerin, J.R.; Sweeney, S.M.; Collins, G.G.; Sedgley, M. The development of a genetic database to identify olive cultivars. J. Am. Soc. Hortic. Sci. 2002, 127, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Abdessemed, S.; Muzzalupo, I.; Benbouza, H. Assessment of genetic diversity among Algerian olive (Olea europaea L.) cultivars using SSR marker. Sci. Hortic. 2015, 192, 10–20. [Google Scholar] [CrossRef]
- Khadari, B.; El Bakkali, A.; Essalouh, L.; Tollon, C.; Pinatel, C.; Besnard, G. Cultivated olive diversification at local and regional scales: Evidence from the genetic characterization of French genetic resources. Front. Plant Sci. 2019, 10, 1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saddoud Deddabi, O.; Montemurro, C.; Ben Maachia, S.; Ben Amar, F.; Fanelli, V.; Gadaleta, S.; El Riachy, M.; Chehade, A.; Siblini, M.; Boucheffa, S.; et al. A Hot Spot of Olive Biodiversity in the Tunisian Oasis of Degache. Diversity 2020, 12, 358. [Google Scholar] [CrossRef]
- Saddoud Debbabi, O.; Rahmani Mnasri, S.; Ben Amar, F.; Ben Naceur, M.; Montemurro, C.; Miazzi, M.M. Applications of microsatellite markers for the characterization of olive genetic resources of Tunisia. Genes 2021, 12, 286. [Google Scholar] [CrossRef]
- Abuzayed, M.; Frary, A.; Doganlar, S. Genetic diversity of some Palestinian and Turkish olive (Olea europaea L.) germplasm determined with SSR markers. IUG J. Nat. Sci. 2018, 26, 10–17. [Google Scholar]
- Dastkar, E.; Soleimani, A.; Jafary, H.; Naghavi, M.R. Genetic and morphological variation in Iranian olive (Olea europaea L.) germplasm. Crop Breed. J. 2013, 3, 99–106. [Google Scholar]
- Ercisli, S.; Bencic, D.; Ipek, A.; Barut, E.; Liber, Z. Genetic relationships among olive (Olea europaea L.) cultivars native to Croatia and Turkey. J. Appl. Bot. 2013, 85, 144–149. [Google Scholar]
- Dervishi, A.; Jakše, J.; Ismaili, H.; Javornik, B.; Štajner, N. Comparative assessment of genetic diversity in Albanian olive (Olea europaea L.) using SSRs from anonymous and transcribed genomic regions. Tree Genet. Genomes 2018, 14, 53. [Google Scholar] [CrossRef]
- Gomes, S.; Martins-Lopes, P.; Lopes, J.; Guedes-Pinto, H. Assessing genetic diversity in Olea europaea L. using ISSR and SSR markers. Plant Mol. Biol. Rep. 2009, 27, 365–373. [Google Scholar] [CrossRef]
- Fernández i Martí, A.; Font i Forcada, C.; Socias i Company, R.; Rubio-Cabetas, M.J. Genetic relationships and population structure of local olive tree accessions from Northeastern Spain revealed by SSR markers. Acta Physiol. Plant. 2015, 37, 1726. [Google Scholar] [CrossRef]
- Rotondi, A.; Ganino, T.; Beghè, D.; Di Virgilio, N.; Morrone, L.; Fabbri, A.; Neri, L. Genetic and landscape characterization of ancient autochthonous olive trees in northern Italy. Plant Biosyst. 2018, 152, 1067–1074. [Google Scholar] [CrossRef]
- Bracci, T.; Busconi, M.; Fogher, C.; Sebastiani, L. Molecular studies in olive (Olea europaea L.): Overview on DNA markers applications and recent advances in genome analysis. Plant Cell Rep. 2011, 30, 449–462. [Google Scholar] [CrossRef]
- Sion, S.; Taranto, F.; Montemurro, C.; Mangini, G.; Camposeo, S.; Falco, V.; Gallo, A.; Mita, G.; Saddoud Debbabi, O.; Ben Amar, F.; et al. Genetic characterization of Apulian olive germplasm as potential source in new breeding programs. Plants 2019, 8, 268. [Google Scholar] [CrossRef] [Green Version]
- Arbeiter, A.; Hladnik, M.; Jakše, J.; Bandelj, D. Identification and validation of novel EST-SSR markers in olives. Sci. Agric. 2017, 74, 215–225. [Google Scholar] [CrossRef]
- Cruz, F.; Julca, I.; Gómez-Garrido, J.; Loska, D.; Marcet-Houben, M.; Cano, E.; Galán, B.; Frias, L.; Ribeca, P.; Derdak, S.; et al. Genome sequence of the olive tree, Olea europaea. Gigascience 2016, 5, 29. [Google Scholar] [CrossRef]
- Unver, T.; Wu, Z.; Sterck, L.; Turktas, M.; Lohaus, R.; Li, Z.; Yang, M.; He, L.; Deng, T.; Escalante, F.J.; et al. Wild olive genome and oil biosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, E9413–E9422. [Google Scholar] [CrossRef] [Green Version]
- Rao, G.; Zhang, J.; Liu, X.; Lin, C.; Xin, H.; Xue, L.; Wang, C. De novo assembly of a new Olea europaea genome accession using nanopore sequencing. Hortic. Res. 2021, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Long, C.; Pang, X.; Ning, D.; Wu, T.; Dong, M.; Han, X.; Guo, H. The newly developed genomic-SSR markers uncover the genetic characteristics and relationships of olive accessions. PeerJ 2020, 8, e8573. [Google Scholar] [CrossRef]
- Rekik, I.; Salimonti, A.; Grati Kamoun, N.; Muzzalupo, I.; Perri, E.; Rebai, A. Characterisation and identification of Tunisian olive tree varieties by microsatellite markers. HortScience 2008, 43, 1371–1376. [Google Scholar] [CrossRef]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Diez, C.M. Identification of the Worldwide Olive Germplasm Bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet. Genomes 2014, 10, 141–155. [Google Scholar] [CrossRef]
- Beghè, D.; Garcìa Molano, J.F.; Fabbri, A.; Ganino, T. Olive biodiversity in Colombia. A molecular study of local germplasm. Sci. Hortic. 2015, 189, 122–131. [Google Scholar] [CrossRef]
- Doveri, S.; Gil, F.G.; Díaz, A.; Reale, S.; Busconi, M.; da Câmara Machado, A.; Martín, A.; Fogher, C.; Donini, P.; Lee, D. Standardization of a set of microsatellite markers for use in cultivar identification studies in olive (Olea europaea L.). Sci. Hortic. 2008, 116, 367–373. [Google Scholar] [CrossRef]
- Baldoni, L.; Cultrera, N.G.; Mariotti, R.; Riccioloni, C.; Arcioni, S.; Vendramin, G.G.; Buonamici, A.; Porceddu, A.; Sarri, V.; Ojeda, M.A.; et al. A consensus list of microsatellites markers for olive genotyping. Mol. Breed. 2009, 24, 213–231. [Google Scholar] [CrossRef]
- Bartolini, G. Olea Databases. 2008. Available online: http://www.oleadb.it (accessed on 9 July 2021).
- Ben Ayed, R.; Ben Hassen, H.; Ennouri, K.; Ben Marzoug, R.; Rebai, A. OGDD (Olive Genetic Diversity Database): A microsatellite markers’ genotypes database of worldwide olive trees for cultivar identification and virgin olive oil traceability. Database 2016, 2016, bav090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.R.; Cornejo, P.; Bertoldi, V.; Ferrer, M.S.; Masuelli, R.W. Development of a microsatellite database for identification of olive (Olea europaea L.) cultivars in Mendoza, Argentina. Acta Hortic. 2014, 1057, 521–524. [Google Scholar] [CrossRef]
- Haddad, B.; Gristina, A.S.; Mercati, F.; Saadi, A.E.; Aiter, N.; Martorana, A.; Sharaf, A.; Carimi, F. Molecular Analysis of the Official Algerian Olive Collection Highlighted a Hotspot of Biodiversity in the Central Mediterranean Basin. Genes 2020, 11, 303. [Google Scholar] [CrossRef] [Green Version]
- Alba, V.; Sabetta, W.; Blanco, A.; Pasqualone, A.; Montemurro, C. Microsatellite markers to identify specific alleles in DNA extracted from monovarietal virgin olive oils. Eur. Food Res. Technol. 2009, 229, 375–382. [Google Scholar] [CrossRef]
- Chedid, E.; Rizou, M.; Kalaitzis, P. Application of high resolution melting combined with DNA-based markers for quantitative analysis of olive oil authenticity and adulteration. Food Chem. X 2020, 6, 100082. [Google Scholar] [CrossRef]
- Pasqualone, A.; di Rienzo, V.; Miazzi, M.M.; Fanelli, V.; Caponio, F.; Montemurro, C. High resolution melting analysis of DNA microsatellites in olive pastes and virgin olive oils obtained by talc addition. Eur. J. Lipid Sci. Technol. 2015, 117, 2044–2048. [Google Scholar] [CrossRef]
- Xanthopoulou, A.; Ganopoulos, I.; Bosmali, I.; Tsaftaris, A.; Madesis, P. DNA fingerprinting as a novel tool for olive and olive oil authentication, traceability, and detection of functional compounds. In Olives and Olive Oil as Functional Foods: Bioactivity, Chemistry and Processing; Shahidi, F., Kiritsakis, A., Eds.; Wiley: New York, NY, USA, 2017; pp. 587–601. [Google Scholar] [CrossRef]
- Batrinou, A.; Strati, I.F.; Houhoula, D.; Tsaknis, J.; Sinanoglou, V.J. Authentication of olive oil based on DNA analysis. Grasas y Aceites 2020, 71, e366. [Google Scholar] [CrossRef]
- Duran, C.; Appleby, N.; Edwards, D.; Batley, J. Molecular genetic markers: Discovery, applications, data storage and visualisation. Curr. Bioinform. 2009, 4, 16–27. [Google Scholar] [CrossRef]
- Yang, J.; Dai, P.; Zhou, T.; Huang, Z.; Feng, L.; Su, H.; Liu, Z.; Zhao, G. Genetic diversity and structure of wintersweet (Chimonanthus praecox) revealed by EST-SSR markers. Sci. Hortic. 2013, 150, 1–10. [Google Scholar] [CrossRef]
- Belaj, A.; del Carmen Dominguez-García, M.; Atienza, S.G.; Urdíroz, N.M.; De la Rosa, R.; Satovic, Z.; Martín, A.; Kilian, A.; Trujillo, I.; Valpuesta, V.; et al. Developing a core collection of olive (Olea europaea L.) based on molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree Genet. Genomes 2012, 8, 365–378. [Google Scholar] [CrossRef]
- Domínguez-García, M.C.; Belaj, A.; De la Rosa, R.; Satovic, Z.; Heller-Uszynska, K.; Kilian, A.; Martín, A.; Atienza, S.G. Development of DArT markers in olive (Olea europaea L.) and usefulness in variability studies and genome mapping. Sci. Hortic. 2012, 136, 50–60. [Google Scholar] [CrossRef]
- Ayed, R.; Ennouri, K.; Hassen, H.; Rebai, A. Molecular phylogeny to specify Zalmati and Chemlali Tataouine Tunisian olive cultivars. J. New Sci. Agric. Biotechnol. 2015, 18, 689–694. [Google Scholar]
- Biton, I.; Doron-Faigenboim, A.; Jamwal, M.; Mani, Y.; Eshed, R.; Rosen, A.; Sherman, A.; Ophir, R.; Lavee, S.; Avidan, B.; et al. Development of a large set of SNP markers for assessing phylogenetic relationships between the olive cultivars composing the Israeli olive germplasm collection. Mol. Breed. 2015, 35, 107. [Google Scholar] [CrossRef]
- İpek, A.; Yılmaz, K.; Sıkıcı, P.; Tangu, N.A.; Öz, A.T.; Bayraktar, M.; İpek, M.; Gülen, H. SNP discovery by GBS in olive and the construction of a high-density genetic linkage map. Biochem. Genet. 2016, 54, 313–325. [Google Scholar] [CrossRef]
- Belaj, A.; De La Rosa, R.; Lorite, I.J.; Mariotti, R.; Cultrera, N.G.; Beuzón, C.R.; González-Plaza, J.J.; Muñoz-Mérida, A.; Trelles, O.; Baldoni, L. Usefulness of a new large set of high throughput EST-SNP markers as a tool for olive germplasm collection management. Front. Plant Sci. 2018, 9, 1320. [Google Scholar] [CrossRef] [Green Version]
- Kaya, H.B.; Akdemir, D.; Lozano, R.; Cetin, O.; Kaya, H.S.; Sahin, M.; Smith, J.L.; Tanyolac, B.; Jannink, J.L. Genome wide association study of 5 agronomic traits in olive (Olea europaea L.). Sci. Rep. 2019, 9, 18764. [Google Scholar] [CrossRef] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Rowe, H.C.; Renaut, S.; Guggisberg, A. RAD in the realm of next-generation sequencing technologies. Mol. Ecol. 2011, 20, 3499–3502. [Google Scholar] [CrossRef] [Green Version]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef]
- Marchese, A.; Marra, F.P.; Caruso, T.; Mhelembe, K.; Costa, F.; Fretto, S.; Sargent, D.J. The first high-density sequence characterized SNP-based linkage map of olive (‘Olea europaea’ L. subsp. ’europaea’) developed using genotyping by sequencing. Aust. J. Crop Sci. 2016, 10, 857–863. [Google Scholar] [CrossRef]
- Taranto, F.; D’Agostino, N.; Pavan, S.; Fanelli, V.; di Rienzo, V.; Sabetta, W.; Miazzi, M.M.; Zelasco, S.; Perri, E.; Montemurro, C. Single nucleotide polymorphism (SNP) diversity in an olive germplasm collection. Acta Hortic. 2018, 1199, 27–32. [Google Scholar] [CrossRef]
- Zhu, S.; Niu, E.; Shi, A.; Mou, B. Genetic diversity analysis of olive germplasm (Olea europaea L.) with genotyping-by-sequencing technology. Front. Genet. 2019, 10, 755. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, R.; Fornasiero, A.; Mousavi, S.; Cultrera, N.G.M.; Brizioli, F.; Pandolfi, S.; Passeri, V.; Rossi, M.; Magris, G.; Scalabrin, S.; et al. Genetic mapping of the incompatibility locus in olive and development of a linked Sequence-Tagged Site marker. Front. Plant. Sci. 2020, 10, 1760. [Google Scholar] [CrossRef]
- Ben Ayed, R.; Rebai, A. Tunisian Table Olive Oil Traceability and Quality Using SNP Genotyping and Bioinformatics Tools. BioMed Res. Int. 2019, 2019, 8291341. [Google Scholar] [CrossRef]
- Imadi, S.R.; Kazi, A.G.; Ahanger, M.A.; Gucel, S.; Ahmad, P. Plant transcriptomics and responses to environmental stress: An overview. J. Genet. 2015, 94, 525–537. [Google Scholar] [CrossRef]
- Sabetta, W.; Mascio, I.; Squeo, G.; Gadaleta, S.; Flamminii, F.; Conte, P.; Di Mattia, C.D.; Piga, A.; Caponio, F.; Montemurro, C. Bioactive potential of minor italian olive genotypes from Apulia, Sardinia and Abruzzo. Foods 2021, 10, 1371. [Google Scholar] [CrossRef]
- Agarwal, P.; Parida, S.K.; Mahto, A.; Das, S.; Mathew, I.E.; Malik, N.; Tyagi, A.K. Expanding frontiers in plant transcriptomics in aid of functional genomics and molecular breeding. Biotechnol. J. 2014, 9, 1480–1492. [Google Scholar] [CrossRef]
- Alagna, F.; Mariotti, R.; Panara, F.; Caporali, S.; Urbani, S.; Veneziani, G.; Esposto, S.; Taticchi, A.; Rosati, A.; Rao, R.; et al. Olive phenolic compounds: Metabolic and transcriptional profiling during fruit development. BMC Plant.Biol. 2012, 12, 162. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Mérida, A.; González-Plaza, J.J.; Cañada, A.; Blanco, A.M.; García-López, M.d.C.; Rodríguez, J.M.; Pedrola, L.; Sicardo, M.D.; Hernández, M.L.; De la Rosa, R.; et al. De novo assembly and functional annotation of the olive (Olea europaea) transcriptome. DNA Res. 2013, 20, 93–108. [Google Scholar] [CrossRef]
- Parra, R.; Paredes, M.A.; Sanchez-Calle, I.M.; Gomez-Jimenez, M.C. Comparative transcriptional profiling analysis of olive ripe-fruit pericarp and abscission zone tissues shows expression differences and distinct patterns of transcriptional regulation. BMC Genom. 2013, 14, 866. [Google Scholar] [CrossRef] [Green Version]
- Carmona, R.; Zafra, A.; Seoane, P.; Castro, A.J.; Guerrero-Fernández, D.; Castillo-Castillo, T.; Medina-García, A.; Cánovas, F.M.; Aldana-Montes, J.F.; Navas-Delgado, I.; et al. ReprOlive: A database with linked data for the olive tree (Olea europaea L.) reproductive transcriptome. Front. Plant Sci. 2015, 6, 625. [Google Scholar] [CrossRef] [Green Version]
- Iaria, D.L.; Chiappetta, A.; Muzzalupo, I. A de novo transcriptomic approach to identify flavonoids and anthocyanins “switch-off” in olive (Olea europaea L.) drupes at different stages of maturation. Front. Plant Sci. 2016, 6, 1246. [Google Scholar] [CrossRef] [Green Version]
- Mariotti, R.; Belaj, A.; De La Rosa, R.; Leòn, L.; Brizioli, F.; Baldoni, L.; Mousavi, S. EST–SNP Study of Olea europaea L. Uncovers Functional Polymorphisms between Cultivated and Wild Olives. Genes 2020, 11, 916. [Google Scholar] [CrossRef]
- Olofsson, J.K.; Cantera, I.; Van de Paer, C.; Hong-Wa, C.; Zedane, L.; Dunning, L.T.; Alberti, A.; Christin, P.A.; Besnard, G. Phylogenomics using low-depth whole genome sequencing: A case study with the olive tribe. Mol. Ecol. Resour. 2019, 19, 877–892. [Google Scholar] [CrossRef]
- Kyriakopoulou, C.I.; Kalogianni, D.P. Genetic identification of the wild form of olive (Olea europaea var. sylvestris) using allele-specific real-time PCR. Foods 2020, 9, 467. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Ruiz, J.; Ramírez-Tejero, J.A.; Fernández-Pozo, N.; de la O Leyva-Pérez, M.; Yan, H.; de la Rosa, R.; Belaj, A.; Montes, E.; Rodríguez-Ariza, M.O.; Navarro, F.; et al. Transposon activation is a major driver in the genome evolution of cultivated olive trees (Olea europaea L.). Plant Genome 2020, 13, e20010. [Google Scholar] [CrossRef] [Green Version]
- Mutz, K.; Heilkenbrinker, A.; Lönne, M.; Walter, J.; Stahl, F. Transcriptome analysis using next-generation sequencing. Curr. Opin. Biotechnol. 2013, 24, 22–30. [Google Scholar] [CrossRef]
- Metzker, M.L. Sequencing technologies—The next generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef] [Green Version]
- Zafra, A.; Carmona, R.; Traverso, J.A.; Hancock, J.T.; Goldman, M.H.; Claros, M.G.; Simon, H.J.; Alche, J.D. Identification and functional annotation of genes differentially expressed in the reproductive tissues of the olive tree (Olea europaea L.) through the generation of subtractive libraries. Front. Plant Sci. 2017, 8, 1576. [Google Scholar] [CrossRef] [Green Version]
- Bruno, L.; Picardi, E.; Pacenza, M.; Chiappetta, A.; Muto, A.; Gagliardi, O.; Muzzalupo, I.; Pesole, G.; Bitonti, M.B. Changes in gene expression and metabolic profile of drupes of Olea europaea L. cv Carolea in relation to maturation stage and cultivation area. BMC Plant Biol. 2019, 19, 428. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Tejero, J.A.; Jiménez-Ruiz, J.; Leyva-Pérez, M.d.l.O.; Barroso, J.B.; Luque, F. Gene Expression Pattern in Olive Tree Organs (Olea europaea L.). Genes 2020, 11, 544. [Google Scholar] [CrossRef]
- Leyva-Perez, M.d.l.O.; Valverde-Corredor, A.; Valderrama, R.; Jiménez-Ruiz, J.; Muñoz-Merida, A.; Trelles, O.; Barroso, J.B.; Mercado-Blanco, J.; Luque, F. Early and delayed long-term transcriptional changes and short-term transient responses during cold acclimation in olive leaves. DNA Res. 2015, 22, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, S.; Regni, L.; Bocchini, M.; Mariotti, R.; Cultrera, N.G.; Mancuso, S.; Googlani, J.; Chakerolhosseini, M.R.; Guerrero, C.; Albertini, E.; et al. Physiological, epigenetic and genetic regulation in some olive cultivars under salt stress. Sci. Rep. 2019, 9, 1093. [Google Scholar] [CrossRef] [Green Version]
- Moretti, S.; Francini, A.; Hernández, M.L.; Martínez-Rivas, J.M.; Sebastiani, L. Effect of saline irrigation on physiological traits, fatty acid composition and desaturase genes expression in olive fruit mesocarp. Plant Physiol. Biochem. 2019, 141, 423–430. [Google Scholar] [CrossRef]
- Hernández, M.L.; Velázquez-Palmero, D.; Sicardo, M.D.; Fernández, J.E.; Diaz-Espejo, A.; Martínez-Rivas, J.M. Effect of a regulated deficit irrigation strategy in a hedgerow ‘Arbequina’ olive orchard on the mesocarp fatty acid composition and desaturase gene expression with respect to olive oil quality. Agric. Water Manag. 2018, 204, 100–106. [Google Scholar] [CrossRef]
- Gros-Balthazard, M.; Besnard, G.; Sarah, G.; Holtz, Y.; Leclercq, J.; Santoni, S.; Wegmann, D.; Glémin, S.; Khadari, B. Evolutionary transcriptomics reveals the origins of olives and the genomic changes associated with their domestication. Plant J. 2019, 100, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Grasso, F.; Coppola, M.; Carbone, F.; Baldoni, L.; Alagna, F.; Perrotta, G.; Pérez-Pulido, A.J.; Garonna, A.; Facella, P.; Daddiego, L.; et al. The transcriptional response to the olive fruit fly (Bactrocera oleae) reveals extended differences between tolerant and susceptible olive (Olea europaea L.) varieties. PLoS ONE 2017, 12, e0183050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyva-Pérez, M.d.l.O.; Jiménez-Ruiz, J.; Gómez-Lama Cabanás, C.; Valverde-Corredor, A.; Barroso, J.B.; Luque, F.; Mercado-Blanco, J. Tolerance of olive (Olea europaea) cv Frantoio to Verticillium dahliae relies on both basal and pathogen-induced differential transcriptomic responses. New Phytol. 2018, 217, 671–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, A.; León, L.; Belaj, A.; Román, B. Nucleotide diversity analysis of candidate genes for Verticillium wilt resistance in olive. Sci. Hortic. 2020, 274, 109653. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Morelli, M.; Saponari, M.; Loconsole, G.; Chiumenti, M.; Boscia, D.; Savino, V.N.; Martelli, G.P.; Saldarelli, P. Transcriptome profiling of two olive cultivars in response to infection by the CoDiRO strain of Xylella fastidiosa subsp. pauca. BMC Genom. 2016, 17, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novelli, S.; Gismondi, A.; Di Marco, G.; Canuti, L.; Nanni, V.; Canini, A. Plant defense factors involved in Olea europaea resistance against Xylella fastidiosa infection. J. Plant Res. 2019, 132, 439–455. [Google Scholar] [CrossRef]
- Sabella, E.; Luvisi, A.; Aprile, A.; Negro, C.; Vergine, M.; Nicolì, F.; Miceli, A.; De Bellis, L. Xylella fastidiosa induces differential expression of lignification related-genes and lignin accumulation in tolerant olive trees cv. Leccino. J. Plant Physiol. 2018, 220, 60–68. [Google Scholar] [CrossRef]
- Neale, D.B.; Williams, C.G. Restriction fragment length polymorphism mapping in conifers and applications to forest genetics and tree improvement. Can. J. For. Res. 1991, 21, 545–554. [Google Scholar] [CrossRef]
- Besnard, G.; Khadari, B.; Villemur, P.; Bervillé, A. Cytoplasmic male sterility in the olive (Olea europaea L.). Theor. Appl. Genet. 2000, 100, 1018–1024. [Google Scholar] [CrossRef]
- Mariotti, R.; Cultrera, N.G.; Diez, C.M.; Baldini, L.; Rubini, A. Identification of new polymorphic regions and differentiation of cultivated olives (Olea europaea L.) through plastome sequence comparison. BMC Plant Biol. 2010, 10, 211. [Google Scholar] [CrossRef] [Green Version]
- Bronzini de Caraffa, V.; Giannettini, J.; Gambotti, C.; Maury, J. Genetic relationships between cultivated and wild olives of Corsica and Sardinia using RAPD markers. Euphytica 2002, 123, 263–271. [Google Scholar] [CrossRef]
- García-Díaz, A.; Oya, R.; Sánchez, A.; Luque, F. Effect of prolonged vegetative reproduction of olive tree cultivars (Olea europaea L.) in mitochondrial homoplasmy and heteroplasmy. Genome 2003, 46, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Intrieri, M.C.; Muleo, R.; Buiatti, M. Chloroplast DNA polymorphisms as molecular markers to identify cultivars of Olea europaea L. J. Hortic. Sci. Biotechnol. 2007, 82, 109–113. [Google Scholar] [CrossRef]
- Besnard, G.; Henry, P.; Wille, L.; Cooke, D.; Chapuis, E. On the origin of the invasive olives (Olea europaea L., Oleaceae). Heredity 2007, 99, 608–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besnard, G.; Rubio de Casas, R.; Christin, P.; Vargas, P. Phylogenetics of Olea (Oleaceae) based on plastid and nuclear ribosomal DNA sequences: Tertiary climatic shifts and lineage differentiation times. Ann. Bot. 2009, 104, 143–160. [Google Scholar] [CrossRef] [Green Version]
- Niu, E.; Jiang, C.; Wang, W.; Zhang, Y.; Zhu, S. Chloroplast Genome Variation and Evolutionary Analysis of Olea europaea L. Genes 2020, 11, 879. [Google Scholar] [CrossRef]
- Dong, W.; Sun, J.; Liu, Y.; Xu, C.; Wang, Y.; Suo, Z.; Zhou, S.; Zhang, Z.; Wen, J. Phylogenomic relationships and species identification of the olive genus Olea (Oleaceae). J. Syst. Evol. 2021. [Google Scholar] [CrossRef]
- Fanelli, V.; Mascio, I.; Miazzi, M.M.; Savoia, M.A.; De Giovanni, C.; Montemurro, C. Molecular Approaches to Agri-Food Traceability and Authentication: An Updated Review. Foods 2021, 10, 1644. [Google Scholar] [CrossRef] [PubMed]
- Ayed, R.B.; Grati-Kamoun, N.; Moreau, F.; Rebaï, A. Comparative study of microsatellite profiles of DNA from oil and leaves of two Tunisian olive cultivars. Eur. Food Res. Technol. 2009, 229, 757–762. [Google Scholar] [CrossRef]
- Uncu, A.T.; Frary, A.; Doganlar, S. Cultivar origin and admixture detection in Turkish olive oils by SNP-based CAPS assays. J. Agric. Food Chem. 2015, 63, 2284–2295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muzzalupo, I.; Perri, E. Recovery and characterisation of DNA from virgin olive oil. Eur. Food Res. Technol. 2002, 214, 528–531. [Google Scholar] [CrossRef] [Green Version]
- Consolandi, C.; Palmieri, L.; Severgnini, M.; Maestri, E.; Marmiroli, N.; Agrimonti, C.; Baldoni, L.; Donini, P.; De Bellis, G.; Castiglioni, B. A procedure for olive oil traceability and authenticity: DNA extraction, multiplex PCR and LDR-universal array analysis. Eur. Food Res. Technol. 2008, 227, 1429–1438. [Google Scholar] [CrossRef]
- Montemurro, C.; Miazzi, M.; Pasqualone, A.; Fanelli, V.; Sabetta, W.; di Rienzo, V. Traceability of PDO Olive Oil “Terra di Bari” Using High Resolution Melting. J. Chem. 2015, 2015, 496986. [Google Scholar] [CrossRef] [Green Version]
- Gomes, S.; Breia, R.; Carvalho, T.; Carnide, V.; Martins-Lopes, P. Microsatellite High-Resolution Melting (SSR-HRM) to Track Olive Genotypes: From Field to Olive Oil. J. Food Sci. 2018, 83, 2415–2423. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Gomes, S.; Barrias, S.; Fernandes, J.R.; Martins-Lopes, P. Applying high-resolution melting (HRM) technology to olive oil and wine authenticity. Food Res. Int. 2018, 103, 170–181. [Google Scholar] [CrossRef] [PubMed]


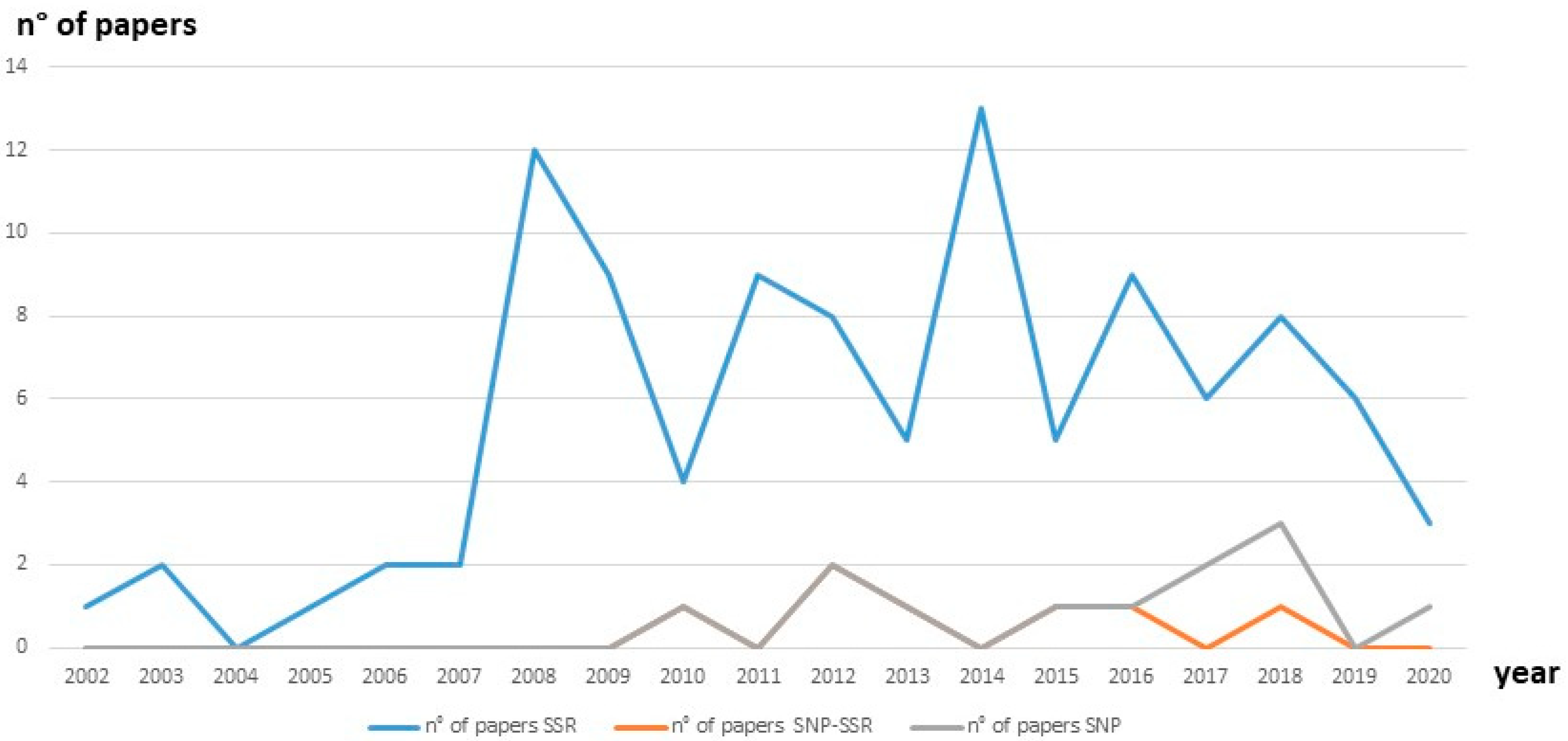{kind=link}
| Year | Descriptors | References |
|---|---|---|
| 1940 | Fruits, leaves, inflorescences and endocarp | [29] |
| 1950 | Leaves, drupes and stones | [30,31] |
| 1960 | ||
| 1970 | ||
| 1980 | Whole plant, fruiting branches, leaves, inflorescences, fruits and endocarp | [32,33] |
| 1990 | Changes to the list of UPOV descriptors and addition of agronomic characters | [39,40] |
| 2000 | Plant passport data, qualitative and quantitative morphological descriptors | [41,42] |
| [35,36] | ||
| 2010 | Morpho-geometric analysis on existing and fossil olive stones | [37] |
| Analysis and image processing of leaves, fruits and endocarp | ||
| 2020 | High resolution imagery for analysis of olive canopy traits | [38] |
| Molecular Marker | Developers | Application in Olea europea L. | References |
|---|---|---|---|
| RFLP | Williams et al., 1989 | Wild and cultivated olea variability | [49] |
| Phylogenetic studies | [50,51,52,53] | ||
| Genetic maps | [54,55,56,57,58] | ||
| Development of organelle-based markers | [53,192,193,194] | ||
| RAPD | Williams et al., 1990 | DNA fingerprinting of cultivars | [18,60,61,62] |
| Phylogenetic studies | [7,63] | ||
| AFLP | Vos et al., 1995 | DNA fingerprinting of cultivars | [65,66] |
| Phylogenetic studies | [67,68] | ||
| Construction of linkage map | [55,69,70] | ||
| QTL identification | [71,72] | ||
| SCAR and CAPS | Paran and Michelmore, 1993 | DNA fingerprinting of cultivars | [73,74] |
| Cultivar traceability in olive oil | [76,203] | ||
| ISSR | Zietkiewicz et al., 1994 | Phylogenetic studies | [79] |
| Germplasm characterization | [80,81,82,83] | ||
| SSR | Morgante and Olivieri, 1993 | Phylogenetic studies | [9,95,96,97,98,99,100,101,102,103,104] |
| Subspecies analysis | [10,21,105,106] | ||
| DNA fingerprinting of cultivars | [54,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123] | ||
| Cultivar traceability in olive oil | [142,143,144,145,201] | ||
| EST-SSR | Germplasm characterization | [22,121,127] | |
| SNP | Wang et al., 1998 | Cultivar identification | [12,107,148,153,159] |
| Genetic maps | [149,152,158] | ||
| Phylogenetic studies | [150,151,160] | ||
| Cultivar traceability in olive oil | [48] |
| Marker | Detection System | Advantages | Disadvantages | * Cost Per Sample |
|---|---|---|---|---|
| AFLP | Capillary electrophoresis | High genomic abundance High polymorphism No sequence information is required | Laboriousness of the technique Dominant markers Expensive | 50 euro |
| ISSR | Capillary electrophoresis | High genomic abundance No sequence information is required | Slightly informative Dominant markers | 10 euro |
| SSR | Capillary electrophoresis | High polymorphism | Reduced genomic abundance | 10 euro |
| High Resolution Melting | High polymorphism Low cost | Reduced genomic abundance Require optimization | 5 euro | |
| SNP | High Resolution Melting | High genomic abundance Low cost | Require optimization | 5 euro |
| CAPS | High genomic abundance Easy to perform | Expensive Low reliability | 20 euro | |
| Sequencing | High genomic abundance High reliability | Specialized personnel | 15 euro |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sion, S.; Savoia, M.A.; Gadaleta, S.; Piarulli, L.; Mascio, I.; Fanelli, V.; Montemurro, C.; Miazzi, M.M. How to Choose a Good Marker to Analyze the Olive Germplasm (Olea europaea L.) and Derived Products. Genes 2021, 12, 1474. https://doi.org/10.3390/genes12101474
Sion S, Savoia MA, Gadaleta S, Piarulli L, Mascio I, Fanelli V, Montemurro C, Miazzi MM. How to Choose a Good Marker to Analyze the Olive Germplasm (Olea europaea L.) and Derived Products. Genes. 2021; 12(10):1474. https://doi.org/10.3390/genes12101474
Chicago/Turabian StyleSion, Sara, Michele Antonio Savoia, Susanna Gadaleta, Luciana Piarulli, Isa Mascio, Valentina Fanelli, Cinzia Montemurro, and Monica Marilena Miazzi. 2021. "How to Choose a Good Marker to Analyze the Olive Germplasm (Olea europaea L.) and Derived Products" Genes 12, no. 10: 1474. https://doi.org/10.3390/genes12101474
APA StyleSion, S., Savoia, M. A., Gadaleta, S., Piarulli, L., Mascio, I., Fanelli, V., Montemurro, C., & Miazzi, M. M. (2021). How to Choose a Good Marker to Analyze the Olive Germplasm (Olea europaea L.) and Derived Products. Genes, 12(10), 1474. https://doi.org/10.3390/genes12101474