Deciphering the Impact of a Bacterial Infection on Meiotic Recombination in Arabidopsis with Fluorescence Tagged Lines


Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of Arabidopsis NLR Genes
2.2. Arabidopsis Fluorescent Tagged Lines
2.3. Plant Growth Conditions
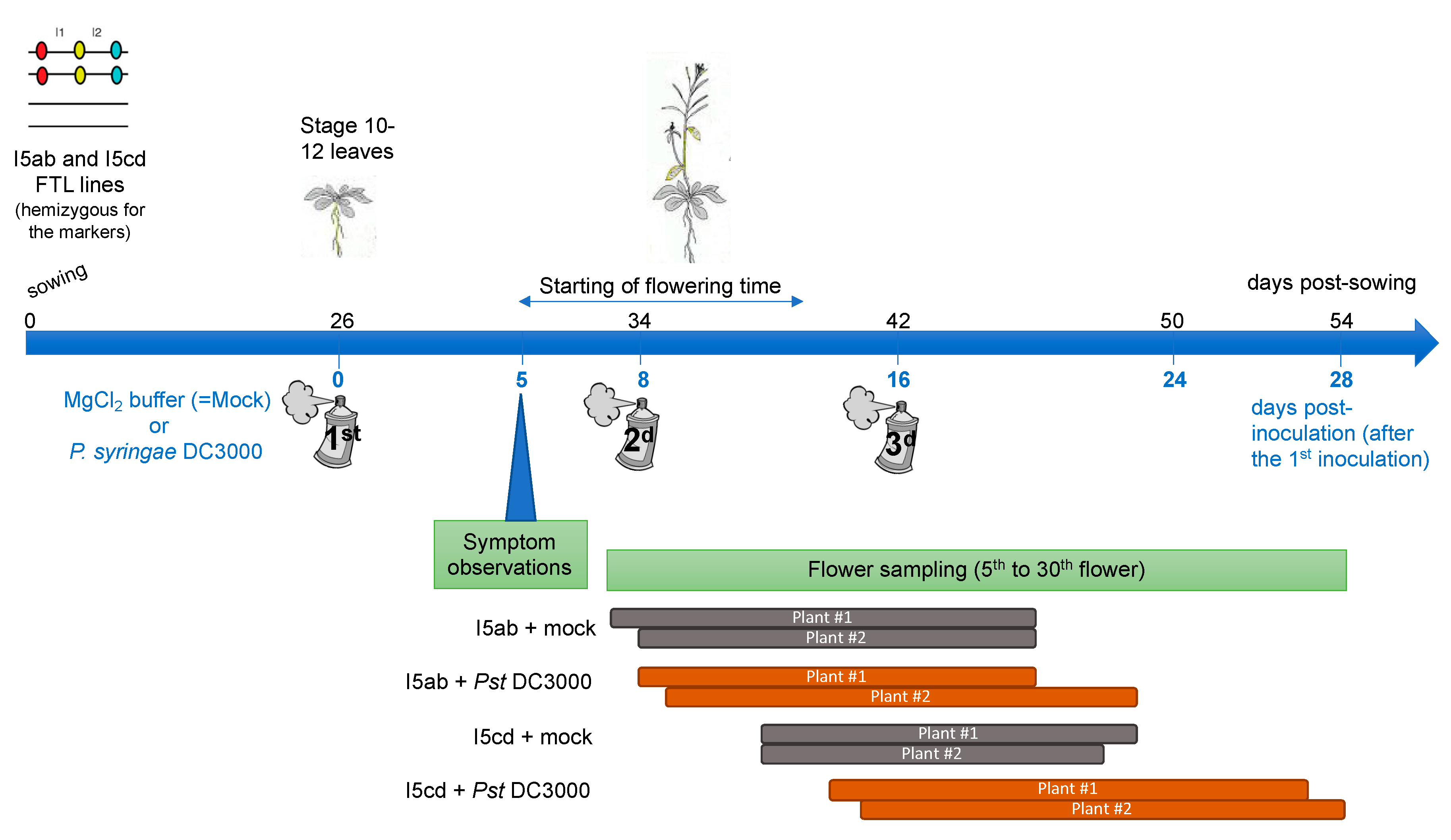
2.4. Inoculation Assay with Pseudomonas syringae
2.5. Analysis of Pollen Tetrads of Fluorescent Tagged Lines
3. Results
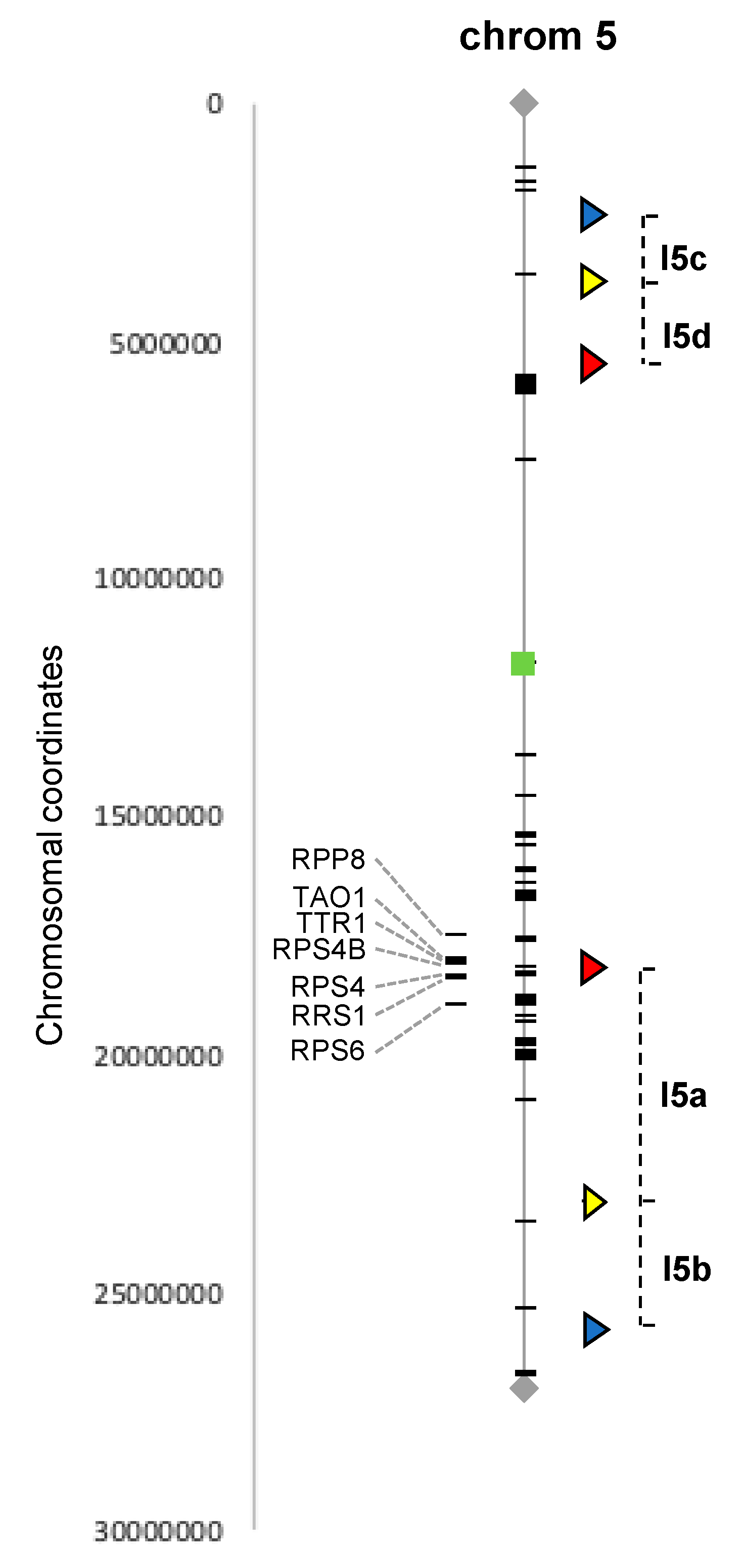
3.1. Selection of the Target Regions Based on the Available Arabidopsis FTL Lines
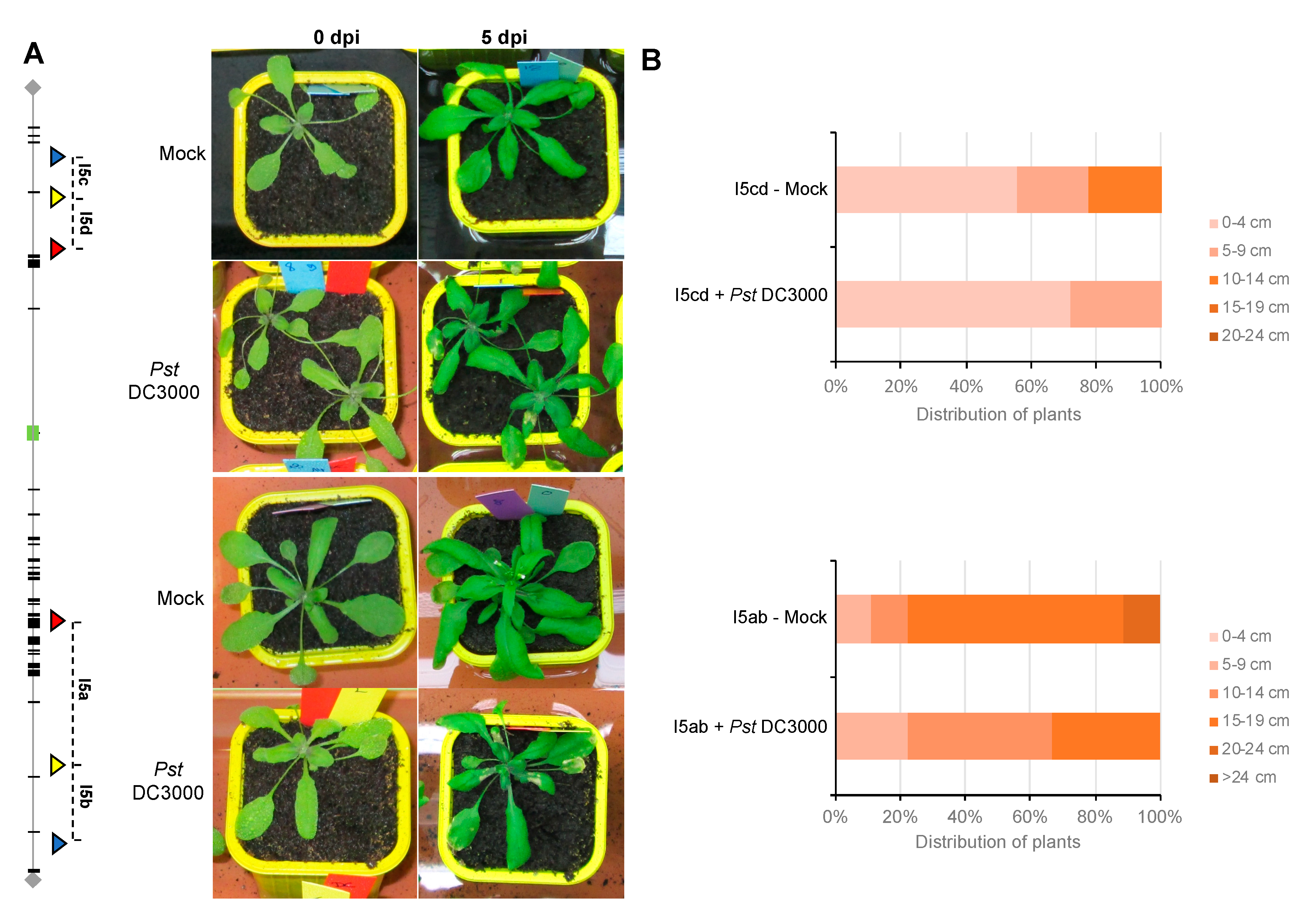
3.2. Floral Stem Development is Slightly Delayed in Inoculated Plants
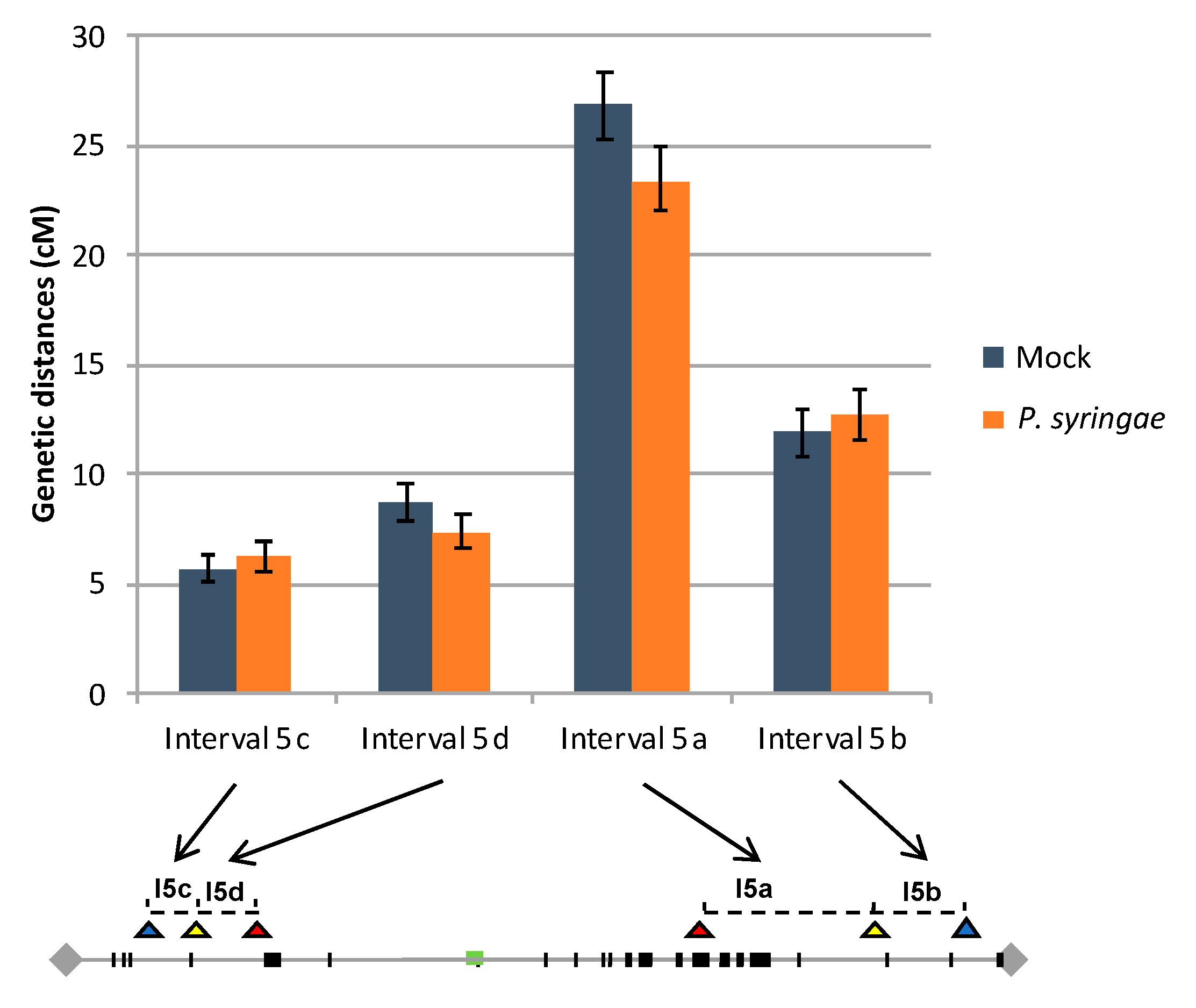
3.3. Meiotic Recombination May Not Be Affected in Inoculated Plants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Baggs, E.; Dagdas, G.; Krasileva, K.V. NLR diversity, helpers and integrated domains: Making sense of the NLR IDentity. Curr. Opin. Plant Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Michelmore, R.W.; Christopoulou, M.; Caldwell, K.S. Impacts of Resistance Gene Genetics, Function, and Evolution on a Durable Future. Annu. Rev. Phytopathol. 2013, 51, 291–319. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Vance, R.E.; Dangl, J.L. Intracellular innate immune surveillance devices in plants and animals. American Association for the Advancement of Science. Science 2016. [Google Scholar] [CrossRef] [PubMed]
- Meyers, B.C.; Kaushik, S.; Nandety, R.S. Evolving disease resistance genes. Curr. Opin. Plant Biol. 2005, 8, 129–134. [Google Scholar] [CrossRef]
- Friedman, A.R.; Baker, B.J. The evolution of resistance genes in multi-protein plant resistance systems. Curr. Opin. Genet. Dev. 2007, 17, 493–499. [Google Scholar] [CrossRef]
- Krasileva, K.V. The role of transposable elements and DNA damage repair mechanisms in gene duplications and gene fusions in plant genomes. Curr. Opin. Plant Biol. 2019, 48, 18–25. [Google Scholar] [CrossRef]
- David, P.; Chen, N.W.G.; Pedrosaharand, A.; Thareau, V.; Sevignac, M.; Cannon, S.B.; Debouck, D.; Langin, T.; Geffroy, V. A Nomadic Subtelomeric Disease Resistance Gene Cluster in Common Bean. Plant Physiol. 2009, 151, 1048–1065. [Google Scholar] [CrossRef]
- Chen, N.W.G.; Thareau, V.; Ribeiro, T.; Magdelenat, G.; Ashfield, T.; Innes, R.W.; Pedrosaharand, A.; Geffroy, V. Common Bean Subtelomeres Are Hot Spots of Recombination and Favor Resistance Gene Evolution. Front. Plant Sci. 2018, 9, 1185. [Google Scholar] [CrossRef]
- Richter, T.E.; Pryor, T.J.; Bennetzen, J.L.; Hulbert, S.H. New rust resistance specificities associated with recombination in the Rp1 complex in maize. Genetics 1995, 141, 373–381. [Google Scholar] [PubMed]
- Parniske, M.; Hammondkosack, K.E.; Golstein, C.; Thomas, C.M.; Jones, D.A.; Harrison, K.; Wulff, B.B.H.; Jones, J.D.G. Novel disease resistance specificities result from sequence exchange between tandemly repeated genes at the Cf-4/9 locus of tomato. Cell 1997, 91, 821–832. [Google Scholar] [CrossRef]
- Mcdowell, J.M.; Dhandaydham, M.; Long, T.A.; Aarts, M.G.M.; Goff, S.A.; Holub, E.B.; Dangl, J.L. Intragenic recombination and diversifying selection contribute to the evolution of downy mildew resistance at the RPP8 locus of Arabidopsis. Plant Cell 1998, 10, 1861–1874. [Google Scholar] [CrossRef] [PubMed]
- Noel, L.D.; Moores, T.L.; van der Biezen, E.A.; Parniske, M.; Daniels, M.J.; Parker, J.E.; Jones, J.D.G. Pronounced intraspecific haplotype divergence at the RPP5 complex disease resistance locus of Arabidopsis. Plant Cell 1999, 11, 2099–2111. [Google Scholar] [CrossRef]
- McDowell, J.M.; Simon, S.A. Recent insights into R gene evolution. Mol. Plant Pathol. 2006, 7, 437–448. [Google Scholar] [CrossRef]
- Geffroy, V.; Macadre, C.; David, P.; Pedrosa-Harand, A.; Sevignac, M.; Dauga, C.; Langin, T. Molecular analysis of a large subtelomeric nucleotide-binding-site-leucine-rich-repeat family in two representative genotypes of the major gene pools of Phaseolus vulgaris. Genetics 2009, 181, 405–419. [Google Scholar] [CrossRef]
- Chin, D.B.; Arroyo-Garcia, R.; Ochoa, O.E.; Kesseli, R.V.; Lavelle, D.O.; Richelmore, R.W. Recombination and spontaneous mutation at the major cluster of resistance genes in lettuce (Lactuca sativa). Genetics 2001, 157, 831–849. [Google Scholar]
- Choi, K.; Reinhard, C.; Serra, H.; Ziolkowski, P.; Underwood, C.J.; Zhao, X.; Hardcastle, T.J.; Yelina, N.E.; Griffin, C.H.; Jackson, M.A.; et al. Recombination Rate Heterogeneity within Arabidopsis Disease Resistance Genes. PLoS Genet. 2016, 12, e1006179. [Google Scholar] [CrossRef]
- Molinier, J.; Ries, G.; Zipfel, C.; Hohn, B. Transgeneration memory of stress in plants. Nature 2006, 442, 1046–1049. [Google Scholar] [CrossRef]
- Lucht, J.M.; Mauch-Mani, B.; Steiner, H.Y.; Metraux, J.P.; Ryals, J.; Hohn, B. Pathogen stress increases somatic recombination frequency in Arabidopsis. Nat. Genet. 2002, 30, 311–314. [Google Scholar] [CrossRef]
- Kovalchuk, I.; Kovalchuk, O.; Kalck, V.; Boyko, V.; Filkowski, J.; Heinlein, M.; Hohn, B. Pathogen-induced systemic plant signal triggers DNA rearrangements. Nature 2003, 423, 760–762. [Google Scholar] [CrossRef] [PubMed]
- Nisa, M.U.; Huang, Y.; Benhamed, M.; Raynaud, C. The plant DNA damage response: Signaling pathways leading to growth inhibition and putative role in response to stress conditions. Front. Plant Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Bent, A.F. Microbial pathogens trigger host DNA double-strand breaks whose abundance is reduced by plant defense responses. PLoS Pathog. 2014, 10, e1004030. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wang, W.; Marques, J.; Mohan, R.; Saleh, A.; Durrant, W.E.; Song, J.; Dong, X. Salicylic acid activates DNA damage responses to potentiate plant immunity. Mol. Cell 2013, 52, 602–610. [Google Scholar] [CrossRef]
- Yao, Y.; Bilichak, A.; Golubov, A.; Kovalchuk, I. Local infection with oilseed rape mosaic virus promotes genetic rearrangements in systemic Arabidopsis tissue. Mutat. Res. 2011, 709, 7–14. [Google Scholar] [CrossRef]
- Mercier, R.; Mézard, C.; Jenczewski, E.; Macaisne, N.; Grelon, M. The molecular biology of meiosis in plants. Annu. Rev. Plant Biol. 2015, 66, 297–327. [Google Scholar] [CrossRef]
- Boyko, A.; Kathiria, P.; Zemp, F.J.; Yao, Y.; Pogribny, I.; Kovalchuk, I. Transgenerational changes in the genome stability and methylation in pathogen-infected plants: (virus-induced plant genome instability). Nucleic Acids Res. 2007, 35, 1714–1725. [Google Scholar] [CrossRef]
- Andronic, L. Viruses as triggers of DNA rearrangements in host plants. Can. J. Plant Sci. 2012, 92, 1083–1091. [Google Scholar] [CrossRef]
- Francis, K.E.; Lam, S.Y.; Harrison, B.D.; Bey, A.L.; Berchowitz, L.E.; Copenhaver, G.P. Pollen tetrad-based visual assay for meiotic recombination in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 3913–3918. [Google Scholar] [CrossRef]
- Berchowitz, L.E.; Copenhaver, G.P. Fluorescent Arabidopsis tetrads: A visual assay for quickly developing large crossover and crossover interference data sets. Nat. Protoc. 2008, 3, 41–50. [Google Scholar] [CrossRef]
- Meyers, B.C.; Kozik, A.; Griego, A.; Kuang, H.H.; Michelmore, R.W. Genome-wide analysis of NBS-LRR-encoding genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef] [PubMed]
- Lambing, C.; Osman, K.; Nuntasoontorn, K.; West, A.; Higgins, J.D.; Copenhaver, G.P.; Yang, J.; Armstrong, S.J.; Mechtler, K.; Roitinger, E.; et al. Arabidopsis PCH2 Mediates Meiotic Chromosome Remodeling and Maturation of Crossovers. PLoS Genet. 2015, 11, e1005372. [Google Scholar] [CrossRef] [PubMed]
- Girard, C.; Chelysheva, L.; Choinard, S.; Froger, N.; Macaisne, N.; Lehmemdi, A.; Mazel, J.; Crismani, W.; Mercier, R. AAA-ATPase FIDGETIN-LIKE 1 and Helicase FANCM Antagonize Meiotic Crossovers by Distinct Mechanisms. PLoS Genet. 2015, 11, e1005369. [Google Scholar] [CrossRef]
- Jahns, M.T.; Vezon, D.; Chambon, A.; Pereira, L.; Falque, M.; Martin, O.; Chelysheva, L.; Grelon, M. Crossover localisation is regulated by the neddylation posttranslational regulatory pathway. PLoS Biol. 2014, 12, e1001930. [Google Scholar] [CrossRef]
- Crismani, W.; Portemer, V.; Froger, N.; Chelysheva, L.; Horlow, C.; Vrielynck, N.; Mercier, R. MCM8 Is Required for a Pathway of Meiotic Double-Strand Break Repair Independent of DMC1 in Arabidopsis thaliana. PLoS Genet. 2013, 9. [Google Scholar] [CrossRef]
- Sun, Y.; Ambrose, J.H.; Haughey, B.S.; Webster, T.D.; Pierrie, S.N.; Munoz, D.F.; Wellman, E.C.; Cherian, S.; Lewis, S.M.; Berchowitz, L.E.; et al. Deep Genome-Wide Measurement of Meiotic Gene Conversion Using Tetrad Analysis in Arabidopsis thaliana. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef]
- Crismani, W.; Girard, C.; Froger, N.; Pradillo, M.; Santos, J.L.; Chelysheva, L.; Copenhaver, G.P.; Horlow, C.; Mercier, R. FANCM Limits Meiotic Crossovers. Science 2012, 336, 1588–1590. [Google Scholar] [CrossRef]
- Chelysheva, L.; Vezon, D.; Chambon, A.; Gendrot, G.; Pereira, L.; Lemhemdi, A.; Vrielynck, N.; le Guin, S.; Novatchkova, M.; Grelon, M. The Arabidopsis HEI10 Is a New ZMM Protein Related to Zip3. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef]
- Macaisne, N.; Vignard, J.; Mercier, R. SHOC1 and PTD form an XPF-ERCC1-like complex that is required for formation of class I crossovers. J. Cell Sci. 2011, 124, 2687–2691. [Google Scholar] [CrossRef]
- Berchowitz, L.E.; Francis, K.E.; Bey, A.L.; Copenhaver, G.P. The role of AtMUS81 in interference-insensitive crossovers in A. thaliana. PLoS Genet. 2007, 3, e132. [Google Scholar] [CrossRef]
- Yelina, N.E.; Ziolkowski, P.; Miller, N.; Zhao, X.; Kelly, K.A.; Munoz, D.F.; Mann, D.J.; Copenhaver, G.P.; Henderson, I.R. High-throughput analysis of meiotic crossover frequency and interference via flow cytometry of fluorescent pollen in Arabidopsis thaliana. Nat. Protoc. 2013, 8, 2119–2134. [Google Scholar] [CrossRef] [PubMed]
- Perkins, D. Biochemical mutants in the smut fungus Ustilago maydis. Genetics 1949, 34, 607–626. [Google Scholar] [PubMed]
- Yao, Y.; Kathiria, P.; Kovalchuk, I. A systemic increase in the recombination frequency upon local infection of Arabidopsis thaliana plants with oilseed rape mosaic virus depends on plant age, the initial inoculum concentration and the time for virus replication. Front. Plant Sci. 2013, 4, 61. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.D. Wolbachia Infection Associated with Increased Recombination in Drosophila. G3 Genes Genomes Genet. 2019, 9, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Rowan, B.A.; Heavens, D.; Feuerborn, T.R.; Tock, A.J.; Henderson, I.R.; Weigel, D. An ultra high-density Arabidopsis thaliana crossover map that refines the influences of structural variation and epigenetic features. Genetics 2019, 213, 771–787. [Google Scholar] [CrossRef]
- Sun, H.; Rowan, B.A.; Flood, P.J.; Brandt, R.; Fuss, J.; Hancock, A.M.; Michelmore, R.W.; Huettel, B.; Schneeberger, K. Linked-read sequencing of gametes allows efficient genome-wide analysis of meiotic recombination. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Giraut, L.; Falque, M.; Drouaud, J.; Pereira, L.; Martin, O.C.; Mezard, C. Genome-wide crossover distribution in Arabidopsis thaliana meiosis reveals sex-specific patterns along chromosomes. PLoS Genet. 2011, 7, e1002354. [Google Scholar] [CrossRef]
- Li, F.; Liu, P.; Wang, T.; Bian, P.; Wu, Y.; Wu, L.; Yu, Z. The induction of bystander mutagenic effects in vivo by α-particle irradiation in whole Arabidopsis thaliana plants. Radiat. Res. 2010, 174, 228–237. [Google Scholar] [CrossRef]
- Filkowski, J.; Yeoman, A.; Kovalchuk, O.; Kovalchuk, I. Systemic plant signal triggers genome instability. Plant J. 2004, 38, 1–11. [Google Scholar] [CrossRef]
- Dumont, B.L.; Devlin, A.A.; Truempy, D.M.; Miller, J.C.; Singh, N.D. No Evidence that Infection Alters Global Recombination Rate in House Mice. PLoS ONE 2015, 10, e0142266. [Google Scholar] [CrossRef]
- Singh, N.D.; Criscoe, D.R.; Skolfield, S.; Kohl, K.P.; Keebaugh, E.S.; Schlenke, T.A. EVOLUTION. Fruit flies diversify their offspring in response to parasite infection. Science 2015, 349, 747–750. [Google Scholar] [CrossRef] [PubMed]
- Zilio, G.; Moesch, L.; Bovet, N.; Sarr, A.; Koella, J.C. The effect of parasite infection on the recombination rate of the mosquito Aedes aegypti. PLoS ONE 2018, 13, e0203481. [Google Scholar] [CrossRef] [PubMed]
- DeBolt, S. Copy number variation shapes genome diversity in Arabidopsis over immediate family generational scales. Genome Biol. Evol. 2010, 2, 441–453. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
INTERVAL I5ab |  |  |  |  |  |  |  |  |  |  |  |  | |||
| Nb of tetrads | Nb of plants | Nb of flowers | 0 CO | 1 CO | 1 CO | 2CO | 2CO | 2CO | 2CO | 2CO | 2 CO | 3CO | 3CO | 4CO | |
| Tetrad Types | Class 1 | Class 2 | Class 3 | Class 4 | Class 5 | Class 6 | Class 7 | Class 8 | Class 9 | Class 10 | Class 11 | Class 12 | Total Tetrads | ||
| Control Plants | 2 | 33 | 167 | 212 | 81 | 11 | 7 | 6 | 6 | 3 | 1 | 1 | 0 | 0 | 495 |
| Inoculated plants | 2 | 31 | 206 | 203 | 94 | 7 | 8 | 13 | 4 | 3 | 2 | 0 | 0 | 0 | 540 |
INTERVAL I5cd |  |  |  |  |  |  |  |  |  |  |  |  | |||
| Nb of tetrads | Nb of plants | Nb of flowers | 0 CO | 1 CO | 1 CO | 2CO | 2CO | 2CO | 2CO | 2CO | 2 CO | 3CO | 3CO | 4CO | |
| Tetrad Types | Class 1 | Class 2 | Class 3 | Class 4 | Class 5 | Class 6 | Class 7 | Class 8 | Class 9 | Class 10 | Class 11 | Class 12 | Total Tetrads | ||
| Control Plants | 2 | 33 | 582 | 78 | 119 | 2 | 2 | 2 | 0 | 1 | 2 | 0 | 0 | 0 | 788 |
| Inoculated plants | 2 | 37 | 443 | 68 | 82 | 2 | 4 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 600 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gratias, A.; Geffroy, V. Deciphering the Impact of a Bacterial Infection on Meiotic Recombination in Arabidopsis with Fluorescence Tagged Lines. Genes 2020, 11, 832. https://doi.org/10.3390/genes11070832
Gratias A, Geffroy V. Deciphering the Impact of a Bacterial Infection on Meiotic Recombination in Arabidopsis with Fluorescence Tagged Lines. Genes. 2020; 11(7):832. https://doi.org/10.3390/genes11070832
Chicago/Turabian StyleGratias, Ariane, and Valérie Geffroy. 2020. "Deciphering the Impact of a Bacterial Infection on Meiotic Recombination in Arabidopsis with Fluorescence Tagged Lines" Genes 11, no. 7: 832. https://doi.org/10.3390/genes11070832
APA StyleGratias, A., & Geffroy, V. (2020). Deciphering the Impact of a Bacterial Infection on Meiotic Recombination in Arabidopsis with Fluorescence Tagged Lines. Genes, 11(7), 832. https://doi.org/10.3390/genes11070832