Negative Regulation of Serine Threonine Kinase 11 (STK11) through miR-100 in Head and Neck Cancer

 ,
,  ,
,  , ,
, ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Samples
2.2. Nucleic Acid Isolation and Assessment
2.3. HPV Detection and Genotyping
2.4. Bisulfite Conversion and STK11 Gene Methylation
2.5. Real-Time PCR Analysis
2.6. Cell Culture and Transfection
2.7. Luciferase Reporter Assay
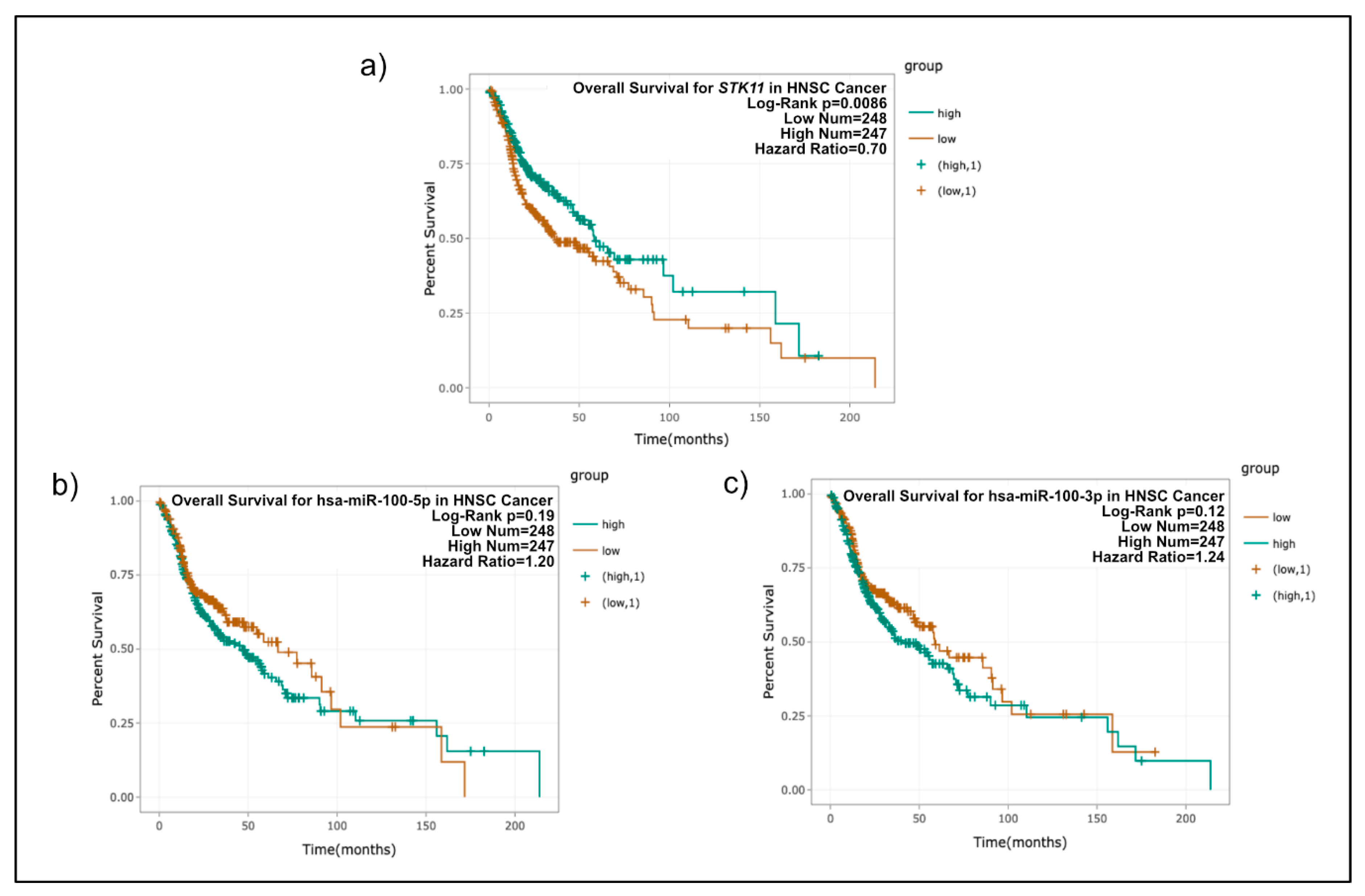
2.8. Patient Survival Analysis of STK11, miR-100-3p, and miR-100-5p
2.9. Statistical Analysis
3. Results
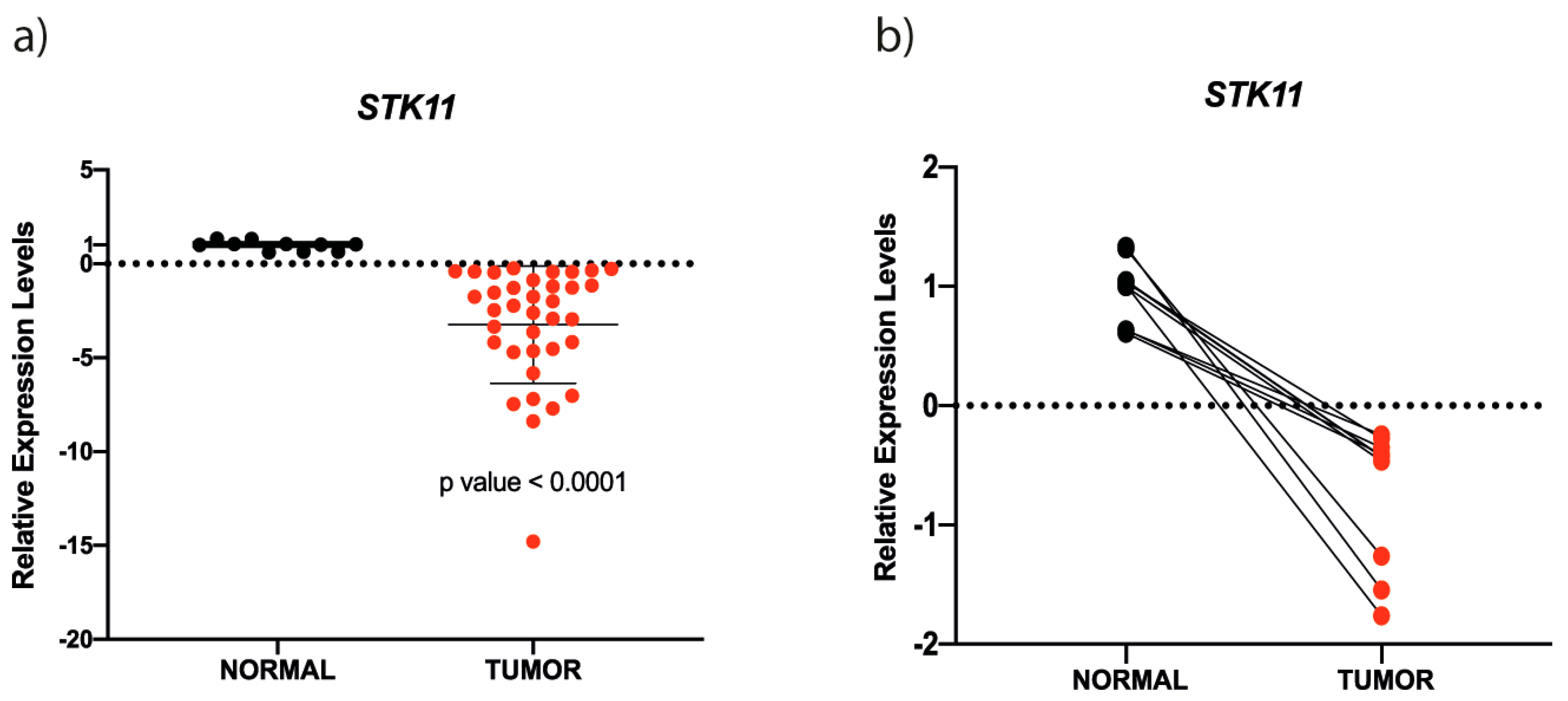
3.1. STK11 mRNA is Down-Regulated in HNC
3.2. STK11 Promoter Methylation Status
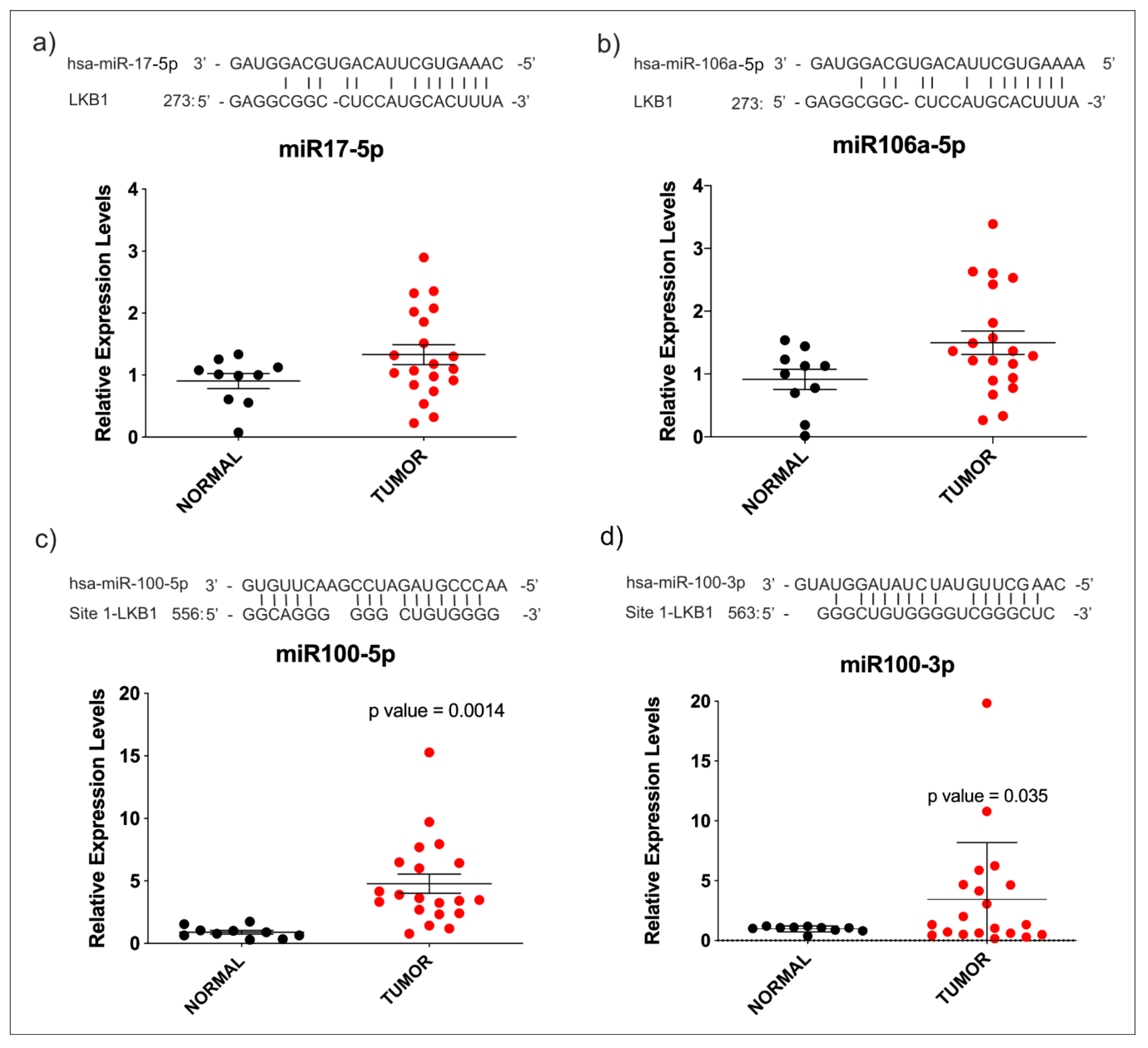
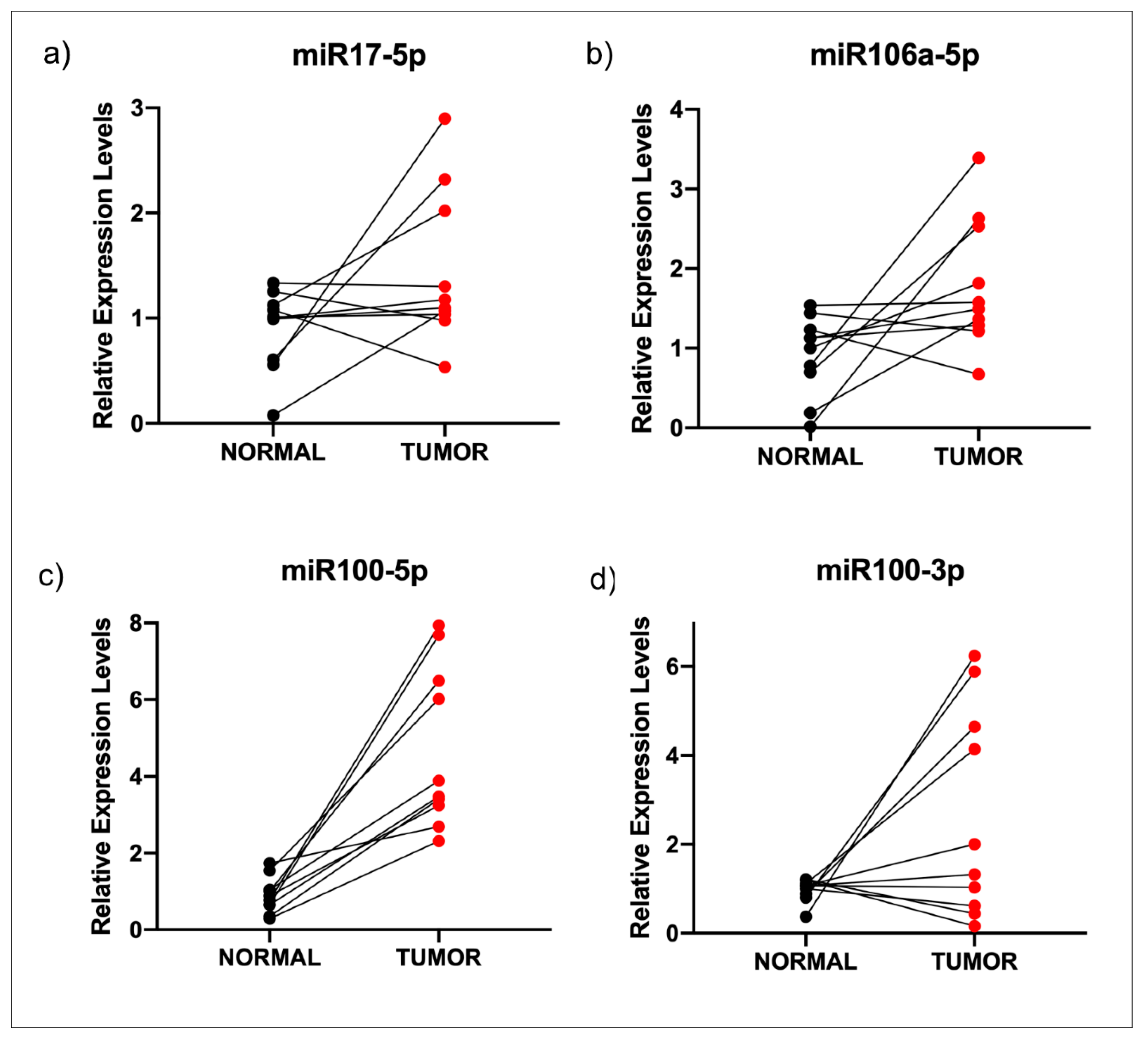
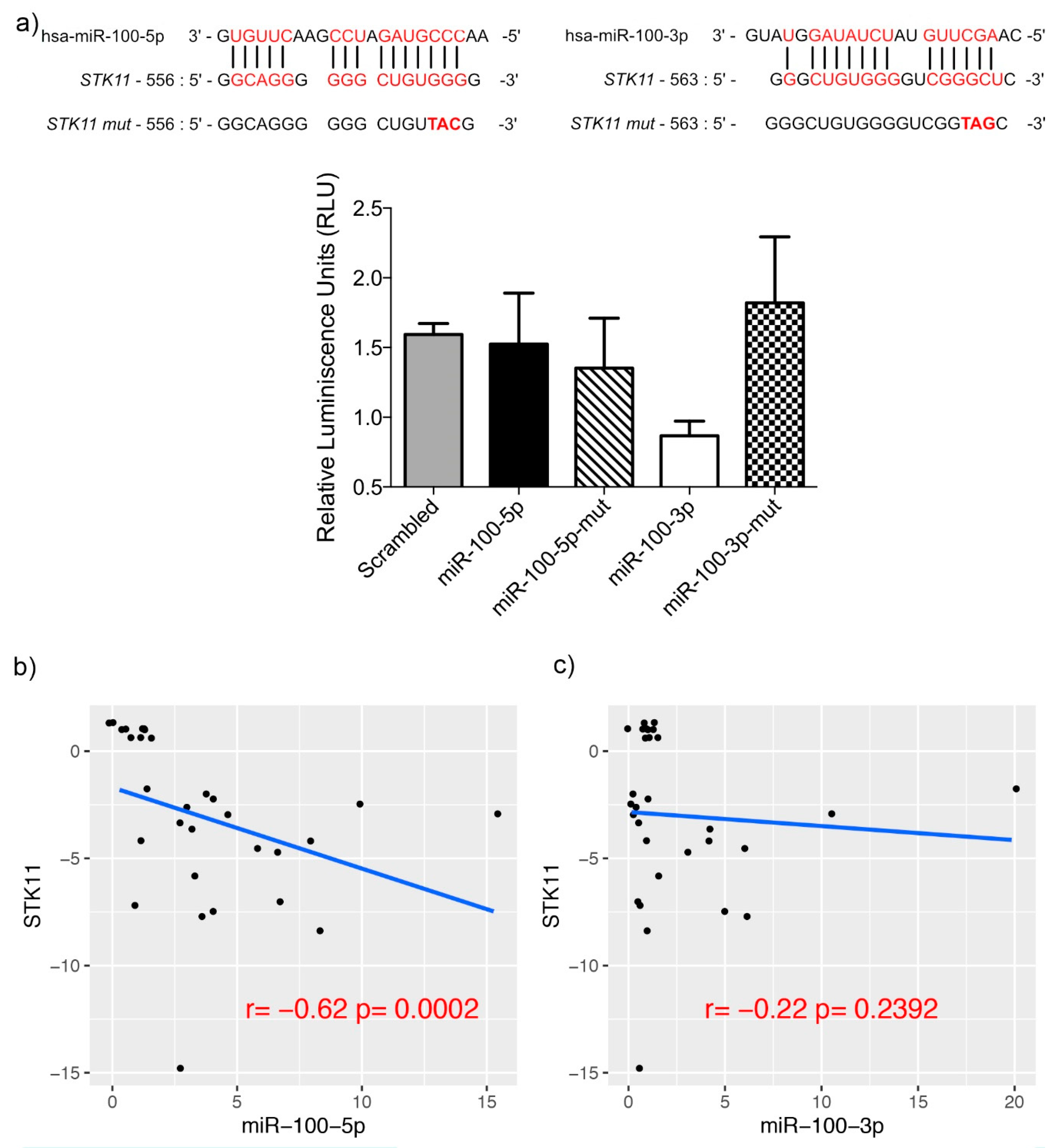
3.3. miR-100-3p Interacts with STK11 mRNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ferlay, J.; Shin, H.-R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Siegel, R.; Ward, E.; Murray, T.; Xu, J.; Smigal, C.; Thun, M.J. Cancer statistics, 2006. CA Cancer J. Clin. 2006, 56, 106–130. [Google Scholar] [CrossRef] [PubMed]
- Leemans, C.R.; Braakhuis, B.J.M.; Brakenhoff, R.H. The molecular biology of head and neck cancer. Nat. Rev. Cancer 2011, 11, 9–22. [Google Scholar] [CrossRef]
- Jethwa, A.R.; Khariwala, S.S. Tobacco-related carcinogenesis in head and neck cancer. Cancer Metastasis Rev. 2017, 36, 411–423. [Google Scholar] [CrossRef]
- Ragin, C.C.R.; Modugno, F.; Gollin, S.M. The epidemiology and risk factors of head and neck cancer: A focus on human papillomavirus. J. Dent. Res. 2007, 86, 104–114. [Google Scholar] [CrossRef]
- Gillison, M.L.; Koch, W.M.; Capone, R.B.; Spafford, M.; Westra, W.H.; Wu, L.; Zahurak, M.L.; Daniel, R.W.; Viglione, M.; Symer, D.E.; et al. Evidence for a causal association between human papillomavirus and a subset of head and neck cancers. J. Natl. Cancer Inst. 2000, 92, 709–720. [Google Scholar] [CrossRef]
- Puram, S.V.; Rocco, J.W. Molecular aspects of head and neck cancer therapy. Hematol. Oncol. Clin. N. Am. 2015, 29, 971–992. [Google Scholar] [CrossRef]
- Gary, C.; Hajek, M.; Biktasova, A.; Bellinger, G.; Yarbrough, W.G.; Issaeva, N. Selective antitumor activity of roscovitine in head and neck cancer. Oncotarget 2016, 7, 38598–38611. [Google Scholar] [CrossRef]
- Karuman, P.; Gozani, O.; Odze, R.D.; Zhou, X.C.; Zhu, H.; Shaw, R.; Brien, T.P.; Bozzuto, C.D.; Ooi, D.; Cantley, L.C.; et al. The Peutz-Jegher gene product LKB1 is a mediator of p53-dependent cell death. Mol. Cell 2001, 7, 1307–1319. [Google Scholar] [CrossRef]
- Hawley, S.A.; Boudeau, J.; Reid, J.L.; Mustard, K.J.; Udd, L.; Makela, T.P.; Alessi, D.R.; Hardie, D.G. Complexes between the LKB1 tumor suppressor, STRAD α/β and MO25 α/β are upstream kinases in the AMP-activated protein kinase cascade. J. Biol. 2003, 2, 28. [Google Scholar] [CrossRef]
- Thomson, D.M.; Brown, J.D.; Fillmore, N.; Condon, B.M.; Kim, H.-J.; Barrow, J.R.; Winder, W.W. LKB1 and the regulation of malonyl-CoA and fatty acid oxidation in muscle. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1572–E1579. [Google Scholar] [CrossRef] [PubMed]
- Tiainen, M.; Ylikorkala, A.; Makela, T.P. Growth suppression by Lkb1 is mediated by a G(1) cell cycle arrest. Proc. Natl. Acad. Sci. USA 1999, 96, 9248–9251. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Huang, D.; Lu, N.; Luo, L. Role of the LKB1/AMPK pathway in tumor invasion and metastasis of cancer cells (Review). Oncol. Rep. 2015, 34, 2821–2826. [Google Scholar] [CrossRef] [PubMed]
- Mehenni, H.; Gehrig, C.; Nezu, J.; Oku, A.; Shimane, M.; Rossier, C.; Guex, N.; Blouin, J.L.; Scott, H.S.; Antonarakis, S.E. Loss of LKB1 kinase activity in Peutz-Jeghers syndrome, and evidence for allelic and locus heterogeneity. Am. J. Hum. Genet. 1998, 63, 1641–1650. [Google Scholar] [CrossRef]
- Avizienyte, E.; Loukola, A.; Roth, S.; Hemminki, A.; Tarkkanen, M.; Salovaara, R.; Arola, J.; Butzow, R.; Husgafvel-Pursiainen, K.; Kokkola, A.; et al. LKB1 somatic mutations in sporadic tumors. Am. J. Pathol. 1999, 154, 677–681. [Google Scholar] [CrossRef]
- Sanchez-Cespedes, M.; Parrella, P.; Esteller, M.; Nomoto, S.; Trink, B.; Engles, J.M.; Westra, W.H.; Herman, J.G.; Sidransky, D. Inactivation of LKB1/STK11 is a common event in adenocarcinomas of the lung. Cancer Res. 2002, 62, 3659–3662. [Google Scholar]
- Bignell, G.R.; Barfoot, R.; Seal, S.; Collins, N.; Warren, W.; Stratton, M.R. Low frequency of somatic mutations in the LKB1/Peutz-Jeghers syndrome gene in sporadic breast cancer. Cancer Res. 1998, 58, 1384–1386. [Google Scholar]
- Nakanishi, C.; Yamaguchi, T.; Iijima, T.; Saji, S.; Toi, M.; Mori, T.; Miyaki, M. Germline mutation of the LKB1/STK11 gene with loss of the normal allele in an aggressive breast cancer of Peutz-Jeghers syndrome. Oncology 2004, 67, 476–479. [Google Scholar] [CrossRef]
- Petersen, G.M. Familial pancreatic cancer. Semin. Oncol. 2016, 43, 548–553. [Google Scholar] [CrossRef]
- Su, G.H.; Hruban, R.H.; Bansal, R.K.; Bova, G.S.; Tang, D.J.; Shekher, M.C.; Westerman, A.M.; Entius, M.M.; Goggins, M.; Yeo, C.J.; et al. Germline and somatic mutations of the STK11/LKB1 Peutz-Jeghers gene in pancreatic and biliary cancers. Am. J. Pathol. 1999, 154, 1835–1840. [Google Scholar] [CrossRef]
- Guldberg, P.; thor Straten, P.; Ahrenkiel, V.; Seremet, T.; Kirkin, A.F.; Zeuthen, J. Somatic mutation of the Peutz-Jeghers syndrome gene, LKB1/STK11, in malignant melanoma. Oncogene 1999, 18, 1777–1780. [Google Scholar] [CrossRef] [PubMed]
- Rowan, A.; Bataille, V.; MacKie, R.; Healy, E.; Bicknell, D.; Bodmer, W.; Tomlinson, I. Somatic mutations in the Peutz-Jeghers (LKB1/STKII) gene in sporadic malignant melanomas. J. Investig. Dermatol. 1999, 112, 509–511. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.J.; Cho, Y.G.; Park, J.Y.; Kim, T.Y.; Lee, J.H.; Kim, H.S.; Lee, J.W.; Song, Y.H.; Nam, S.W.; Lee, S.H.; et al. Genetic analysis of the LKB1/STK11 gene in hepatocellular carcinomas. Eur. J. Cancer 2004, 40, 136–141. [Google Scholar] [CrossRef]
- Qiu, W.; Schonleben, F.; Thaker, H.M.; Goggins, M.; Su, G.H. A novel mutation of STK11/LKB1 gene leads to the loss of cell growth inhibition in head and neck squamous cell carcinoma. Oncogene 2006, 25, 2937–2942. [Google Scholar] [CrossRef] [PubMed]
- Zeng, P.-Y.; Berger, S.L. LKB1 is recruited to the p21/WAF1 promoter by p53 to mediate transcriptional activation. Cancer Res. 2006, 66, 10701–10708. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Choi, J.E.; Na, Y.K.; Lee, E.J.; Lee, W.K.; Choi, Y.Y.; Yoon, G.S.; Jeon, H.-S.; Kim, D.S.; Park, J.Y. Genetic and epigenetic alterations of the LKB1 gene and their associations with mutations in TP53 and EGFR pathway genes in Korean non-small cell lung cancers. Lung Cancer 2013, 81, 194–199. [Google Scholar] [CrossRef]
- Kullmann, L.; Krahn, M.P. Controlling the master-upstream regulation of the tumor suppressor LKB1. Oncogene 2018, 37, 3045–3057. [Google Scholar] [CrossRef]
- Zheng, F.; Yuan, X.; Chen, E.; Ye, Y.; Li, X.; Dai, Y. Methylation of STK11 promoter is a risk factor for tumor stage and survival in clear cell renal cell carcinoma. Oncol. Lett. 2017, 14, 3065–3070. [Google Scholar] [CrossRef]
- Trojan, J.; Brieger, A.; Raedle, J.; Esteller, M.; Zeuzem, S. 5’-CpG island methylation of the LKB1/STK11 promoter and allelic loss at chromosome 19p13.3 in sporadic colorectal cancer. Gut 2000, 47, 272–276. [Google Scholar] [CrossRef]
- Lee, C.G.; Kim, Y.W.; Kim, E.H.; Meng, Z.; Huang, W.; Hwang, S.J.; Kim, S.G. Farnesoid X receptor protects hepatocytes from injury by repressing miR-199a-3p, which increases levels of LKB1. Gastroenterology 2012, 142, 1206–1217.e7. [Google Scholar] [CrossRef]
- Chen, H.; Untiveros, G.M.; McKee, L.A.K.; Perez, J.; Li, J.; Antin, P.B.; Konhilas, J.P. Micro-RNA-195 and -451 regulate the LKB1/AMPK signaling axis by targeting MO25. PLoS ONE 2012, 7, e41574. [Google Scholar] [CrossRef] [PubMed]
- Lao, G.; Liu, P.; Wu, Q.; Zhang, W.; Liu, Y.; Yang, L.; Ma, C. Mir-155 promotes cervical cancer cell proliferation through suppression of its target gene LKB1. Tumour. Biol. 2014, 35, 11933–11938. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.-N.; Jiang, M.-J.; Mei, Z.; Dai, J.-J.; Dai, C.-Y.; Fang, C.; Huang, Q.; Tian, L. microRNA-7 impairs autophagy-derived pools of glucose to suppress pancreatic cancer progression. Cancer Lett. 2017, 400, 69–78. [Google Scholar] [CrossRef]
- Sotlar, K.; Diemer, D.; Dethleffs, A.; Hack, Y.; Stubner, A.; Vollmer, N.; Menton, S.; Menton, M.; Dietz, K.; Wallwiener, D.; et al. Detection and typing of human papillomavirus by e6 nested multiplex PCR. J. Clin. Microbiol. 2004, 42, 3176–3184. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 18, 1427–1431. [Google Scholar] [CrossRef]
- Dweep, H.; Gretz, N. MiRWalk2.0: A comprehensive atlas of microRNA-target interactions. Nat. Methods 2015, 12, 697. [Google Scholar] [CrossRef]
- Krü, J.; Rehmsmeier, M. RNAhybrid: Microrna target prediction easy, fast and flexible. Nucleic Acids Res. 2006. [Google Scholar] [CrossRef]
- Lawrence, M.S.; Sougnez, C.; Lichtenstein, L.; Cibulskis, K.; Lander, E.; Gabriel, S.B.; Getz, G.; Ally, A.; Balasundaram, M.; Birol, I.; et al. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2015, 44, 71. [Google Scholar] [CrossRef]
- Walboomers, J.M.; Jacobs, M.V.; Manos, M.M.; Bosch, F.X.; Kummer, J.A.; Shah, K.V.; Snijders, P.J.; Peto, J.; Meijer, C.J.; Munoz, N. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J. Pathol. 1999, 189, 12–19. [Google Scholar] [CrossRef]
- Khode, S.R.; Dwivedi, R.C.; Rhys-Evans, P.; Kazi, R. Exploring the link between human papilloma virus and oral and oropharyngeal cancers. J. Cancer Res. Ther. 2014, 10, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Paz, I.B.; Cook, N.; Odom-Maryon, T.; Xie, Y.; Wilczynski, S.P. Human papillomavirus (HPV) in head and neck cancer. An association of HPV 16 with squamous cell carcinoma of Waldeyer’s tonsillar ring. Cancer 1997, 79, 595–604. [Google Scholar] [CrossRef]
- Haraf, D.J.; Nodzenski, E.; Brachman, D.; Mick, R.; Montag, A.; Graves, D.; Vokes, E.E.; Weichselbaum, R.R. Human papilloma virus and p53 in head and neck cancer: Clinical correlates and survival. Clin. Cancer Res. 1996, 2, 755–762. [Google Scholar] [PubMed]
- Schwartz, S.M.; Daling, J.R.; Doody, D.R.; Wipf, G.C.; Carter, J.J.; Madeleine, M.M.; Mao, E.J.; Fitzgibbons, E.D.; Huang, S.; Beckmann, A.M.; et al. Oral cancer risk in relation to sexual history and evidence of human papillomavirus infection. J. Natl. Cancer Inst. 1998, 90, 1626–1636. [Google Scholar] [CrossRef]
- Fouret, P.; Monceaux, G.; Temam, S.; Lacourreye, L.; St Guily, J.L. Human papillomavirus in head and neck squamous cell carcinomas in nonsmokers. Arch. Otolaryngol. Head Neck Surg. 1997, 123, 513–516. [Google Scholar] [CrossRef]
- Gillison, M.L.; Castellsague, X.; Chaturvedi, A.; Goodman, M.T.; Snijders, P.; Tommasino, M.; Arbyn, M.; Franceschi, S. Eurogin Roadmap: Comparative epidemiology of HPV infection and associated cancers of the head and neck and cervix. Int. J. Cancer 2014, 134, 497–507. [Google Scholar] [CrossRef]
- Kreimer, A.R.; Clifford, G.M.; Boyle, P.; Franceschi, S. Human papillomavirus types in head and neck squamous cell carcinomas worldwide: A systematic review. Cancer Epidemiol. Biomark. Prev. 2005, 14, 467–475. [Google Scholar] [CrossRef]
- Resta, N.; Pierannunzio, D.; Lenato, G.M.; Stella, A.; Capocaccia, R.; Bagnulo, R.; Lastella, P.; Susca, F.C.; Bozzao, C.; Loconte, D.C.; et al. Cancer risk associated with STK11/LKB1 germline mutations in Peutz-Jeghers syndrome patients: Results of an Italian multicenter study. Dig. Liver Dis. 2013, 45, 606–611. [Google Scholar] [CrossRef]
- Rowan, A.; Churchman, M.; Jefferey, R.; Hanby, A.; Poulsom, R.; Tomlinson, I. In situ analysis of LKB1/STK11 mRNA expression in human normal tissues and tumours. J. Pathol. 2000, 192, 203–206. [Google Scholar] [CrossRef]
- Ekizoglu, S.; Dalay, N.; Karaman, E.; Akdeniz, D.; Ozaydin, A.; Buyru, N. LKB1 downregulation may be independent of promoter methylation or FOXO3 expression in head and neck cancer. Transl. Res. 2013, 162, 122–129. [Google Scholar] [CrossRef]
- Dai, W.; Teodoridis, J.M.; Zeller, C.; Graham, J.; Hersey, J.; Flanagan, J.M.; Stronach, E.; Millan, D.W.; Siddiqui, N.; Paul, J.; et al. Systematic CpG islands methylation profiling of genes in the wnt pathway in epithelial ovarian cancer identifies biomarkers of progression-free survival. Clin. Cancer Res. 2011, 17, 4052–4062. [Google Scholar] [CrossRef] [PubMed]
- Claus, R.; Wilop, S.; Hielscher, T.; Sonnet, M.; Dahl, E.; Galm, O.; Jost, E.; Plass, C. A systematic comparison of quantitative high-resolution DNA methylation analysis and methylation-specific PCR. Epigenetics 2012, 7, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Summers, T.; Langan, R.C.; Nissan, A.; Brucher, B.L.D.M.; Bilchik, A.J.; Protic, M.; Daumer, M.; Avital, I.; Stojadinovic, A. Serum-based DNA methylation biomarkers in colorectal cancer: Potential for screening and early detection. J. Cancer 2013, 4, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, T.; Sheng, Y.; Zhang, C.; Peng, Y.; Wang, X.; Zhang, C. Methylation Profiling of Multiple Tumor Suppressor Genes in Hepatocellular Carcinoma and the Epigenetic Mechanism of 3OST2 Regulation. J. Cancer 2015, 6, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Sartor, M.A.; Dolinoy, D.C.; Jones, T.R.; Colacino, J.A.; Prince, M.E.P.; Carey, T.E.; Rozek, L.S. Genome-wide methylation and expression differences in HPV(+) and HPV(-) squamous cell carcinoma cell lines are consistent with divergent mechanisms of carcinogenesis. Epigenetics 2011, 6, 777–787. [Google Scholar] [CrossRef]
- Henson, B.J.; Bhattacharjee, S.; O’Dee, D.M.; Feingold, E.; Gollin, S.M. Decreased expression of miR-125b and miR-100 in oral cancer cells contributes to malignancy. Genes Chromosomes Cancer 2009, 48, 569–582. [Google Scholar] [CrossRef]
- Peng, D.-X.; Luo, M.; Qiu, L.-W.; He, Y.-L.; Wang, X.-F. Prognostic implications of microRNA-100 and its functional roles in human epithelial ovarian cancer. Oncol. Rep. 2012, 27, 1238–1244. [Google Scholar] [CrossRef]
- Xu, C.; Zeng, Q.; Xu, W.; Jiao, L.; Chen, Y.; Zhang, Z.; Wu, C.; Jin, T.; Pan, A.; Wei, R.; et al. miRNA-100 inhibits human bladder urothelial carcinogenesis by directly targeting mTOR. Mol. Cancer Ther. 2013, 12, 207–219. [Google Scholar] [CrossRef]
- Ge, Y.-Y.; Shi, Q.; Zheng, Z.-Y.; Gong, J.; Zeng, C.; Yang, J.; Zhuang, S.-M. MicroRNA-100 promotes the autophagy of hepatocellular carcinoma cells by inhibiting the expression of mTOR and IGF-1R. Oncotarget 2014, 5, 6218–6228. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer/Cocktail | HPV Genotype | Amplicon (bp) | Sequence (5′-3′) |
|---|---|---|---|
| GP-E7-5B | CTG AGC TGT CAR NTA ATT GCT CA | ||
| GP-E6-3F | 630 | GGG WGK KAC TGA AAT CGG T | |
| GP-E7-6B | TCC TCT GAG TYG YCP AAT TGC TC | ||
| Cocktail I | 16 | 457 | Forward CAC AGT TATGCA CAG AGC TGC Reverse CAT ATA TTC ATG CAA TGT AGG TGT A |
| 18 | 322 | Forward CAC TTC ACT GCA AGA CAT AGA Reverse GTT GTG AAA TCGTCGTTT TTC A | |
| 31 | 263 | Forward GAA ATT GCA TGA ACT AAG CTC G Reverse CAC ATA TAC CTT TGTTTG TCA A | |
| 59 | 215 | Forward CAA AGG GGA ACT GCA AGA AAG Reverse TAT AAC AGC GTA TCA GCA GC | |
| 45 | 151 | Forward GTG GAA AAG TGC ATT ACA GG Reverse ACC TCT GTG GGT CCC AAT GT | |
| Cocktail II | 33 | 398 | Forward ACT ATA CAC AAC ATT GAA CTA Reverse GTT TTT ACA CGT CAC AGT GCA |
| 6/11 | 334 | Forward TGC AAG AAT GCA CTG ACC AC Reverse TGC ATG TTG TCC AGC AGT GT | |
| 58 | 274 | Forward GTA AAG TGT GCT TAC GAT TGC Reverse GTTGTTACA GGT TAC ACT TGT | |
| 52 | 229 | Forward TAA GGC TCG AGT GTG TGC AG Reverse CTAATA GTT ATT TCA CTT AAT GGT |
| Clinical Parameters | Patients n = 59 (100%) | STK11 Promoter Methylation n = 59 | STK11 Expression n = 36 | miRNA Expression n = 20 |
|---|---|---|---|---|
| Gender | ||||
| Male | 40 (67.8%) | |||
| Female | 19 (32.2%) | |||
| Age | ||||
| <50 | 10 (16.9%) | |||
| ≥50 | 49 (83.1%) | |||
| Clinical stage | ||||
| II | 5 (8.5%) | 5 (8.5%) | 4 (11.1%) | 2 (10%) |
| III | 11 (18.6%) | 11, 3a (18.6%) | 6, 3a (16.7%) | 4, 3a (20%) |
| IVA | 28 (47.5%) | 28, 6a (47.5%) | 15, 6a (41.7%) | 8, 6a (40%) |
| IVB | 10 (16.9%) | 10 (16.9%) | 8 (22.2%) | 4 (20%) |
| IVC | 5 (8.5%) | 5, 1a (8.5%) | 3, 1a (8.3%) | 2, 1a (10%) |
| Anatomic region | ||||
| Lip and oral cavity | 50 (84.7%) | |||
| Larynx | 1 (1.7%) | |||
| Pharynx | 7 (11.9%) | |||
| Nasal cavity | 1 (1.7%) | |||
| Histologic grade | ||||
| Low | 10 (16.9%) | |||
| Moderate | 48 (81.4%) | |||
| High | 1 (1.7%) | |||
| High-risk HPV | ||||
| 16 | 4 (6.8%) | |||
| Negative for HPV | 55 (93.2%) | |||
| Smoking Habit | ||||
| Positive | 34 (57.6%) | |||
| Negative | 25 (42.4%) | |||
| Alcoholism | ||||
| Positive | 34 (57.6%) | |||
| Negative | 25 (42.4%) | |||
| Tumor size | ||||
| >5 cm | 29 (49.2%) | |||
| <5 cm | 30 (50.8%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueroa-González, G.; Carrillo-Hernández, J.F.; Perez-Rodriguez, I.; Cantú de León, D.; Campos-Parra, A.D.; Martínez-Gutiérrez, A.D.; Coronel-Hernández, J.; García-Castillo, V.; López-Camarillo, C.; Peralta-Zaragoza, O.; et al. Negative Regulation of Serine Threonine Kinase 11 (STK11) through miR-100 in Head and Neck Cancer. Genes 2020, 11, 1058. https://doi.org/10.3390/genes11091058
Figueroa-González G, Carrillo-Hernández JF, Perez-Rodriguez I, Cantú de León D, Campos-Parra AD, Martínez-Gutiérrez AD, Coronel-Hernández J, García-Castillo V, López-Camarillo C, Peralta-Zaragoza O, et al. Negative Regulation of Serine Threonine Kinase 11 (STK11) through miR-100 in Head and Neck Cancer. Genes. 2020; 11(9):1058. https://doi.org/10.3390/genes11091058
Chicago/Turabian StyleFigueroa-González, Gabriela, José F. Carrillo-Hernández, Itzel Perez-Rodriguez, David Cantú de León, Alma D. Campos-Parra, Antonio D. Martínez-Gutiérrez, Jossimar Coronel-Hernández, Verónica García-Castillo, César López-Camarillo, Oscar Peralta-Zaragoza, and et al. 2020. "Negative Regulation of Serine Threonine Kinase 11 (STK11) through miR-100 in Head and Neck Cancer" Genes 11, no. 9: 1058. https://doi.org/10.3390/genes11091058
APA StyleFigueroa-González, G., Carrillo-Hernández, J. F., Perez-Rodriguez, I., Cantú de León, D., Campos-Parra, A. D., Martínez-Gutiérrez, A. D., Coronel-Hernández, J., García-Castillo, V., López-Camarillo, C., Peralta-Zaragoza, O., Jacobo-Herrera, N. J., Guardado-Estrada, M., & Pérez-Plasencia, C. (2020). Negative Regulation of Serine Threonine Kinase 11 (STK11) through miR-100 in Head and Neck Cancer. Genes, 11(9), 1058. https://doi.org/10.3390/genes11091058