Identification of Loci and Pathways Associated with Heifer Conception Rate in U.S. Holsteins

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animals and Phenotypes
2.2. DNA Extraction and Genotyping
2.3. Quality Control
2.4. Genome-Wide Association Analysis
2.5. Positional Candidate Genes
2.6. Transcription Factor Binding Sites
2.7. Pathway Analysis
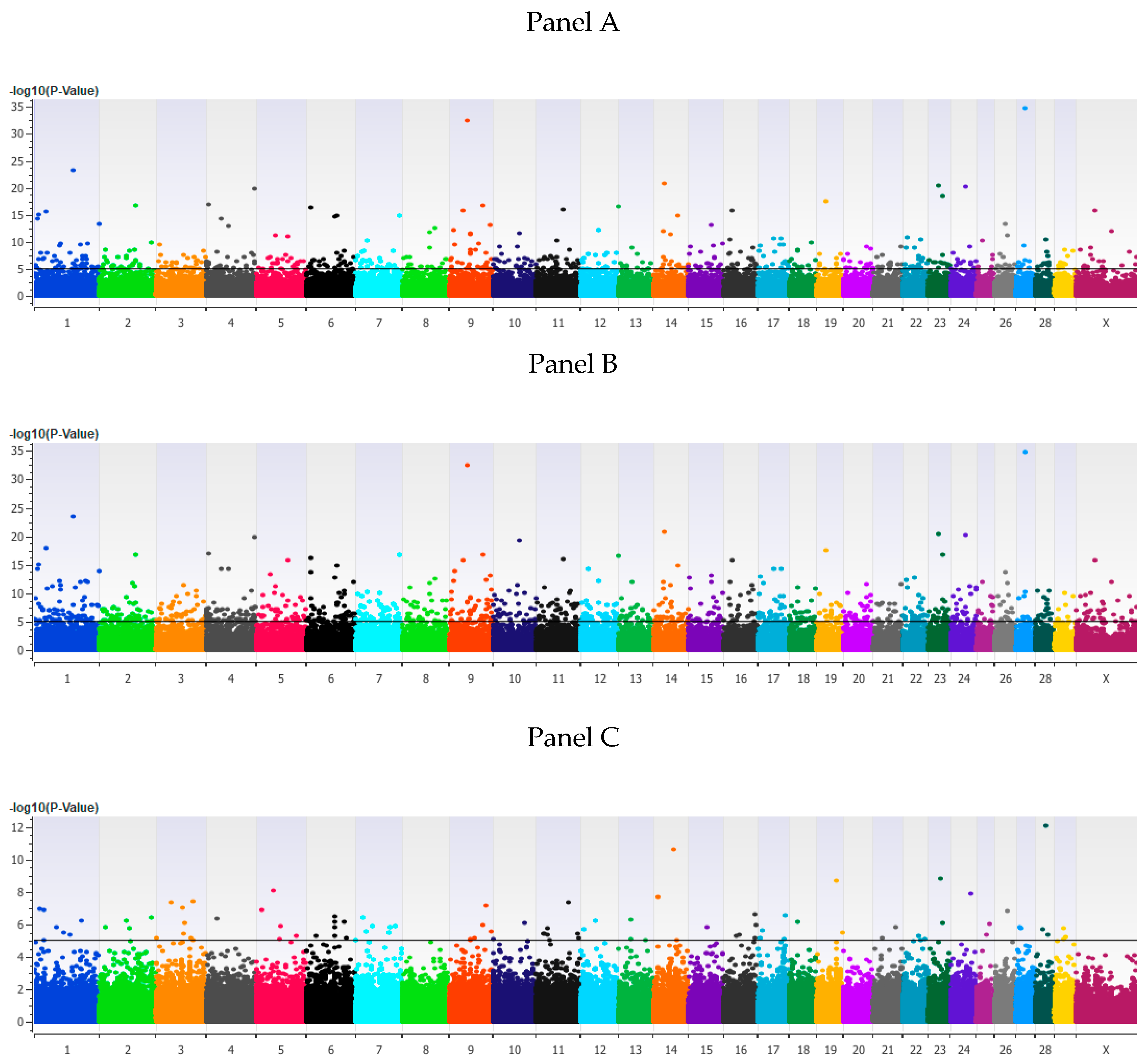
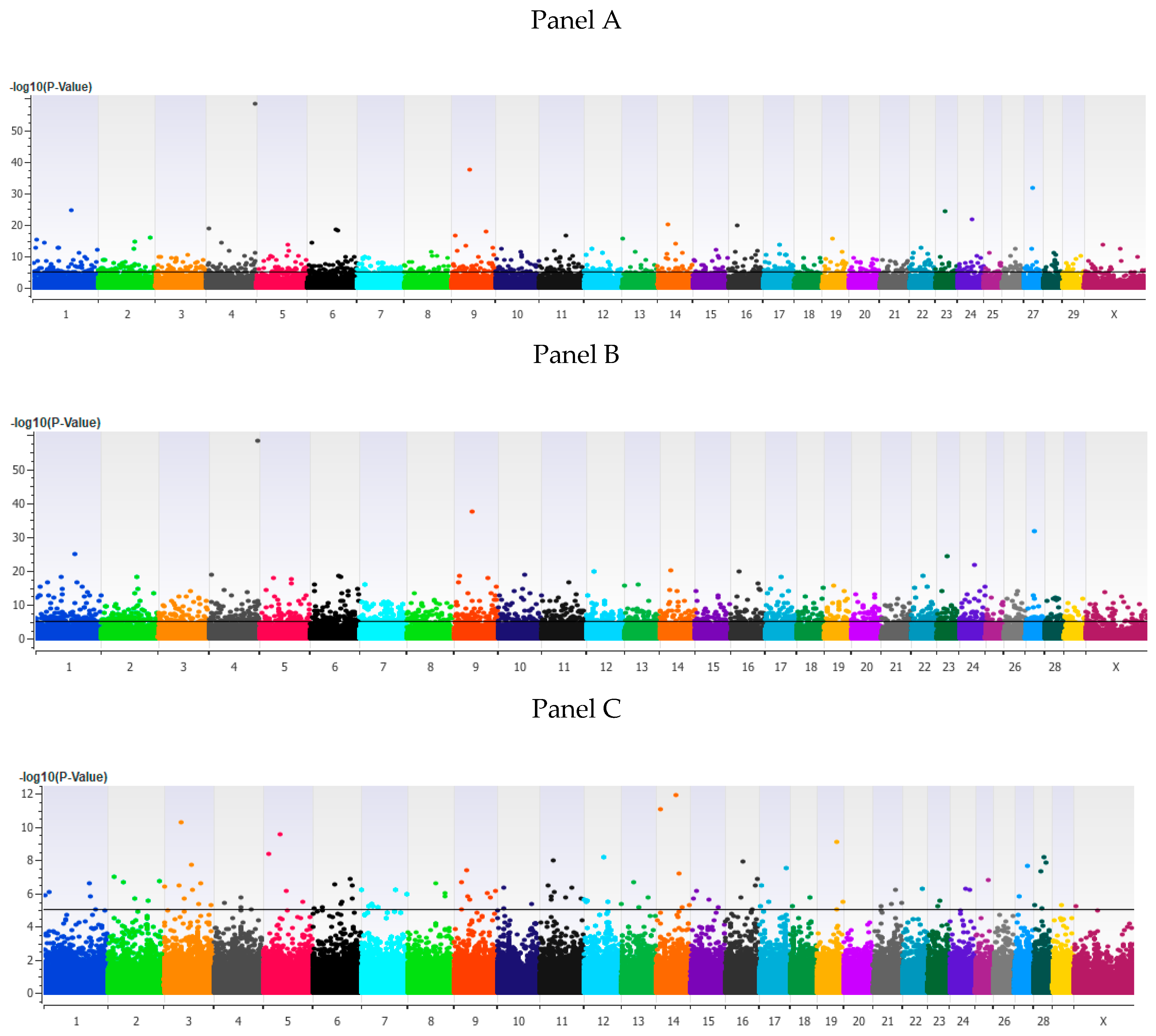
3. Results
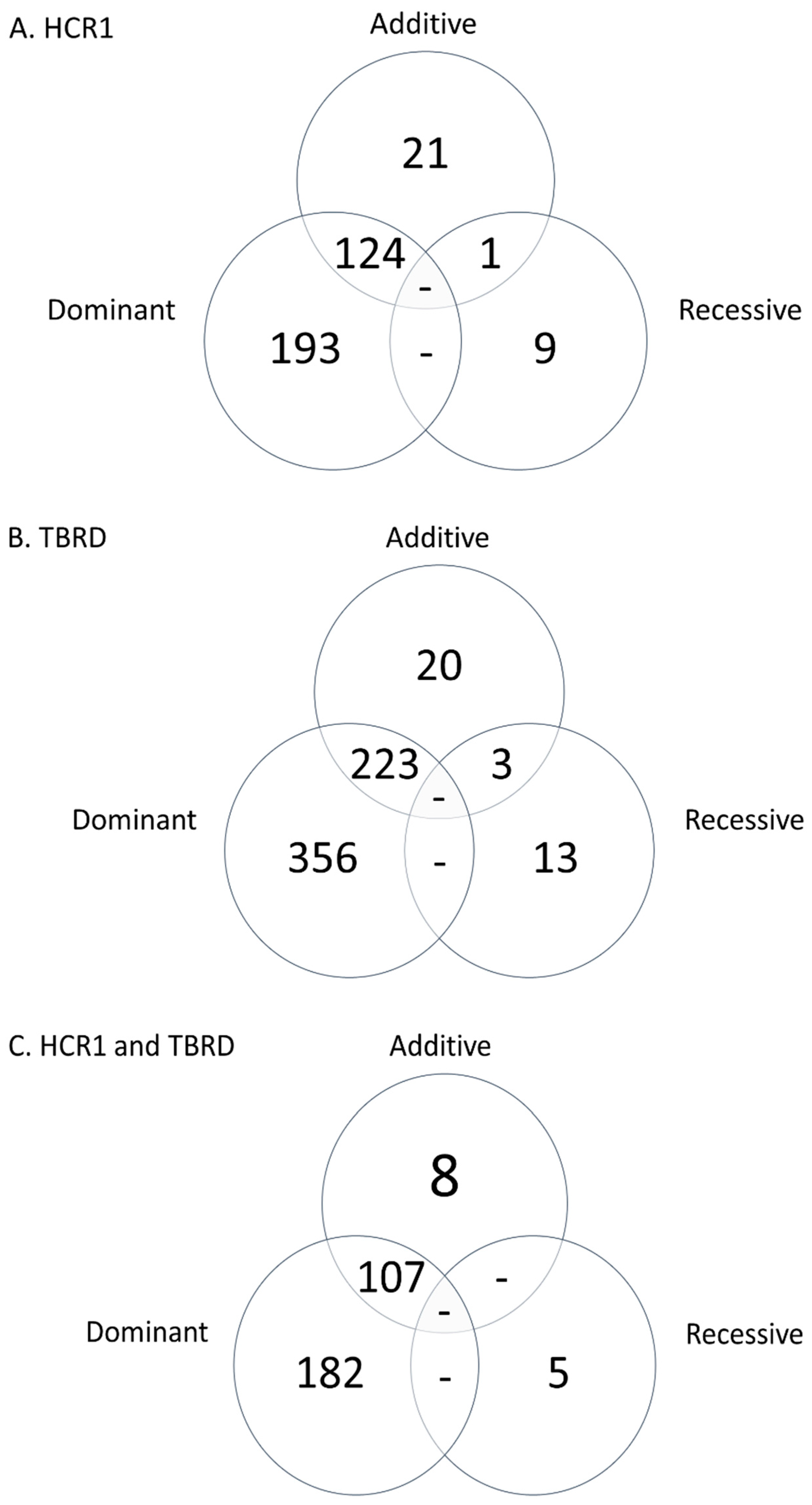
3.1. Genome-Wide Association Analysis
3.2. Validation of Associated Loci
3.3. Pathway Analysis
4. Discussion
4.1. Supporting SNPs in Fertility Associated Loci
4.2. Ingenuity Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Veerkamp, R.F.; Beerda, B. Genetics and genomics to improve fertility in high producing dairy cows. Theriogenology 2007, 68, S266–S273. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Madsen, P.; Lund, M.S.; Zhang, Y.; Nielsen, U.S.; Su, G. Improvement in genetic evaluation of female fertility in dairy cattle using multiple-trait models including milk production traits1. J. Anim. Sci. 2010, 88, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Boichard, D. Estimation of the economic value of conception rate in dairy cattle. Livest. Prod. Sci. 1990, 24, 187–204. [Google Scholar] [CrossRef]
- USDA Daughter Pregnancy Rate Evaluation–2005. Available online: https://www.aipl.arsusda.gov/reference/fertility/dpr.htm (accessed on 29 March 2019).
- Council on Dairy Cattle Breeding. Trend in Heifer Conception Rate for Holstein or Red & White. In: Trends & Stats: Genetic and Phenotypic Trend: Council on Dairy Cattle Breeding. Available online: https://queries.uscdcb.com/eval/summary/trend.cfm?R_Menu=HO.h#StartBody (accessed on 25 April 2018).
- Lucy, M. Reproductive Loss in High-Producing Dairy Cattle: Where Will It End? J. Dairy Sci. 2001, 84, 1277–1293. [Google Scholar] [CrossRef]
- Walsh, S.; Williams, E.; Evans, A. A review of the causes of poor fertility in high milk producing dairy cows. Anim. Reprod. Sci. 2011, 123, 127–138. [Google Scholar] [CrossRef]
- Dobson, H.; Smith, R.; Royal; Knight, C.; Sheldon, I.M. The high-producing dairy cow and its reproductive performance. Reprod. Domest. Anim. 2007, 42, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kiser, J.N.; Keuter, E.M.; Seabury, C.M.; Neupane, M.; Moraes, J.; Dalton, J.; Burns, G.; Spencer, T.; Neibergs, H.L. Validation of 46 loci associated with female fertility traits in cattle. BMC Genom. 2019, 20, 576. [Google Scholar] [CrossRef]
- Kiser, J.N.; Clancey, E.; Moraes, J.G.N.; Dalton, J.; Burns, G.W.; Spencer, T.E.; Neibergs, H.L. Identification of loci associated with conception rate in primiparous Holstein cows. BMC Genom. 2019, 20, 840. [Google Scholar] [CrossRef] [PubMed]
- Levy, C.S.; Slomiansky, V.; Gattelli, A.; Nahmod, K.; Pelisch, F.; Blaustein, M.; Srebrow, A.; Coso, O.A.; Kordon, E.C. Tumor necrosis factor alpha induces LIF expression through ERK1/2 activation in mammary epithelial cells. J. Cell. Biochem. 2010, 110, 857–865. [Google Scholar] [CrossRef]
- Browning, S.R.; Browning, B.L.; Daviglus, M.L.; Durazo-Arvizu, R.A.; Schneiderman, N.; Kaplan, R.C.; Laurie, C.C. Ancestry-specific recent effective population size in the Americas. PLoS Genet. 2018, 14, e1007385. [Google Scholar] [CrossRef]
- Kang, H.M.; Zaitlen, N.A.; Wade, C.M.; Kirby, A.; Heckerman, D.; Daly, M.J.; Eskin, E. Efficient Control of Population Structure in Model Organism Association Mapping. Genetics 2008, 178, 1709–1723. [Google Scholar] [CrossRef] [PubMed]
- Dudbridge, F.; Gusnanto, A. Estimation of significance thresholds for genomewide association scans. Genet. Epidemiol. 2008, 32, 227–234. [Google Scholar] [CrossRef]
- Gibbs, R.; Belmont, J.W.; Hardenbol, P.; Willis, T.D.; Fuli, Y.; Yang, H.; Ch’ang, L.Y.; Huang, W.; Liu, B.; Shen, Y.; et al. Initial planning groups: Populations and ethical, and Methods. The International HapMap Project. Nature 2008, 426, 789–796. [Google Scholar] [CrossRef]
- Mills, R.E.; 1000 Genomes Project; Walter, K.; Stewart, C.; Handsaker, R.E.; Chen, K.; Alkan, C.; Abyzov, A.; Yoon, S.C.; Ye, K.; et al. Mapping copy number variation by population-scale genome sequencing. Nature 2011, 470, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; The GIANT Consortium; Weedon, M.N.; Purcell, S.; Lettre, G.; Estrada, K.; Willer, C.J.; Smith, A.V.; Ingelsson, E.; O’Connell, J.R.; et al. Genomic inflation factors under polygenic inheritance. Eur. J. Hum. Genet. 2011, 19, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Weiss, K.M.; Clark, A.G. Linkage disequilibrium and the mapping of complex human traits. Trends Genet. 2002, 18, 19–24. [Google Scholar] [CrossRef]
- Sutter, N.B.; Eberle, M.A.; Parker, H.G.; Pullar, B.J.; Kirkness, E.F.; Kruglyak, L.; Ostrander, E.A. Extensive and breed-specific linkage disequilibrium in Canis familiaris. Genome Res. 2004, 14, 2388–2396. [Google Scholar] [CrossRef]
- Keel, B.N.; Lindholm-Perry, A.K.; Snelling, W.M. Evolutionary and Functional Features of Copy Number Variation in the Cattle Genome. Front. Genet. 2016, 7, 6. [Google Scholar] [CrossRef]
- SNP & Variation Suite v8.8.3 Manual. Available online: https://doc.goldenhelix.com/SVS/latest/svsmanual/mixedModelMethods/overview.html (accessed on 4 July 2020).
- Ensembl. Available online: https://uswest.ensembl.org/index.html (accessed on 4 July 2020).
- Gabriel, S.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The Structure of Haplotype Blocks in the Human Genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef]
- The Bovine Genome Database. Available online: https://bovinegenome.elsiklab.missouri.edu/?q=node/61 (accessed on 4 July 2020).
- Messeguer, X.; Escudero, R.; Farré, D.; Núñez, O.; Martinez, J.; Albà, M.M.; Martínez, J. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef] [PubMed]
- Farré, D.; Roset, R.; Huerta, M.; Adsuara, J.; Roselló, L.; Albà, M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef]
- Matys, V.; Kel-Margoulis, O.; Fricke, E.; Liebich, I.; Land, S.; Barre-Dirrie, A.; Reuter, I.; Chekmenev, D.; Krull, M.; Hornischer, K.; et al. TRANSFAC(R) and its module TRANSCompel(R): Transcriptional gene regulation in eukaryotes. Nucleic Acids Res. 2006, 34, 108. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2013, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Hayes, B.J.; Bowman, P.J.; Chamberlain, A.C.; Verbyla, K.; Goddard, M.E. Accuracy of genomic breeding values in multi-breed dairy cattle populations. Genet. Sel. Evol. 2009, 41, 51. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information SNP Database. Available online: https://www.ncbi.nlm.nih.gov/snp/ (accessed on 4 July 2020).
- Azizieh, F.; Raghupathy, R.G. Tumor necrosis factor-α and pregnancy complications: A prospective study. Med. Princ. Pract. 2014, 24, 165–170. [Google Scholar] [CrossRef]
- Prasad, A.; Schnabel, R.D.; McKay, S.D.; Murdoch, B.M.; Stothard, P.; Kolbehdari, D.; Wang, Z.; Taylor, J.F.; Moore, S. Linkage disequilibrium and signatures of selection on chromosomes 19 and 29 in beef and dairy cattle. Anim. Genet. 2008, 39, 597–605. [Google Scholar] [CrossRef]
- Pausch, H.; Schwarzenbacher, H.; Burgstaller, J.; Flisikowski, K.; Wurmser, C.; Jansen, S.; Jung, S.; Schnieke, A.; Wittek, T.; Fries, R. Homozygous haplotype deficiency reveals deleterious mutations compromising reproductive and rearing success in cattle. BMC Genom. 2015, 16, 312. [Google Scholar] [CrossRef]
- Moore, S.; Pryce, J.; Hayes, B.J.; Chamberlain, A.J.; Kemper, K.E.; Berry, D.P.; McCabe, M.; Cormican, P.; Lonergan, P.; Fair, T.; et al. Differentially Expressed Genes in Endometrium and Corpus Luteum of Holstein Cows Selected for High and Low Fertility Are Enriched for Sequence Variants Associated with Fertility1. Boil. Reprod. 2016, 94, 19. [Google Scholar] [CrossRef]
- Cole, J.B.; Wiggans, G.; Ma, L.; Sonstegard, T.S.; Lawlor, T.J.; Crooker, B.; Van Tassell, C.P.; Yang, J.; Wang, S.; Matukumalli, L.K.; et al. Genome-wide association analysis of thirty one production, health, reproduction and body conformation traits in contemporary U.S. Holstein cows. BMC Genom. 2011, 12, 408. [Google Scholar] [CrossRef]
- Höglund, J.K.; Sahana, G.; Guldbrandtsen, B.; Lund, M.S. Validation of associations for female fertility traits in Nordic Holstein, Nordic Red and Jersey dairy cattle. BMC Genet. 2014, 15, 8. [Google Scholar] [CrossRef]
- Fonseca, P.A.D.S.; Id-Lahoucine, S.; Reverter, A.; Medrano, J.F.; Fortes, M.R.S.; Casellas, J.; Miglior, F.; Brito, L.; Carvalho, M.R.S.; Schenkel, F.S.; et al. Combining multi-OMICs information to identify key-regulator genes for pleiotropic effect on fertility and production traits in beef cattle. PLoS ONE 2018, 13, e0205295. [Google Scholar] [CrossRef] [PubMed]
- Nayeri, S.; Sargolzaei, M.; Abo-Ismail, M.K.; May, N.; Miller, S.P.; Schenkel, F.; Moore, S.; Stothard, P. Genome-wide association for milk production and female fertility traits in Canadian dairy Holstein cattle. BMC Genet. 2016, 17, 75. [Google Scholar] [CrossRef] [PubMed]
- Minozzi, G.; Nicolazzi, E.L.; Stella, A.; Biffani, S.; Negrini, R.; Lazzari, B.; Marsan, P.A.; Williams, J.L. Genome Wide Analysis of Fertility and Production Traits in Italian Holstein Cattle. PLoS ONE 2013, 8, e80219. [Google Scholar] [CrossRef] [PubMed]
- Neupane, M.; Geary, T.W.; Kiser, J.N.; Burns, G.W.; Hansen, P.J.; Spencer, T.E.; Neibergs, H.L. Loci and pathways associated with uterine capacity for pregnancy and fertility in beef cattle. PLoS ONE 2017, 12, e0188997. [Google Scholar] [CrossRef] [PubMed]
- Cochran, S.D.; Cole, J.B.; Null, D.J.; Hansen, P.J. Discovery of single nucleotide polymorphisms in candidate genes associated with fertility and production traits in Holstein cattle. BMC Genet. 2013, 14, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.S.; Denicol, A.C.; Cole, J.B.; Null, D.J.; Hansen, P.J. Use of single nucleotide polymorphisms in candidate genes associated with daughter pregnancy rate for prediction of genetic merit for reproduction in Holstein cows. Anim. Genet. 2016, 47, 288–297. [Google Scholar] [CrossRef]
- Geary, T.W.; Burns, G.W.; Moraes, J.G.; Moss, J.I.; Denicol, A.C.; Dobbs, K.B.; Ortega, M.S.; Hansen, P.J.; Wehrman, M.E.; Neibergs, H.; et al. Identification of Beef Heifers with Superior Uterine Capacity for Pregnancy1. Boil. Reprod. 2016, 95, 47. [Google Scholar] [CrossRef] [PubMed]
- Moraes, J.G.N.; Behura, S.K.; Geary, T.W.; Hansen, P.J.; Neibergs, H.L.; Spencer, T. Uterine influences on conceptus development in fertility-classified animals. Proc. Natl. Acad. Sci. USA 2018, 115, E1749–E1758. [Google Scholar] [CrossRef]
- Sargolzaei, M.; Schenkel, F.S.; Jansen, G.; Schaeffer, L. Extent of Linkage Disequilibrium in Holstein Cattle in North America. J. Dairy Sci. 2008, 91, 2106–2117. [Google Scholar] [CrossRef]
- Yokoyama, M.; Ichinoe, M.; Okina, S.; Sakurai, Y.; Nakada, N.; Yanagisawa, N.; Jiang, S.-X.; Numata, Y.; Umezawa, A.; Miyazaki, K.; et al. CD109, a negative regulator of TGF-β signaling, is a putative risk marker in diffuse large B-cell lymphoma. Int. J. Hematol. 2016, 105, 614–622. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Z.; Wang, J.; Lv, D.; Zhu, T.; Wang, F.; Tian, X.; Yao, Y.; Ji, P.; Liu, G. Melatonin regulates the activities of ovary and delays the fertility decline in female animals via MT1/AMPK pathway. J. Pineal Res. 2019, 66, e12550. [Google Scholar] [CrossRef] [PubMed]
- Gaddis, K.P.; Null, D.; Cole, J.B. Explorations in genome-wide association studies and network analyses with dairy cattle fertility traits. J. Dairy Sci. 2016, 99, 6420–6435. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Luo, Y.; Liang, B.; Ye, L.; Lu, G.; He, W. Potential applications of MEG3 in cancer diagnosis and prognosis. Oncotarget 2017, 8, 73282–73295. [Google Scholar] [CrossRef] [PubMed]
- García-Acero, M.L.; Acosta, J. Whole-Exome Sequencing Identifies a de novo AHDC1 Mutation in a Colombian Patient with Xia-Gibbs Syndrome. Mol. Syndr. 2017, 8, 308–312. [Google Scholar] [CrossRef] [PubMed]
- Ballester, M.; Molist, J.; López-Béjar, M.; Sánchez, A.; Santalo, J.; Folch, J.M.; Ibáñez, E. Disruption of the mouse phospholipase C-β1 gene in a β-lactoglobulin transgenic line affects viability, growth, and fertility in mice. Gene 2004, 341, 279–289. [Google Scholar] [CrossRef]
- Makker, A.; Goel, M.M.; Mahdi, A.A. PI3K/PTEN/Akt and TSC/mTOR signaling pathways, ovarian dysfunction, and infertility: An update. J. Mol. Endocrinol. 2014, 53, R103–R118. [Google Scholar] [CrossRef]
- Tak, Y.G.; Farnham, P.J. Making sense of GWAS: Using epigenomics and genome engineering to understand the functional relevance of SNPs in non-coding regions of the human genome. Epigenetics Chromatin 2015, 8, 1–18. [Google Scholar] [CrossRef]
- Bhanuprakash, V.; Chhotaray, S.; Pruthviraj, D.R.; Rawat, C.; Karthikeyan, A.; Panigrahi, M. Copy number variation in livestock: A mini review. Vet. World 2018, 11, 535–541. [Google Scholar] [CrossRef]
- Singh, H.; Aplin, J.D. Adhesion molecules in endometrial epithelium: Tissue integrity and embryo implantation. J. Anat. 2009, 215, 3–13. [Google Scholar] [CrossRef]
- Onuchic, L.F.; Furu, L.; Nagasawa, Y.; Hou, X.; Eggermann, T.; Ren, Z.; Bergmann, C.; Senderek, J.; Esquivel, E.; Zeltner, R.; et al. PKHD1, the Polycystic Kidney and Hepatic Disease 1 Gene, Encodes a Novel Large Protein Containing Multiple Immunoglobulin-Like Plexin-Transcription–Factor Domains and Parallel Beta-Helix 1 Repeats. Am. J. Hum. Genet. 2002, 70, 1305–1317. [Google Scholar] [CrossRef]
- Song, J.-Y.; Ichtchenko, K.; Südhof, T.C.; Brose, N. Neuroligin 1 is a postsynaptic cell-adhesion molecule of excitatory synapses. Proc. Natl. Acad. Sci. USA 1999, 96, 1100–1105. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Long, L.K.; Hatch, M.M.; Hoffman, D.A. DPP6 Domains Responsible for Its Localization and Function*. J. Boil. Chem. 2014, 289, 32153–32165. [Google Scholar] [CrossRef] [PubMed]
- Brainard, A.M.; Korovkina, V.P.; England, S.K. Potassium channels and uterine function. Semin. Cell Dev. Boil. 2007, 18, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.C.; McClure, M.C.; Smith, M.A.; Abel, P.; Bradley, M.E. The role of voltage-gated potassium channels in the regulation of mouse uterine contractility. Reprod. Boil. Endocrinol. 2007, 5, 41. [Google Scholar] [CrossRef]
- Zhu, R.; Xiao, D.; Zhang, L. Potassium channels and uterine vascular adaptation to pregnancy and chronic hypoxia. Curr. Vasc. Pharmacol. 2013, 11, 737–747. [Google Scholar] [CrossRef]
- Tribe, R.M.; Hassoni, A.A.; Aaronson, P.I.; Knock, G.A. Modulation of Potassium Current Characteristics in Human Myometrial Smooth Muscle by 17β-Estradiol and Progesterone1. Boil. Reprod. 2001, 64, 1526–1534. [Google Scholar] [CrossRef]
- Clabaut, M.; Duclos, R.; Acritopoulou-Fourcroy, S. Increase of uterine motility and simultaneous decrease of progesterone concentrations in the rat after bilateral ovariectomy at mid-pregnancy. Reproduction 1988, 82, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Jeschke, U.; Walzel, H.; Mylonas, I.; Papadopoulos, P.; Shabani, N.; Kuhn, C.; Schulze, S.; Friese, K.; Karsten, U.; Anz, D.; et al. The Human Endometrium Expresses the Glycoprotein Mucin-1 and Shows Positive Correlation for Thomsen-Friedenreich Epitope Expression and Galectin-1 Binding. J. Histochem. Cytochem. 2009, 57, 871–881. [Google Scholar] [CrossRef] [PubMed]
- DeSouza, M.M.; Lagow, E.; Carson, D.D. Mucin Functions and Expression in Mammalian Reproductive Tract Tissues. Biochem. Biophys. Res. Commun. 1998, 247, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dharmaraj, N.; Gendler, S.J.; Carson, D. Expression of human MUC1 during early pregnancy in the human MUC1 transgenic mouse model. Boil. Reprod. 2009, 81, 1182–1188. [Google Scholar] [CrossRef]
- Minten, M.A.; Bilby, T.R.; Bruno, R.G.S.; Allen, C.C.; Madsen, C.A.; Wang, Z.; Sawyer, J.E.; Tibary, A.; Neibergs, H.L.; Geary, T.W.; et al. Effects of Fertility on Gene Expression and Function of the Bovine Endometrium. PLoS ONE 2013, 8, e69444. [Google Scholar] [CrossRef] [PubMed]
- Bygdeman, M.; Swahn, M. 2 Uterine contractility during pregnancy and the effect of abortifacient drugs. Baillière’s Clin. Obstet Gynaecol. 1990, 4, 249–261. [Google Scholar] [CrossRef]
- Ellinger, I. The Calcium-Sensing Receptor and the Reproductive System. Front Physiol. 2016, 7, 371. [Google Scholar] [CrossRef] [PubMed]
- Sötje, B.; Haffner, D.; Ziegler, W.H. Fibrocystin in MDCK cells: Impact on cell adhesion and epithelial morphogenesis. Mol. Cell. Pediatr. 2015, 2, A26. [Google Scholar] [CrossRef][Green Version]
- Bergmann, C.; Senderek, J.; Küpper, F.; Schneider, F.; Dornia, C.; Windelen, E.; Eggermann, T.; Rudnik-Schöneborn, S.; Kirfel, J.; Furu, L.; et al. PKHD1mutations in autosomal recessive polycystic kidney disease (ARPKD). Hum. Mutat. 2004, 23, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.-L.; Zhao, H.-B.; Li, M.-Q.; Du, M.-R.; Meng, Y.-H.; Li, D.-J. Focal Adhesion Kinase Signaling is Necessary for the Cyclosporin A-Enhanced Migration and Invasion of Human Trophoblast Cells. Placenta 2012, 33, 704–711. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2016, 45, D353–D361. [Google Scholar] [CrossRef]
- Talukder, A.K.; Yousef, M.S.; Rashid, M.B.; Awai, K.; Acosta, T.J.; Shimizu, T.; Okuda, K.; Shimada, M.; Imakawa, K.; Miyamoto, A. Bovine embryo induces an anti-inflammatory response in uterine epithelial cells and immune cells in vitro: Possible involvement of interferon tau as an intermediator. J. Reprod. Dev. 2017, 63, 425–434. [Google Scholar] [CrossRef]
- Liu, X.-P.; Yin, X.-H.; Yan, X.-H.; Zeng, X.-T.; Wang, X. The Clinical Relevance of Fragile Histidine Triad Protein (FHIT) in Patients with Bladder Cancer. Med. Sci. Monit. 2018, 24, 3113–3118. [Google Scholar] [CrossRef]
- Kadri, N.K.; Sahana, G.; Charlier, C.; Iso-Touru, T.; Guldbrandtsen, B.; Karim, L.; Nielsen, U.S.; Panitz, F.; Aamand, G.P.; Schulman, N.; et al. A 660-Kb Deletion with Antagonistic Effects on Fertility and Milk Production Segregates at High Frequency in Nordic Red Cattle: Additional Evidence for the Common Occurrence of Balancing Selection in Livestock. PLoS Genet. 2014, 10, e1004049. [Google Scholar] [CrossRef]
- McDaneld, T.G.; Kuehn, L.A.; Thomas, M.G.; Pollak, E.J.; Keele, J.W. Deletion on chromosome 5 associated with decreased reproductive efficiency in female cattle1,2,3,4. J. Anim. Sci. 2014, 92, 1378–1384. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; Endicott, R.M.; Parghi, S.S.; Walker, M.; Kidd, J.M.; Trask, B.J. Extensive Copy-Number Variation of the Human Olfactory Receptor Gene Family. Am. J. Hum. Genet. 2008, 83, 228–242. [Google Scholar] [CrossRef]
- Fair, T. The Contribution of the Maternal Immune System to the Establishment of Pregnancy in Cattle. Front. Immunol. 2015, 6, 7. [Google Scholar] [CrossRef]
- Faubert, B.L.; Kaminski, N.E. AP-1 activity is negatively regulated by cannabinol through inhibition of its protein components, c-fos and c-jun. J. Leukoc. Boil. 2000, 67, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Lye, S.J. Differential expression of activator protein-1 transcription factors in pregnant rat myometrium. Boil. Reprod. 2002, 67, 240–246. [Google Scholar] [CrossRef]
- Mudduwa, L.K.; Peiris, H.; Gunasekara, S.; Abeysiriwardhana, D.; Liyanage, N.; Rayala, S.K.; Liyanage, T.G. KIBRA; a novel biomarker predicting recurrence free survival of breast cancer patients receiving adjuvant therapy. BMC Cancer 2018, 18, 589. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, X.; Huang, J.; Feng, L.; Dolinta, K.G.; Chen, J. Defining the protein-protein interaction network of the human hippo pathway. Mol. Cell. Proteom. 2013, 13, 119–131. [Google Scholar] [CrossRef]
- Issaragrisil, S.; Lorthongpanich, C. Emerging Role of the Hippo Signaling Pathway in Position Sensing and Lineage Specification in Mammalian Preimplantation Embryos1. Boil. Reprod. 2015, 92, 6. [Google Scholar] [CrossRef]
- Banerjee, A.; Meenakumari, K.J.; Udin, S.; Krishna, A. Melatonin regulates delayed embryonic development in the short-nosed fruit bat, Cynopterus sphinx. Reproduction 2009, 138, 935–944. [Google Scholar] [CrossRef][Green Version]
- Olcese, J.M.; Beesley, S. Clinical significance of melatonin receptors in the human myometrium. Fertil. Steril. 2014, 102, 329–335. [Google Scholar] [CrossRef]
- Carlomagno, G.; Minini, M.; Tilotta, M.; Unfer, V. From Implantation to Birth: Insight into Molecular Melatonin Functions. Int. J. Mol. Sci. 2018, 19, 2802. [Google Scholar] [CrossRef] [PubMed]
- Salleh, N.; Giribabu, N. Leukemia Inhibitory Factor: Roles in Embryo Implantation and in Nonhormonal Contraception. Sci. World J. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| BTA 1 | BP Position 2 | SNP ID 3 | Model 4 | HCR1 p-Value 5 | TBRD p-Values 6 | Positional Candidate Genes 7 |
|---|---|---|---|---|---|---|
| 1 | 94,300,486 | rs41606324 | additive dominant | 6.28 × 10−24 3.65 × 10−24 | 3.97 × 10−25 2.01 × 10−25 | NLGN1 |
| 4 | 117,606,629 | rs137620917 | additive dominant | 1.47 × 10−20 1.47 × 10−20 | 4.67 × 10−59 4.67 × 10−59 | DPP6 |
| 5 | 40,576,651 | rs43434026 | recessive | 8.48 × 10−9 | 3.03 × 10−10 | MUC19 |
| 9 | 45,148,825 | rs41609220 | additive | 4.89 × 10−33 | 5.72 × 10−38 | - |
| 10 | 63,217,046 | rs133388647 | dominant | 6.98 × 10−20 | 1.36 × 10−19 | - |
| 14 | 25,633,578 | rs134826452 | additive dominant | 1.67 × 10−21 1.67 × 10−21 | 7.83 × 10−21 7.83 × 10−21 | - |
| 14 | 50,291,072 | rs41913814 | recessive | 2.64 × 10−11 | 1.21 × 10−21 | - |
| 19 | 47,475,942 | rs41917870 | recessive | 2.14 × 10−9 | 8.34 × 10−10 | EFCAB3 |
| 23 | 23,965,902 | rs41634508 | additive dominant | 4.77 × 10−21 4.77 × 10−21 | 5.55 × 10−25 5.55 × 10−25 | PKHD1 |
| 27 | 21,375,791 | rs132728892 | additive dominant | 1.84 × 10−35 1.84 × 10−35 | 2.90 × 10−32 2.90 × 10−32 | - |
| BTA 1 | BP Position 2 | SNP ID 3 | Study 4 | Trait in Other Studies 5 |
|---|---|---|---|---|
| 1 | 1–6 | rs136767715 | Pausch et al. 2015 [33] | FH |
| 1 | 25–27 | rs133334228 | Moore et al. 2016 [34] | GMF |
| 1 | 129–130 | rs136894301 | Cole et al. 2011 [35] | DPR |
| 1 | 61–62 | rs42760220 rs42675527 | Hoglund 2014 [36] | AISC |
| 1 | 61–63 | rs43652271 | Fonseca et al. 2018 [37] | PLAF |
| 1 | 84–85 | rs109101339 | Hoglund 2014 [36] | AISC/AISH |
| 1 | 94–96 | rs135160436 | Hoglund 2014 [36] | AISH |
| 1 | 88–89 | rs109891698 | Hoglund 2014 [36] | AISC |
| 3 | 32–33 | rs43336503 | Fonseca et al. 2018 [37] | PLAF |
| 3 | 119–121 | rs41615294 | Hoglund 2014 [36] | AISH |
| 3 | 96–97 | rs41585055 | Hoglund 2014 [36] | AISC |
| 3 | 98–99 | rs43356386 | Hoglund 2014 [36] | AISC |
| 4 | 29–30 | rs43181427 | Hoglund 2014 [36] | AISC |
| 4 | 37–38 | rs133417267 | Fonseca et al. 2018 [37] | PLAF |
| 5 | 88–89 | rs137120693 | Nayeri et al. 2016 [38] | CTFS |
| 6 | 92–94 | rs110063753 rs43477811 | Hoglund 2014 [36] | AISC |
| 7 | 12–13 | rs133465587 | Cole et al. 2011 [35] | FH |
| 7 | 57–59 | 57,914,820 | Hoglund 2014 [36] | AISC |
| 8 | 59–61 | rs134066757 | Hoglund 2014 [36] | AISC |
| 8 | 72–74 | rs133191466 | Hoglund 2014 [36] | AISC |
| 8 | 84–86 | rs43137599 | Hoglund 2014 [36] | AISC |
| 9 | 28–30 | rs109636996 | Hoglund 2014 [36] | AISC |
| 9 | 61–61 | rs43601819 | Hoglund 2014 [36] | AISC |
| 9 | 90–91 | rs43608400 | Hoglund 2014 [36] | AISC |
| 10 | 26–35 | rs110325782 | Cole et al. 2011 [35] | FH |
| 10 | 63–65 | rs133765760 | Hoglund 2014 [36] | AISC |
| 11 | 17–18 | rs134981474 | Hoglund 2014 [36] | AISC |
| 11 | 21–22 | rs43669974 | Moore et al. 2016 [34] | GMF |
| 11 | 57–59 | rs42234541 rs136444067 | Hoglund 2014 [36] | AISH |
| 11 | 93–95 | rs134709354 | Hoglund 2014 [36] | AISC |
| 11 | 101–102 | rs136026124 | Fonseca et al. 2018 [37] | PLAF |
| 12 | 81–82 | rs135307240 | Fonseca et al. 2018 [37] | PLAF |
| 13 | 45–46 | rs42628484 | Hoglund 2014 [36] | AISC |
| 14 | 45–46 | rs41630614 | Moore et al. 2016 [34] | GMF |
| 14 | 44–46 | rs136545426 | Minnozi et al. 2013 [39] | NR56 |
| 15 | 63–64 | rs135885524 | Fonseca et al. 2018 [37] | PLAF |
| 16 | 16–17 | rs133881641 | Hoglund 2014 [36] | AISC |
| 16 | 21–21 | rs108994652 | Neupane et al. 2017 [40] | ET |
| 16 | 69–70 | rs42385478 | Hoglund 2014 [36] | AISC |
| 17 | 14–15 | rs110372003 | Hoglund 2014 [36] | AISC |
| 17 | 57–58 | rs137751476 | Hoglund 2014 [36] | AISC |
| 18 | 25–26 | rs135881758 | Cochran 2013 [41] | DPR |
| 18 | 25–26 | rs135881758 | Ortega et al. 2016 [42] | CCR |
| 18 | 48–49 | rs137310621 | Minnozi et al. 2013 [39] | NR56 |
| 20 | 46–47 | rs41949865 | Hoglund 2014 [36] | AISC |
| 20 | 53–54 | rs135839614 | Hoglund 2014 [36] | AISC |
| 21 | 52–54 | rs137802601 | Nayeri et al. 2016 [38] | CTFS |
| 26 | 6–8 | rs133146678 | Fonseca et al. 2018 [37] | PLAF |
| 26 | 25–26 | rs110088444 | Hoglund 2014 [36] | AISC |
| 26 | 40–41 | rs136057362 | Cole et al. 2011 [35] | FH |
| X | 0–1 | rs42069602 | Cole et al. 2011 [35] | DPR |
| Genes 1 | Tissue 2 | Phenotype 3 |
|---|---|---|
| ABLIM3 | Pregnant Endometrium HF vs. SF | TBRD |
| AHCYL2 | HF vs. SF Conceptuses | TBRD |
| AHDC1 | HF—Pregnant vs. Open -Endometrium | HCR1 TBRD |
| CADPS | SF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| CD109 | HF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| CHD9 | HF vs. SF Conceptuses | HCR1 TBRD |
| DCP1A | HF vs. SF Conceptuses | HCR1 TBRD |
| EPHA3 | Pregnant Endometrium HF vs. SF | TBRD |
| FHIT | HF vs. SF Conceptuses | TBRD |
| FNIP2 | HF vs. SF Conceptuses | HCR1 TBRD |
| GRIA4 | HF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| KIAA0825 | HF vs. SF Conceptuses | TBRD |
| MAP6 | HF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| MEG3 | HF—Pregnant vs. Open Endometrium HF vs. SF Conceptuses | HCR1 TBRD |
| NTRK2 | Pregnant Endometrium HF vs. SF and HF—Pregnant vs. Open Endometrium SF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| PIK3R1 | HF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| PKHD1 | HF vs. SF Conceptuses | HCR1 TBRD |
| PLCB1 | HF—Pregnant vs. Open Endometrium and SF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| POLD3 | SF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| PRKG1 | HF vs. SF Conceptuses | HCR1 TBRD |
| RAB3C | SF—Pregnant vs. Open Endometrium | TBRD |
| ROBO1 | SF—Pregnant vs. Open Endometrium | TBRD |
| SDK2 | HF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| SORCS3 | HF vs. SF Conceptuses | HCR1 TBRD |
| STC1 | SF—Pregnant vs. Open Endometrium | TBRD |
| STX16 | HF vs. SF Conceptuses | TBRD |
| SYNE1 | HF—Pregnant vs. Open Endometrium | HCR1 TBRD |
| TDRD1 | HF vs. SF Conceptuses | HCR1 TBRD |
| TIAM1 | HF vs. SF Conceptuses | HCR1 TBRD |
| UPK1B | SF—Pregnant vs. Open Endometrium HF vs. SF Conceptuses Open Endometrium HF vs. SF | HCR1 TBRD |
| ZCCHC14 | HF vs. SF Conceptuses | TBRD |
| Ingenuity Canonical Pathways 1 | Significance (p-Value) 2 | Number of Target Molecules 3 |
|---|---|---|
| Neuropathic Pain Signaling In Dorsal Horn Neurons | 0.002213 | 12 |
| PI3K Signaling in B Lymphocytes | 0.002291 | 13 |
| Axonal Guidance Signaling | 0.003428 | 25 |
| Role of Macrophages, Fibroblasts and Endothelial Cells in Rheumatoid Arthritis | 0.003673 | 20 |
| Prolactin Signaling | 0.003981 | 10 |
| VDR/RXR Activation | 0.005821 | 9 |
| Thrombin Signaling | 0.005888 | 15 |
| P2Y Purigenic Receptor Signaling Pathway | 0.006026 | 12 |
| Aryl Hydrocarbon Receptor Signaling | 0.006237 | 12 |
| Endothelin-1 Signaling | 0.007943 | 14 |
| Gap Junction Signaling | 0.00912 | 14 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galliou, J.M.; Kiser, J.N.; Oliver, K.F.; Seabury, C.M.; Moraes, J.G.N.; Burns, G.W.; Spencer, T.E.; Dalton, J.; Neibergs, H.L. Identification of Loci and Pathways Associated with Heifer Conception Rate in U.S. Holsteins. Genes 2020, 11, 767. https://doi.org/10.3390/genes11070767
Galliou JM, Kiser JN, Oliver KF, Seabury CM, Moraes JGN, Burns GW, Spencer TE, Dalton J, Neibergs HL. Identification of Loci and Pathways Associated with Heifer Conception Rate in U.S. Holsteins. Genes. 2020; 11(7):767. https://doi.org/10.3390/genes11070767
Chicago/Turabian StyleGalliou, Justine M., Jennifer N. Kiser, Kayleen F. Oliver, Christopher M. Seabury, Joao G. N. Moraes, Gregory W. Burns, Thomas E. Spencer, Joseph Dalton, and Holly L. Neibergs. 2020. "Identification of Loci and Pathways Associated with Heifer Conception Rate in U.S. Holsteins" Genes 11, no. 7: 767. https://doi.org/10.3390/genes11070767
APA StyleGalliou, J. M., Kiser, J. N., Oliver, K. F., Seabury, C. M., Moraes, J. G. N., Burns, G. W., Spencer, T. E., Dalton, J., & Neibergs, H. L. (2020). Identification of Loci and Pathways Associated with Heifer Conception Rate in U.S. Holsteins. Genes, 11(7), 767. https://doi.org/10.3390/genes11070767