Sustentacular Cell Enwrapment of Olfactory Receptor Neuronal Dendrites: An Update

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
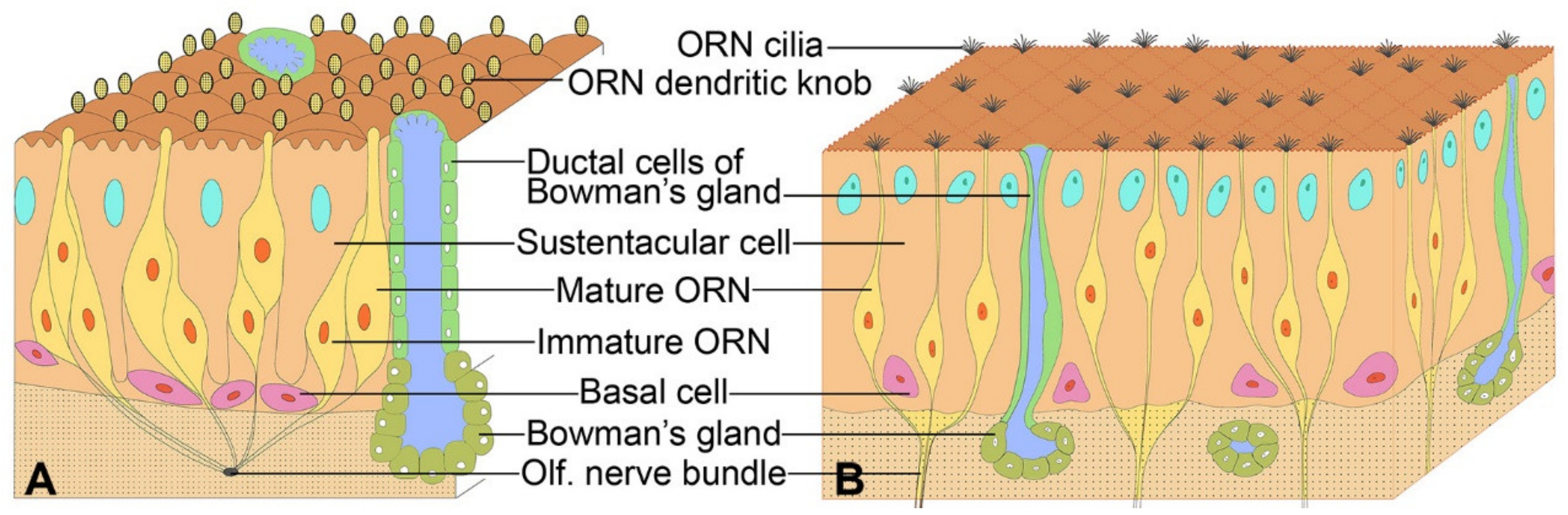
2. Early Cytological and Cytoarchitectonic Studies of the Olfactory Epithelium
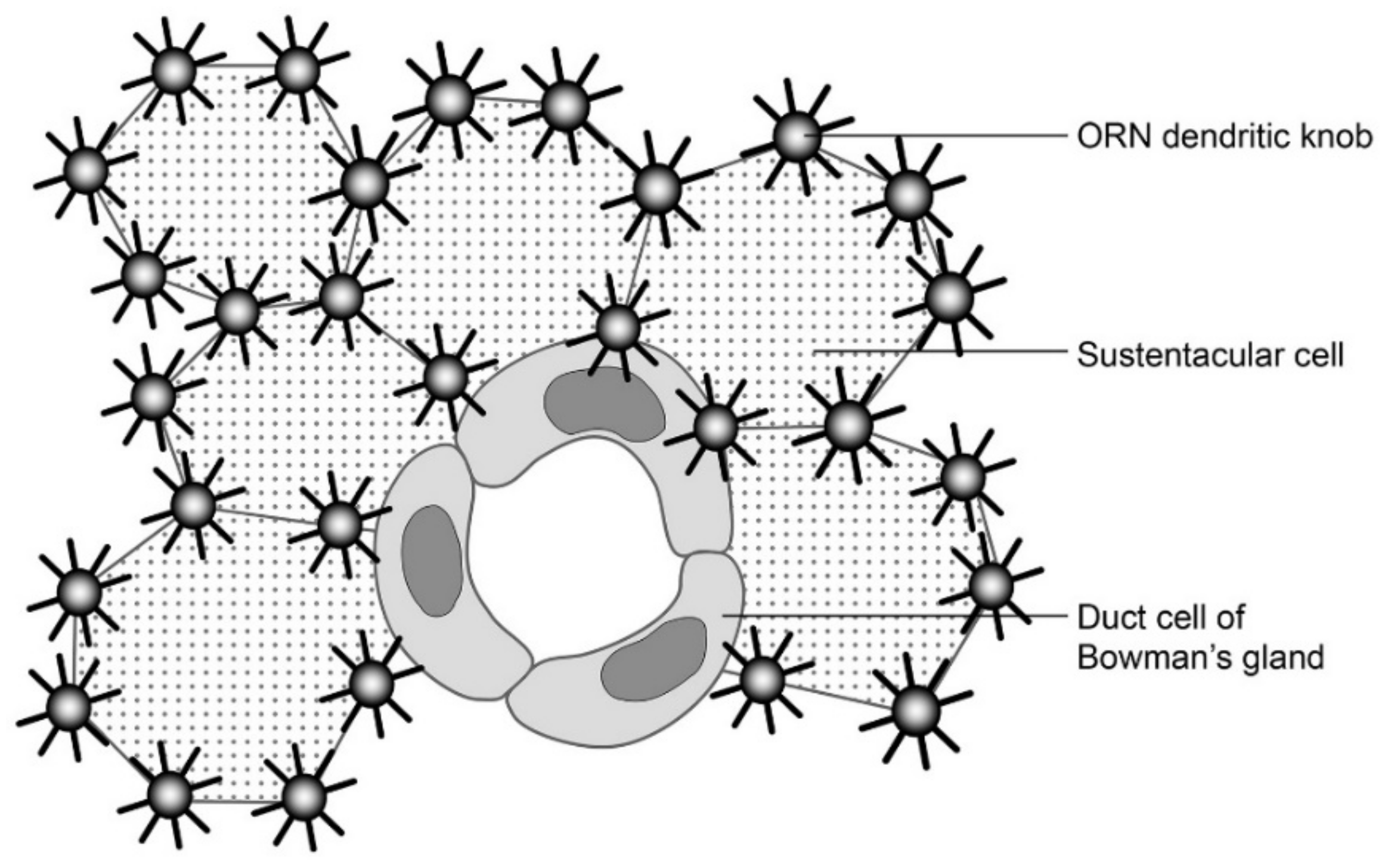
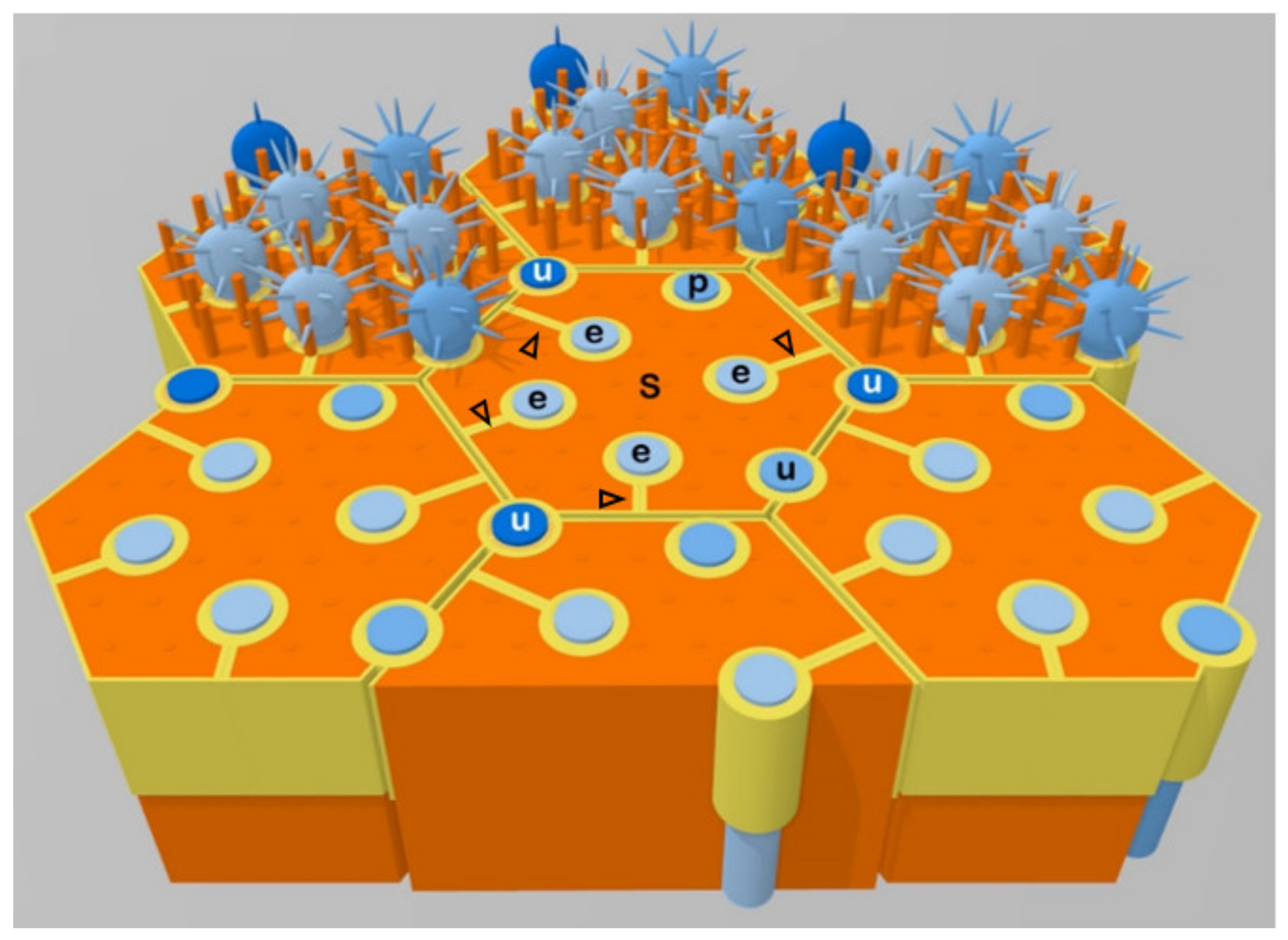
3. A Majority of Olfactory Receptor Neuron Dendrites are Enwrapped by Sustentacular Cells
4. The Enwrapment, Sideways Migration and Terminal Maturation of ORN Dendrites
5. Comparison of ORN Dendritic Enwrapment, Neurite Myelination, Remak’s Bundling, and Possible Molecular Mechanisms
6. Some Other Questions and Future Studies
Funding
Conflicts of Interest
References
- Schultze, M. Über die endigungsweise des geruchsnerven und die epithelialgebilde der nasenschleimhaut. Monatsberichte der Konigl Preußs. Akad der Wissen Berlin 1856, 21, 504–515. [Google Scholar]
- Allison, A.C. The morphology of the olfactory system in vertebrates. Biol. Rev. 1953, 28, 195–244. [Google Scholar] [CrossRef]
- Breipohl, W.; Laugwitz, H.J.; Bornfeld, N. Topological relations between the dendrites of olfactory sensory cells and sustentacular cells in different vertebrates. An ultrastructural study. J. Anat. 1974, 117, 89–94. [Google Scholar] [PubMed]
- Suzuki, Y.; Takeda, M.; Farbman, A.I. Supporting cells as phagocytes in the olfactory epithelium after bulbectomy. J. Comp. Neurol. 1996, 376, 509–517. [Google Scholar] [CrossRef]
- Schwob, J.E. Neural regeneration and the peripheral olfactory system. Anat. Rec. 2002, 269, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Menco, B.P.H.M.; Morrison, E.E. Morphology of the mammalian olfactory epithelium: Form, fine structure and pathology. In Handbook of Olfaction and Gustation, 2nd ed.; Doty, R., Ed.; Marcel Dekker: New York, NY, USA, 2003; pp. 17–49. [Google Scholar]
- Asan, E.; Drenckhahn, D. Immunocytochemical characterization of two types of microvillar cells in rodent olfactory epithelium. Histochem. Cell Biol. 2005, 123, 157–168. [Google Scholar] [CrossRef]
- Elsaesser, R.; Paysan, J. The sense of smell, its signalling pathways, and the dichotomy of cilia and microvilli in olfactory sensory cells. BMC Neurosci. 2007, 8, S1. [Google Scholar] [CrossRef]
- Steinke, A.; Meier-Stiegen, S.; Drenckhahn, D.; Asan, E. Molecular composition of tight and adherens junctions in the rat olfactory epithelium and fila. Histochem. Cell Biol. 2008, 130, 339. [Google Scholar] [CrossRef]
- Hegg, C.C.; Irwin, M.; Lucero, M.T. Calcium store-mediated signaling in sustentacular cells of the mouse olfactory epithelium. Glia 2009, 57, 634–644. [Google Scholar] [CrossRef]
- Liang, F. Olfactory receptor neuronal dendrites become mostly intra-sustentacularly enwrapped upon maturity. J. Anat. 2018, 232, 674–685. [Google Scholar] [CrossRef]
- Buck, L.B. Unraveling the sense of smell (Nobel lecture). Angew. Chem. Int. Ed. Engl. 2005, 44, 6128–6140. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; von Campenhause, H.; Yoshihara, Y. Zonal organization of the mammalian main and accessory olfactory systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Takagi, S.F. Secretion and electrogenesis of the supporting cell in the olfactory epithelium. J. Physiol. 1974, 242, 353–370. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Takeda, M.; Obara, N.; Suzuki, N.; Takeichi, N. Olfactory epithelium consisting of supporting cells and horizontal basal cells in the posterior nasal cavity of mice. Cell Tissue Res. 2000, 299, 313–325. [Google Scholar] [CrossRef]
- Hassenklöver, T.; Kurtanska, S.; Bartoszek, I.; Junek, S.; Schild, D.; Manzini, I. Nucleotide-induced Ca2+ signaling in sustentacular supporting cells of the olfactory epithelium. Glia 2008, 56, 1614–1624. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Tang, J.H.; Ling, E.A.; Wu, Y.; Liang, F. Juxtanodin in the rat olfactory epithelium: Specific expression in sustentacular cells and preferential subcellular positioning at the apical junctional belt. Neuroscience 2009, 161, 249–258. [Google Scholar] [CrossRef]
- Graziadei, P.P.C.; Graziadei, G.A.M. Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J. Neurocytol. 1979, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Shipley, M.T.; Ennis, M.; Puche, A.C. Olfactory system. In The Rat Nervous System, 3rd ed.; Paxinos, G., Ed.; Elsevier: San Diego, CA, USA, 2004; pp. 923–964. [Google Scholar]
- Ashwell, K. The olfactory system. In The Mouse Nervous System; Watson, C., Paxinos, G., Puelles, L., Eds.; Academic Press: Amsterdam, The Netherland, 2012; pp. 653–660. [Google Scholar]
- Brann, J.H.; Firestein, S.J. A lifetime of neurogenesis in the olfactory system. Front. Neurosci. 2014, 8, 182. [Google Scholar] [CrossRef]
- Kwon, B.S.; Kim, M.K.; Kim, W.H.; Pyo, J.S.; Cheon, Y.H.; Cha, C.I.; Nam, S.Y.; Baik, T.K.; Lee, B.L. Age-related changes in microvillar cells of rat olfactory epithelium. Neurosci. Lett. 2005, 378, 65–69. [Google Scholar] [CrossRef]
- Montani, G.; Tonelli, S.; Elsaesser, R.; Paysan, J.; Tirindelli, R. Neuropeptide Y in the olfactory microvillar cells. Eur. J. Neurosci. 2006, 24, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Lemons, K.; Fu, Z.; Aoudé, I.; Ogura, T.; Sun, J.; Chang, J.; Mbonu, K.; Matsumoto, I.; Arakawa, H.; Lin, W. Lack of TRPM5-expressing microvillous cells in mouse main olfactory epithelium leads to impaired odor-evoked responses and olfactory-guided behavior in a challenging chemical environment. eNeuro 2017, 4, ENEURO.0135-17.2017. [Google Scholar] [CrossRef] [PubMed]
- Genovese, F.; Tizzano, M. Microvillous cells in the olfactory epithelium express elements of the solitary chemosensory cell transduction signaling cascade. PLoS ONE 2018, 13, e0202754. [Google Scholar] [CrossRef] [PubMed]
- Graziadei, P.P.C. The olfactory mucosa of vertebrates. In Olfaction. Handbook of Sensory Physiology; Beidler, L.M., Ed.; Springer: Berlin, Germany, 1971; Volume 4/1. [Google Scholar]
- Morrison, E.E.; Costanzo, R.M. Morphology of olfactory epithlium in humans and other vertebrates. Microsc. Res. Tech. 1992, 23, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Takahashi, S.; Ushiki, T. Cytoarchitecture of the normal rat olfactory epithelium: Light and scanning electron microscopic studies. Arch. Histol. Cytol. 2004, 67, 159–170. [Google Scholar] [CrossRef]
- Ma, M.; Shepherd, G.M. Functional mosaic organization of mouse olfactory receptor neurons. Proc. Natl. Acad. Sci. USA 2000, 97, 12869–12874. [Google Scholar] [CrossRef]
- Barral, J.-P.; Croibier, A. Chapter 10: Olfactory nerve. In Manual Therapy for the Cranial Nerves; Churchill Livingstone: London, UK, 2009; p. 61. ISBN 9780702031007. [Google Scholar]
- Farbman, A.I. Cell Biology of Olfaction; Cambridge University Press: New York, UK, USA, 1992. [Google Scholar]
- Smutzer, G.S.; Doty, R.L.; Arnold, S.E.; Trojanowski, J.Q. Olfactory system neuropathology in Alzheimer’s disease, Parkinson’s disease, and schizophrenia. In Handbook of Olfaction and Gustation, 2nd ed.; Doty, R., Ed.; Marcel Dekker: New York, USA, 2003; pp. 503–523. [Google Scholar]
- Standring, S. Gray’s Anatomy: The Anatomical Basis of Clinical Practice, 40th ed.; Churchill-Livingstone: London, UK; Elsevier: Amsterdam, The Netherlands, 2008; pp. 552–553. [Google Scholar]
- Salazar, I.; Sanchez-Quinteiro, P.; Barrios, A.W.; López Amado, M.; Vega, J.A. Anatomy of the olfactory mucosa. Handb. Clin. Neurol. 2019, 164, 47–65. [Google Scholar] [CrossRef]
- Menco, B.P.H.M. Qualitative and quantitative freeze-fracture studies on olfactory and nasal respiratory epithelial surfaces of frog, ox, rat, and dog III. Tight-junctions. Cell Tissue Res. 1980, 211, 361–373. [Google Scholar] [CrossRef]
- Wolburg, H.; Wolburg-Buchholz, K.; Sam, H.; Horvát, S.; Deli, M.A.; Mack, A.F. Epithelial and endothelial barriers in the olfactory region of the nasal cavity of the rat. Histochem. Cell Biol. 2008, 130, 127–140. [Google Scholar] [CrossRef]
- Moran, D.T.; Rowley, J.C., 3rd; Jafek, B.W.; Lovell, M.A. The fine structure of the olfactory mucosa in man. J. Neurocytol. 1982, 11, 721–746. [Google Scholar] [CrossRef]
- Vogalis, F.; Hegg, C.C.; Lucero, M.T. Electrical coupling in sustentacular cells of the mouse olfactory epithelium. J. Neurophysiol. 2005, 94, 1001–1012. [Google Scholar] [CrossRef]
- Zhang, C.B. Gap junctions in olfactory neurons modulate olfactory sensitivity. BMC Neurosci. 2010, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Roskams, A.J.; Cai, X.; Ronnet, G.V. Expression of neuron-specific beta-III tubulin during olfactory neurogenesis in the embryonic and adult rat. Neuroscience 1998, 83, 191–200. [Google Scholar] [CrossRef]
- Czesnik, D.; Schild, D.; Kuduz, J.; Manzini, I. Cannabinoid action in the olfactory epithelium. Proc. Natl. Acad. Sci. USA 2007, 104, 2967–2972. [Google Scholar] [CrossRef] [PubMed]
- Breunig, E.; Manzini, I.; Piscitelli, F.; Gutermann, B.; Di Marzo, V.; Schild, D.; Czesnik, D. The endocannabinoid 2-arachidonoyl-glycerol controls odor sensitivity in larvae of Xenopus laevis. J. Neurosci. 2010, 30, 8965–8973. [Google Scholar] [CrossRef]
- Ogura, T.; Szebenyi, S.A.; Krosnowski, K.; Sathyanesan, A.; Jackson, J.; Lin, W. Cholinergic microvillous cells in the mouse main olfactory epithelium and effect of acetylcholine on olfactory sensory neurons and supporting cells. J. Neurophysiol. 2011, 106, 1274–1287. [Google Scholar] [CrossRef]
- Hutch, C.R.; Hegg, C.C. Cannabinoid receptor signaling induces proliferation but not neurogenesis in the mouse olfactory epithelium. Neurogenesis 2016, 3, e1118177. [Google Scholar] [CrossRef]
- Fletcher, R.B.; Das, D.; Gadye, L.; Street, K.N.; Baudhuin, A.; Wagner, A.; Cole, M.B.; Flores, Q.; Choi, Y.G.; Yosef, N.; et al. Deconstructing olfactory stem cell trajectories at single-cell resolution. Cell Stem Cell 2017, 20, 817–830. [Google Scholar] [CrossRef]
- Coleman, J.H.; Lin, B.; Louie, J.D.; Peterson, J.; Lane, R.P.; Schwob, J.E. Spatial determination of neuronal diversification in the olfactory epithelium. J. Neurosci. 2019, 39, 814–832. [Google Scholar] [CrossRef]
- Li, H.; Li, T.; Horns, F.; Li, J.; Xie, Q.; Xu, C.; Wu, B.; Kebschull, J.M.; McLaughlin, C.N.; Kolluru, S.S.; et al. Single-cell transcriptomes reveal diverse regulatory strategies for olfactory receptor expression and axon targeting. Curr. Biol. 2020, 30, 1189–1198. [Google Scholar] [CrossRef]
- Craig, A.D. How do you feel? Interoception: The sense of the physiological condition of the body. Nat. Rev. Neurosci. 2002, 3, 655–666. [Google Scholar] [CrossRef]
- Murinson, B.B.; Griffin, J.W. C-fiber structure varies with location in peripheral nerve. J. Neuropathol. Exp. Neurol. 2004, 63, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Harty, B.L.; Monk, K.R. Unwrapping the unappreciated: Recent progress in Remak Schwann cell biology. Curr. Opin. Neurobiol. 2017, 47, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.E.; Smutzer, G.S.; Trojanowski, J.Q.; Moberg, P.J. Cellular and molecular neuropathology of the olfactory epithelium and central olfactory pathways in Alzheimer’s disease and schizophrenia. Ann. N. Y. Acad. Sci. 1998, 855, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L. Olfaction in Parkinson’s disease and related disorders. Neurobiol. Dis. 2012, 46, 527–552. [Google Scholar] [CrossRef] [PubMed]
- Buron, E.; Bulbena, A. Olfaction in affective and anxiety disorders: A review of the literature. Psychopathology 2013, 46, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.; Eckert, A.; Woitalla, D.; Ellrichmann, G.; Turewicz, M.; Stephan, C.; Eisenacher, M.; May, C.; Meyer, H.E.; Brüning, T.; et al. Diagnostic value of the impairment of olfaction in Parkinson’s disease. PLoS ONE 2013, 8, e64735. [Google Scholar] [CrossRef]
- Seligman, S.C.; Kamath, V.; Giovannetti, T.; Arnold, S.E.; Moberg, P.J. Olfaction and apathy in Alzheimer’s disease, mild cognitive impairment, and healthy older adults. Aging Ment. Health 2013, 17, 564–570. [Google Scholar] [CrossRef]
- Auster, T.L.; Cohen, A.S.; Callaway, D.A.; Brown, L.A. Objective and subjective olfaction across the schizophrenia spectrum. Psychiatry 2014, 77, 57–66. [Google Scholar] [CrossRef]
- Doty, R.L.; Hawkes, C.H.; Good, K.P.; Duda, J.E. Odor perception and neuropathology in neurodegenerative diseases and schizophrenia. In Handbook of Olfaction and Gustation; Doty, R.L., Ed.; Wiley: Hoboken, NJ, USA, 2015; pp. 93–108. [Google Scholar]
- Field, T. Smell and taste dysfunction as early markers for neurodegenerative and neuropsychiatric diseases. J. Alzheimers Dis. Parkinsonism 2015, 5, 186. [Google Scholar]
- Holbrook, E.H.; Wu, E.; Curry, W.T.; Lin, D.T.; Schwob, J.E. Immunohistochemical characterization of human olfactory tissue. Laryngoscope 2011, 121, 1687–1701. [Google Scholar] [CrossRef] [PubMed]
- Melendez-Vasquez, C.V.; Rios, J.C.; Zanazzi, G.; Lambert, S.; Bretscher, A.; Salzer, J.L. Nodes of Ranvier form in association with ezrin-radixin-moesin (ERM)-positive Schwann cell processes. Proc. Natl. Acad. Sci. USA 2001, 98, 1235–1240. [Google Scholar] [CrossRef]
- Gatto, C.L.; Walker, B.J.; Lambert, S. Local ERM activation and dynamic growth cones at Schwann cell tips implicated in efficient formation of nodes of Ranvier. J. Cell Biol. 2003, 162, 489–498. [Google Scholar] [CrossRef]
- Nawaz, S.; Sánchez, P.; Schmitt, S.; Snaidero, N.; Mitkovski, M.; Velte, C.; Brückner, B.R.; Alexopoulos, I.; Czopka, T.; Jung, S.Y.; et al. Actin filament turnover drives leading edge growth during myelin sheath formation in the central nervous system. Dev. Cell 2015, 34, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Samanta, J.; Salzer, J.L. Myelination: Actin disassembly leads the way. Dev. Cell 2015, 34, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Zuchero, J.B.; Fu, M.M.; Sloan, S.A.; Ibrahim, A.; Olson, A.; Zaremba, A.; Dugas, J.C.; Wienbar, S.; Caprariello, A.V.; Kantor, C.; et al. CNS myelin wrapping is driven by actin disassembly. Dev. Cell 2015, 34, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Maurya, D.K.; Henriques, T.; Marini, M.; Pedemonte, N.; Galietta, L.J.; Rock, J.R.; Harfe, B.D.; Menini, A. development of the olfactory epithelium and nasal glands in TMEM16A-/- and TMEM16A+/+ mice. PLoS ONE 2015, 10, e0129171. [Google Scholar] [CrossRef]
- Zhang, B.; Cao, Q.; Guo, A.; Chu, H.; Chan, Y.G.; Buschdorf, J.P.; Low, B.C.; Ling, E.A.; Liang, F. Juxtanodin: An oligodendroglial protein that promotes cellular arborization and 2’,3’-cyclic nucleotide-3’-phosphodiesterase trafficking. Proc. Natl. Acad. Sci. USA 2005, 102, 11527–11532. [Google Scholar] [CrossRef]
- Brockschnieder, D.; Sabanay, H.; Riethmacher, D.; Peles, E. Ermin, a myelinating oligodendrocyte-specific protein that regulates cell morphology. J. Neurosci. 2006, 26, 757–762. [Google Scholar] [CrossRef]
- Meng, J.; Xia, W.; Tang, J.H.; Tang, B.L.; Liang, F. Dephosphorylation-dependent inhibitory activity of juxtanodin on filamentous actin disassembly. J. Biol. Chem. 2010, 285, 28838–28849. [Google Scholar] [CrossRef]
- Ruskamo, S.; Chukhlieb, M.; Vahokoski, J.; Bhargav, S.P.; Liang, F.; Kursula, I.; Kursula, P. Juxtanodin is an intrinsically disordered F-actin-binding protein. Sci. Rep. 2012, 2, 899. [Google Scholar] [CrossRef]
- Liang, F.; Hwang, J.H.; Tang, N.W.; Hunziker, W. Juxtanodin in retinal pigment epithelial cells: Expression and biological activities in regulating cell morphology and actin cytoskeleton organization. J. Comp. Neurol. 2018, 526, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, B.; Defoe, D.M.; Besharse, J.C. Membrane turnover in rod photoreceptors: Ensheathment and phagocytosis of outer segment distal tips by pseudopodia of the retinal pigment epithelium. Proc. R. Soc. Lond. B Biol. Sci. 1987, 230, 339–354. [Google Scholar] [PubMed]
- Buck, L.B.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 157–167. [Google Scholar] [CrossRef]
- Fleischer, J.; Breer, H.; Strotmann, J. Mammalian olfactory receptors. Front. Cell. Neurosci. 2009, 3, 9. [Google Scholar] [CrossRef]
- Johnson, M.A.; Tsai, L.; Roy, D.S.; Valenzuela, D.H.; Mosley, C.; Magklara, A.; Lomvardas, S.; Liberles, S.D.; Barnea, G. Neurons expressing trace amine-associated receptors project to discrete glomeruli and constitute an olfactory subsystem. Proc. Natl. Acad. Sci. USA 2012, 109, 13410–13415. [Google Scholar] [CrossRef]
- Ihara, S.; Yoshikawa, K.; Touhara, K. Chemosensory signals and their receptors in the olfactory neural system. Neuroscience 2013, 254, 45–60. [Google Scholar] [CrossRef]
- Bear, D.M.; Lassance, J.M.; Hoekstra, H.E.; Datta, S.R. The evolving neural and genetic architecture of vertebrate olfaction. Curr. Biol. 2016, 26, R1039–R1049. [Google Scholar] [CrossRef]



© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, F. Sustentacular Cell Enwrapment of Olfactory Receptor Neuronal Dendrites: An Update. Genes 2020, 11, 493. https://doi.org/10.3390/genes11050493
Liang F. Sustentacular Cell Enwrapment of Olfactory Receptor Neuronal Dendrites: An Update. Genes. 2020; 11(5):493. https://doi.org/10.3390/genes11050493
Chicago/Turabian StyleLiang, Fengyi. 2020. "Sustentacular Cell Enwrapment of Olfactory Receptor Neuronal Dendrites: An Update" Genes 11, no. 5: 493. https://doi.org/10.3390/genes11050493
APA StyleLiang, F. (2020). Sustentacular Cell Enwrapment of Olfactory Receptor Neuronal Dendrites: An Update. Genes, 11(5), 493. https://doi.org/10.3390/genes11050493