VviUCC1 Nucleotide Diversity, Linkage Disequilibrium and Association with Rachis Architecture Traits in Grapevine

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Phenotyping
2.2. Genotypic Data and Sequence Analyses
2.3. Association Analyses
3. Results
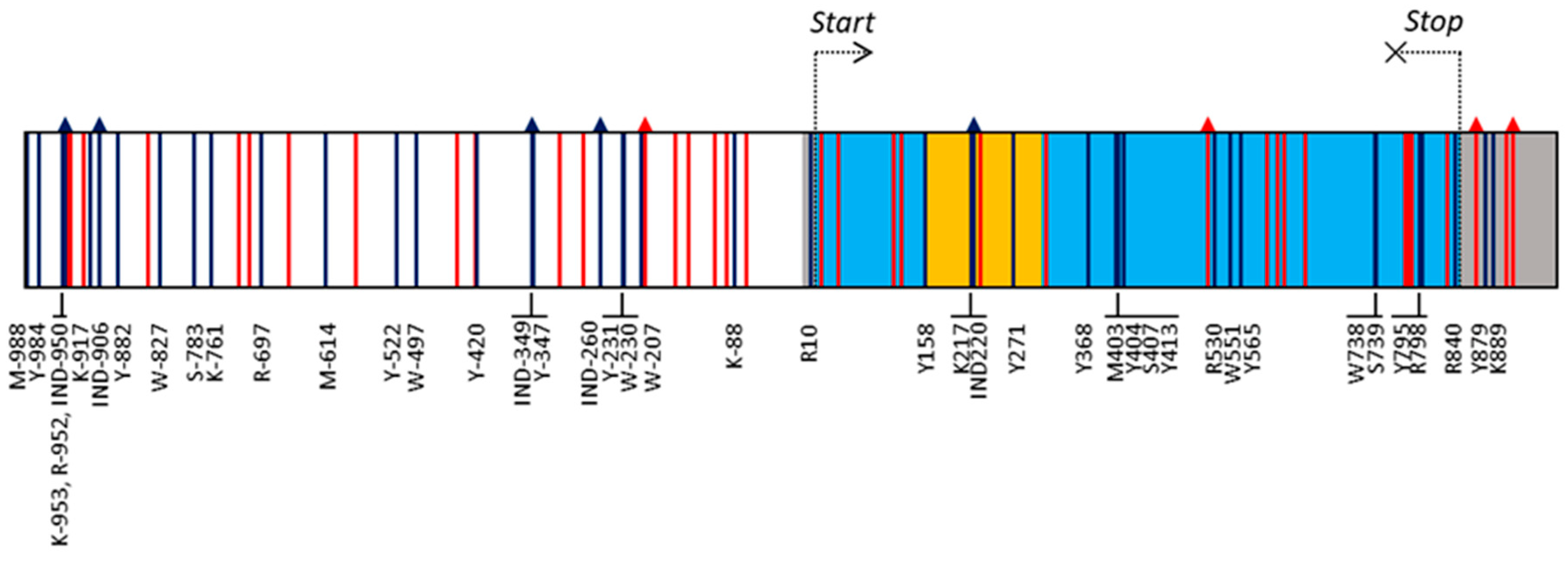
3.1. VviUCC1 Sequence Analysis
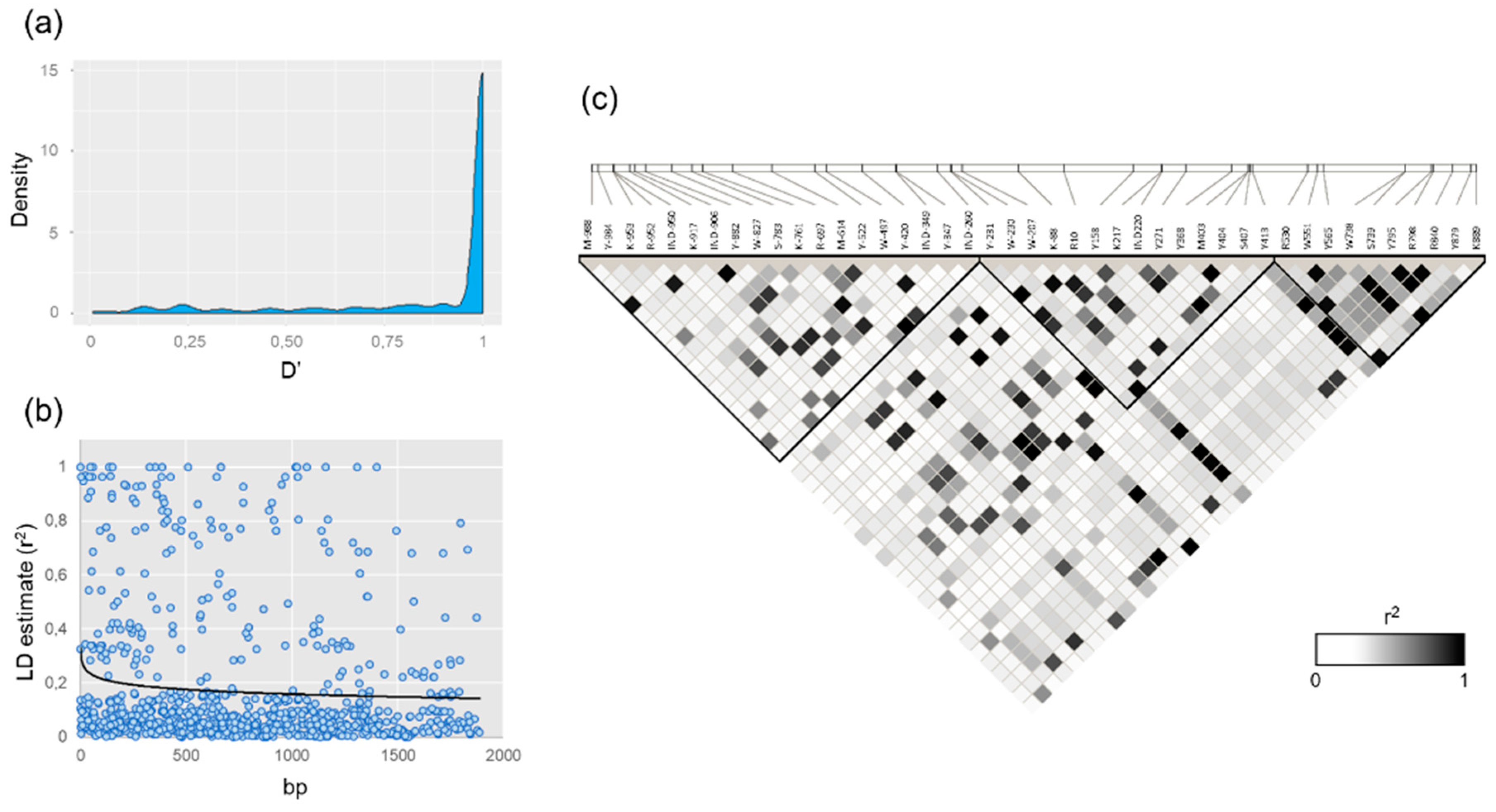
3.2. VviUCC1 Linkage Disequilibrium (LD) Evaluation
3.3. VviUCC1 Association Analyses with Cluster Compactness and Rachis Architecture Traits
3.3.1. Phenotypic Diversity for Cluster Compactness and Rachis Architecture Traits
3.3.2. Marker-Trait Association Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Myles, S.; Boyko, A.R.; Owens, C.L.; Brown, P.J.; Grassi, F.; Aradhya, M.K.; Prins, B.; Reynolds, A.; Chia, J.-M.; Ware, D.; et al. Genetic structure and domestication history of the grape. Proc. Natl. Acad. Sci. USA 2011, 108, 3457–3458. [Google Scholar]
- Wolkovich, E.M.; García de Cortázar-Atauri, I.; Morales-Castilla, I.; Nicholas, K.A.; Lacombe, T. From Pinot to Xinomavro in the world’s future wine-growing regions. Nat. Clim. Chang. 2018, 8, 29–37. [Google Scholar]
- Migicovsky, Z.; Sawler, J.; Gardner, K.M.; Aradhya, M.K.; Prins, B.H.; Schwaninger, H.R.; Bustamante, C.D.; Buckler, E.S.; Zhong, G.-Y.; Brown, P.J.; et al. Patterns of genomic and phenomic diversity in wine and table grapes. Hortic. Res. 2017, 4, 17035. [Google Scholar] [CrossRef] [PubMed]
- Houel, C.; Martin-Magniette, M.L.; Nicolas, S.D.; Lacombe, T.; Le Cunff, L.; Franck, D.; Torregrosa, L.; Conejero, G.; Lalet, S.; This, P.; et al. Genetic variability of berry size in the grapevine (Vitis vinifera L.). Aust. J. Grape Wine Res. 2013, 19, 208–220. [Google Scholar]
- Laucou, V.; Launay, A.; Bacilieri, R.; Lacombe, T.; Adam-Blondon, A.F.; Bérard, A.; Chauveau, A.; de Andrés, M.T.; Hausmann, L.; Ibáñez, J.; et al. Extended diversity analysis of cultivated grapevine Vitis vinifera with 10K genome-wide SNPs. PLoS ONE 2018, 13, e0192540. [Google Scholar] [CrossRef]
- Liang, Z.C.; Duan, S.; sheng, J.; Zhu, S.; Ni, X.; Shao, J.; Liu, C.; Nick, P.; Du, F.; Fan, P.; et al. Whole-genome resequencing of 472 Vitis accessions for grapevine diversity and demographic history analyses. Nat. Commun. 2019, 10, 1190. [Google Scholar] [CrossRef]
- Roscher, R.; Herzog, K.; Kunkel, A.; Kicherer, A.; Topfer, R.; Förstner, W. Automated image analysis framework for high-throughput determination of grapevine berry sizes using conditional random fields. Comput. Electron. Agric. 2014, 100, 148–158. [Google Scholar]
- Tello, J.; Aguirrezábal, R.; Hernaiz, S.; Larreina, B.; Montemayor, M.I.; Vaquero, E.; Ibáñez, J. Multicultivar and multivariate study of the natural variation for grapevine bunch compactness. Aust. J. Grape Wine Res. 2015, 21, 277–289. [Google Scholar]
- Vail, M.E.; Marois, J.J. Grape cluster architecture and the susceptibility of berries to Botrytis cinerea. Phytopathology 1991, 81, 188–191. [Google Scholar]
- Tello, J.; Ibáñez, J. What do we know about grapevine bunch compactness? A state-of-the-art review. Aust. J. Grape Wine Res. 2018, 24, 6–23. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P. Alteration of wine quality caused by Botrytis damages. Vignevini 1983, 10, 48–52. [Google Scholar]
- Molitor, D.; Baus, O.; Hoffmann, L.; Beyer, M. Meteorological conditions determine the thermal-termporal position of the annual Botrytis bunch rot epidemic on Vitis vinifera L. cv. Riesling grapes. OENO ONE 2016, 50, 4. [Google Scholar] [CrossRef]
- Molitor, D.; Rothmeier, M.; Behr, M.; Fischer, S.; Hoffman, L.; Evers, D. Crop cultural and chemical methods to control grey mould on grapes. Vitis 2011, 50, 81–87. [Google Scholar]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Rodríguez-Lorenzo, M.; Cibriáin, J.F.; Sagües, A.; Abad, F.J.; Martínez-Zapater, J.M.; Ibáñez, J. Intra-varietal diversity for agronomic traits in “Garnacha Blanca”. Vitis 2019, 58, 33–35. [Google Scholar]
- Silvestre, J.P.; Roberto, S.R.; Colombo, R.C.; Goncalves, L.S.A.; Koyama, R.; Shahab, M.; Ahmed, S.; De Souza, R.T. Bunch sizing of ‘BRS Nubia’ table grape by inflorescence management, shoot tipping and berry thinning. Sci. Hort. 2017, 225, 764–770. [Google Scholar] [CrossRef]
- Ziegler, M.; Wegmann-Herr, P.; Schmarr, H.-G.; Gök, R.; Winterhalter, P.; Fischer, U. Impact of rootstock, clonal selection, and berry size of Vitis vinifera sp. Riesling on the formation of TDN, vitispiranes, and other volatile compounds. J. Agric. Food Chem. 2020, 68, 3834–3849. [Google Scholar] [CrossRef]
- Tello, J.; Forneck, A. A double-sigmoid model for grapevine bunch compactness development. OENO ONE 2018, 52, 307–316. [Google Scholar] [CrossRef]
- Guilpart, N.; Metay, A.; Gary, C. Grapevine bud fertility and number of berries per bunch are determined by water and nitrogen stress around flowering in the previous year. Eur. J. Agron. 2014, 54, 9–20. [Google Scholar] [CrossRef]
- Li-Mallet, A.; Rabot, A.; Geny, L. Factors controlling inflorescence primordia formation of grapevine: Their role in latent bud fruitfulness? A review. Botany 2016, 94, 147–163. [Google Scholar] [CrossRef]
- Ibáñez, J.; Baroja, E.; Grimplet, J.; Ibáñez, S. Cultivated grapevine displays a great diversity for reproductive performance variables. Crop Breed. Genet. Genom. 2020, 2, e200003. [Google Scholar]
- Tello, J.; Torres-Pérez, R.; Grimplet, J.; Ibáñez, J. Association analysis of grapevine bunch traits using a comprehensive approach. Theor. Appl. Genet. 2016, 129, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Fanizza, G.; Lamaj, F.; Costantini, L.; Chaabane, R.; Grando, M.S. QTL analysis for fruit yield components in table grapes (Vitis vinifera). Theor. Appl. Genet. 2005, 111, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Vezzulli, S.; Doligez, A.; Bellin, D. Molecular mapping of grapevine genes. In The Grape Genome; Cantu, D., Walker, M.A., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 103–136. [Google Scholar]
- Richter, R.; Gabriel, D.; Rist, F.; Topfer, R.; Zyprian, E. Identification of co-located QTLs and genomic regions affecting grapevine cluster architecture. Theor. Appl. Genet. 2018. [Google Scholar] [CrossRef] [PubMed]
- Rossmann, S.; Richter, R.; Sun, H.; Schneeberger, K.; Topfer, R.; Zyprian, E.; Theres, K. Mutations in the miR396 binding site of the growth-regulating factor gene VvGRF4 modulate inflorescence architecture in grapevine. Plant J. 2020, 101, 1234–1248. [Google Scholar] [CrossRef] [PubMed]
- Correa, J.; Mamani, M.; Muñoz-Espinoza, C.; Laborie, D.; Muñoz, C.; Pinto, M.; Hinrichsen, P. Heritability and identification of QTLs and underlying candidate genes associated with the architecture of the grapevine cluster (Vitis vinifera L.). Theor. Appl. Genet. 2014, 127, 1143–1162. [Google Scholar] [CrossRef] [PubMed]
- Molitor, D.; Behr, M.; Hoffman, L.; Evers, D. Benefits and drawbacks of pre-bloom applications of gibberellic acid (GA3) for stem elongation in Sauvignon blanc. S. Afr. J. Enol. Vitic. 2012, 33, 198–202. [Google Scholar] [CrossRef][Green Version]
- Marguerit, E.; Boury, C.; Manicki, A.; Donnart, M.; Butterlin, G.; Nemorin, A.; Wiedemann-Merdinoglu, S.; Merdinoglu, D.; Ollat, N.; Decroocq, S. Genetic dissection of sex determinism, inflorescence morphology and downy mildew resistance in grapevine. Theor. Appl. Genet. 2009, 118, 1261–1278. [Google Scholar] [CrossRef]
- Grimplet, J.; Tello, J.; Laguna, N.; Ibáñez, J. Differences in flower transcriptome between grapevine clones are related to their cluster compactness, fruitfulness and berry size. Front. Plant Sci. 2017, 8, 632. [Google Scholar] [CrossRef]
- Grimplet, J.; Ibanez, S.; Baroja, E.; Tello, J.; Ibáñez, J. Phenotypic, hormonal, and genomic variation among Vitis vinifera clones with different cluster compactness and reproductive performance. Front. Plant Sci. 2019, 9, 1917. [Google Scholar] [CrossRef]
- Nersissian, A.M.; Immoos, C.; Hill, M.G.; Hart, P.J.; Williams, G.; Herrmann, R.G.; Valentine, J.S. Uclacyanins, stellacyanins, and plantacyanins are distinct subfamilies of phytocyanins: Plant-specific mononuclear blue copper proteins. Protein Sci. 1998, 7, 1915–1929. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Hu, W.; Wang, Y.; Liu, B.-Y.; Yan, H.; Xiang, Y. Genome-wide identification, classification, and expression of phytocyanins in Populus trichocarpa. Planta 2018, 247, 1133–1148. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Li, X.; Lv, Y.; Ding, L. Comparative analysis of the phytocyanin gene family in 10 plant species: A focus on Zea mays. Front. Plant Sci. 2015, 6, 515. [Google Scholar] [CrossRef] [PubMed]
- Trupiano, D.; Rocco, M.; Renzone, G.; Scaloni, A.; Viscosi, V.; Chiatante, D.; Scippa, G.S. The proteome of Populus nigra woody root: Response to bending. Ann. Bot. 2012, 110, 415–432. [Google Scholar]
- Ruan, X.-M.; Luo, F.; Li, D.-D.; Zhang, J.; Liu, Z.-H.; Xu, W.-L.; Huang, G.-Q.; Li, X.-B. Cotton BCP genes encoding putative blue copper-binding proteins are functionally expressed in fiber development and involved in response to high-salinity and heavy metal stresses. Physiol. Plant. 2011, 141, 71–83. [Google Scholar]
- Ma, H.; Zhao, H.; Liu, Z.; Zhao, J. The phytocyanin gene family in rice (Oriza sativa L.): Genome-wide identification, classification and transcriptional analysis. PLoS ONE 2011, 6, e25184. [Google Scholar] [CrossRef]
- Drew, J.E.; Gatehouse, J.A. Isolation and characterization of a pea pod cDNA encoding a putative blue copper protein correlated with lignin deposition. J. Exp. Bot. 1997, 45, 1873–1884. [Google Scholar] [CrossRef]
- Li, X.; Yang, X.; Wu, H.X. Transcriptomic profiling of radiata pine branches reveals new insights into reaction wood formation with implications in plant gravitropism. BMC Genom. 2013, 14, 768. [Google Scholar] [CrossRef]
- Kovalchuk, A.; Raffaello, T.; Jaber, E.; Kerio, S.; Ghimire, R.; Lorenz, W.W.; Dean, J.F.; Holopainen, J.K.; Asiegbu, F.O. Activation of defence pathways in Scots pine bark after feeding by pine weevil (Hylobius abietis). BMC Genom. 2015, 16, 352. [Google Scholar] [CrossRef]
- Jamet, E.; Canut, H.; Boudart, G.; Pont-Lezica, R.F. Cell wall proteins: A new insight through proteomics. Trends Plant Sci. 2006, 11, 33–39. [Google Scholar] [CrossRef]
- Fasoli, M.; Dal Santo, S.; Zenoni, S.; Tomielli, G.B.; Farina, L.; Zamboni, A.; Porceddu, A.; Venturini, L.; Bicego, M.; Murino, V.; et al. The grapevine expression atlas reveals a deep transcriptome shift driving the entire plant into a maturation program. Plant Cell 2012, 24, 3489–3505. [Google Scholar] [PubMed]
- Diaz-Riquelme, J.; Martínez-Zapater, J.M.; Carmona, M.J. Transcriptional analysis of tendril and inflorescence development in grapevine (Vitis vinifera L.). PLoS ONE 2014, 9, e92339. [Google Scholar]
- Tello, J.; Torres-Pérez, R.; Grimplet, J.; Carbonell-Bejerano, P.; Martínez-Zapater, J.M.; Ibáñez, J. Polymorphisms and minihaplotypes in the VvNAC26 gene associate with berry size variation in grapevine. BMC Plant Biol. 2015, 15, 253. [Google Scholar] [CrossRef] [PubMed]
- OIV. OIV Descriptor List for Grape Varieties and Vitis Species, 2nd ed.; Organisation Internationale de la Vigne et du Vin: Paris, France, 2009. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- De Andrés, M.T.; Benito, A.; Pérez-Rivera, G.; Ocete, R.; López, M.A.; Gaforio, L.; Muñoz, G.; Cabello, F.; Martinez-Zapater, J.M.; Arroyo-Garcia, R. Genetic diversity of wild grapevine populations in Spain and their genetic relationships with cultivated grapevines. Mol. Ecol. 2012, 21, 800–816. [Google Scholar]
- Vélez, M.D.; Ibáñez, J. Assessment of the uniformity and stability of grapevine cultivars using a set of microsatellite markers. Euphytica 2012, 184, 419–432. [Google Scholar] [CrossRef]
- Lacombe, T.; Boursiquot, J.-M.; Laucou, V.; Di Vecchi Staraz, M.; Péros, J.P.; This, P. Large-scale parentage analysis in an extended set of grapevine cultivars (Vitis vinifera L.). Theor. Appl. Genet. 2013, 126, 401–414. [Google Scholar] [CrossRef]
- Canaguier, A.; Grimplet, J.; Di Gaspero, G.; Scalabrin, S.; Duchene, E.; Choisne, N.; Mohellibi, N.; Guichard, C.; Rombauts, S.; Le Clainche, I.; et al. A new version of the grapevine reference genome assembly (12X.v2) and of its annotation (VCost.v3). Genom. Data 2017, 14, 56–62. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alingment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-performance genomics data visualization and exploration. Brief. Bioinform. 2012, 14, 178–192. [Google Scholar]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.M.; Wang, L.; Land, S.J.; Ruden, D.M.; Liu, X. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain W1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar]
- Bromberg, Y.; Rost, B. SNAP: Predict effect of non-synonymous polymorphisms on function. Nucleic Acids Res. 2007, 35, 3823–3835. [Google Scholar] [CrossRef]
- Choi, Y.A.; Sims, G.E.; Murphy, S.; Miller, J.R.; Chan, A.P. Predicting the functional effect of amino acid substitutions and indels. PLoS ONE 2012, 7, e46688. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Stenberg, M.J.E. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulaturo elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Stephens, M.; Donnelly, P. A comparison of bayesian methods for haplotype reconstruction from population genotype data. Am. J. Hum. Genet. 2003, 73, 1162–1169. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar]
- Barret, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef]
- Duggal, P.; Gillanders, E.M.; Holmes, T.N.; Bailey-Wilson, J.E. Establishing an adjusted p-value threshold to control the family-wide type I error in genome wide association studies. BMC Genom. 2008, 9, 516. [Google Scholar] [CrossRef]
- Wong, D.C.J. Network aggregation improves gene function prediction of grapevine gene co-expression networks. Plant Mol. Biol. 2020. [Google Scholar] [CrossRef]
- Grimplet, J.; Cramer, G.R.; Dickerson, J.A.; Mathiason, K.; Van Hemert, J.; Fennell, A.Y. VitisNet: “Omics” Integration through Grapevine Molecular Networks. PLoS ONE 2009, 4, e8365. [Google Scholar] [CrossRef]
- Yu, J.M.; Pressoir, G.; Briggs, W.H.; Bi, I.V.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnely, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar]
- Wang, J. An estimator for pairwise relatedness using molecular markers. Genetics 2002, 160, 1203–1215. [Google Scholar]
- Pew, J.; Muir, P.H.; Wang, J.; Frasier, T.R. related: An R package for analysing pairwise relatedness from codominant molecular markers. Mol. Ecol. Resour. 2015, 15, 557–561. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar]
- Theiler, R.; Coombe, B.G. Influence of berry growth and growth regulators on the development of grape peduncles in Vitis vinifera L. Vitis 1985, 24, 1–11. [Google Scholar]
- Shavrukov, Y.N.; Dry, I.B.; Thomas, M.R. Inflorescence and bunch architecture development in Vitis vinifera L. Aust. J. Grape Wine Res. 2004, 10, 116–124. [Google Scholar]
- Carvajal-Millán, E.; Carvallo, T.; Orozco, J.A.; Martínez, M.A.; Tapia, I.; Guerrero, V.M.; Rascón-Chu, A.; Llamas, J.; Gardea, A.A. Polyphenol oxidase activity, color changes, and dehydration in table grape rachis during development and storage as affected by N-(2-chloro-4-pyridyl)-N-phenylurea. J. Agric. Food Chem. 2001, 49, 946–951. [Google Scholar]
- Gourieroux, A.M.; Holzapfel, B.P.; McCully, M.E.; Scollary, G.R.; Rogiers, S.Y. Vascular development of the grapevine (Vitis vinifera L.) inflorescence rachis in response to flower number, plant growth regulators and defoliation. J. Plant Res. 2017, 130, 873–883. [Google Scholar] [CrossRef]
- Spigno, G.; Maggi, L.; Amendola, D.; Dragoni, M.; De Faveri, D.M. Influence of cultivar on the lignocellulosic fractionation of grape stalks. Ind. Crop. Prod. 2013, 46, 283–289. [Google Scholar] [CrossRef]
- Riahi, L.; Zoghlami, N.; Dereeper, A.; Laucou, V.; Mliki, A.; This, P. Molecular characterization and evolutionary pattern of the 9-cis-epoxycarotenoid dioxygenase NCED1 gene in grapevine. Mol. Breed. 2013, 32, 253–266. [Google Scholar] [CrossRef]
- Riahi, L.; Zoghlami, N.; Dereeper, A.; Laucou, V.; Mliki, A.; This, P. Single nucleotide polymorphism and haplotype diversity of the gene NAC4 in grapevine. Ind. Crop. Prod. 2013, 43, 718–724. [Google Scholar] [CrossRef]
- Tatarinova, T.V.; Chekalin, E.; Nikolsky, Y.; Bruskin, S.; Chebotarov, D.; McNally, K.L.; Alexandrov, N. Nucleotide diversity analysis highlights functionally important genomic regions. Sci. Rep. 2016, 6, 35730. [Google Scholar] [CrossRef]
- Li, Y.; Vinckenbosch, N.; Tian, G.; Huerta-Sanchez, E.; Jiang, T.; Jiang, H.; Albrechtsen, A.; Andersen, G.; Cao, H.; Korneliussen, T.; et al. Resequencing of 200 human exomes identifies an excess of low-frequency non-synonymous coding variants. Nat. Genet. 2010, 42, 969–972. [Google Scholar] [CrossRef]
- Seifert, S.; Vornam, B.; Finkeldey, R. DNA sequence variation and development of SNP markers in beech (Fagus sylvatica L.). Eur. J. For. Res. 2012, 131, 1761–1770. [Google Scholar] [CrossRef][Green Version]
- Yang, Y.; Cuenca, J.; Wang, N.; Liang, Z.; Sun, H.; Gutierrez, B.; Xi, X.; Arro, J.; Wang, Y.; Fan, P.; et al. A key ‘foxy’ aroma gene is regulated by homology-induced promoter indels in the iconic juice grape ‘Concord’. Hortic. Res. 2020, 7, 67. [Google Scholar] [CrossRef]
- Novina, C.D.; Roy, A.L. Core promoters and transcriptional control. Trends Genet. 1996, 12, 351–355. [Google Scholar] [CrossRef]
- Liu, C.-J. Deciphering the enigma of lignification: Precursor transport, oxidation, and the topochemistry of lignin assembly. Mol. Plant 2012, 5, 304–317. [Google Scholar] [CrossRef]
- Del Río, J.C.; Rencoret, J.; Gutiérrez, A.; Elder, T.; Kim, H.; Ralph, J. Lignin monomers from beyond the canonical monolignol biosynthetic pathway: Another brick in the wall. ACS Sustain. Chem. Eng. 2020, 8, 4997–5012. [Google Scholar] [CrossRef]
- Miao, Y.-C.; Liu, C.-J. ATP-binding cassette-like transporters are involved in the transport of lignin precursors across plasma and vacuolar membranes. Proc. Natl. Acad. Sci. USA 2010, 107, 22728–22733. [Google Scholar] [CrossRef]
- Deluc, L.; Bogs, J.; Walker, A.R.; Ferrier, T.; Decendit, A.; Merillon, J.-M.; Robinson, S.P.; Barrieu, F. The transcription factor VvMYB5b contributes to the regulation of anthocyanin and proanthocyanidin biosynthesis in developing grape berries. Plant Physiol. 2008, 147, 2041–2053. [Google Scholar] [CrossRef]
- Verma, V.; Ravindram, P.; Kumar, P.P. Plant hormone-mediated regulation of stress response. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Zhao, Q.; Dixon, R.A. Transcriptional networks for lignin biosynthesis: More complex than we thought? Trends Plant Sci. 2011, 16, 227–233. [Google Scholar] [CrossRef]
- Zufferey, V.; Spring, J.L.; Voinesco, F.; Viret, O.; Gindro, K. Physiological and histological approaches to study berry shrivel in grapes. J. Int. Sci. Vigne Vin. 2015, 49, 113–125. [Google Scholar] [CrossRef]
- Upadhyay, A.; Maske, S.; Jogaiah, S.; Kadoo, N.Y.; Gupta, V.S. GA3 application in grapes (Vitis vinifera L.) modulates different sets of genes at cluster emergence, full bloom, and berry stage as revealed by RNA sequence-based transcriptome analysis. Funct. Integr. Genom. 2018, 18, 439–455. [Google Scholar] [CrossRef]
- Nakamura, M.; Hori, Y. Postharvest berry drop of seedless berries produced by GA treatment in grape cultivar “Kyoho”. III. Effect of GA on xylem differentiation and lignification of rachis. Tohoku J. Agric. Res. 1984, 34, 55–64. [Google Scholar]
- García-Riojas, M.; Meneses, M.; Oviedo, K.; Carrasco, C.; Defilippi, B.; González-Agüero, M.; León, G.; Hinrichsen, P. Exogenous gibberellic acid application induces the overexpression of key genes for pedicel lignification and an increase in berry drop in table grape. Plant Physiol. Biochem. 2018, 126, 32–38. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trait | Acronym | Description |
|---|---|---|
| Cluster compactness | COMP | Visual compactness of the cluster (O.I.V. rating) |
| Peduncle length | PDULE | Distance from insertion point on the shoot to the first ramification of the cluster |
| Rachis length | RALE | Length of the stalk, peduncle excluded |
| First ramification length | RM1LE | Length of the first ramification of the rachis |
| Second ramification length | RM2LE | Length of the second ramification of the rachis |
| Third ramification length | RM3LE | Length of the third ramification of the rachis |
| Fourth ramification length | RM4LE | Length of the fourth ramification of the rachis |
| Pedicel length | PDILE | Distance from insertion to ramification (15 independent measurements per cluster) |
| W738 Genotype | S739 Genotype | Possible Codon Combinations | Possible Amino Acid Combinations | N Observed Individuals |
|---|---|---|---|---|
| A:A | C:C | ACA | Thr | 4 |
| G:G | AGA | Arg | 2 | |
| C:G | ACA/AGA | Thr/Arg | 4 | |
| T:T | C:C | TCA | Ser | 46 |
| A:T | C:C | ACA/TCA | Thr/Ser | 33 |
| C:G | ACA1/TCA/AGA/TGA 1 | Thr 2/Ser/Arg/STOP 2 | 25 |
| Trait | 2011 | 2012 | 2013 | H2 (%) 1 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± s.d. | Mx | Mn | Mean ± s.d. | Mx | Mn | Mean ± s.d. | Mx | Mn | ||
| COMP (OIV rating) | 5.6 ± 1.4 | 9 | 2.6 | 4.9 ± 1.5 | 8.6 | 1.6 | 5.4 ± 1.5 | 8.8 | 2.4 | 32.1 |
| PDULE (cm) | 4.6 ± 1.1 | 8.0 | 2.3 | 3.8 ± 0.9 | 7.0 | 1.9 | 4.9 ± 1.3 | 8.3 | 2.6 | 38.9 |
| RALE (cm) | n.a. | n.a. | n.a. | 11.3 ± 3.8 | 24.3 | 5.3 | 13.7 ± 4.5 | 27 | 5.2 | - |
| RM1LE (cm) | 4.8 ± 2 | 10.3 | 1.6 | 4.2 ± 2 | 10.7 | 1.0 | 5.2 ± 2.6 | 13.1 | 1.2 | 47.2 |
| RM2LE (cm) | 4.2 ± 1.8 | 10.7 | 1.4 | 3.5 ± 1.7 | 9.2 | 1.0 | 4.5 ± 2.3 | 10.8 | 1.1 | 45.1 |
| RM3LE (cm) | n.a. | n.a. | n.a. | 2.7 ± 1.4 | 8.2 | 0.4 | 3.2 ± 1.8 | 8.6 | 0.6 | - |
| RM4LE (cm) | n.a. | n.a. | n.a. | 2.3 ± 1.3 | 7.5 | 0.4 | 2.7 ± 1.5 | 7.2 | 0.4 | - |
| PDILE (mm) | 6.9 ± 1.2 | 10.7 | 4.4 | 5.6 ± 0.8 | 8.1 | 3.0 | 6.9 ± 1.3 | 10.7 | 3.8 | 31.6 |
| Polymorphism | Trait | 2011 | 2012 | 2013 | BLUP | ||||
|---|---|---|---|---|---|---|---|---|---|
| p-Value 1 | Trait (%) | p-Value 1 | Trait (%) | p-Value 1 | Trait (%) | p-Value 1 | Trait (%) | ||
| Y-984 | COMP | 8.3 × 10−4 ** | 14.1 | 3.4 × 10−3 * | 11.1 | 2.9 × 10−2 NS | - | 6.9 × 10−4 ** | 13.2 |
| RM1LE | 4.1 × 10−2 NS | - | 9.9 × 10−2 NS | - | 5.3 × 10−3 * | 10 | 7.0 × 10−3 * | 8.3 | |
| RM2LE | 1.7 × 10−2 NS | - | 2.1 × 10−1 NS | - | 1.2 × 10−2 * | 8.2 | 1.0 × 10−2 * | 7.4 | |
| RM3LE | n.a | n.a | 3.5 × 10−1 NS | - | 2.3 × 10−3 ** | 11.6 | n.a | n.a | |
| RM4LE | n.a | n.a | 2.8 × 10−1 NS | - | 1.6 × 10−3 ** | 12.9 | n.a | n.a | |
| Y-231/K-88 | COMP | 3.1 × 10−3 ** | 11.3 | 2.3 × 10−3 ** | 11.9 | 4.8 × 10−2 NS | - | 6.9 × 10−4 ** | 13.3 |
| RM1LE | 1.0 × 10−2 * | 9.3 | 4.5 × 10−2 NS | - | 1.3 × 10−4 *** | 17.8 | 1.3 × 10−3 ** | 11.2 | |
| RM2LE | 1.3 × 10−2 * | 8.5 | 7.6 × 10−2 NS | - | 7.9 × 10−5 *** | 19 | 2.5 × 10−3 ** | 9.9 | |
| RM3LE | n.a | n.a | 3.1 × 10−1 NS | - | 1.8 × 10−4 *** | 17 | n.a | n.a | |
| RM4LE | n.a | n.a | 2.5 × 10−1 NS | - | 3.6 × 10−4 ** | 16.1 | n.a | n.a | |
| Y158 | COMP | 2.1 × 10−3 ** | 12.1 | 3.1 × 10−3 ** | 11.3 | 4.3 × 10−2 NS | - | 7.7 × 10−4 ** | 13 |
| RM1LE | 6.1 × 10−2 NS | - | 8.9 × 10−2 NS | - | 1.7 × 10−3 ** | 12.3 | 1.7 × 10−2 NS | - | |
| RM2LE | 9.1 × 10−2 NS | - | 1.4 × 10−1 NS | - | 1.1 × 10−3 ** | 13.1 | 2.7 × 10−2 NS | - | |
| RM3LE | n.a | n.a | 6.1 × 10−1 NS | - | 1.4 × 10−3 ** | 12.6 | n.a | n.a | |
| RM4LE | n.a | n.a | 3.7 × 10−1 NS | - | 1.8 × 10−3 ** | 12.7 | n.a | n.a | |
| K217 | COMP | 1.0 × 10−2 * | 8.9 | 4.3 × 10−3 * | 10.7 | 4.9 × 10−2 NS | - | 1.0 × 10−3 ** | 12.5 |
| RM1LE | 8.8 × 10−2 NS | - | 1.2 × 10−1 NS | - | 4.1 × 10−3 * | 10.5 | 3.2 × 10−2 NS | - | |
| RM2LE | 4.0 × 10−2 NS | - | 1.3 × 10−1 NS | - | 2.6 × 10−3 ** | 11.4 | 2.8 × 10−2 NS | - | |
| RM3LE | n.a | n.a | 6.1 × 10−1 NS | - | 3.1 × 10−3 ** | 11 | n.a | n.a | |
| RM4LE | n.a | n.a | 2.4 × 10−1 NS | - | 7.0 × 10−3 * | 9.7 | n.a | n.a | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tello, J.; Torres-Pérez, R.; Flutre, T.; Grimplet, J.; Ibáñez, J. VviUCC1 Nucleotide Diversity, Linkage Disequilibrium and Association with Rachis Architecture Traits in Grapevine. Genes 2020, 11, 598. https://doi.org/10.3390/genes11060598
Tello J, Torres-Pérez R, Flutre T, Grimplet J, Ibáñez J. VviUCC1 Nucleotide Diversity, Linkage Disequilibrium and Association with Rachis Architecture Traits in Grapevine. Genes. 2020; 11(6):598. https://doi.org/10.3390/genes11060598
Chicago/Turabian StyleTello, Javier, Rafael Torres-Pérez, Timothée Flutre, Jérôme Grimplet, and Javier Ibáñez. 2020. "VviUCC1 Nucleotide Diversity, Linkage Disequilibrium and Association with Rachis Architecture Traits in Grapevine" Genes 11, no. 6: 598. https://doi.org/10.3390/genes11060598
APA StyleTello, J., Torres-Pérez, R., Flutre, T., Grimplet, J., & Ibáñez, J. (2020). VviUCC1 Nucleotide Diversity, Linkage Disequilibrium and Association with Rachis Architecture Traits in Grapevine. Genes, 11(6), 598. https://doi.org/10.3390/genes11060598