Liver Transcriptome Responses to Heat Stress and Newcastle Disease Virus Infection in Genetically Distinct Chicken Inbred Lines

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
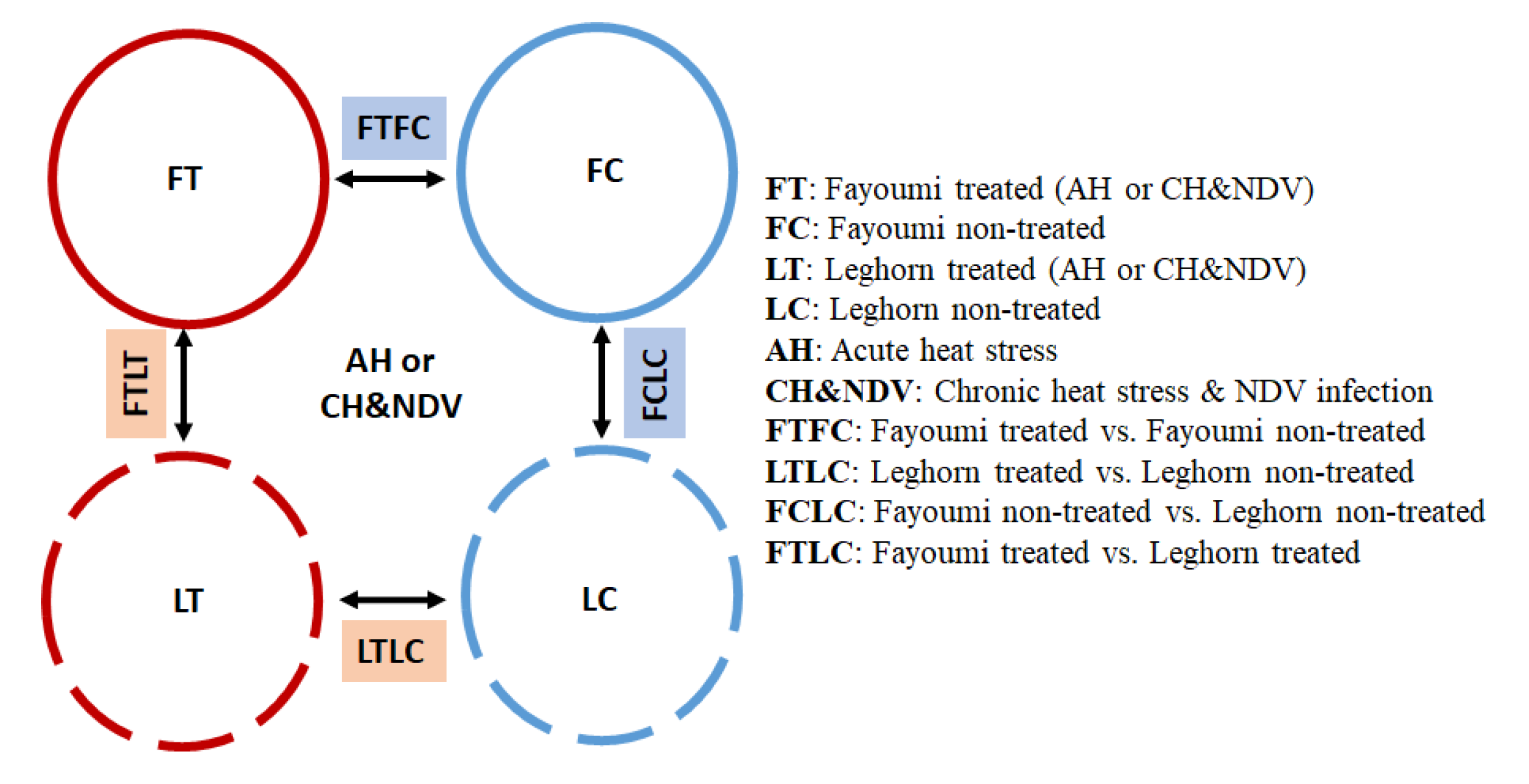
2.1. Experimental Populations and Design
2.2. Sample Collection and RNA Isolation
2.3. RNA-seq Library Preparation
2.4. RNA-seq Analysis
2.5. Gene Ontology
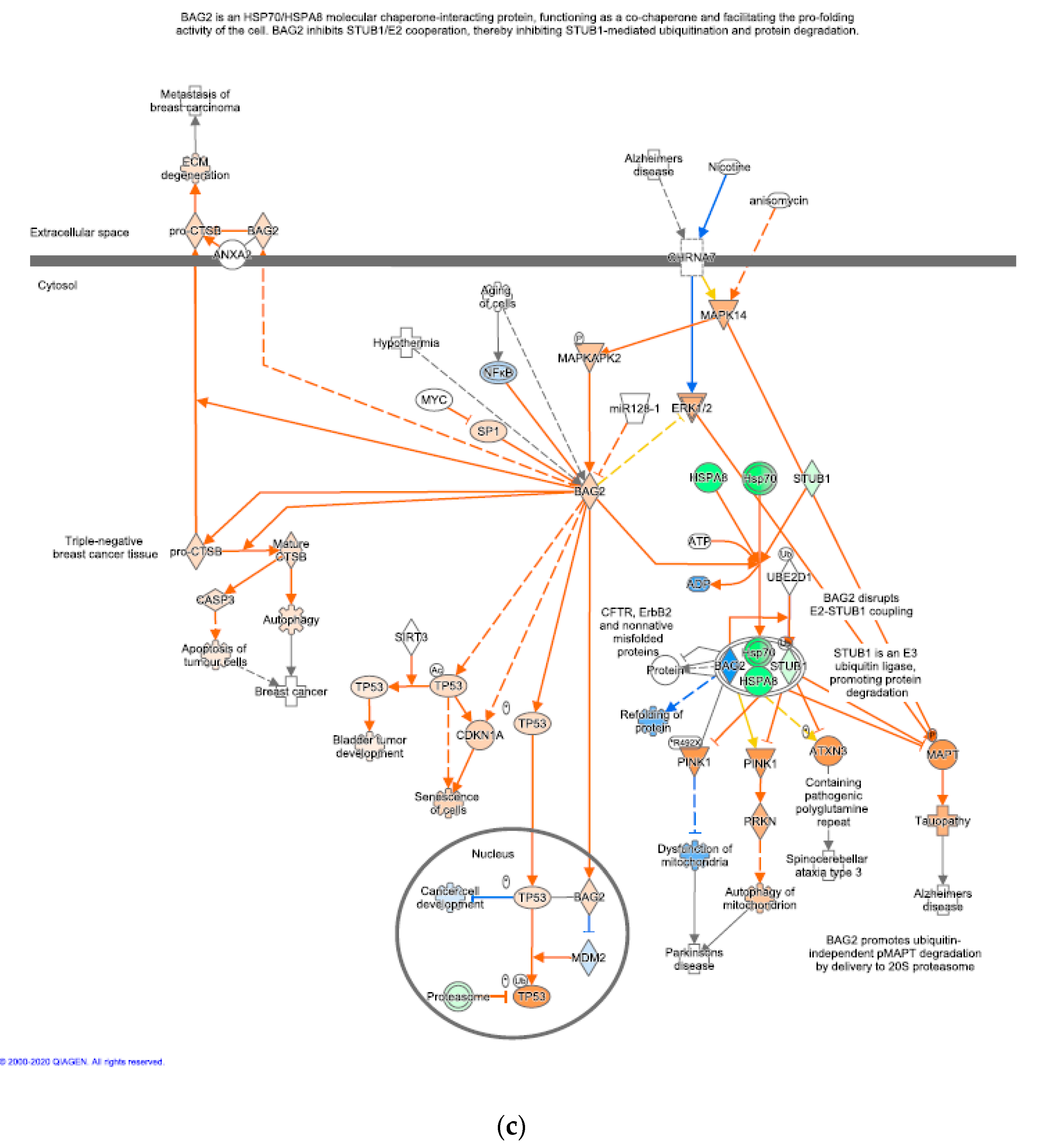
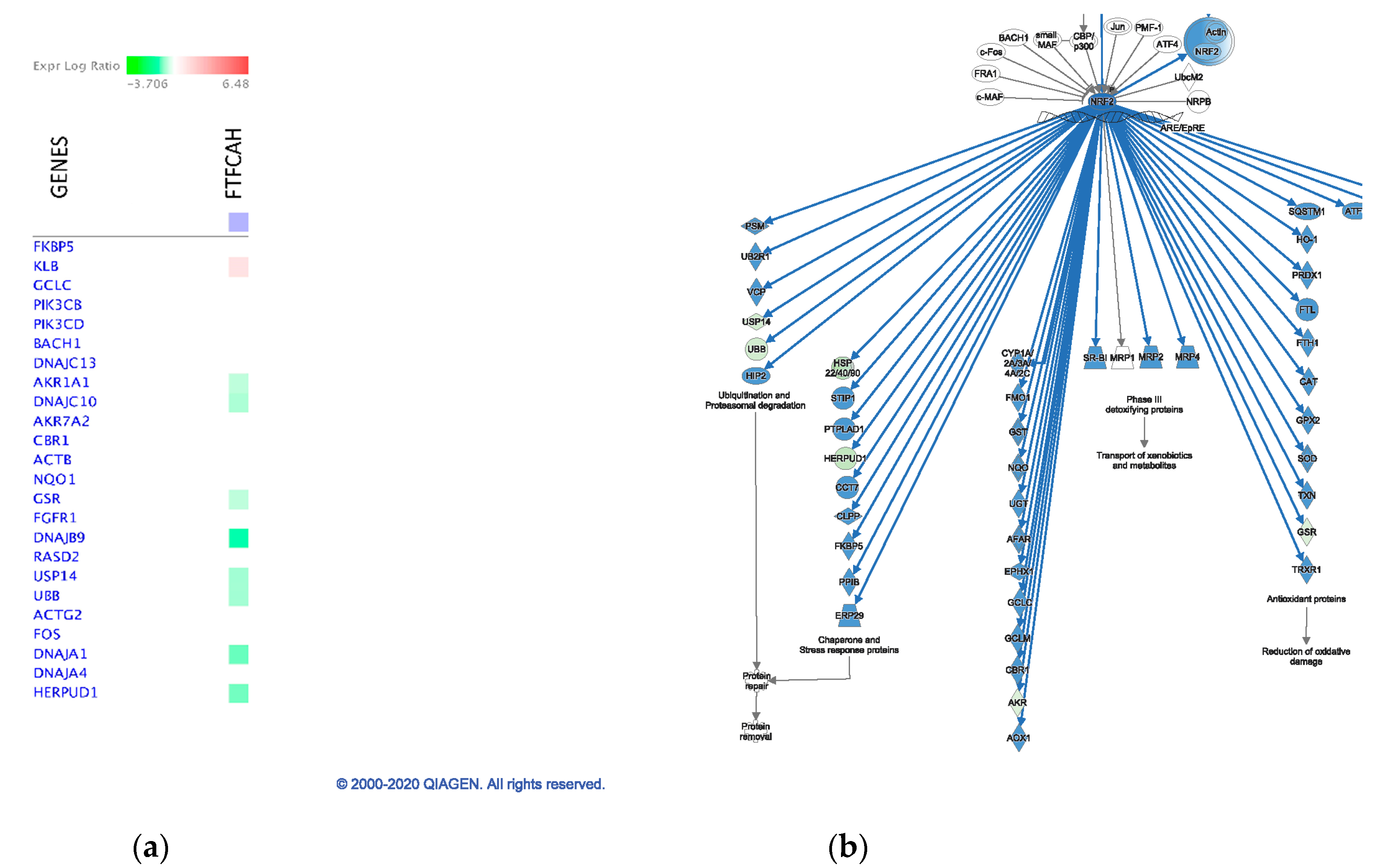
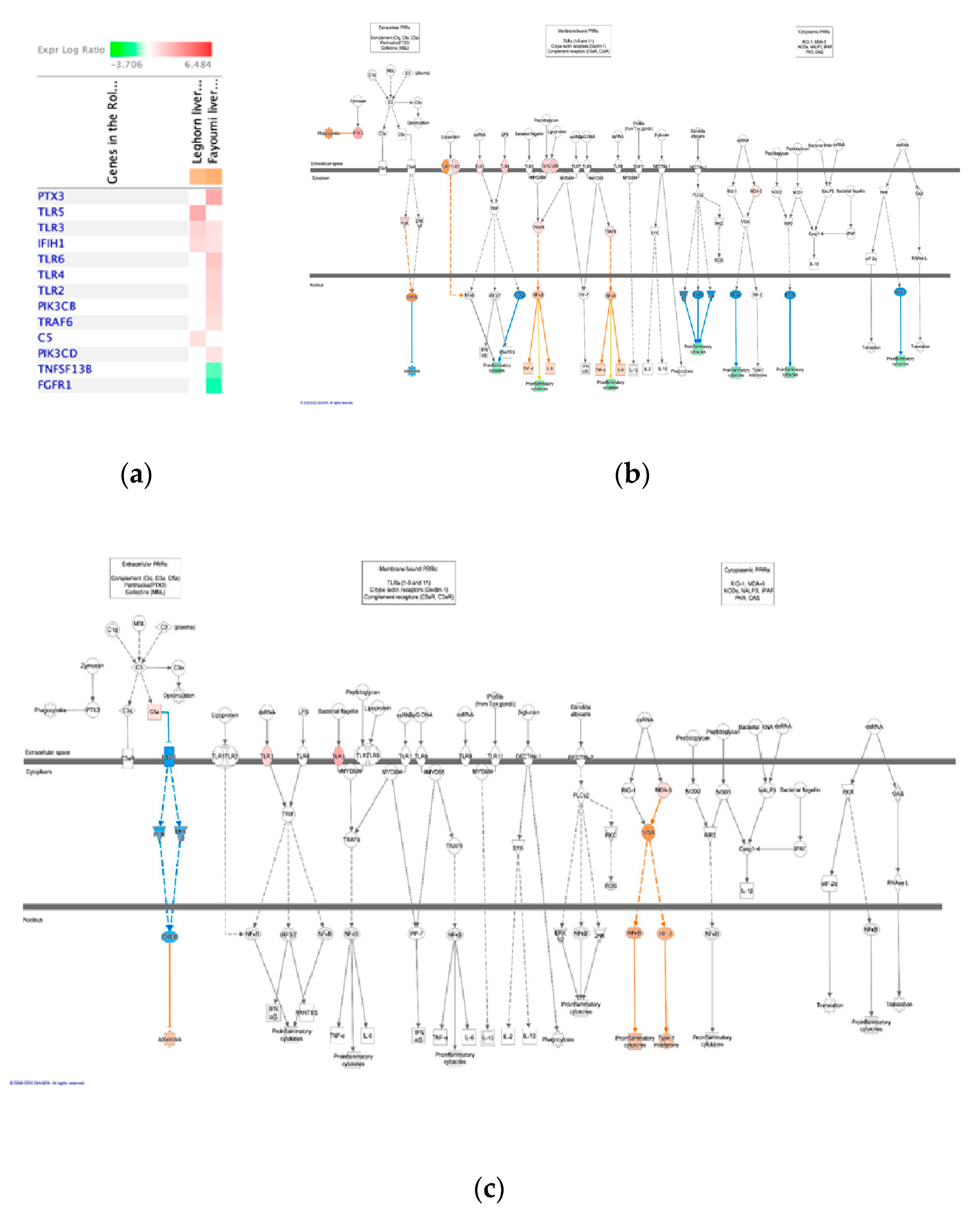
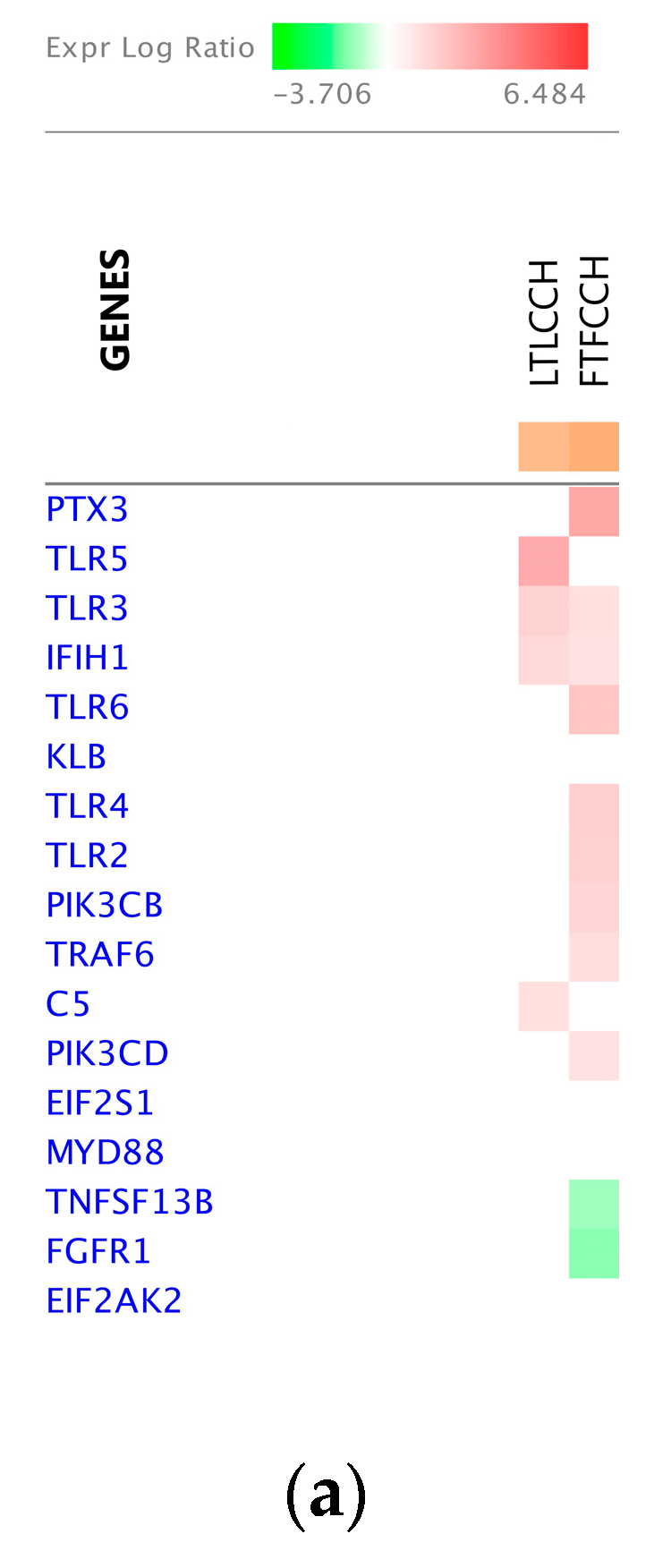
2.6. Pathway Analysis
2.7. Gene Co-Expression Network Analysis
3. Results
3.1. Effects of Heat Stress and NDV Infection on Gene Expression
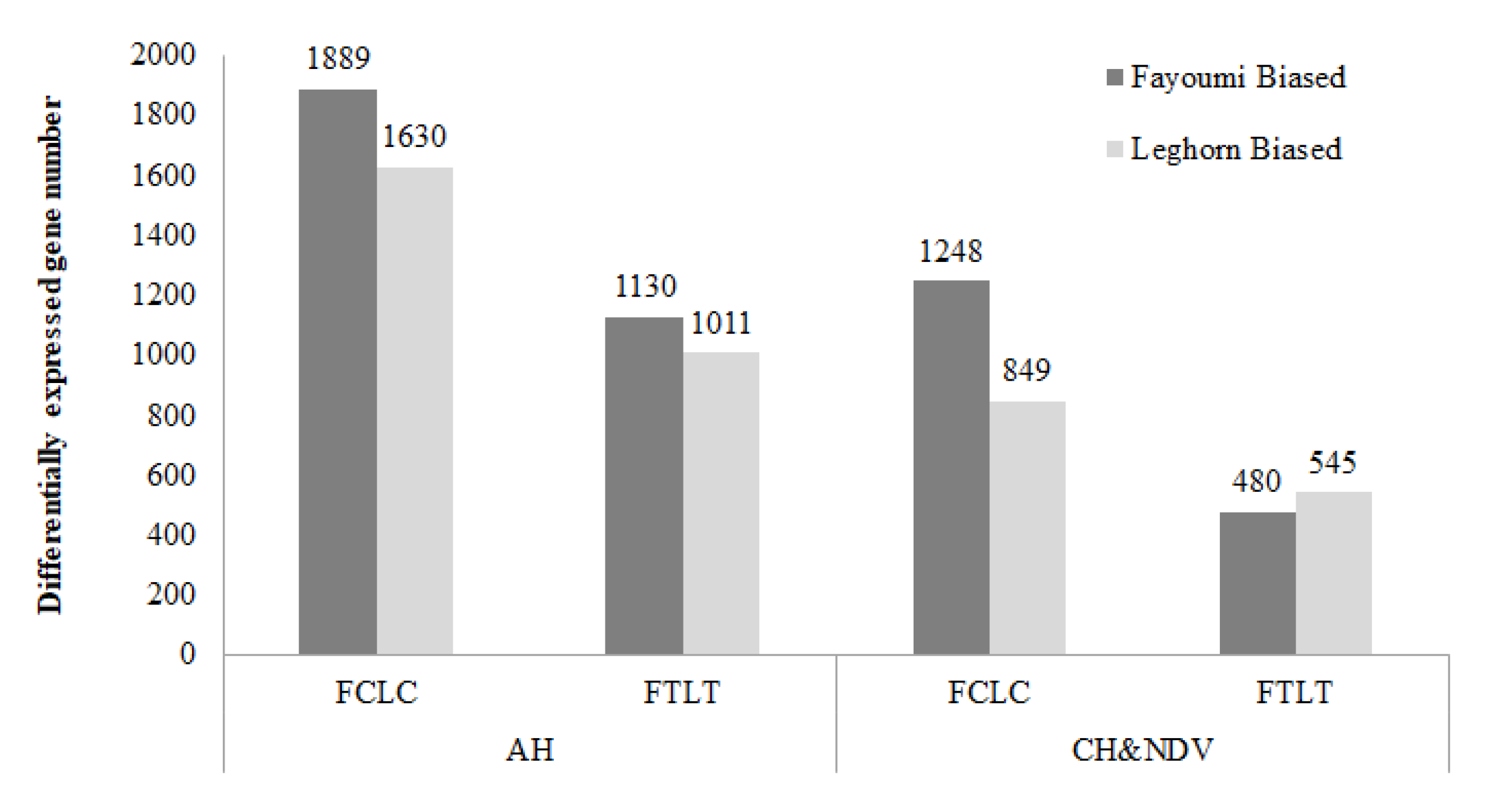
3.1.1. Between Genetic Lines
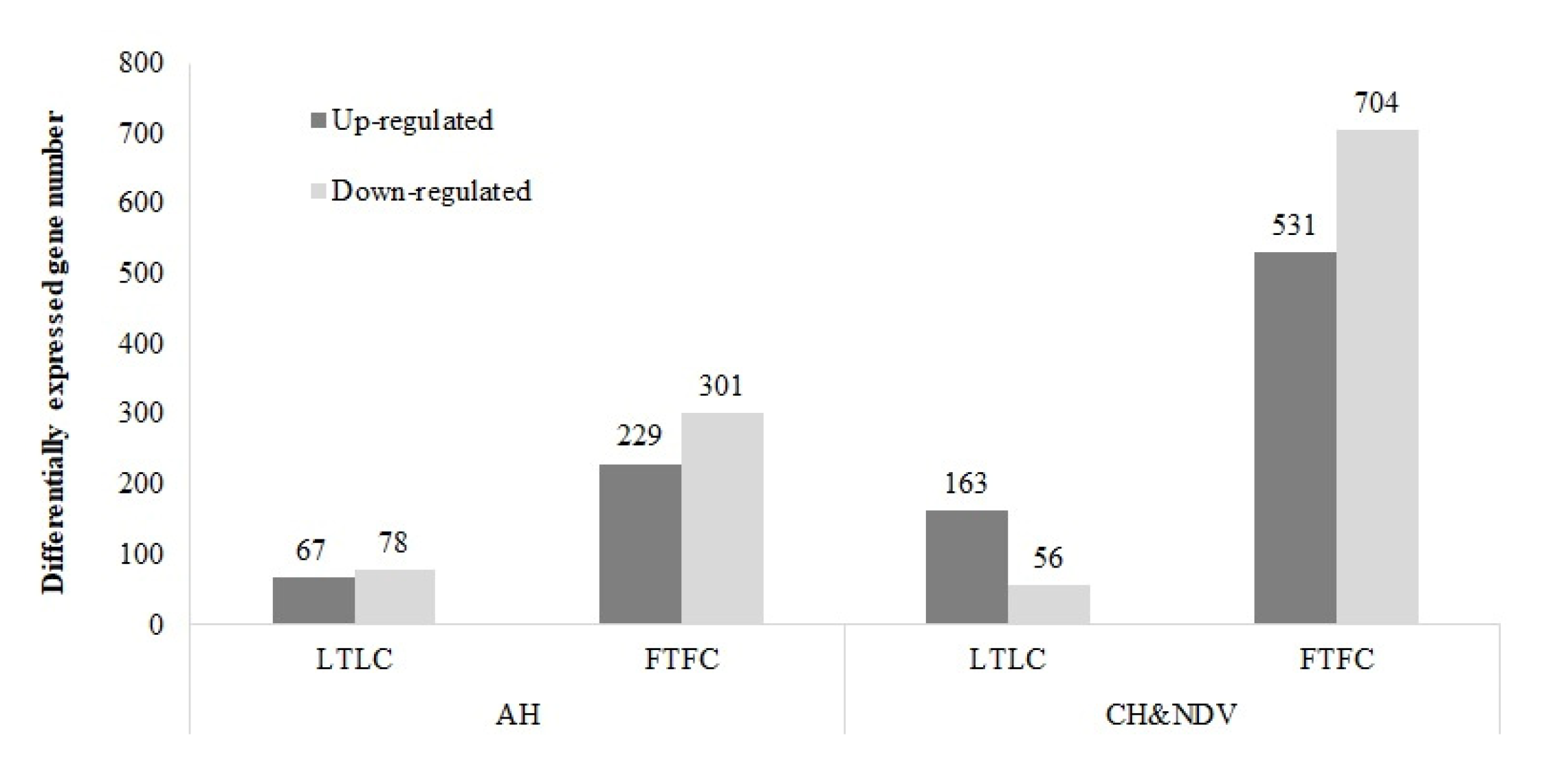
3.1.2. Within Genetic Lines
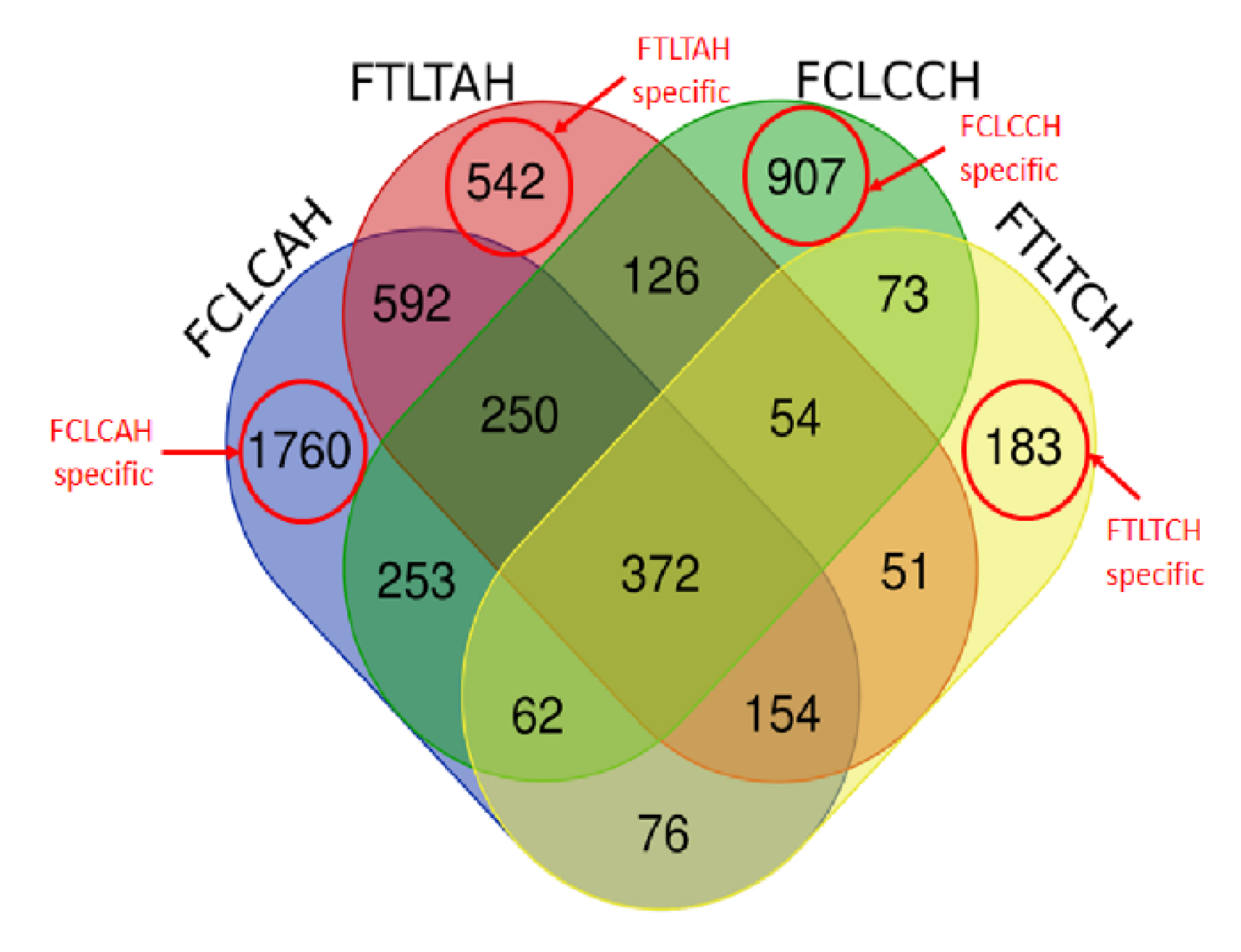
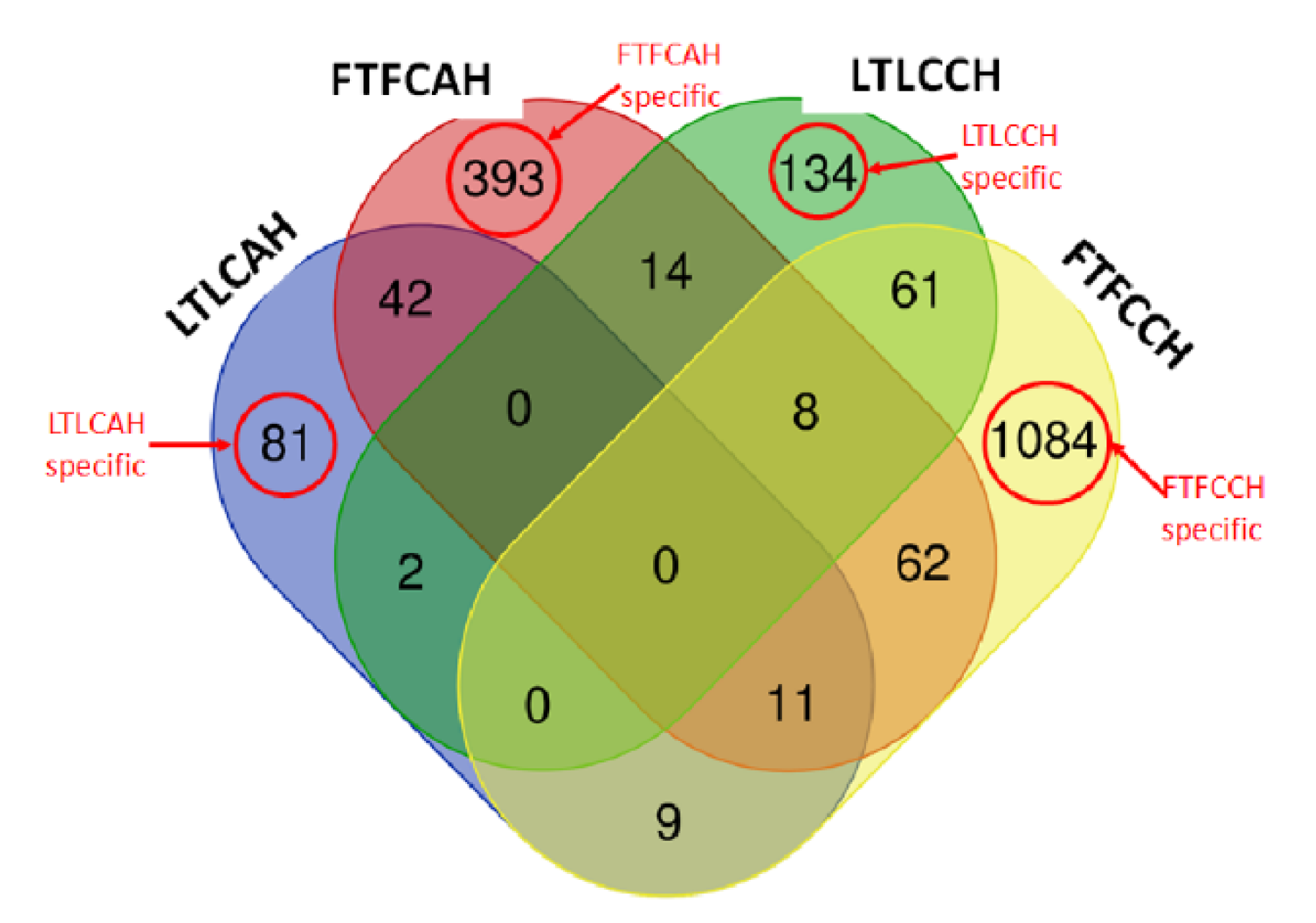
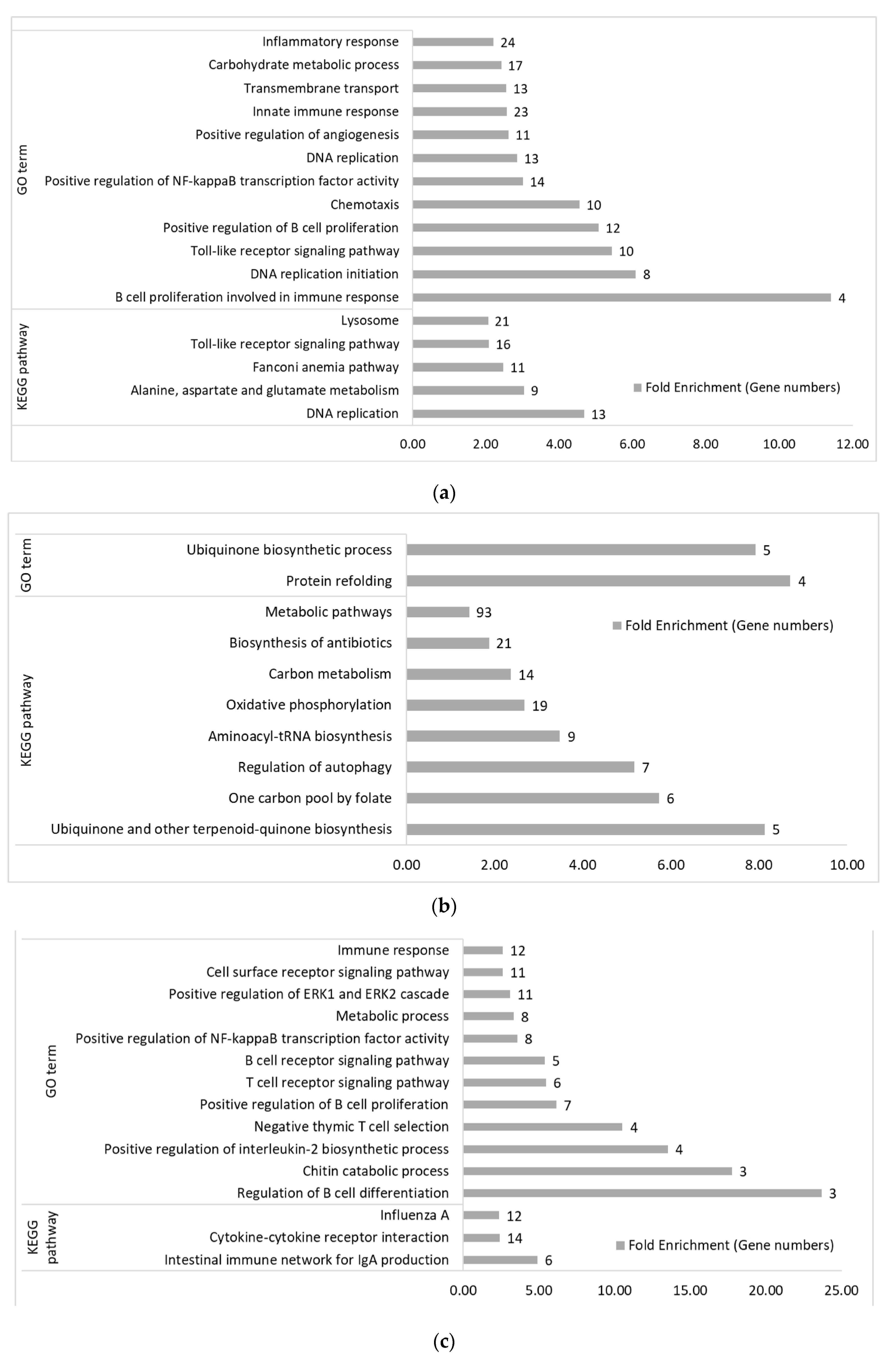
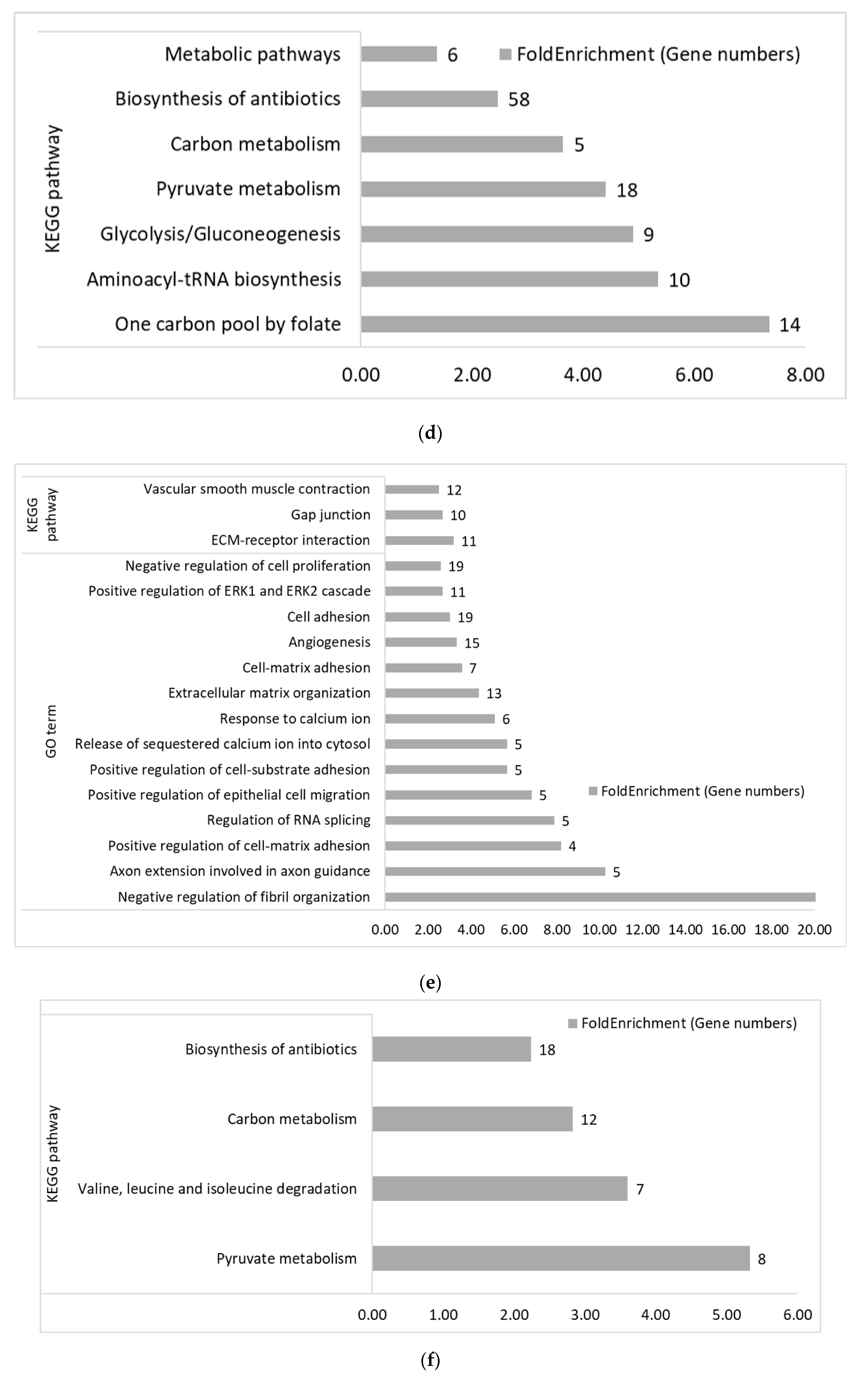
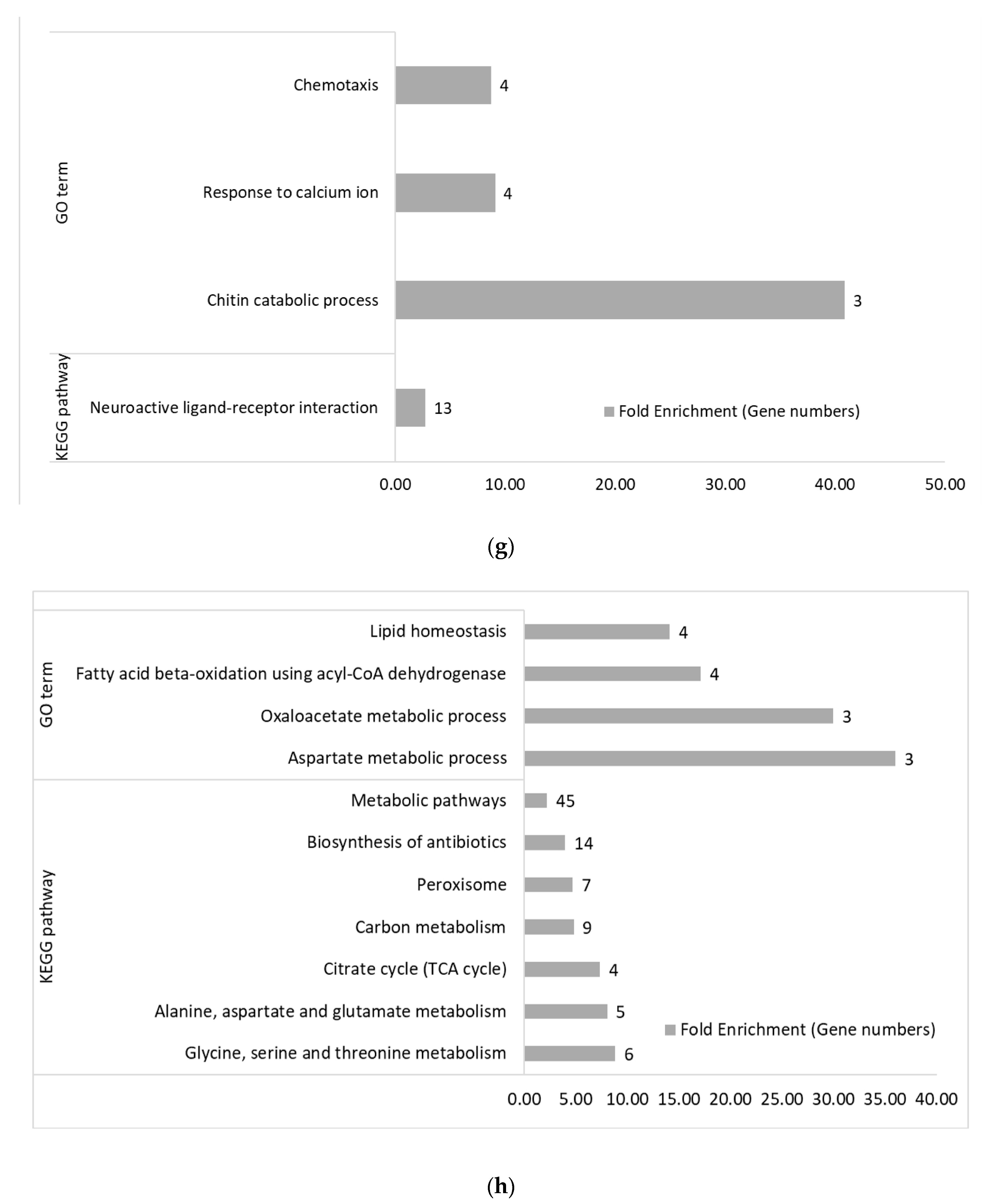
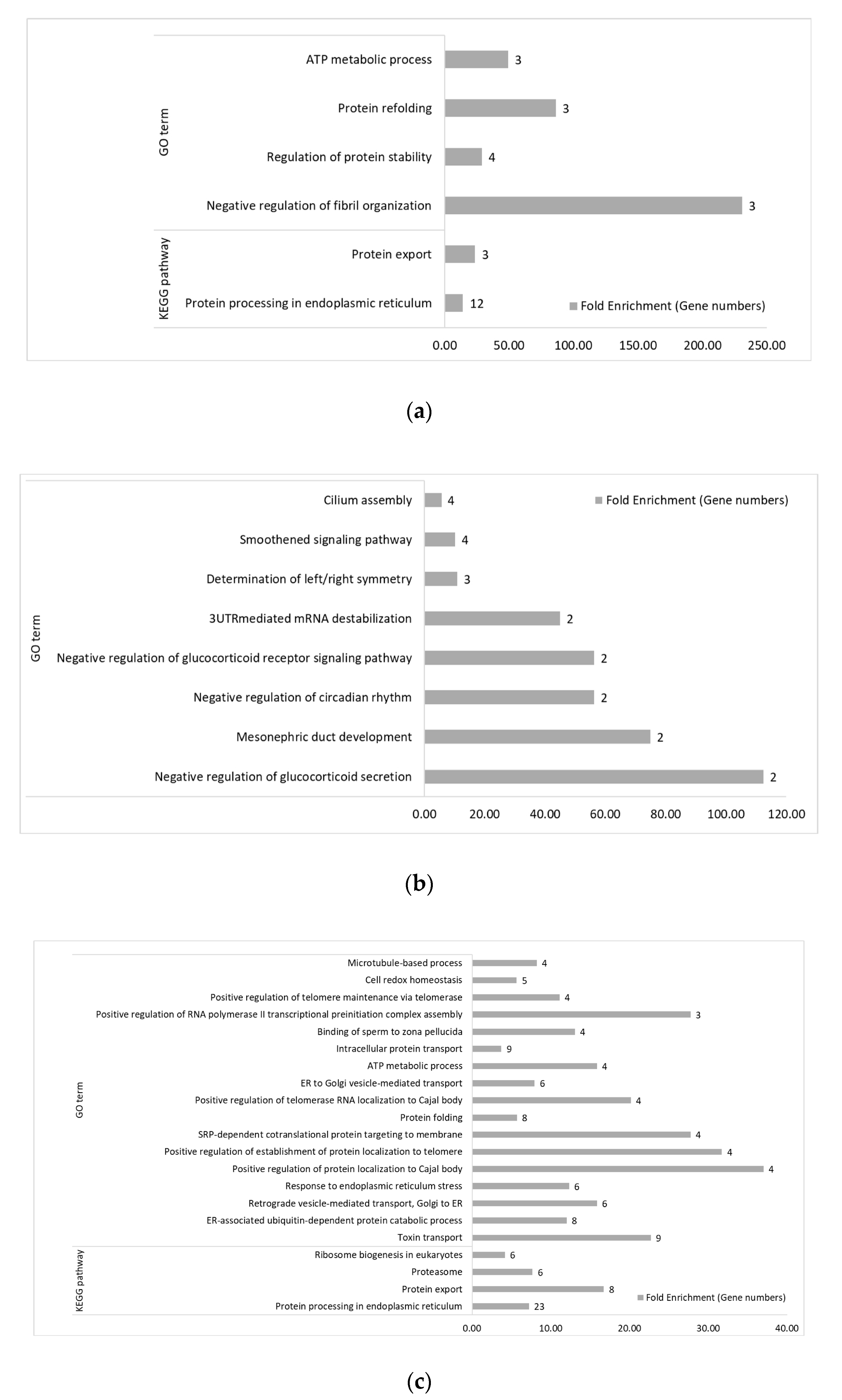
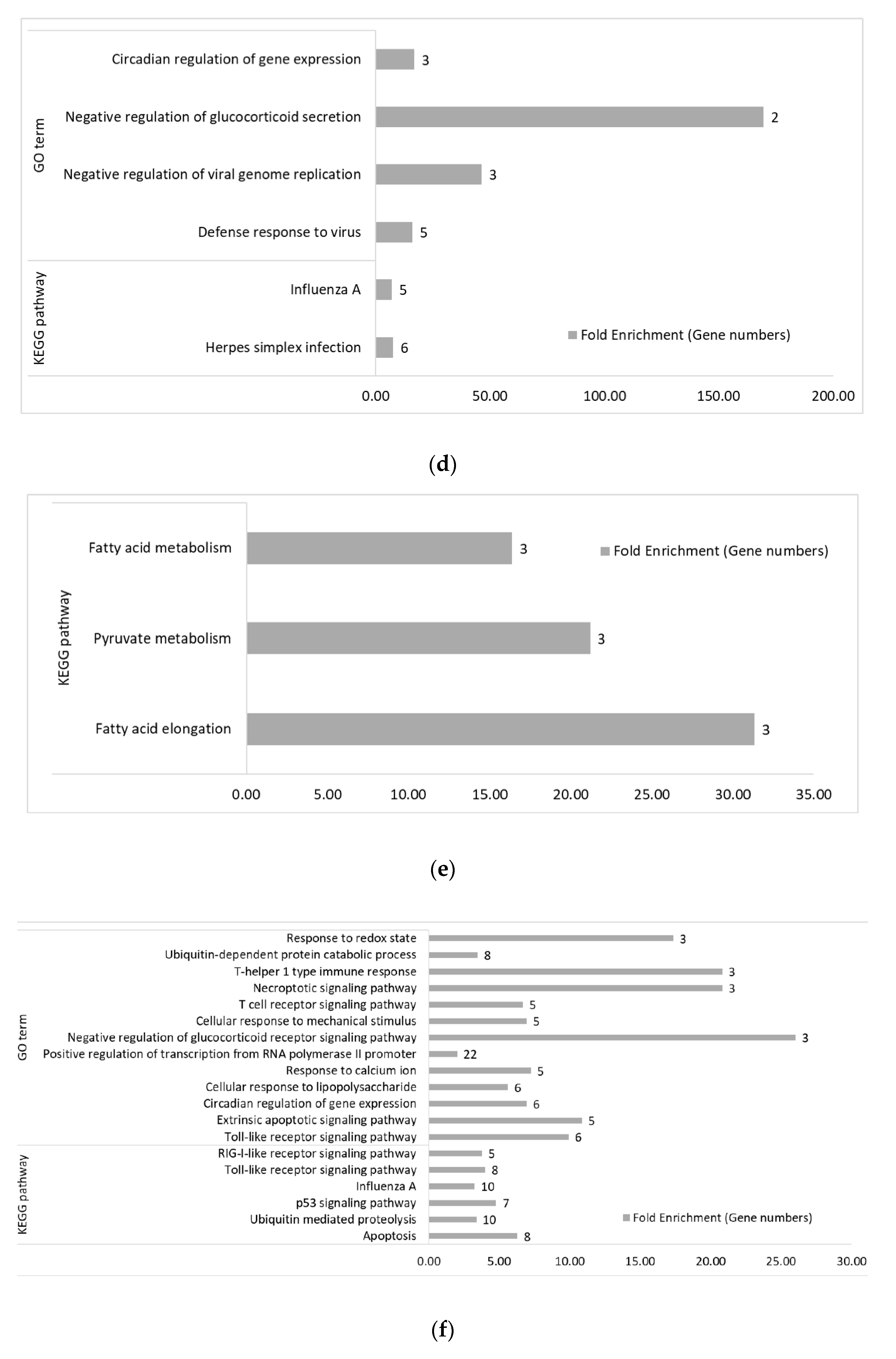
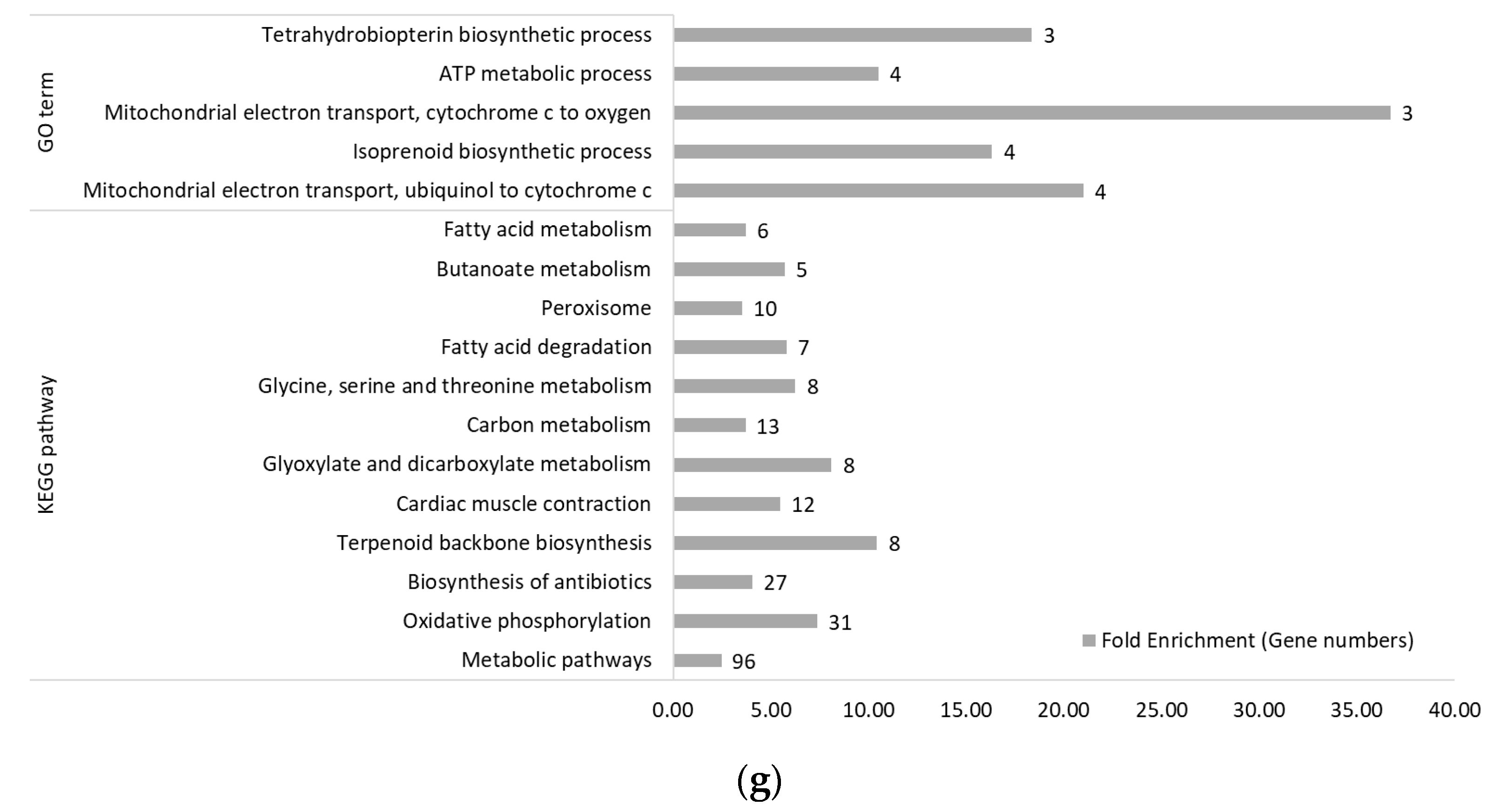
3.2. Functional Categories of Differentially Expressed Genes
3.2.1. Between Genetic Lines
3.2.2. Within Genetic Lines
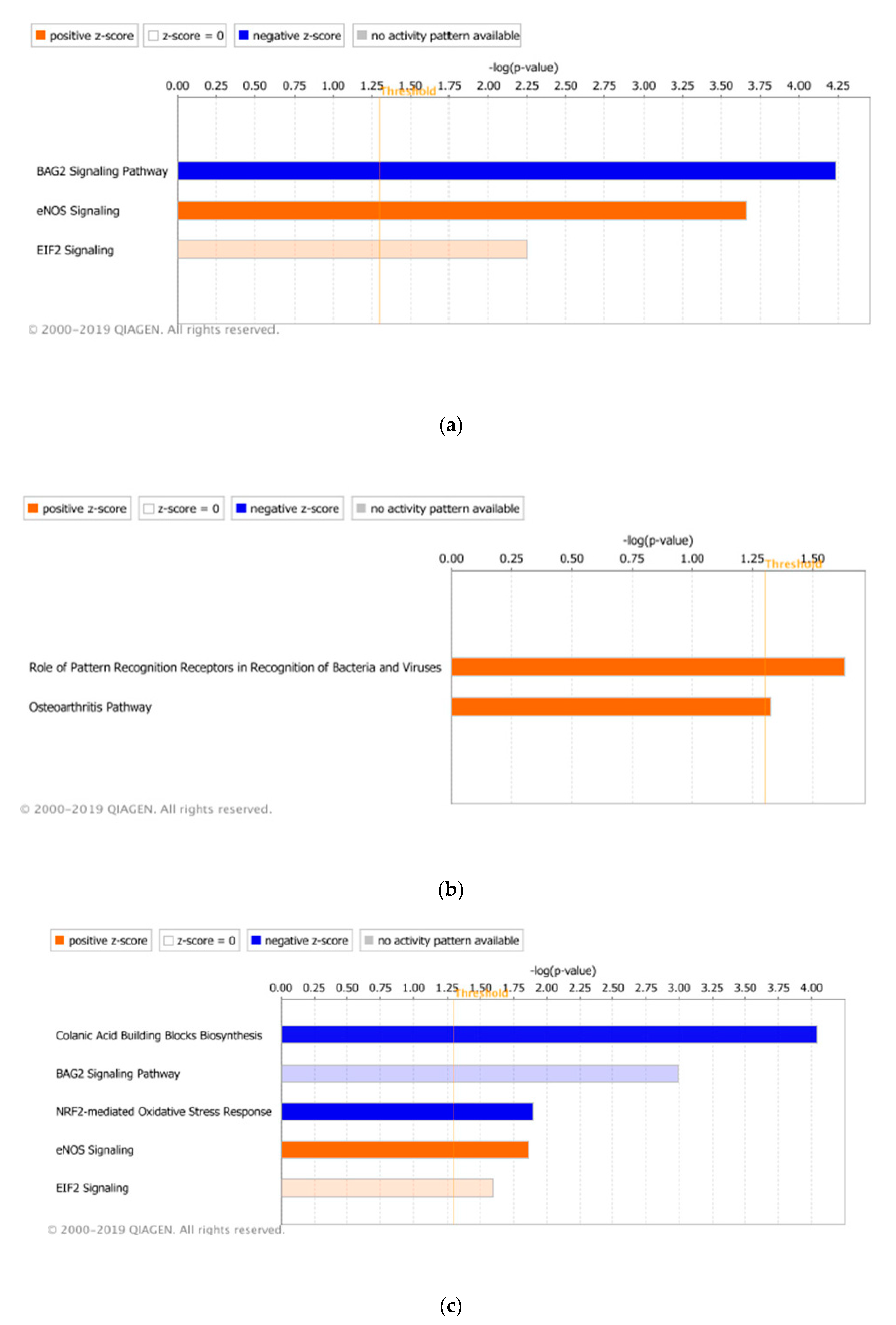
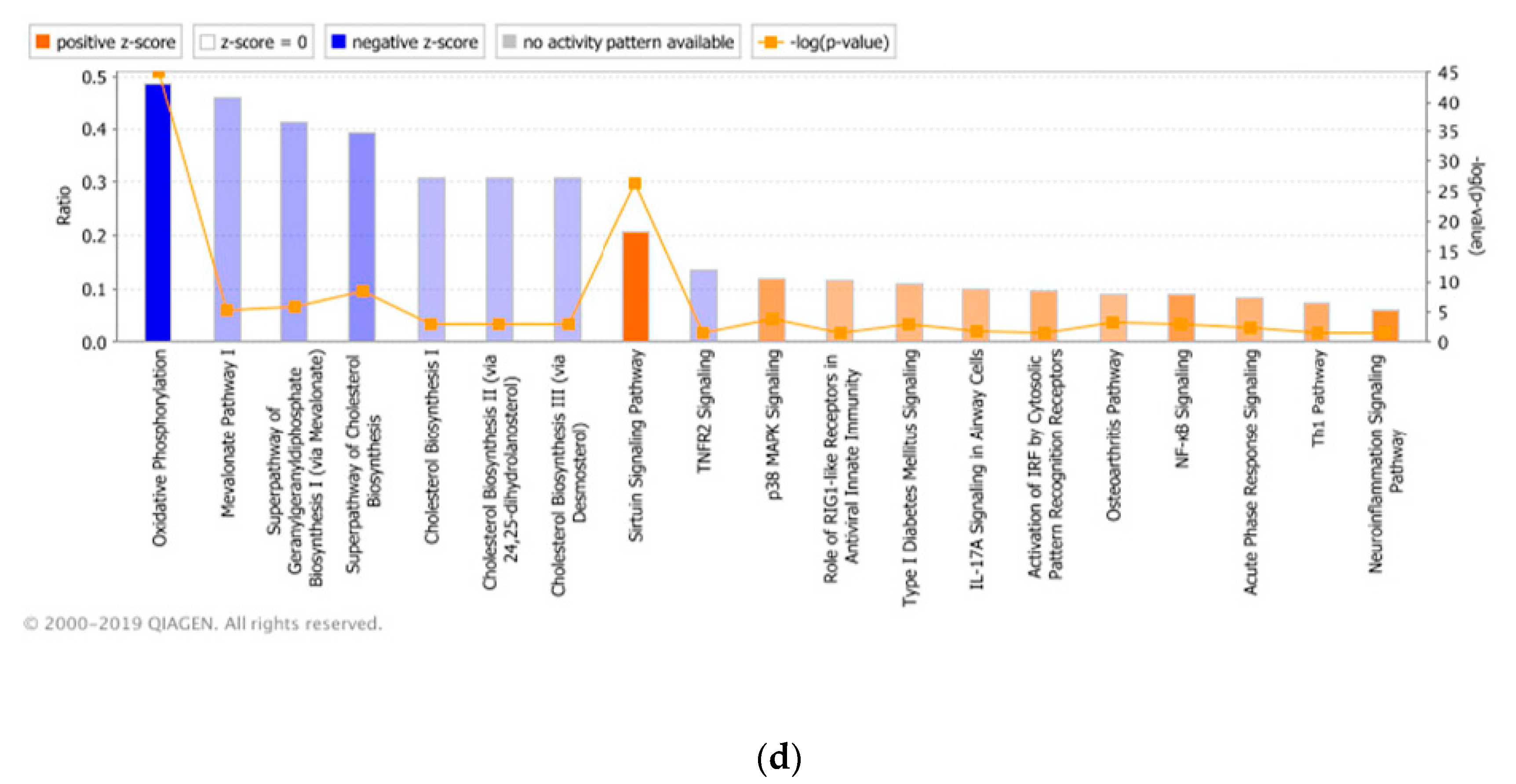
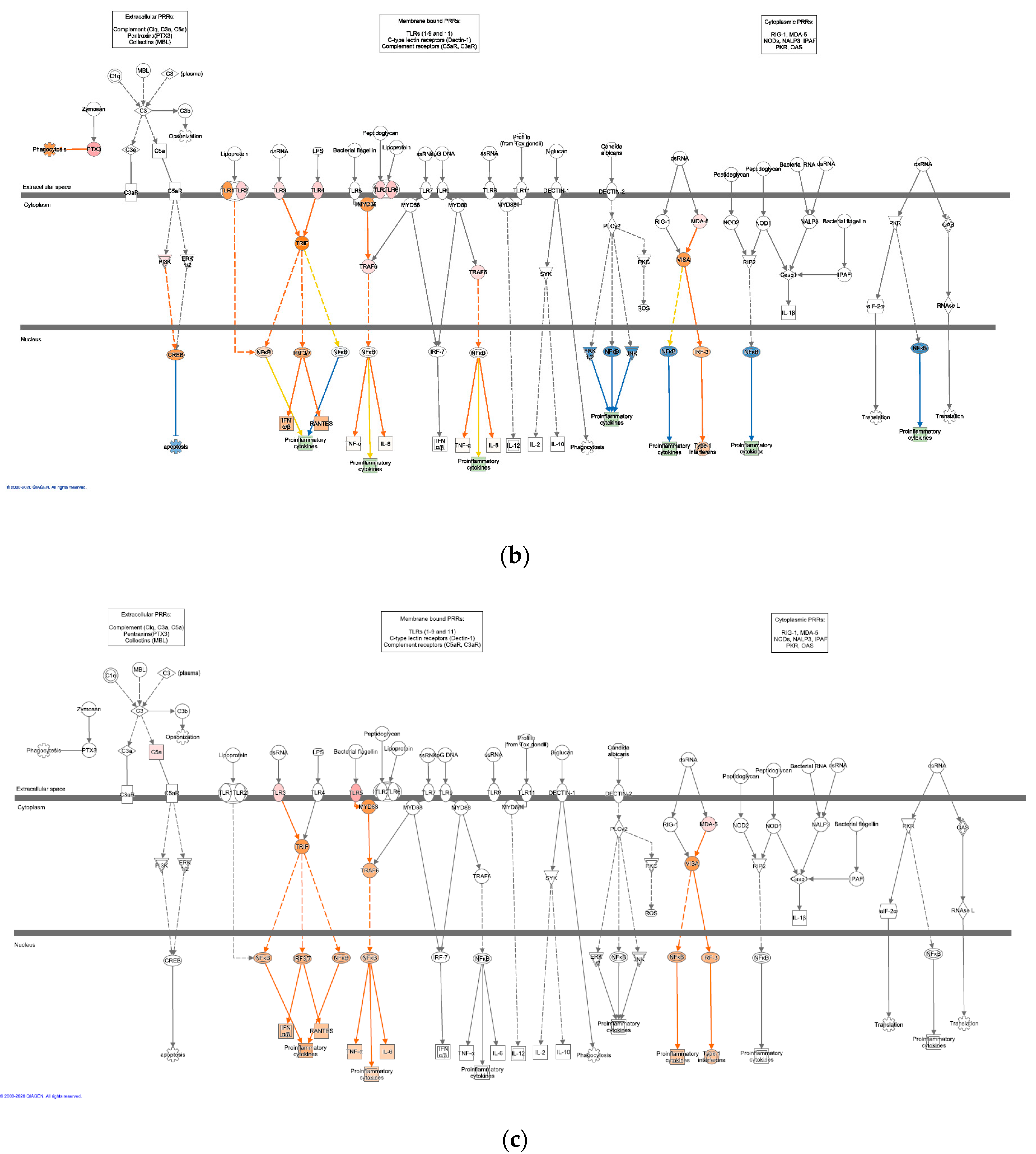
3.3. Significant Canonical Pathways Identified in Each Genetic Line with Treatments
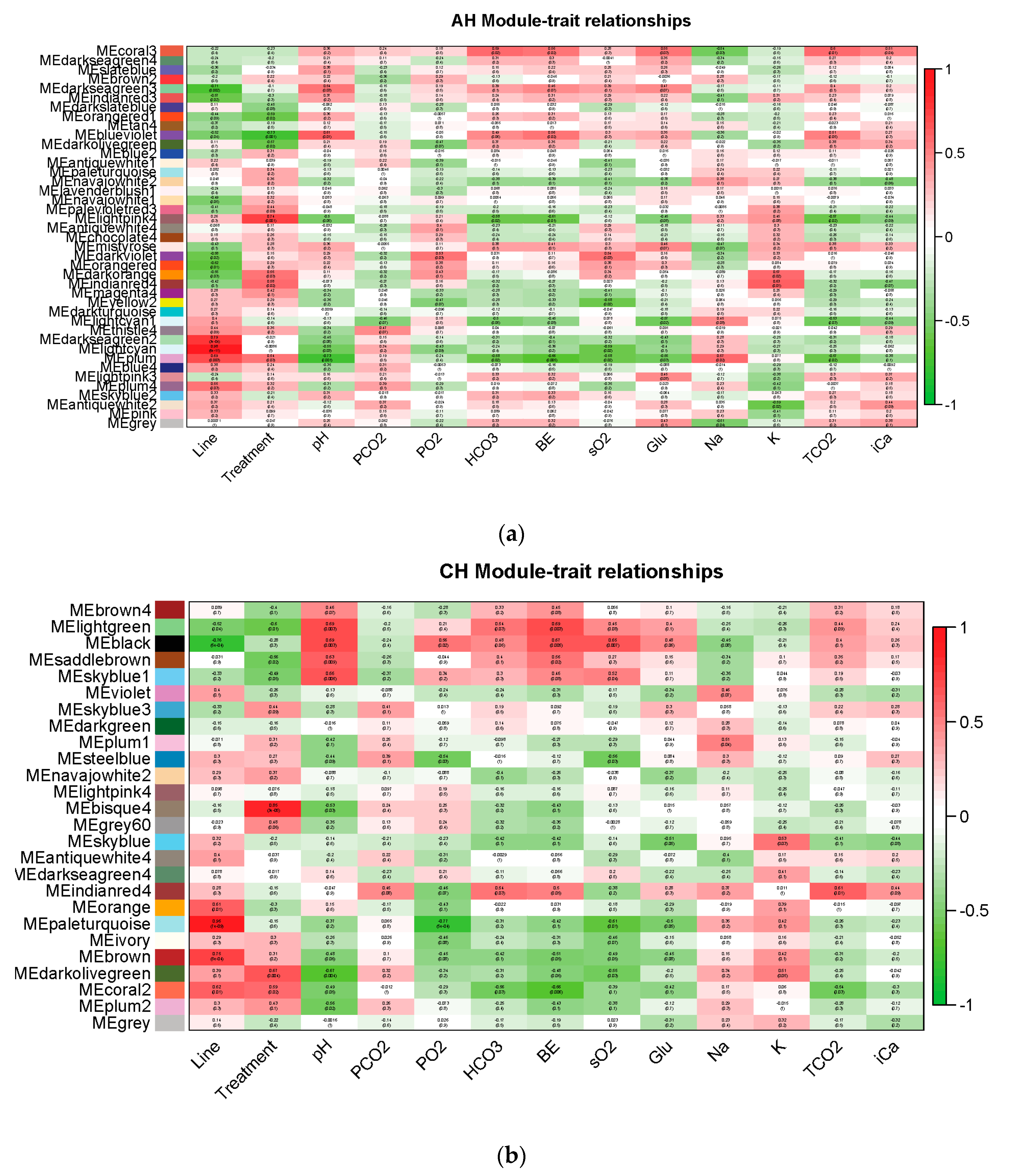
3.4. Weighted Gene Co-Expression Network Analysis (WGCNA)
4. Discussion
4.1. Host Response to Acute Heat Stress
4.1.1. The Leghorn line
4.1.2. The Fayoumi Line
4.2. Host Response to Chronic Heat Stress Combined with NDV Infection
4.2.1. The Leghorn Line
4.2.2. The Fayoumi Line
4.3. The Fayoumi Line had More Active Immune Response than the Leghorn Line with both Treatments
4.4. WGCNA Identified Potentially Important Gene Modules and Driver Genes
4.4.1. Significant Correlated Gene Modules
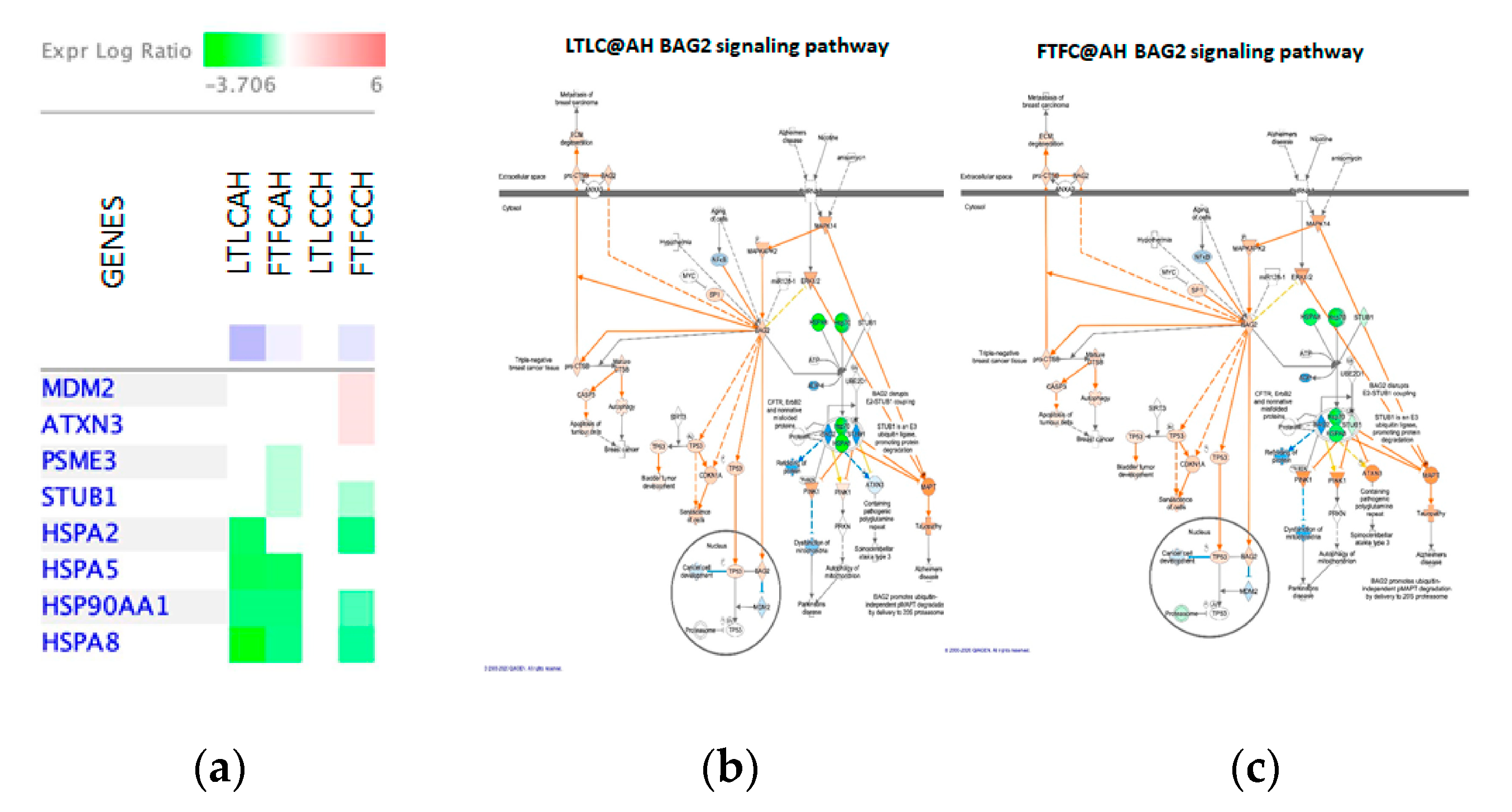
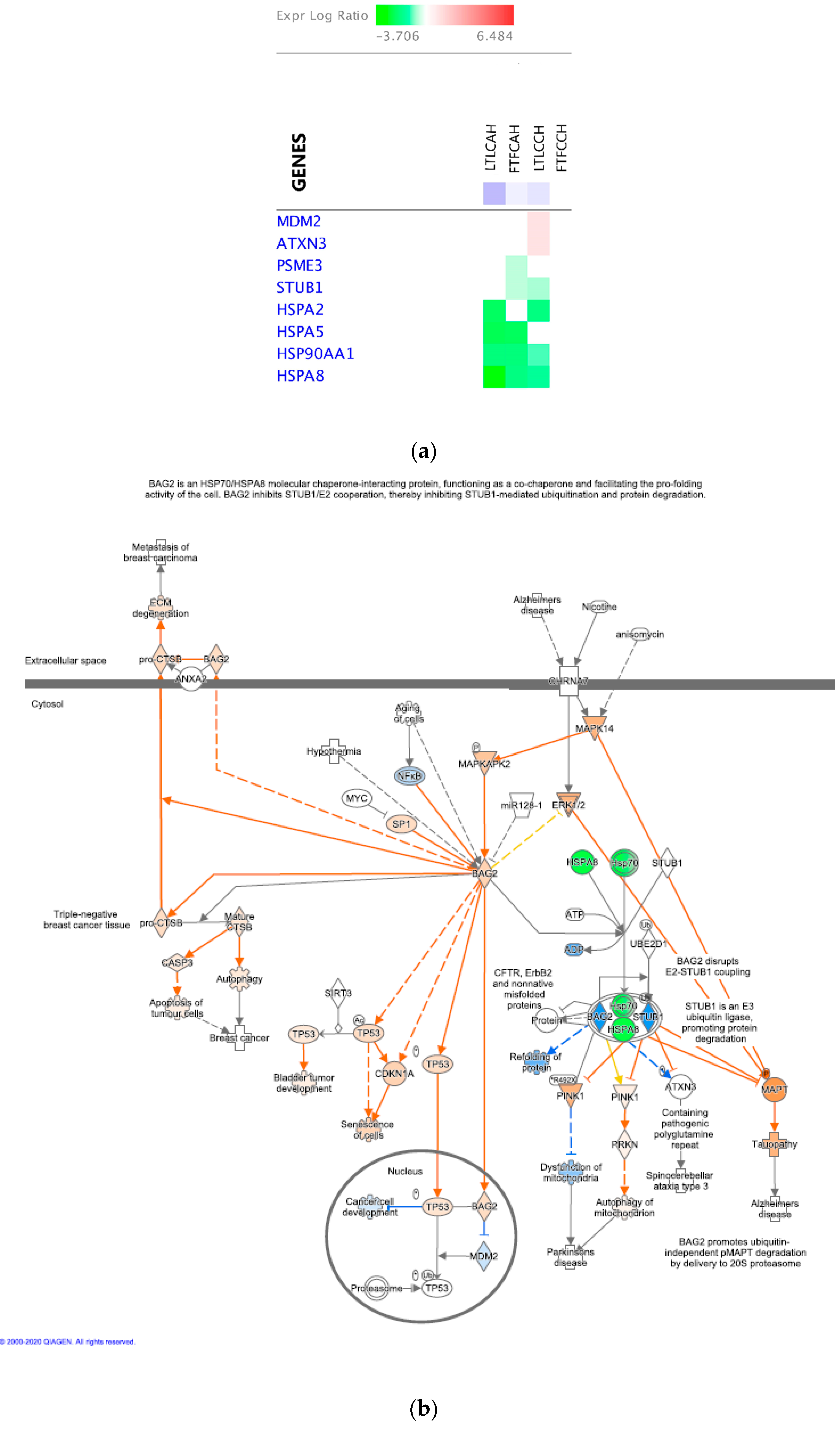
4.4.2. Other Potentially Important Driver Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rojas-Downing, M.M.; Nejadhashemi, A.P.; Harrigan, T.; Woznicki, S.A. Climate change and livestock: Impacts, adaptation, and mitigation. Clim. Risk Manag. 2017, 16, 145–163. [Google Scholar] [CrossRef]
- Lara, L.J.; Rostagno, M.H. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Settar, P.; Yalcin, S.; Turkmut, L.; Ozkan, S.; Cahanar, A. Season by genotype interaction related to broiler growth rate and heat tolerance. Poult. Sci. 1999, 78, 1353–1358. [Google Scholar] [CrossRef]
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic Losses from Heat Stress by US Livestock Industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef]
- Borges, S.A.; da Silva, A.V.F.; Majorka, A.; Hooge, D.M.; Cummings, K.R. Physiological responses of broiler chickens to heat stress and dietary electrolyte balance (sodium plus potassium minus chloride, milliequivalents per kilogram). Poult. Sci. 2004, 83, 1551–1558. [Google Scholar] [CrossRef]
- Brobeck, J.R. Food intake as a mechanism of temperature regulation. Obes. Res. 1997, 5, 641–645. [Google Scholar] [CrossRef]
- Wilson, T.E. Renal sympathetic nerve, blood flow, and epithelial transport responses to thermal stress. Auton. Neurosci. Basic Clin. 2017, 204, 25–34. [Google Scholar] [CrossRef]
- Ahmad, T.; Sarwar, M. Dietary electrolyte balance: Implications in heat stressed broilers. World Poult. Sci. J. 2006, 62, 638–653. [Google Scholar] [CrossRef]
- Bagath, M.; Krishnan, G.; Devaraj, C.; Rashamol, V.P.; Pragna, P.; Lees, A.M.; Sejian, V. The impact of heat stress on the immune system in dairy cattle: A review. Res. Vet. Sci. 2019, 126, 94–102. [Google Scholar] [CrossRef]
- Monson, M.S.; Van Goor, A.G.; Ashwell, C.M.; Persia, M.E.; Rothschild, M.F.; Schmidt, C.J.; Lamont, S.J. Immunomodulatory effects of heat stress and lipopolysaccharide on the bursal transcriptome in two distinct chicken lines. BMC Genom. 2018, 19, 643. [Google Scholar] [CrossRef]
- Kitalyi, A.J. Village Chicken Production Systems in Rural Africa: Household Food Security and Gender Issues; FAO: Rome, Italy, 1998. [Google Scholar]
- Abolnik, C. History of Newcastle disease in South Africa. Onderstepoort J. Vet. Res. 2017, 84, e1–e7. [Google Scholar] [CrossRef]
- Mack, L.A.; Felver-Gant, J.N.; Dennis, R.L.; Cheng, H.W. Genetic variations alter production and behavioral responses following heat stress in 2 strains of laying hens. Poult. Sci. 2013, 92, 285–294. [Google Scholar] [CrossRef]
- Wang, Y.; Saelao, P.; Chanthavixay, K.; Gallardo, R.; Bunn, D.; Lamont, S.J.; Dekkers, J.M.; Kelly, T.; Zhou, H. Physiological responses to heat stress in two genetically distinct chicken inbred lines. Poult. Sci. 2018, 97, 770–780. [Google Scholar] [CrossRef]
- Rui, L.Y. Energy Metabolism in the Liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic. Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinformatics 2008, 9, 559. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. Fast R Functions for Robust Correlations and Hierarchical Clustering. J. Stat. Softw. 2012, 46, 1–17. [Google Scholar] [CrossRef]
- Horvath, S.; Dong, J. Geometric interpretation of gene coexpression network analysis. PLoS Comput. Biol. 2008, 4, e1000117. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kawakami, E.; Shoemaker, J.E.; Lopes, T.J.S.; Matsuoka, Y.; Tomita, Y.; Kozuka-Hata, H.; Gorai, T.; Kuwahara, T.; Takeda, E.; et al. Influenza Virus-Host Interactome Screen as a Platform for Antiviral Drug Development. Cell Host Microbe 2014, 16, 795–805. [Google Scholar] [CrossRef] [PubMed]
- Livant, E.J.; Avendano, S.; McLeod, S.; Ye, X.; Lamont, S.J.; Dekkers, J.C.M.; Ewald, S.J. MX1 exon 13 polymorphisms in broiler breeder chickens and associations with commercial traits. Anim. Genet. 2007, 38, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Saelao, P.; Wang, Y.; Gallardo, R.A.; Lamont, S.J.; Dekkers, J.M.; Kelly, T.; Zhou, H. Novel insights into the host immune response of chicken Harderian gland tissue during Newcastle disease virus infection and heat treatment. BMC Vet. Res. 2018, 14, 280. [Google Scholar] [CrossRef]
- Saelao, P.; Wang, Y.; Chanthavixay, G.; Yu, V.; Gallardo, R.A.; Dekkers, J.C.M.; Lamont, S.J.; Kelly, T.; Zhou, H. Integrated Proteomic and Transcriptomic Analysis of Differential Expression of Chicken Lung Tissue in Response to NDV Infection during Heat Stress. Genes 2018, 9, 579. [Google Scholar] [CrossRef]
- Jastrebski, S.F.; Lamont, S.J.; Schmidt, C.J. Chicken hepatic response to chronic heat stress using integrated transcriptome and metabolome analysis. PLoS ONE 2017, 12, e0181900. [Google Scholar] [CrossRef]
- Schonbuhler, B.; Schmitt, V.; Huesmann, H.; Kern, A.; Gamerdinger, M.; Behl, C. BAG2 Interferes with CHIP-Mediated Ubiquitination of HSP72. Int. J. Mol. Sci. 2016, 18, 69. [Google Scholar] [CrossRef]
- Zhang, H.T.; Zeng, L.F.; He, Q.Y.; Tao, W.A.; Zha, Z.G.; Hu, C.D. The E3 ubiquitin ligase CHIP mediates ubiquitination and proteasomal degradation of PRMT5. Biochim. Biophys. Acta 2016, 1863, 335–346. [Google Scholar] [CrossRef]
- Kim, J.; Cha, Y.N.; Surh, Y.J. A protective role of nuclear factor-erythroid 2-related factor-2 (Nrf2) in inflammatory disorders. Mutat. Res. 2010, 690, 12–23. [Google Scholar] [CrossRef]
- Battino, M.; Giampieri, F.; Pistollato, F.; Sureda, A.; de Oliveira, M.R.; Pittala, V.; Fallarino, F.; Nabavi, S.F.; Atanasov, A.G.; Nabavi, S.M. Nrf2 as regulator of innate immunity: A molecular Swiss army knife! Biotechnol. Adv. 2018, 36, 358–370. [Google Scholar] [CrossRef]
- Wang, Y.; Lupiani, B.; Reddy, S.M.; Lamont, S.J.; Zhou, H. RNA-seq analysis revealed novel genes and signaling pathway associated with disease resistance to avian influenza virus infection in chickens. Poult. Sci. 2014, 93, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Katibah, G.E.; Qin, Y.; Sidote, D.J.; Yao, J.; Lambowitz, A.M.; Collins, K. Broad and adaptable RNA structure recognition by the human interferon-induced tetratricopeptide repeat protein IFIT5. Proc. Natl. Acad. Sci. USA 2014, 111, 12025–12030. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Li, Y.; Zhang, Y.; Zhang, L.; Wang, Z.; Zhang, X.; Gui, L.; Huang, J. Correction: IFI6 Inhibits Apoptosis via Mitochondrial-Dependent Pathway in Dengue Virus 2 Infected Vascular Endothelial Cells. PLoS ONE 2015, 10, e0138896. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fang, P.; Yang, W.Y.; Chan, K.; Lavallee, M.; Xu, K.; Gao, T.; Wang, H.; Yang, X. Mitochondrial ROS, uncoupled from ATP synthesis, determine endothelial activation for both physiological recruitment of patrolling cells and pathological recruitment of inflammatory cells. Can. J. Physiol. Pharmacol. 2017, 95, 247–252. [Google Scholar] [CrossRef]
- Akbarian, A.; Michiels, J.; Degroote, J.; Majdeddin, M.; Golian, A.; De Smet, S. Association between heat stress and oxidative stress in poultry; mitochondrial dysfunction and dietary interventions with phytochemicals. J. Anim. Sci. Biotechnol. 2016, 7, 37. [Google Scholar] [CrossRef]
- Yang, L.; Tan, G.Y.; Fu, Y.Q.; Feng, J.H.; Zhang, M.H. Effects of acute heat stress and subsequent stress removal on function of hepatic mitochondrial respiration, ROS production and lipid peroxidation in broiler chickens. Comp. Biochem. Physiol. C Toxico.l Pharmacol. 2010, 151, 204–208. [Google Scholar] [CrossRef]
- Mishra, B.; Jha, R. Oxidative Stress in the Poultry Gut: Potential Challenges and Interventions. Front. Vet. Sci. 2019, 6, 60. [Google Scholar] [CrossRef]
- Van Goor, A.; Bolek, K.J.; Ashwell, C.M.; Persia, M.E.; Rothschild, M.F.; Schmidt, C.J.; Lamont, S.J. Identification of quantitative trait loci for body temperature, body weight, breast yield, and digestibility in an advanced intercross line of chickens under heat stress. Genet. Sel. Evol. 2015, 47, 96. [Google Scholar] [CrossRef]
- Fleming, D.S.; Koltes, J.E.; Fritz-Waters, E.R.; Rothschild, M.F.; Schmidt, C.J.; Ashwell, C.M.; Persia, M.E.; Reecy, J.M.; Lamont, S.J. Single nucleotide variant discovery of highly inbred Leghorn and Fayoumi chicken breeds using pooled whole genome resequencing data reveals insights into phenotype differences. BMC Genom. 2016, 17, 812. [Google Scholar] [CrossRef]
- Bereshchenko, O.; Bruscoli, S.; Riccardi, C. Glucocorticoids, Sex Hormones, and Immunity. Front. Immunol. 2018, 9, 1332. [Google Scholar] [CrossRef]
- Messaoudi, L.; Yang, Y.G.; Kinomura, A.; Stavreva, D.A.; Yan, G.; Bortolin-Cavaille, M.L.; Arakawa, H.; Buerstedde, J.M.; Hainaut, P.; Cavaille, J.; et al. Subcellular distribution of human RDM1 protein isoforms and their nucleolar accumulation in response to heat shock and proteotoxic stress. Nucleic. Acids Res. 2007, 35, 6571–6587. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Kimura, M.; Miyazaki, H.; Okawa, K.; Onuki, R.; Nemoto, C.; Tabata, E.; Wakita, S.; Kashimura, A.; Sakaguchi, M.; et al. Acidic mammalian chitinase is a proteases-resistant glycosidase in mouse digestive system. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shen, Z.; Wu, J. Expression, purification and in vitro antifungal activity of acidic mammalian chitinase against Candida albicans, Aspergillus fumigatus and Trichophyton rubrum strains. Clin. Exp. Dermatol. 2009, 34, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Hebert, S.C.; Mount, D.B.; Gamba, G. Molecular physiology of cation-coupled Cl- cotransport: The SLC12 family. Pflug. Arch. Eur. J. Physiol. 2004, 447, 580–593. [Google Scholar] [CrossRef] [PubMed]
- Mount, D.B.; Romero, M.F. The SLC26 gene family of multifunctional anion exchangers. Pflug. Arch. Eur. J. Physiol. 2004, 447, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Blackman, S.M.; Commander, C.W.; Watson, C.; Arcara, K.M.; Strug, L.J.; Stonebraker, J.R.; Wright, F.A.; Rommens, J.M.; Sun, L.; Pace, R.G.; et al. Genetic Modifiers of Cystic Fibrosis-Related Diabetes. Diabetes 2013, 62, 3627–3635. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Treatment | Time Point | Raw Reads | Aligned Reads | Alignment Rate |
|---|---|---|---|---|---|
| Leghorn | Non-treated | Acute Heat | 199,693,733 | 183,292,871 | 91.79% |
| Leghorn | Treated | Acute Heat | 187,612,958 | 170,964,578 | 91.13% |
| Fayoumi | Non-treated | Acute Heat | 166,442,903 | 152,955,781 | 91.90% |
| Fayoumi | Treated | Acute Heat | 195,204,499 | 178,279,792 | 91.33% |
| Leghorn | Non-treated | CH&NDV | 106,510,459 | 98,128,436 | 92.13% |
| Leghorn | Treated | CH&NDV | 120,871,158 | 111,479,701 | 92.23% |
| Fayoumi | Non-treated | CH&NDV | 94,966,874 | 88,659,977 | 93.36% |
| Fayoumi | Treated | CH&NDV | 115,633,054 | 108,848,582 | 94.13% |
| Fayoumi DEG 1 | Gene Description | AH 2 (log2foldchange) | CH&NDV 3 (log2foldchange) |
|---|---|---|---|
| ZNFX1 | Zinc finger NFX1-type containing 1 | −1.03 | 1.61 |
| USP18 | Ubiquitin specific peptidase 18 | −1.64 | 1.22 |
| MX1 | MX dynamin like GTPase 1 | −1.36 | 2.07 |
| CMPK2 | Cytidine/uridine monophosphate kinase 2 | −1.49 | 1.68 |
| Novel gene | Chromosome 3 open reading frame, human C8orf80 | −1.19 | 1.24 |
| TLX1 | T cell leukemia homeobox 1 | 2.56 | −1.87 |
| HSPB9 | Heat shock protein family B (small) member 9 | 1.59 | −2.42 |
| FAM222A | Family with sequence similarity 222 member A | 1.35 | −1.85 |
| Factor | Positive Correlation | Negative Correlation | ||
|---|---|---|---|---|
| AH | CH&NDV | AH | CH&NDV | |
| Line 1 | Lightcyan (0.98 2) | Paleturquoise (0.96) | Darkseagreen3 (−0.71) | Black (−0.76) |
| Treatment | Lightpink4 (0.74) | Bisque4 (0.85) | Blueviolet (−0.73) | Lightgreen(−0.60) |
| pH | Blueviolet (0.61) | Black (0.69)/ Lightgreen (0.69) | Plum (−0.73) | Darkolivergreen (−0.67) |
| PCO2 | NS 3 | NS | NS | NS |
| HCO3 | Coral3 (0.59) | Lightgreen (0.54)/ Indianred4 (0.54) | Plum (−0.58)/ Lightpink4 (−0.58) | Coral2 (−0.56) |
| TCO2 | Coral3 (0.60) | Indianred4 (0.61) | Plum (−0.57)/ Lightpink4 (−0.57) | Coral2 (−0.54) |
| BE | Coral3 (0.56)/ Blueviolet (0.56) | Lightgreen (0.69) | Plum (−0.66) | Coral2 (−0.66) |
| Glu | Coral3 (0.55) | NS | Lightcyan1 (−0.57) | Skyblue (−0.51) |
| iCa2+ | Coral3 (0.51) | NS | NS | NS |
| PO2 | Darkviolet (0.53) | Black (0.56) | NS | Paleturquoise (−0.77) |
| sO2 | Darkviolet (0.54) | Black (0.65) | Lightcyan (−0.59) | Paleturquoise (−0.61) |
| Na+ | Plum (0.57) | Plum1 (0.51) | Coral3 (−0.54) | NS |
| K+ | Indianred4 (0.63) | Skyblue (0.53) | Antiquewhite2 (−0.59) | NS |
| Module | Lightcyan | Plum | Lightpink4 | Coral3 | Blueviolet | Darkviolet | |
|---|---|---|---|---|---|---|---|
| Factor | |||||||
| Line | 0.98 1 | 0.69 | 0.26 | −0.22 | −0.52 | −0.58 | |
| Treatment | −0.01 | 0.54 | 0.74 | −0.23 | −0.73 | 0.15 | |
| pH | −0.55 | −0.73 | −0.5 | 0.36 | 0.61 | 0.29 | |
| PCO2 | 0.34 | 0.19 | −0.09 | 0.24 | −0.15 | −0.32 | |
| TCO2 | −0.25 | −0.57 | −0.57 | 0.60 | 0.51 | 0.02 | |
| HCO3 | −0.24 | −0.58 | −0.58 | 0.59 | 0.49 | 0.03 | |
| BE | −0.36 | −0.66 | −0.61 | 0.56 | 0.56 | 0.11 | |
| PO2 | −0.43 | −0.24 | 0.21 | 0.18 | −0.05 | 0.53 | |
| sO2 | −0.59 | −0.58 | −0.12 | 0.28 | 0.3 | 0.54 | |
| Na+ | 0.29 | 0.57 | 0.33 | −0.54 | −0.36 | −0.34 | |
| K+ | −0.27 | 0.077 | 0.45 | −0.19 | −0.02 | 0.33 | |
| iCa2+ | −0.06 | −0.38 | −0.44 | 0.51 | 0.27 | −0.05 | |
| Glu | −0.40 | −0.55 | −0.45 | 0.55 | 0.35 | 0.16 | |
| Module | Paleturquoise | Coral2 | Skyblue | Indianred4 | Lightgreen | Black | |
|---|---|---|---|---|---|---|---|
| Factor | |||||||
| Line | 0.96 1 | 0.62 | 0.32 | 0.28 | −0.52 | −0.76 | |
| Treatment | −0.15 | 0.59 | −0.20 | −0.15 | −0.60 | −0.28 | |
| pH | −0.37 | −0.49 | −0.14 | −0.05 | 0.69 | 0.69 | |
| PCO2 | 0.07 | −0.01 | −0.21 | 0.45 | −0.20 | −0.24 | |
| TCO2 | −0.26 | −0.54 | −0.41 | 0.61 | 0.44 | 0.40 | |
| HCO3 | −0.31 | −0.56 | −0.42 | 0.54 | 0.54 | 0.48 | |
| BE | −0.42 | −0.66 | −0.42 | 0.50 | 0.69 | 0.67 | |
| PO2 | −0.77 | −0.29 | −0.23 | −0.46 | 0.21 | 0.56 | |
| sO2 | −0.61 | −0.39 | −0.14 | −0.38 | 0.45 | 0.65 | |
| Na+ | 0.35 | 0.17 | 0.10 | 0.37 | −0.25 | −0.45 | |
| K+ | 0.42 | 0.06 | 0.53 | 0.01 | −0.26 | −0.21 | |
| iCa2+ | −0.23 | −0.30 | −0.44 | 0.44 | 0.24 | 0.26 | |
| Glu | −0.50 | −0.42 | −0.51 | 0.28 | 0.40 | 0.48 | |
| Factor | Correlation | Module Color | Gene Name (GS 1, MM 2) |
|---|---|---|---|
| Line | Positive | Lightcyan | RDM1 (1.00,0.98), CHIA (0.99,0.95), SMDT1 (0.98,0.95), IFT57 (0.98,0.96), ERMP1 (0.98,0.95) |
| Negative | Darkseagreen3 | FAXDC2 (0.81, −0.93), FMO3 (0.80, −0.95), CYP2J21 (0.80, −0.91), PLXNA4 (0.79, −0.85), KLHL8 (0.78, −0.85) | |
| Treatment | Positive | Lightpink4 | BTBD7 (0.80, 0.67), MCF2 (0.78, 0.63), PLD1 (0.78, 0.82), GRIP2 (0.75, 0.64), CECR2 (0.75, 0.80) |
| Negative | Blueviolet | CACNG5 (0.95, −0.65), ERLEC1 (−0.91, 0.82), PSMC1 (−0.90, 0.88), EIF2A (−0.90, 0.77), HSPA5 (−0.89, 0.86) | |
| pH | Positive | Blueviolet | gga-mir−1723 (0.72, 0.81), NEMF (0.72, 0.79), HSP90AA1 (0.71, 0.93), SF3B3 (0.71, 0.71), VKORC1L1 (0.71, 0.92) |
| Negative | Plum | TTC30B (−0.85, 0.81), ENSGALG00000040869 (−0.80, 0.88), PPCDC (−0.79, 0.88), HSP90AB1 (0.78, −0.88), TTC17 (−0.77, 0.88) | |
| HCO3 | Positive | Coral3 | ENSGALG00000045967 (0.76, 0.73), ARHGEF33 (0.71, 0.88), SLC24A1 (0.67, 0.63), PRSS12 (0.66, 0.50), SPTAN1 (0.64, 0.65) |
| Negative | Lightpink4 | TGS1 (0.75, −0.82), UPF3B (0.73, −0.94), ZBTB8OS (0.73, −0.79), SRSF2 (0.73, −0.86), gga-mir−6585 (0.72, −0.86) | |
| Plum | ENSGALG00000007624 (0.82, −0.56), TTC30B (−0.76, 0.81), BARX2 (−0.71, 0.70), RF00392 (0.70, −0.77), NLE1 (0.69, −0.79) | ||
| TCO2 | Positive | Coral3 | ARHGEF33 (0.70, 0.88), SLC24A1 (0.64, 0.63), CFAP58 (0.63, 0.78), PRSS12 (0.63, 0.50), SPTAN1 (0.62, 0.65) |
| Negative | Lightpink4 | TGS1 (0.76, −0.82), ZBTB8OS (0.74, −0.79), SRSF2 (0.72, −0.86), UPF3B (0.71, −0.94), GTF2B (0.70, −0.81) | |
| Plum | ENSGALG00000007624 (0.78, −0.56), TTC30B (−0.75, 0.81), RF00392 (0.71, −0.77), BARX2 (−0.70, 0.70), NLE1 (0.68, −0.79) | ||
| BE | Positive | Coral3 | ENSGALG00000045967 (0.79, 0.73), PRSS12 (0.75, 0.50), ARHGEF33 (0.73, 0.88), SPTAN1 (0.71, 0.65), SLC24A1 (0.70, 0.63) |
| Blueviolet | INSIG1 (0.76, 0.65), SUCO (0.74, 0.59), ENOX2 (0.72, 0.73), NABP1 (0.69, 0.86), USP14 (0.69, 0.94) | ||
| Negative | Plum | TT30B (−0.85, 0.81), ENSGALG00000007624 (0.80, −0.56), PPCDC (−0.73, 0.88), NLE1 (0.72, −0.79), RF00392 (0.71, −0.77) | |
| Glu | Positive | Coral3 | CFAP58 (0.58, 0.78), ENSGALG00000045967 (0.57, 0.73), CAMK2N1 (0.57, 0.72), ENSGALG00000035761 (0.54, 0.78), ENSGALG00000040923 (0.53, 0.68) |
| Negative | Lightcyan1 | ST6GALNAC3 (−0.82, 0.65), COL20A1 (−0.82, 0.69), SLC10A4 (0.67, 0.68), MYOM2 (−0.66, 0.81), HTR7L (−0.60, 0.80) | |
| iCa2+ | Positive | Coral3 | ENSGALG00000035761 (0.60, 0.78), SPTAN1 (0.58, 0.65), ENSGALG00000045967 (0.52, 0.73), ENSGALG00000011252 (0.51, 0.91), CAMK2N1 (−0.55, 0.51) |
| PO2 | Positive | Darkviolet | VEZF1 (0.76, 0.59), CDON (0.76, 0.80), ENSGALG00000028262 (0.75, 0.81), CNGA2 (0.72, 0.58), MN1 (0.72, 0.83) |
| sO2 | Positive | Darkviolet | DDR2 (0.76, 0.77), CNGA2 (0.75, 0.58), ENSGALG00000026460 (0.72, 0.71), MAPK8IP1 (0.72, 0.88), KLF10 (0.71, 0.57) |
| Negative | lightcyan | TRIM37 (−0.82, 0.80), SEPTIN10 (−0.81, 0.87), C11H16ORF87 (−80, 0.77), ACE2 (−0.79, 0.84), BLOC1S4 (−0.78, 0.88) | |
| Na+ | Positive | Plum | MXD4 (0.81, 0.75), MID1 (0.75, 0.88), BARX2 (0.74, 0.70), TTC30B (0.73, 0.81), MAP2K5 (0.66, 0.84) |
| Negative | Coral3 | ENSGALG00000045967 (−0.79 0.73), FAM81A (0.72, −0.83), ARHGEF33 (−0.66, 0.88), ENSGALG00000040923 (−0.65, 0.68), SLC24A1 (−0.64, 0.63) | |
| K+ | Positive | Indianred4 | CEMIP (0.75, 0.92), FAM189A2 (0.74, 0.81), ELK3 (0.72, 0.76), TMEM204 (0.71, 0.94), ALPL (0.71, 0.86) |
| Negative | Antiquewhite2 | KCNMB4 (0.77, −0.79), HEATR6 (−0.75, 0.59), UBE2T (−0.73, 0.68), UNC93A (−0.69, 0.83), FLRT3 (−0.60, 0.51) |
| Trait | Correlation | Module Color | Gene Name (GS 1, MM 2) |
|---|---|---|---|
| Line | Positive | Paleturquoise | RDM1 (0.99, 0.95), CHIA (0.99,0.95), DPT (0.98,0.94), GALNT16 (0.98, 0.93), COMTD1 (0.97, 0.93) |
| Negative | Black | MRPL3 (−0.90, 0.87), HAAO (−0.88, 0.88), LPAR2 (0.86, −0.91), GNA11 (−0.85, 0.90), ENSGALG00000041512 (0.85, −0.77) | |
| Treatment | Positive | Bisque4 | FAM46A (0.92, 0.81), DTX3L (0.92, 0.81), ARNTL (0.91, 0.77), ENSGALG00000042001 (0.90, 0.79), MX1 (0.89, 0.78) |
| Negative | Lightgreen | PSMD1 (−0.89, 0.66), PNPLA7 (−0.89, 0.70), IFIH1 (0.83, −0.80), LCAT (−0.82, 0.84), HBA1 (−0.81, 0.82) | |
| pH | Positive | Black | COX7A2L (0.82, 0.88), CCDC25 (0.81, 0.86), ND4L (0.79, 0.91), RAB11A (0.79, 0.79), TIMM44 (0.79, 0.88) |
| Lightgreen | ENSGALG00000034218 (0.87, 0.90), COMMD9 (0.87, 0.88), ENSGALG00000042254 (0.85, 0.89), TPM3 (0.85, 0.86), ISCU (0.84, 0.87) | ||
| Negative | Darkloivergreen | PRIMPOL (−0.84, 0.79), HELB (−0.83, 0.92), ENSGALG00000045606 (−0.82, 0.86), INTS6 (−0.81, 0.88), PCF11 (−0.81, 0.90) | |
| HCO3 | Positive | Lightgreen | ZP4 (0.73, 0.55), HS6ST1 (0.72, 0.87), RBP3 (0.72, 0.57), HIST1H46 (0.72, 0.87), PCCB (0.70, 0.86) |
| Indianred4 | TNNC2 (0.68, 0.77), ZP2 (0.62, 0.56), ENSGALG00000026655 (0.61, 0.98), ENSGALG00000041238 (−0.61, −0.81), ENSGALG00000012416 (−0.61, −0.76) | ||
| Negative | Coral2 | TMEM154 (−0.66, 0.90), FAH (0.66, −0.93), MGARP (0.64, −0.92), AAMDC (0.61, −0.89), FANCL (0.61, −0.86) | |
| TCO2 | Positive | Indianred4 | TNNC2 (0.64, 0.77), ENSGALG00000026655 (0.62, 0.98), CCN3 (0.61, 0.69), ENSGALG00000029695 (0.60, 0.99), ST6GAL2 (0.58, 0.67) |
| Negative | Coral2 | PNPLA3 (−0.70, 0.82), TMEM154 (−0.69, 0.90), CYP1AC1 (0.65, −0.70), REEP2 (−0.63, 0.82), MANBAL4 (−0.62, 0.73) | |
| BE | Positive | Lightgreen | HS6ST1 (0.82, 0.87), HIST1H46 (0.80, 0.87), CPS1 (0.80, 0.85), SNX3 (0.80, 0.62), MALSU1 (0.80, 0.80) |
| Negative | Coral2 | MGARP (0.79, −0.92), AAMDC (0.77, −0.89), HADHA (0.76, −0.92), FBXL18 (−0.76, 0.80), REEP2 (−0.74, 0.82) | |
| Glu | Negative | Skyblue | INAVA (−0.69, 0.71), ENSGALG00000007007 (−0.69, 0.75), PTHLH (−0.66, 0.66), SLC12A5 (−0.64, 0.79), SINHCAF (−0.64, 0.60) |
| PO2 | Positive | Black | RAB31 (0.78, 0.60), GAN11 (0.71, 0.81), SEPTIN6 (0.71, 0.62), CRADD (0.69, 0.71), POLR3F (0.67, 0.76) |
| Negative | Paleturquoise | VAMP7 (0.89, −0.63), BMP2 (−0.87, 0.91), AP3B1 (0.85, −0.78), NLN (0.84, −0.83), C3orf33 (−0.84, 0.75) | |
| sO2 | Positive | Black | GAN11 (0.79, 0.90), SEPTIN6 (0.78, 0.75), RAB31 (0.77, 0.68), NEK4 (0.74, 0.90), ENSGALG00000037200 (0.72, 0.75) |
| Negative | Paleturquoise | VAMP7 (0.91, −0.63), MGAT4D (−0.85, 0.73), ABCA4 (−0.80, 0.71), ADAMTS3 (−0.79, 0.81), TRMT61B (−0.78, 0.80) | |
| Na+ | Positive | Plum1 | CNN1 (0.68, 0.70), GRK6 (0.66, 0.63), ENSGALG00000034399 (0.66, 0.87), BAHCC1 (0.65, 0.91), SLC7A3 (0.65, 0.78) |
| K+ | Positive | Skyblue | RF00569 (0.79, 0.65), CACNG2 (0.61, 0.77), SLC26A9 (0.57, 0.85), SCRT1 (0.55, 0.63), ZPLD1 (0.55, 0.52) |
| Treatment | Gene Name | Gene Description | log2 Fold Change |
|---|---|---|---|
| AH | GADD45G | Growth arrest and DNA damage inducible γ | 1.91 |
| TRPC3 | Transient receptor potential cation channel subfamily C member 3 | 1.8 | |
| GPRC5B | G protein-coupled receptor class C group 5 member B | 1.63 | |
| MSC | Musculin | 1.58 | |
| CEMIP | Cell migration inducing hyaluronidase 1 | 1.53 | |
| GPR182 | G protein-coupled receptor 182 | 1.48 | |
| ART7C | Erythroblast NAD--arginine ADP-ribosyltransferase-like | 1.43 | |
| ADAMTS15 | ADAM metallopeptidase with thrombospondin type 1 motif, 15 | 1.38 | |
| DNASE2B | Deoxyribonuclease 2 β | 1.36 | |
| LDLRAD3 | Low density lipoprotein receptor class A domain containing 3 | 1.31 | |
| SOCS1 | Suppressor of cytokine signaling 1 | −1.43 | |
| BAG3 | BCL2 associated athanogene 3 | −1.48 | |
| TTC7A | Tetratricopeptide repeat domain 7A | −1.52 | |
| DNAJA4 | DnaJ heat shock protein family (Hsp40) member A4 | −1.56 | |
| JMJD6 | Jumonji domain containing 6 | −1.57 | |
| HSPH1 | Heat shock protein family H (Hsp110) member 1 | −1.67 | |
| HSP90AA1 | Heat shock protein 90 α family class A member 1 | −1.96 | |
| HSPA2 | Heat shock 70kDa protein 2 | −2.25 | |
| HSPA5 | Heat shock 70kDa protein 5 (glucose-regulated protein, 78kDa) | −2.39 | |
| HSPA8 | Heat shock 70kDa protein 8 | −2.84 | |
| CH&NDV | NSUN7 | NOP2/Sun RNA methyltransferase family member 7 | 6.48 |
| HSPB2 | Heat shock protein family B (small) member 2 | 4.38 | |
| IFI6 | Interferon α inducible protein 6 | 3.62 | |
| OASL | 2’−5’-oligoadenylate synthetase like | 3.32 | |
| IFIT5 | Interferon induced protein with tetratricopeptide repeats 5 | 3.31 | |
| MX1 | MX dynamin like GTPase 1 | 3.28 | |
| ADGRG2 | Adhesion G protein-coupled receptor G2 | 3.24 | |
| CNN1 | Calponin 1 | 3.06 | |
| LRRC56 | Leucine rich repeat containing 56 | 2.8 | |
| CLCA1 | Chloride channel accessory 1 | 2.71 | |
| RPL3L | Ribosomal protein L3 like | −1.64 | |
| ABHD6 | Abhydrolase domain containing 6 | −1.73 | |
| BHLHE40 | Basic helix-loop-helix family member e40 | −1.75 | |
| CYP2C18 | Cytochrome P450 family 2 subfamily C member 18 | −1.77 | |
| EPB42 | Erythrocyte membrane protein band 4.2 | −1.82 | |
| ANGPTL4 | Angiopoietin like 4 | −1.93 | |
| LMBR1L | Limb development membrane protein 1 like | −2.31 | |
| CASP14 | Caspase 14, apoptosis-related cysteine peptidase | −2.34 | |
| ACOT12 | Acyl-CoA thioesterase 12 | −2.35 | |
| DIO2 | Iodothyronine deiodinase 2 | −3.71 |
| Treatment | Gene Name | Gene Description | log2 Fold Change |
|---|---|---|---|
| AH | CLRN2 | Clarin 2 | 6.18 |
| ZBTB16 | Zinc finger and BTB domain containing 16 | 3.46 | |
| TLX1 | T cell leukemia homeobox 1 | 2.56 | |
| GRIP2 | Glutamate receptor interacting protein 2 | 2.42 | |
| CCDC103 | Coiled-coil domain containing 103 | 2.18 | |
| MBOAT2 | Membrane bound O-acyltransferase domain containing 2 | 1.89 | |
| PPARGC1A | PPARG coactivator 1 α | 1.85 | |
| TMEM255A | Transmembrane protein 255A | 1.8 | |
| GPRC5B | G protein-coupled receptor class C group 5 member B | 1.78 | |
| HKDC1 | Hexokinase domain containing 1 | 1.76 | |
| PTPRD | Protein tyrosine phosphatase, receptor type D | −1.53 | |
| USP18 | Ubiquitin specific peptidase 18 | −1.64 | |
| TTC7A | Tetratricopeptide repeat domain 7A | −1.79 | |
| HSPA8 | Heat shock 70kDa protein 8 | −1.88 | |
| SCF14M2 | Solute carrier family 14 member 2 | −1.92 | |
| HSP90AA1 | Heat shock protein 90 α family class A member 1 | −1.96 | |
| FBN2 | Fibrillin 2 | −2.16 | |
| HSPA5 | Heat shock 70kDa protein 5 (glucose-regulated protein, 78kDa) | −2.3 | |
| BHLHA15 | Basic helix-loop-helix family member a15 | −3.12 | |
| PRRX1 | Paired related homeobox 1 | −3.2 | |
| CH&NDV | G0S2 | Gallus gallus G0/G1 switch 2 (G0S2), mRNA | 4.9 |
| LAMB3 | Laminin subunit β 3 | 4.48 | |
| NPAS2 | Gallus gallus neuronal PAS domain protein 2 (NPAS2), mRNA | 4.03 | |
| PLCH1 | Phospholipase C eta 1 | 3.11 | |
| TMEM255A | Transmembrane protein 255A | 3.02 | |
| MYBPC3 | Myosin binding protein C, cardiac | 2.97 | |
| PROK2 | Prokineticin 2 | 2.96 | |
| SCNN1B | Sodium channel epithelial 1 β subunit | 2.86 | |
| SERPINB10B | Gallus gallus serpin peptidase inhibitor, clade B, member 10 B | 2.84 | |
| PTX3 | Pentraxin 3 | 2.79 | |
| E3UPLT | E3 ubiquitin-protein ligase Topors-like | −2.85 | |
| PLCE1 | Phospholipase C epsilon 1 | −3.08 | |
| NKX2-3 | NK2 homeobox 3 | −3.41 | |
| APOV1 | Apovitellenin 1 | −3.53 | |
| DIO2 | Iodothyronine deiodinase 2 | −3.64 | |
| SPARCL1 | SPARC like 1 | −3.68 | |
| CHAC1 | ChaC glutathione specific γ-glutamylcyclotransferase 1 | −4.16 | |
| NOV | Nephroblastoma overexpressed (NOV), mRNA | −4.76 | |
| EDSC | Epidermal differentiation protein rich in serine and cysteine, mRNA | −5.19 | |
| NKX6-3 | NK6 homeobox 3 | −6.76 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Saelao, P.; Kern, C.; Jin, S.; Gallardo, R.A.; Kelly, T.; Dekkers, J.M.; Lamont, S.J.; Zhou, H. Liver Transcriptome Responses to Heat Stress and Newcastle Disease Virus Infection in Genetically Distinct Chicken Inbred Lines. Genes 2020, 11, 1067. https://doi.org/10.3390/genes11091067
Wang Y, Saelao P, Kern C, Jin S, Gallardo RA, Kelly T, Dekkers JM, Lamont SJ, Zhou H. Liver Transcriptome Responses to Heat Stress and Newcastle Disease Virus Infection in Genetically Distinct Chicken Inbred Lines. Genes. 2020; 11(9):1067. https://doi.org/10.3390/genes11091067
Chicago/Turabian StyleWang, Ying, Perot Saelao, Colin Kern, Sihua Jin, Rodrigo A. Gallardo, Terra Kelly, Jack M. Dekkers, Susan J. Lamont, and Huaijun Zhou. 2020. "Liver Transcriptome Responses to Heat Stress and Newcastle Disease Virus Infection in Genetically Distinct Chicken Inbred Lines" Genes 11, no. 9: 1067. https://doi.org/10.3390/genes11091067
APA StyleWang, Y., Saelao, P., Kern, C., Jin, S., Gallardo, R. A., Kelly, T., Dekkers, J. M., Lamont, S. J., & Zhou, H. (2020). Liver Transcriptome Responses to Heat Stress and Newcastle Disease Virus Infection in Genetically Distinct Chicken Inbred Lines. Genes, 11(9), 1067. https://doi.org/10.3390/genes11091067