Distribution of Antibiotic Resistance Genes in the Saliva of Healthy Omnivores, Ovo-Lacto-Vegetarians, and Vegans

 ,
, 
 , ,
, ,  , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
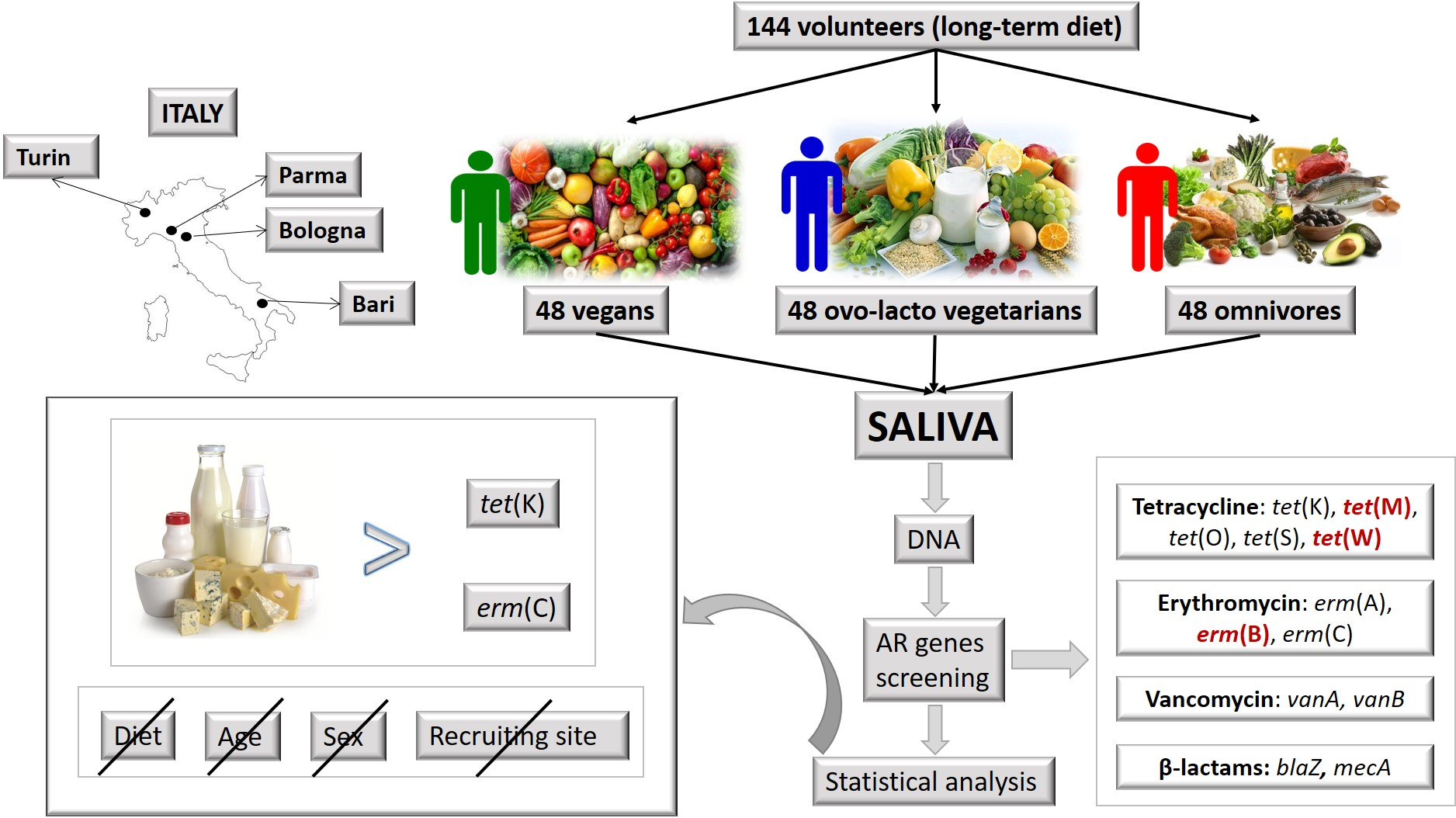
2.1. Recruitment of Healthy Volunteers with Omnivore, Ovo-Lacto-Vegetarian and Vegan Diets
2.2. Saliva Sample Collection and DNA Extraction
2.3. PCR and Nested PCR Screening of the Saliva Samples
2.4. Statistical Analysis
2.4.1. Pearson’s Chi Square (χ2) Tests
2.4.2. Principal Component Analysis (PCA)
3. Results
3.1. Cohort of Heathy Volunteers Following Long-Term Omnivore, Ovo-Lacto-Vegetarian and Vegan Diets
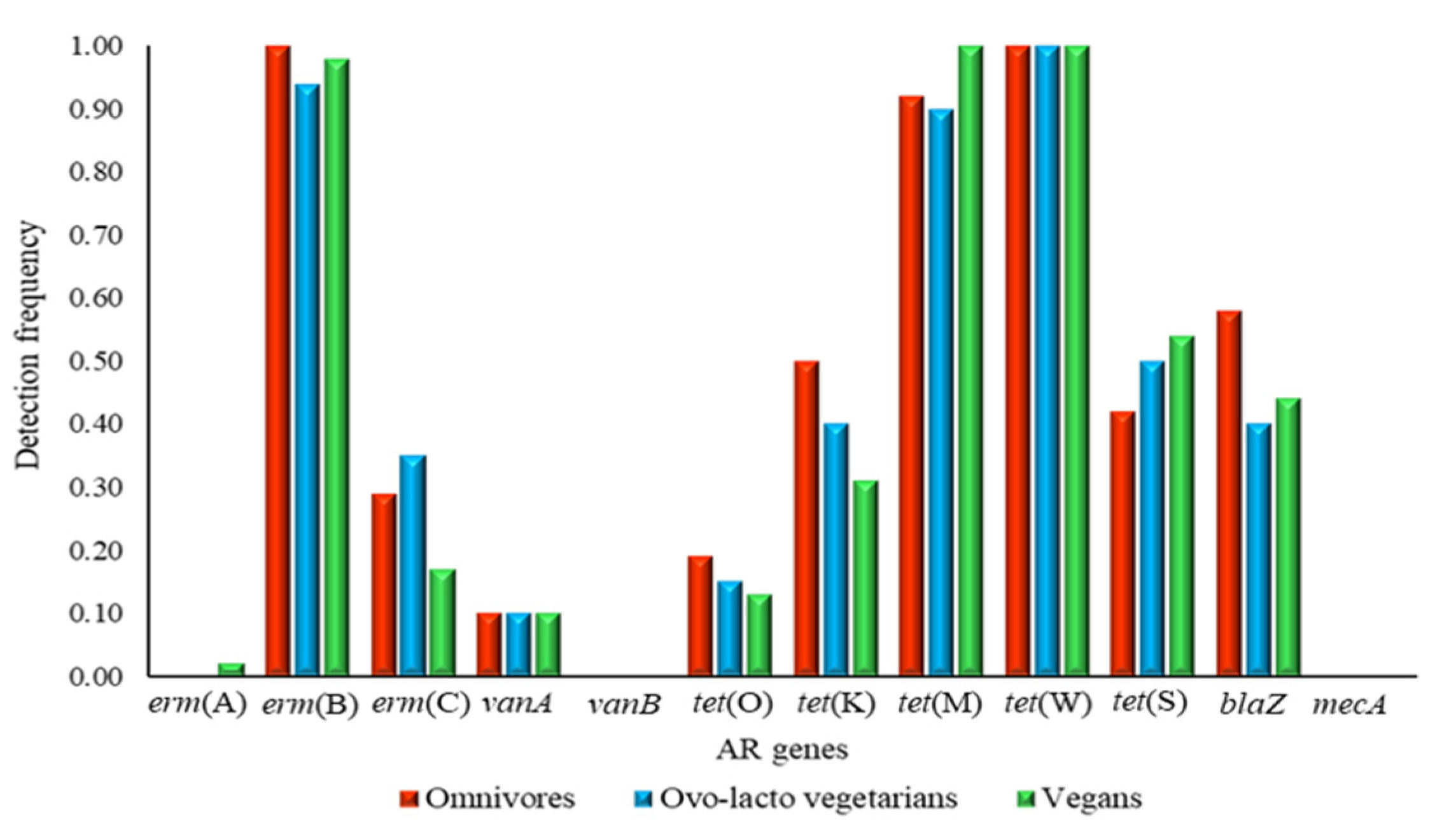
3.2. PCR and Nested PCR
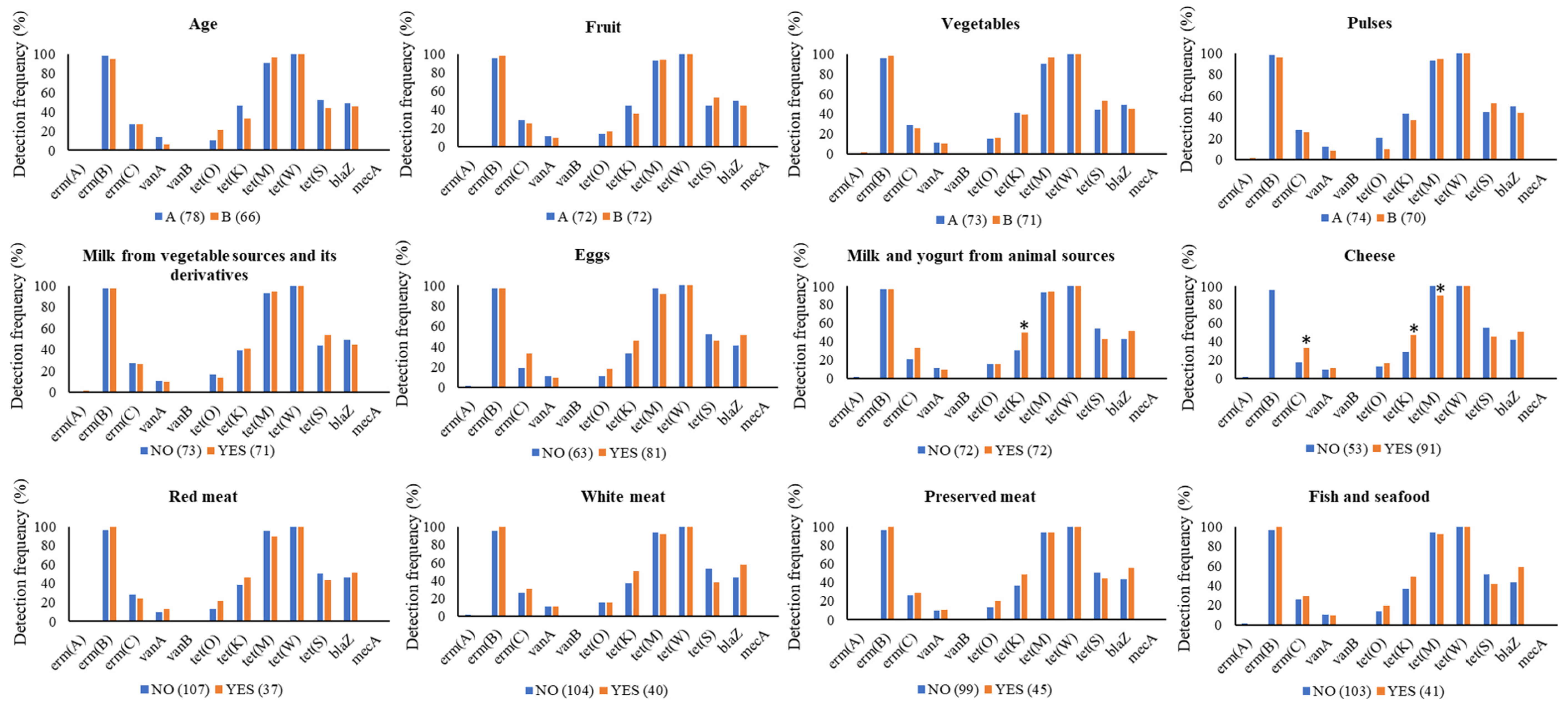
3.3. χ2 Tests
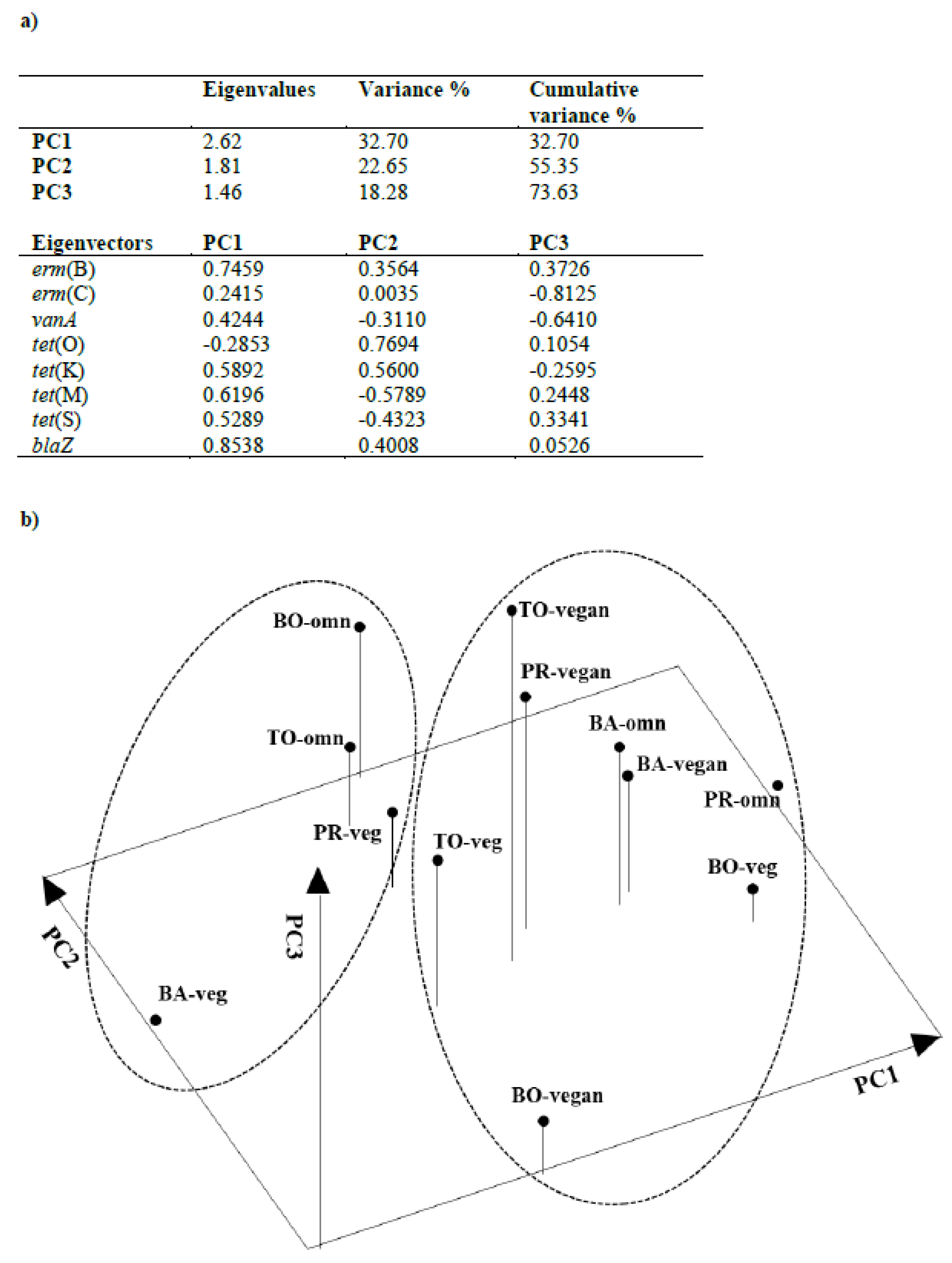
3.4. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Amerongen, A.V.; Veerman, E.C. Saliva-the defender of the oral cavity. Oral Dis. 2002, 8, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Mason, M.R.; Nagaraja, H.N.; Camerlengo, T.; Joshi, V.; Kumar, P.S. Deep sequencing identifies ethnicity-specific bacterial signatures in the oral microbiome. PLoS ONE 2013, 8, e77287. [Google Scholar] [CrossRef] [PubMed]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Willis, J.R.; Gabaldón, T. The human oral microbiome in health and disease: From sequences to ecosystems. Microorganisms 2020, 8, 308. [Google Scholar] [CrossRef]
- Moraes, L.C.; Só, M.V.; Dal Pizzol Tda, S.; Ferreira, M.B.; Montagner, F. Distribution of genes related to antimicrobial resistance in different oral environments: A systematic review. J. Endod. 2015, 41, 434–441. [Google Scholar] [CrossRef]
- Baumgartner, J.C.; Siqueira, J.F., Jr.; Xia, T.; Róças, I.N. Geographical differences in bacteria detected in endodontic infections using polymerase chain reaction. J. Endod. 2004, 30, 141–144. [Google Scholar] [CrossRef]
- Gomes, B.P.; Pinheiro, E.T.; Gadê-Neto, C.R.; Sousa, E.L.; Ferraz, C.C.; Zaia, A.A.; Teixeira, F.B.; Souza-Filho, F.J. Microbiological examination of infected dental root canals. Oral Microbiol. Immunol. 2004, 19, 71–76. [Google Scholar] [CrossRef]
- Santos, A.L.; Siqueira, J.F., Jr.; Rôças, I.N.; Jesus, E.C.; Rosado, A.S.; Tiedje, J.M. Comparing the bacterial diversity of acute and chronic dental root canal infections. PLoS ONE 2011, 6, e28088. [Google Scholar] [CrossRef]
- Piombino, P.; Genovese, A.; Esposito, S.; Moio, L.; Cutolo, P.P.; Chambery, A.; Severino, V.; Moneta, E.; Smith, D.P.; Owens, S.M.; et al. Saliva from obese individuals suppresses the release of aroma compounds from wine. PLoS ONE 2014, 9, e85611. [Google Scholar] [CrossRef]
- De Filippis, F.; Vannini, L.; La Storia, A.; Laghi, L.; Piombino, P.; Stellato, G.; Serrazanetti, D.I.; Gozzi, G.; Turroni, S.; Ferrocino, I.; et al. The same microbiota and a potentially discriminant metabolome in the saliva of omnivore, ovo-lacto-vegetarian and vegan individuals. PLoS ONE 2014, 9, e112373. [Google Scholar] [CrossRef]
- Laiola, M.; De Filippis, F.; Vitaglione, P.; Ercolini, D. A mediterranean diet intervention reduces the levels of salivary periodontopathogenic bacteria in overweight and obese subjects. Appl. Environ. Microbiol. 2020, 86, e00777-20. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.H.; Kern, T.; Bak, E.G.; Kashani, A.; Allin, K.H.; Nielsen, T.; Hansen, T.; Pedersen, O. Impact of a vegan diet on the human salivary microbiota. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [PubMed]
- Kuramitsu, H.K.; Trapa, V. Genetic exchange between oral streptococci during mixed growth. J. Gen. Microbiol. 1984, 130, 2497–2500. [Google Scholar] [CrossRef]
- Wang, H.H.; Manuzon, M.; Lehman, M.; Wan, K.; Luo, H.; Wittum, T.E.; Yousef, A.; Bakaletz, L.O. Food commensal microbes as a potentially important avenue in transmitting antibiotic resistance genes. FEMS Microbiol. Lett. 2006, 254, 226–231. [Google Scholar] [CrossRef]
- Tansirichaiya, S.; Rahman, M.A.; Antepowicz, A.; Mullany, P.; Roberts, A.P. Detection of novel integrons in the metagenome of human saliva. PLoS ONE 2016, 11, e0157605. [Google Scholar] [CrossRef]
- Mercer, D.K.; Scott, K.P.; Bruce-Johnson, W.A.; Glover, L.A.; Flint, H.J. Fate of free DNA and transformation of the oral bacterium Streptococcus gordonii DL1 by plasmid DNA in human saliva. Appl. Environ. Microbiol. 1999, 65, 6–10. [Google Scholar] [CrossRef]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; de Schaetzen, M.A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial resistance in the food chain: A review. Int. J. Environ. Res. Public Health. 2013, 10, 2643–2669. [Google Scholar] [CrossRef]
- Duran, G.M.; Marshall, D.L. Ready-to-eat shrimp as an international vehicle of antibiotic-resistant bacteria. J. Food Prot. 2005, 68, 2395–2401. [Google Scholar] [CrossRef]
- Lancaster, H.; Ready, D.; Mullany, P.; Spratt, D.; Bedi, R.; Wilson, M. Prevalence and identification of tetracycline-resistant oral bacteria in children not receiving antibiotic therapy. FEMS Microbiol. Lett. 2003, 228, 99–104. [Google Scholar] [CrossRef]
- Villedieu, A.; Diaz-Torres, M.L.; Hunt, N.; McNab, R.; Spratt, D.A.; Wilson, M.; Mullany, P. Prevalence of tetracycline resistance genes in oral bacteria. Antimicrob. Agents Chemother. 2003, 47, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Kirtzalidou, E.I.; Mitsou, E.K.; Pramateftaki, P.; Kyriacou, A. Screening fecal enterococci from Greek healthy infants for susceptibility to antimicrobial agents. Microb. Drug Resist. 2012, 18, 578–585. [Google Scholar] [CrossRef]
- Gueimonde, M.; Salminen, S.; Isolauri, E. Presence of specific antibiotic (tet) resistance genes in infant faecal microbiota. FEMS Immunol. Med. Microbiol. 2006, 48, 21–25. [Google Scholar] [CrossRef]
- Mitsou, E.K.; Kirtzalidou, E.; Pramateftaki, P.; Kyriacou, A. Antibiotic resistance in faecal microbiota of Greek healthy infants. Benef. Microbes. 2010, 1, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Aarts, H.; Margolles, A. Antibiotic resistance genes in food and gut (non-pathogenic) bacteria. Bad genes in good bugs. Front. Microbiol. 2015, 5, 754. [Google Scholar] [CrossRef]
- Rolain, J.M. Food and human gut as reservoirs of transferable antibiotic resistance encoding genes. Front. Microbiol. 2013, 4, 173. [Google Scholar] [CrossRef]
- Martinez, J.L.; Coque, T.M.; Baquero, F. What is a resistance gene? Nat. Rev. Microbiol. 2015, 13, 116–123. [Google Scholar] [CrossRef]
- Milanović, V.; Osimani, A.; Aquilanti, L.; Tavoletti, S.; Garofalo, C.; Polverigiani, S.; Litta-Mulondo, A.; Cocolin, L.; Ferrocino, I.; Di Cagno, R.; et al. Occurrence of antibiotic resistance genes in the faecal DNA of healthy omnivores, ovo-lacto vegetarians and vegans. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Carr, V.R.; Witherden, E.A.; Lee, S.; Shoaie, S.; Mullany, P.; Proctor, G.B.; Gomez-Cabrero, D.; Moyes, D.L. Abundance and diversity of resistomes differ between healthy human oral cavities and gut. Nat. Commun. 2020, 11, 693. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- Ferrocino, I.; Di Cagno, R.; De Angelis, M.; Turroni, S.; Vannini, L.; Bancalari, E.; Rantsiou, K.; Cardinali, G.; Neviani, E.; Cocolin, L. Fecal microbiota in healthy subjects following omnivore, vegetarian and vegan diets: Culturable populations and rRNA DGGE profiling. PLoS ONE 2015, 10, e0128669. [Google Scholar] [CrossRef]
- Milanović, V.; Osimani, A.; Cardinali, F.; Litta-Mulondo, A.G.; Vignaroli, C.; Citterio, B.; Mangiaterra, G.; Aquilanti, L.; Garofalo, C.; Biavasco, F.; et al. Erythromycin-resistant lactic acid bacteria in the healthy gut of vegans, ovo-lacto vegetarians and omnivores. PLoS ONE 2019, 14, e0220549. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Vignaroli, C.; Zandri, G.; Aquilanti, L.; Bordoni, D.; Osimani, A.; Clementi, F.; Biavasco, F. Direct detection of antibiotic resistance genes in specimens of chicken and pork meat. Int. J. Food. Microbiol. 2007, 113, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. The antibiotic resistome: The nexus of chemical and genetic diversity. Nat. Rev. Microbiol. 2007, 5, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Hudson, J.A.; Frewer, L.J.; Glyn, J.; Brereton, P.A.; Whittingham, M.J.; Stewart, G. The agri-food chain and antimicrobial resistance: A review. Trends Food. Sci. Tech. 2017, 69, 131–147. [Google Scholar] [CrossRef]
- Caniça, M.; Manageiro, V.; Hikmate, A.; Moran-Gilad, J.; Franz, C.M.A.P. Antibiotic resistance in foodborne bacteria. Trends Food. Sci. Tech. 2019, 84, 41–44. [Google Scholar] [CrossRef]
- Clementi, F.; Aquilanti, L. Recent investigations and updated criteria for the assessment of antibiotic resistance in food lactic acid bacteria. Anaerobe 2011, 17, 394–398. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). ECDC/EFSA/EMA second joint report on the integrated analysis of the consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food-producing animals. EFSA J. 2017, 15, 4872. [Google Scholar]
- Anderson, A.C.; Jonas, D.; Huber, I.; Karygianni, L.; Wölber, J.; Hellwig, E.; Arweiler, N.; Vach, K.; Wittmer, A.; Al-Ahmad, A. Enterococcus faecalis from food, flinical specimens, and oral sites: Prevalence of virulence factors in association with biofilm formation. Front. Microbiol. 2016, 6, 1534. [Google Scholar] [CrossRef]
- Diaz-Torres, M.L.; Villedieu, A.; Hunt, N.; McNab, R.; Spratt, D.A.; Allan, E.; Mullany, P.; Wilsonet, M. Determining the antibiotic resistance potential of the indigenous oral microbiota of humans using a metagenomic approach. FEMS Microbiol. Lett. 2006, 258, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Seville, L.A.; Patterson, A.J.; Scott, K.P.; Mullany, P.; Quail, M.A.; Parkhill, J.; Ready, D.; Wilson, M.; Spratt, D.; Roberts, A.P. Distribution of tetracycline and erythromycin resistance genes among human oral and fecal metagenomic DNA. Microb. Drug Resist. 2009, 15, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Sukumar, S.; Roberts, A.P.; Martin, F.E.; Adler, C.J. Metagenomic insights into transferable antibiotic resistance in oral bacteria. J. Dent. Res. 2016, 95, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Card, R.M.; Warburton, P.J.; MacLaren, N.; Mullany, P.; Allan, E.; Anjum, M.F. Application of microarray and functional-based screening methods for the detection of antimicrobial resistance genes in the microbiomes of healthy humans. PLoS ONE 2014, 9, e86428. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.J.; Colangeli, R.; Spigaglia, P.; Scott, K.P. Distribution of specific tetracycline and erythromycin resistance genes in environmental samples assessed by macroarray detection. Environ. Microbiol. 2007, 9, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Q. Relationship between tetracycline antibiotic susceptibility and genotype in oral cavity lactobacilli clinical isolates. Antimicrob. Resist. Infect. Control. 2019, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Guay, G.G.; Khan, S.A.; Rothstein, D.M. The tet(K) gene of plasmid pT181 of Staphylococcus aureus encodes an efflux protein that contains 14 transmembrane helices. Plasmid 1993, 30, 163–166. [Google Scholar] [CrossRef]
- Moon, K.H.; Kim, W.K.; Yoon, S.J.; Kim, J.M.; Shin, C.K.; Im, S.H. Relationship between two tetracycline resistance plasmids of Staphylococcus aureus in Korea. J. Microbiol. Biotechnol. 1996, 6, 292–294. [Google Scholar]
- Villedieu, A.; Diaz-Torres, M.L.; Roberts, A.P.; Hunt, N.; McNab, R.; Spratt, D.; Wilson, M.; Mullany, P. Genetic basis of erythromycin resistance in oral bacteria. Antimicrob. Agents Chemother. 2004, 48, 2298–2301. [Google Scholar] [CrossRef]
- EMA (European Medicines Agency). Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2017; Trends from 2010 to 2017; Ninth ESVAC report; EMA/294674/2019; European Medicines Agency: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Kim, G.Y.; Lee, C.H. Antimicrobial susceptibility and pathogenic genes of Staphylococcus aureus isolated from the oral cavity of patients with periodontitis. J. Periodontal Implant. Sci. 2015, 45, 223–228. [Google Scholar] [CrossRef]
- Zmantar, T.; Kouidhi, B.; Hentati, H.; Bakhrouf, A. Detection of disinfectant and antibiotic resistance genes in Staphylococcus aureus isolated from the oral cavity of Tunisian children. Ann. Microbiol. 2012, 62, 123–128. [Google Scholar] [CrossRef]
- Koukos, G.; Konstantinidis, A.; Tsalikis, L.; Arsenakis, M.; Slini, T.; Sakellari, D. Prevalence of beta-lactam (bla tem) and metronidazole (nim) resistance genes in the oral cavity of Greek subjects. Open Dent. J. 2016, 10, 89–98. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kühn, I.; Iversen, A.; Burman, L.G.; Olsson-Liljequist, B.; Franklin, A.; Finn, M.; Aarestrup, F.; Seyfarth, A.M.; Franklin, A.; Finn, M.; et al. Comparison of enterococcal populations in animals, humans, and the environment—A European study. Int. J. Food Microbiol. 2003, 88, 133–145. [Google Scholar] [CrossRef]
- Razavi, A.; Gmür, R.; Imfeld, T.; Zehnder, M. Recovery of Enterococcus faecalis from cheese in the oral cavity of healthy subjects. Oral Microbiol. Immunol. 2007, 22, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Takeda, I.; Stretch, C.; Barnaby, P.; Bhatnager, K.; Rankin, K.; Fu, H.; Weljie, A.; Jha, N.; Slupsky, C. Understanding the human salivary metabolome. NMR Biomed. 2009, 22, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Ruaro, A.; Andrighetto, C.; Torriani, S.; Lombardi, A. Biodiversity and characterization of indigenous coagulase-negative staphylococci isolated from raw milk and cheese of North Italy. Food Microbiol. 2013, 34, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Turchi, B.; Bertelloni, F.; Marzoli, F.; Cerri, D.; Tola, S.; Azara, E.; Longheu, C.M.; Tassi, R.; Schiavo, M.; Cilia, G.; et al. Coagulase negative staphylococci from ovine milk: Genotypic and phenotypic characterization of susceptibility to antibiotics, disinfectants and biofilm production. Small Ruminant Res. 2020, 83, 106030. [Google Scholar] [CrossRef]
- Flórez, A.B.; Alegría, Á.; Rossi, F.; Delgado, S.; Felis, G.E.; Torriani, S.; Mayo, B. Molecular identification and quantification of tetracycline and erythromycin resistance genes in Spanish and Italian retail cheeses. Biomed. Res. Int. 2014, 2014, 746859. [Google Scholar] [CrossRef]
- Devirgiliis, C.; Caravelli, A.; Coppola, D.; Barile, S.; Perozzi, G. Antibiotic resistance and microbial composition along the manufacturing process of Mozzarella di Bufala Campana. Int. J. Food Microbiol. 2008, 128, 378–384. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Gajewska, J. S. epidermidis strains from artisanal cheese made from unpasteurized milk in Poland–genetic characterization of antimicrobial resistance and virulence determinants. Int. J. Food Microbiol. 2019, 294, 55–59. [Google Scholar]
- Schlegelova, J.; Vlkova, H.; Babak, V.; Holasova, M.; Jaglic, Z.; Stosova, T.P. Sauer. Resistance to erythromycin of Staphylococcus spp. isolates from the food chain. Vet. Med. 2008, 53, 307–314. [Google Scholar] [CrossRef]
- Ytreland, K.J. Antibiotic Use and Antibiotic Resistance in Dental Practice. Master’s Thesis, UIT, The Arctic University of Norway, Tromsø, Norway, 2016. [Google Scholar]
- Martinez, J.L. Environmental pollution by antibiotics and by antibiotic resistance determinants. Environ. Pollut. 2009, 157, 2893–2902. [Google Scholar] [CrossRef] [PubMed]
- FAO/OIE/WHO (Food and Agriculture Organization of the United Nations, World Organization for Animal Health, World Health Organization). The Fao Action Plan on Antimicrobial Resistance 2016–2020: Supporting The Food and Agriculture Sectors in Implementing the Global Action Plan on Antimicrobial Resistance to Minimize the Impact of Antimicrobial Resistance; FAO: Rome, Italy, 2016. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Categories (g or mL day−1) | Diet | ||
|---|---|---|---|
| Vegans | Ovo-Lacto Vegetarians | Omnivores | |
| Fruits | 689.7 ± 983.1 | 303.7 ± 194.4 | 181.3 ± 108.0 |
| Vegetables | 520.8 ± 210.8 | 438.1 ± 189.0 | 270.1 ± 107.5 |
| Pulses | 115.2 ± 75.5 | 92.4 ± 55.3 | 16.0 ± 26.6 |
| Milk from vegetable sources and its derivatives | 135.1 ± 113.7 | 74.7 ± 89.1 | 8.5 ± 43.4 |
| Eggs | n.c. | 15.8 ± 15.4 | 15.0 ± 12.1 |
| Milk and yogurt from animal sources | n.c. | 60.2 ± 94.0 | 137.6 ± 140.5 |
| Cheese | n.c. | 49.2 ± 39.1 | 61.3 ± 32.3 |
| Red meat | n.c. | n.c. | 35.3 ± 31.9 |
| White meat | n.c. | n.c. | 39.9 ± 43.7 |
| Preserved meat | n.c. | n.c. | 46.3 ± 26.5 |
| Fish and seafood | n.c. | n.c. | 38.9 ± 31.2 |
| Diet (Number of Volunteers) | Assay | Number of Positive Samples | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| erm(A) | erm(B) | erm(C) | vanA | vanB | tet(O) | tet(M) | tet(W) | tet(S) | tet(K) | mecA | blaZ | ||
| Omnivore (48) | PCR | 0 | 31 | 5 | 0 | 0 | 0 | 44 | 46 | 1 | 11 | 0 | 15 |
| n-PCR | 0 | 17 | 9 | 5 | 0 | 9 | 0 | 2 | 19 | 13 | 0 | 13 | |
| Total | 0 | 48 | 14 | 5 | 0 | 9 | 44 | 48 | 20 | 24 | 0 | 28 | |
| Vegan (48) | PCR | 0 | 35 | 1 | 0 | 0 | 0 | 47 | 48 | 0 | 7 | 0 | 11 |
| n-PCR | 1 | 12 | 7 | 5 | 0 | 6 | 1 | n.a. | 26 | 8 | 0 | 10 | |
| Total | 1 | 47 | 8 | 5 | 0 | 6 | 48 | 48 | 26 | 15 | 0 | 21 | |
| Ovo-lacto vegetarian (48) | PCR | 0 | 23 | 0 | 0 | 0 | 0 | 43 | 45 | 0 | 8 | 0 | 13 |
| n-PCR | 0 | 22 | 17 | 5 | 0 | 7 | 0 | 3 | 24 | 11 | 0 | 6 | |
| Total | 0 | 45 | 17 | 5 | 0 | 7 | 43 | 48 | 24 | 19 | 0 | 19 | |
| Whole cohort (144) | PCR | 0 | 89 | 6 | 0 | 0 | 0 | 134 | 139 | 1 | 26 | 0 | 39 |
| n-PCR | 1 | 51 | 33 | 15 | 0 | 22 | 1 | 5 | 69 | 32 | 0 | 29 | |
| Total | 1 | 140 | 39 | 15 | 0 | 22 | 135 | 144 | 70 | 58 | 0 | 68 | |
| (A) Bari | (B) Bari | ||||||
| AR gene | Ovo-lacto vegetarians (n = 12) | Omnivores + Vegans (n = 27) | p | AR gene | Omnivores (n = 14) | Vegans (n = 13) | p |
| erm(B) | 10 (83%) | 27 (100%) | 0.026 * | erm(B) | 14 (100%) | 13 (100%) | n.a. |
| erm(C) | 4 (33%) | 8 (30%) | 0.818 | erm(C) | 4 (29%) | 4 (31%) | 0.901 |
| vanA | 1 (8%) | 3 (11%) | 0.788 | vanA | 1 (7%) | 2 (15%) | 0.493 |
| tet(O) | 3 (25%) | 3 (11%) | 0.283 | tet(O) | 1 (7%) | 2 (15%) | 0.493 |
| tet(K) | 4 (33%) | 12 (44%) | 0.512 | tet(K) | 6 (43%) | 6 (46%) | 0.863 |
| tet(M) | 10 (%) | 26 (96%) | 0.182 | tet(M) | 13 (93%) | 13 (100%) | 0.245 |
| tet(S) | 3 (25%) | 17 (63%) | 0.026 * | tet(S) | 10 (71%) | 7 (54%) | 0.343 |
| blaZ | 3 (25%) | 15 (56%) | 0.072 | blaZ | 8 (57%) | 7 (54%) | 0.863 |
| (C) Bologna | (D) Bologna | ||||||
| AR gene | Ovo-lacto vegetarians (n = 12) | Omnivores + Vegans (n = 24) | p | AR gene | Omnivores (n = 13) | Vegans (n = 11) | p |
| erm(B) | 12 (100%) | 23 (96%) | 0.364 | erm(B) | 13 (100%) | 10 (91%) | 0.204 |
| erm(C) | 7 (58%) | 5 (21%) | 0.026 * | erm(C) | 2 (15%) | 3 (27%) | 0.475 |
| vanA | 2 (17%) | 4 (17%) | 1.000 | vanA | 1 (8%) | 3 (27%) | 0.194 |
| tet(O) | 1 (8%) | 4 (17%) | 0.479 | tet(O) | 4 (31%) | 0 (0%) | 0.018 * |
| tet(K) | 6 (50%) | 8 (33%) | 0.336 | tet(K) | 7 (54%) | 1 (9%) | 0.015 * |
| tet(M) | 12 (100%) | 22 (92%) | 0.195 | tet(M) | 11 (85%) | 11 (100%) | 0.107 |
| tet(S) | 10 (83%) | 10 (42%) | 0.014 * | tet(S) | 5 (38%) | 5 (45%) | 0.729 |
| blaZ | 6 (50%) | 10 (42%) | 0.636 | blaZ | 7 (54%) | 3 (27%) | 0.184 |
| (E) Parma | (F) Parma | ||||||
| AR gene | Ovo-lacto vegetarians + Vegans (n = 24) | Omnivores (n = 9) | p | AR gene | Vegans (n = 12) | Ovo-lacto vegetarians (n = 12) | p |
| erm(B) | 24 (100%) | 9 (100%) | n.a. | erm(B) | 12 (%) | 12 | n.a. |
| erm(C) | 5 (21%) | 3 (33%) | 0.465 | erm(C) | 1 (8%) | 4 (33%) | 0.121 |
| vanA | 2 (8%) | 3 (33%) | 0.092 | vanA | 0 (0%) | 2 (17%) | 0.086 |
| tet(O) | 4 (17%) | 1 (11%) | 0.684 | tet(O) | 1 (8%) | 3 (25%) | 0.264 |
| tet(K) | 10 (42%) | 6 (67%) | 0.198 | tet(K) | 6 (50%) | 4 (33%) | 0.406 |
| tet(M) | 22 (92%) | 9 (100%) | 0.250 | tet(M) | 12 (100%) | 10 (83%) | 0.086 |
| tet(S) | 11 (46%) | 4 (44%) | 0.943 | tet(S) | 6 (50%) | 5 (42%) | 0.682 |
| blaZ | 10 (42%) | 7 (78%) | 0.058 | blaZ | 5 (42%) | 5 (42%) | 1.000 |
| (G) Turin | (H) Turin | ||||||
| AR gene | Ovo-lacto vegetarians +Vegans (n = 24) | Omnivores (n = 12) | p | AR gene | Vegans (n = 12) | Ovo-lacto vegetarians (n = 12) | p |
| erm(B) | 23 (96%) | 12 (100%) | 0.364 | erm(B) | 12 (100%) | 11 (92%) | 0.232 |
| erm(C) | 2 (8%) | 5 (42%) | 0.020 * | erm(C) | 0 (0%) | 2 (17%) | 0.086 |
| vanA | 0 (0%) | 0 (0%) | n.a. | vanA | 0 (0%) | 0 (0%) | n.a. |
| tet(O) | 3 (12%) | 3 (25%) | 0.354 | tet(O) | 3 (25%) | 0 (0%) | 0.032 * |
| tet(K) | 7 (29%) | 5 (42%) | 0.457 | tet(K) | 2 (17%) | 5 (42%) | 0.173 |
| tet(M) | 23 (96%) | 11 (92%) | 0.617 | tet(M) | 12 (100%) | 11 (92%) | 0.232 |
| tet(S) | 14 (58%) | 1 (8%) | 0.002 * | tet(S) | 5 (42%) | 9 (75%) | 0.094 |
| blaZ | 11 (46%) | 6 (50%) | 0.813 | blaZ | 5 (42%) | 6 (50%) | 0.682 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milanović, V.; Aquilanti, L.; Tavoletti, S.; Garofalo, C.; Osimani, A.; De Filippis, F.; Ercolini, D.; Ferrocino, I.; Cagno, R.D.; Turroni, S.; et al. Distribution of Antibiotic Resistance Genes in the Saliva of Healthy Omnivores, Ovo-Lacto-Vegetarians, and Vegans. Genes 2020, 11, 1088. https://doi.org/10.3390/genes11091088
Milanović V, Aquilanti L, Tavoletti S, Garofalo C, Osimani A, De Filippis F, Ercolini D, Ferrocino I, Cagno RD, Turroni S, et al. Distribution of Antibiotic Resistance Genes in the Saliva of Healthy Omnivores, Ovo-Lacto-Vegetarians, and Vegans. Genes. 2020; 11(9):1088. https://doi.org/10.3390/genes11091088
Chicago/Turabian StyleMilanović, Vesna, Lucia Aquilanti, Stefano Tavoletti, Cristiana Garofalo, Andrea Osimani, Francesca De Filippis, Danilo Ercolini, Ilario Ferrocino, Raffaella Di Cagno, Silvia Turroni, and et al. 2020. "Distribution of Antibiotic Resistance Genes in the Saliva of Healthy Omnivores, Ovo-Lacto-Vegetarians, and Vegans" Genes 11, no. 9: 1088. https://doi.org/10.3390/genes11091088
APA StyleMilanović, V., Aquilanti, L., Tavoletti, S., Garofalo, C., Osimani, A., De Filippis, F., Ercolini, D., Ferrocino, I., Cagno, R. D., Turroni, S., Lazzi, C., Pellegrini, N., & Clementi, F. (2020). Distribution of Antibiotic Resistance Genes in the Saliva of Healthy Omnivores, Ovo-Lacto-Vegetarians, and Vegans. Genes, 11(9), 1088. https://doi.org/10.3390/genes11091088