Effects of 5′-3′ Exonuclease Xrn1 on Cell Size, Proliferation and Division, and mRNA Levels of Periodic Genes in Cryptococcus neoformans

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Construction of xrn1∆ and Complementation Strains xrn1∆-C
2.3. Southern Blot Analysis
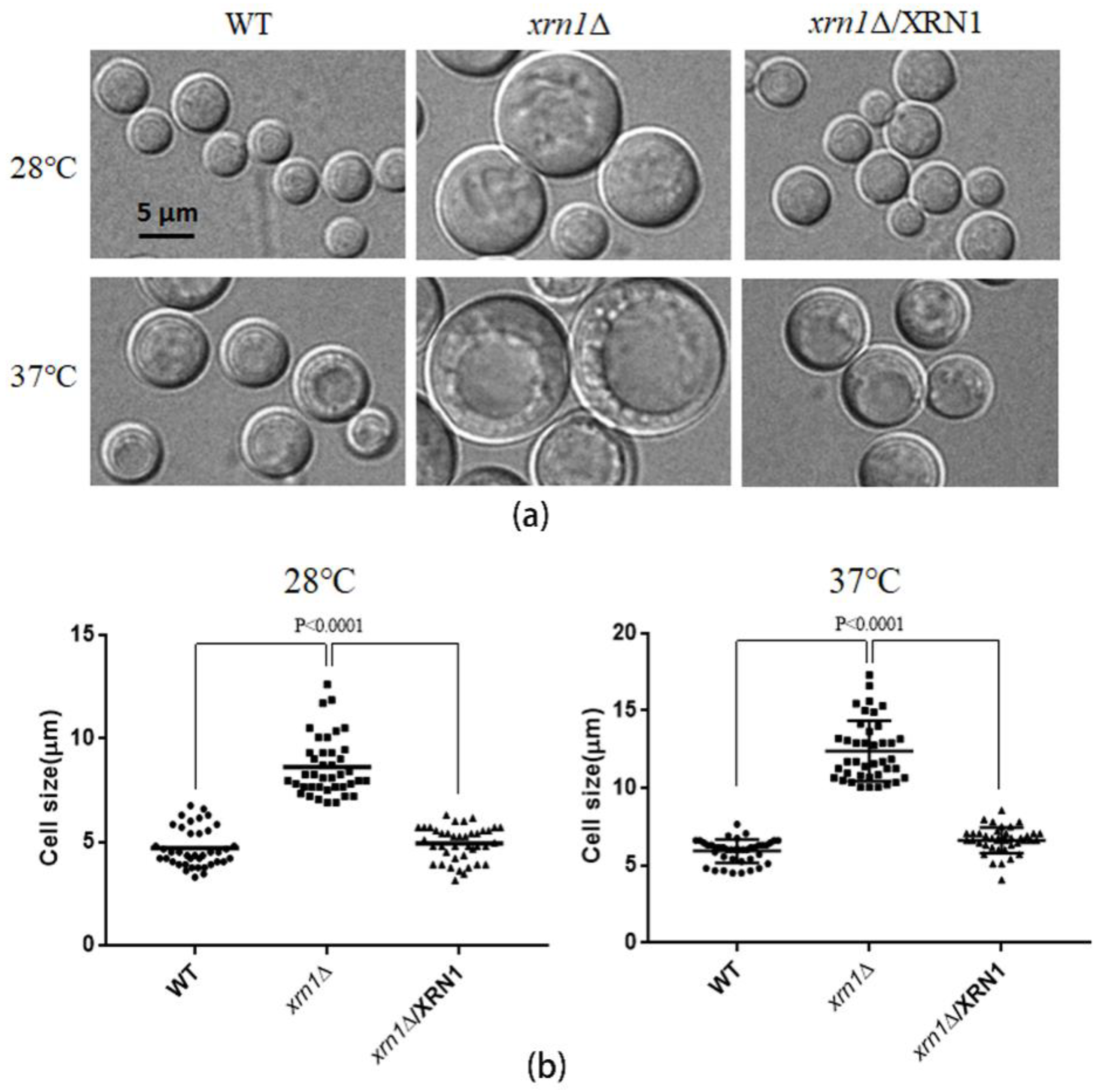
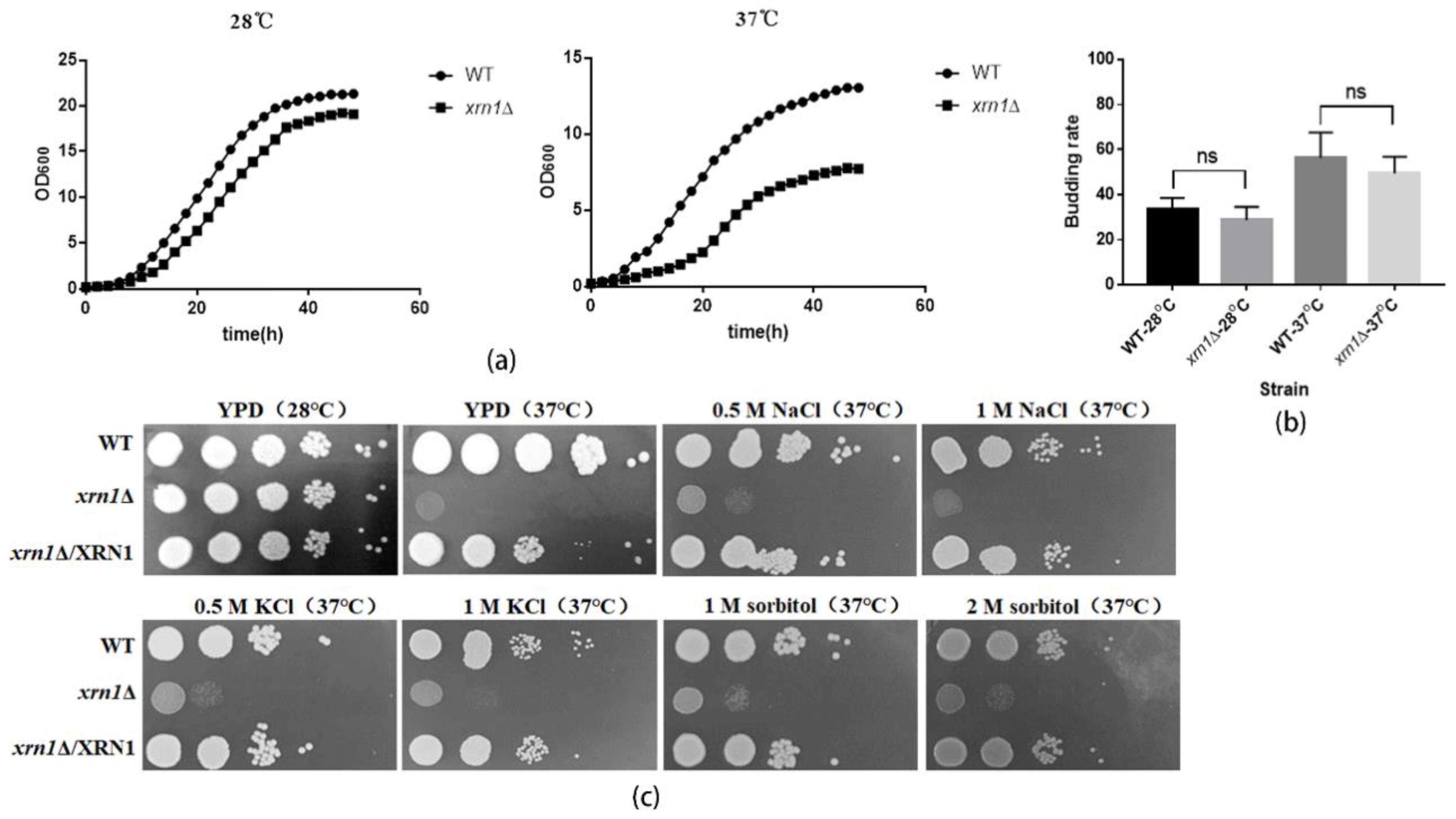
2.4. Determining the Cell Size, Growth Curves and Budding Rate
2.5. Determining the Sensitivity of xrn1∆ Strain to Stress Conditions
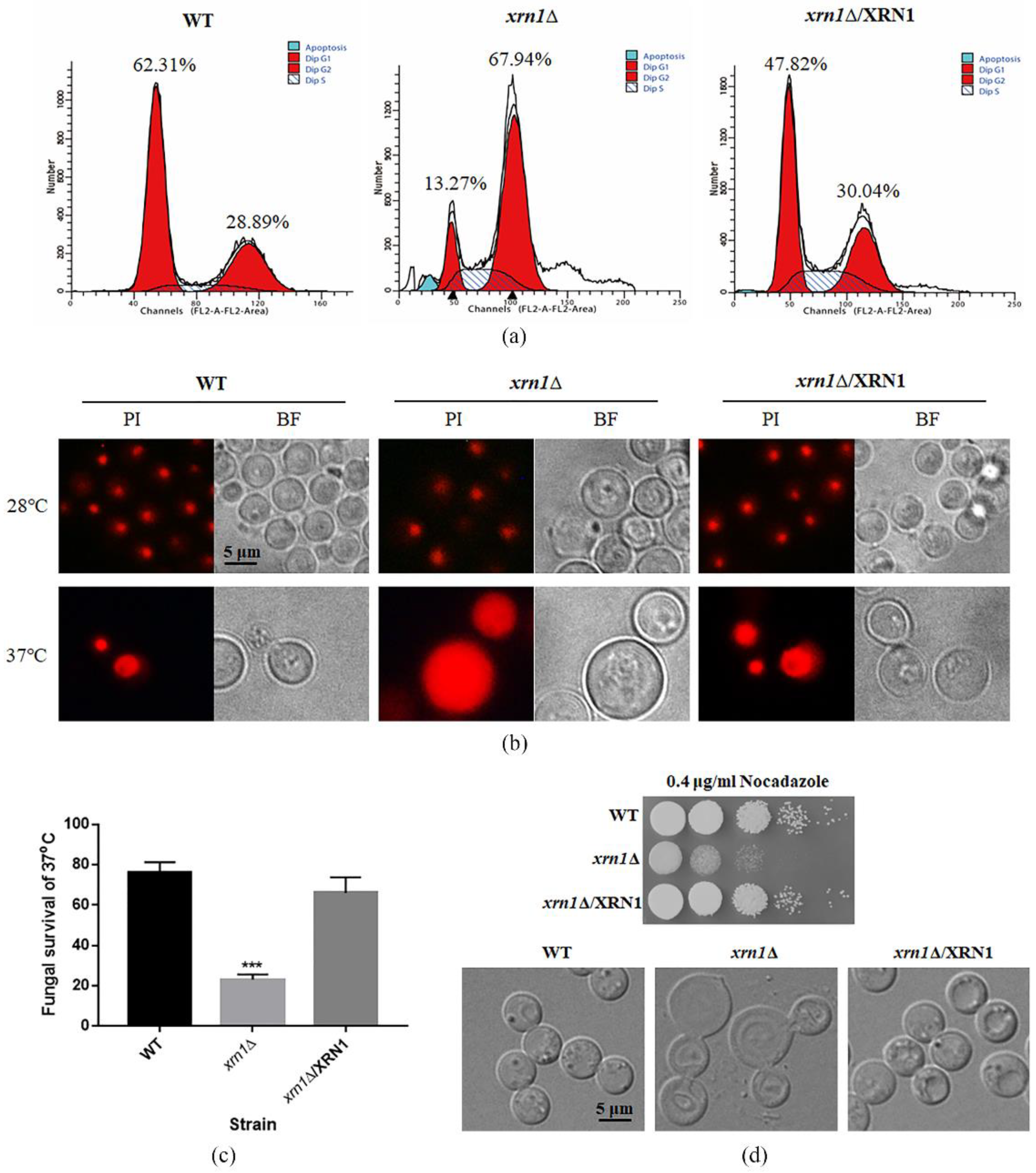
2.6. Observation of Nucleus and Flow Cytometry
2.7. RNA Preparation and Quantitative Real-Time PCR
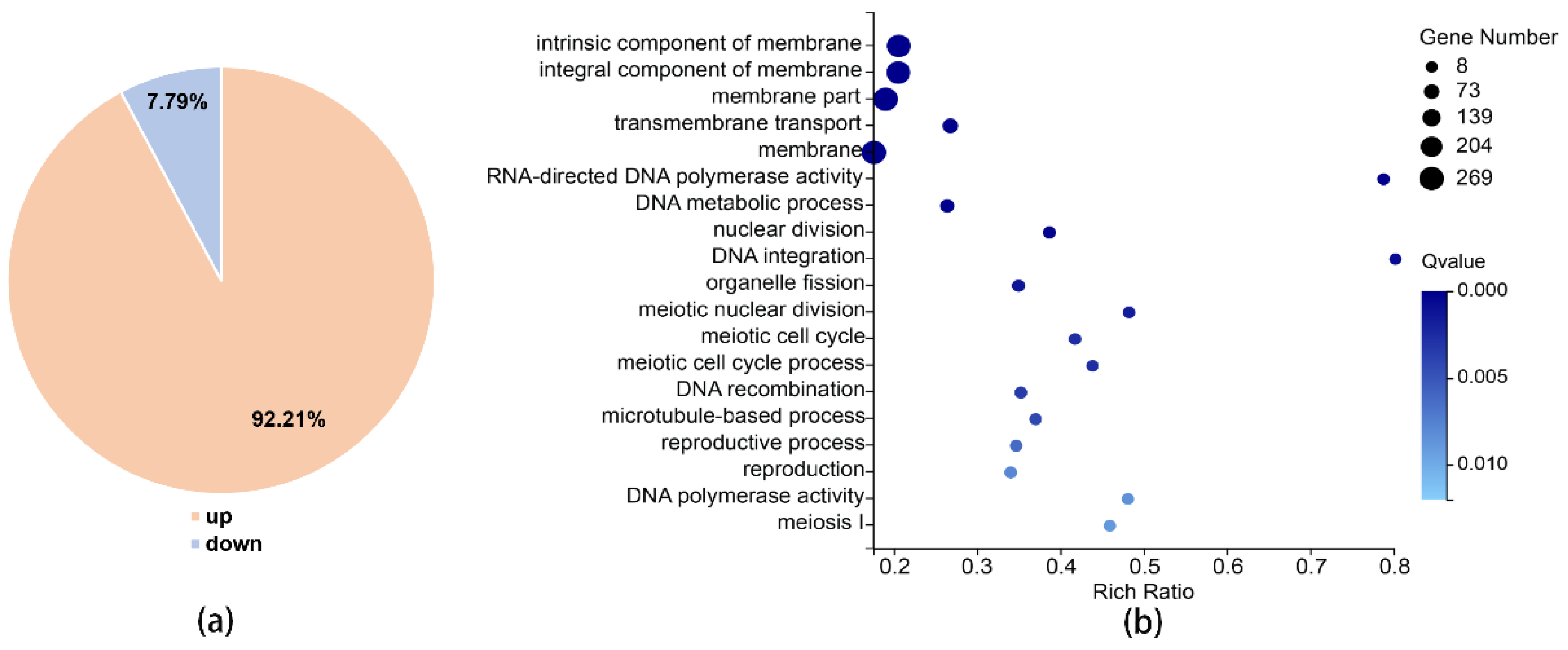
2.8. RNA-Sequencing Analyses
3. Results
3.1. Characterization of XRN1 Gene in C. Neoformans
3.2. XRN1 Deletion Increased the Cell Size
3.3. XRN1 Deletion Altered the DNA Content and Led to Hypersensitivity to Nocodazole
3.4. Multiple Functions of XRN1 in C. Neoformans
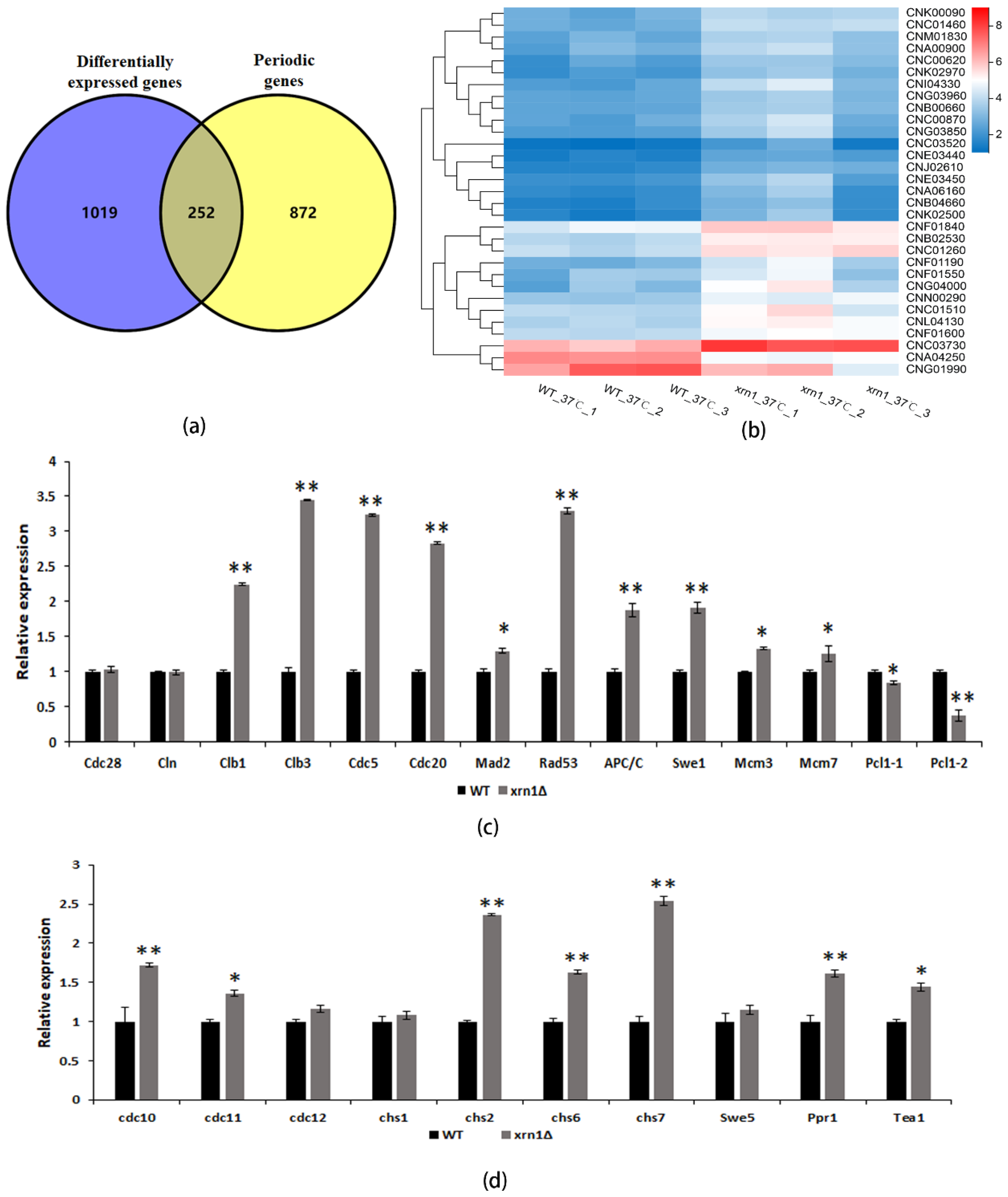
3.5. XRN1 Deletion Up-Regulated the Expression of Periodic Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Neurohr, G.E.; Terry, R.L.; Lengefeld, J.; Bonney, M.; Brittingham, G.P.; Moretto, F.; Miettinen, T.P.; Vaites, L.P.; Soares, L.M.; Paulo, J.A.; et al. Excessive cell growth causes cytoplasm dilution and contributes to senescence. Cell 2019, 176, 1083–1097.e18. [Google Scholar] [CrossRef] [PubMed]
- Aldea, M.; Jenkins, K.; Csikasz-Nagy, A. Growth rate as a direct regulator of the start network to set cell size. Front. Cell Dev. Biol. 2017, 5, 57. [Google Scholar] [CrossRef] [PubMed]
- Goehring, N.V.; Hyman, A.A. Organelle growth vontrol through limiting pools of cytoplasmic components. Curr. Biol. 2012, 22, R330–R339. [Google Scholar] [CrossRef] [PubMed]
- Talia, S.D.; Skotheim, J.M.; Bean, J.M.; Siggia, E.D.; Cross, F.R. The effects of molecular noise and size control on variability in the budding yeast cell cycle. Nature 2007, 448, 947–951. [Google Scholar] [CrossRef]
- Doncic, A.; Falleurfettig, M.; Skotheim, J.M. Distinct interactions select and maintain a specific cell fate. Mol. Cell 2011, 43, 515–527. [Google Scholar] [CrossRef]
- Meyer, S.; Temme, C.; Wahle, E. Messenger RNA turnover in eukaryotes: Pathways and enzymes. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 197–216. [Google Scholar] [CrossRef]
- Nagarajan, V.K.; Jones, C.I.; Newbury, S.F.; Green, P.J. XRN 5′→3′ exoribonucleases: Structure, mechanisms and functions. Biochim. Biophys. Acta 2013, 1829, 590–603. [Google Scholar] [CrossRef]
- Jones, C.I.; Zabolotskaya, M.V.; Newbury, S.F. The 5′→3′ exoribonuclease XRN1/Pacman and its functions in cellular processes and development. Wiley Interdiscip. Rev. Rna 2012, 3, 455–468. [Google Scholar] [CrossRef]
- Braun, J.E.; Truffault, V.; Boland, A.; Huntzinger, E.; Chang, C.-T.; Haas, G.; Weichenrieder, O.; Coles, M.; Izaurralde, E. A direct interaction between DCP1 and XRN1 couples mRNA decapping to 5′ exonucleolytic degradation. Nat. Struct. Mol. Biol. 2012, 19, 1324–1331. [Google Scholar] [CrossRef]
- Larimer, F.W.; Hsu, C.L.; Maupin, M.K.; Stevens, A. Characterization of the XRN1 gene encoding a 5′→3′ exoribonuclease: Sequence data and analysis of disparate protein and mRNA levels of gene-disrupted yeast cells. Gene 1992, 120, 51–57. [Google Scholar] [CrossRef]
- Interthal, H.; Bellocq, C.; Bähler, J.; Bashkirov, V.I.; Edelstein, S.; Heyer, W.D. A role of Sep1 (= Kem1, Xrn1) as a microtubule-associated protein in Saccharomyces cerevisiae. EMBO J. 1995, 14, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Szankasi, P.; Smith, G.R. Requirement of S. pombe exonuclease II, a homologue of S. cerevisiae Sep1, for normal mitotic growth and viability. Curr. Genet. 1996, 30, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Heyer, W.D.; Johnson, A.W.; Reinhart, U.; Kolodner, R.D. Regulation and intracellular localization of Saccharomyces cerevisiae strand exchange protein 1 (Sep1/Xrn1/Kem1), a multifunctional exonuclease. Mol. Cell. Biol. 1995, 15, 2728–2736. [Google Scholar] [CrossRef] [PubMed]
- Tishkoff, D.X.; Johnson, A.W.; Kolodner, R.D. Molecular and genetic analysis of the gene encoding the Saccharomyces cerevisiae strand exchange protein Sep1. Mol. Cell. Biol. 1991, 11, 2593–2608. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, J.; Ljungdahl, P.O.; Fink, G.R. Kem mutations affect nuclear fusion in Saccharomyces cerevisiae. Genetics 1990, 126, 799–812. [Google Scholar]
- Kim, J.; Kim, J. KEM1 is involved in filamentous growth of Saccharomyces cerevisiae. Fems Microbiol. Lett. 2002, 216, 33–38. [Google Scholar] [CrossRef][Green Version]
- An, H.S.; Lee, K.H.; Kim, J. Identification of an exoribonuclease homolog, CaKEM1/CaXRN1, in Candida albicans and its characterization in filamentous growth. Fems Microbiol. Lett. 2004, 235, 297–303. [Google Scholar] [CrossRef]
- Park, B.J.; Wannemuehler, K.A.; Marston, B.J.; Govender, N.; Pappas, P.G.; Chiller, T.M. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS. Aids 2009, 23, 525–530. [Google Scholar] [CrossRef]
- Zhu, X.; Williamson, P.R. Role of laccase in the biology and virulence of Cryptococcus neoformans. Fems Yeast Res. 2004, 5, 1–10. [Google Scholar] [CrossRef]
- Zaragoza, O.; Rodrigues, M.L.; Jesus, M.D.; Frases, S.; Dadachova, E.; Casadevall, A. The capsule of the fungal pathogen Cryptococcus neoformans. Adv. Appl. Microbiol. 2009, 68, 133–216. [Google Scholar]
- Vecchiarelli, A.; Monari, C. Capsular material of Cryptococcus neoformans: Virulence and much more. Mycopathologia 2012, 173, 375–386. [Google Scholar] [CrossRef]
- Wollschlaeger, C.; Trevijano-Contador, N.; Wang, X.; Legrand, M.; Zaragoza, O.; Heitman, J.; Janbon, G. Distinct and redundant roles of exonucleases in Cryptococcus neoformans: Implications for virulence and mating. Fungal Genet. Biol. 2014, 73, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wei, D.; Zhu, X.; Pan, J.; Zhang, P.; Huo, L. A ‘suicide’ CRISPR-Cas9 system to promote gene deletion and restoration by electroporation in Cryptococcus neoformans. Sci. Rep. 2016, 6, 31145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, D.; Zhao, X.; Wei, D.; Wang, Y.; Zhu, X. Effects of CTR4 deletion on virulence and stress response in Cryptococcus neoformans. Antonie Van Leeuwenhoek 2016, 109, 1081–1090. [Google Scholar] [CrossRef]
- Zhao, X.; Feng, W.; Zhu, X.; Li, C.; Ma, X.; Li, X.; Zhu, X.; Wei, D. Conserved autophagy pathway contributes to stress tolerance and virulence and differentially controls autophagic flux upon nutrient starvation in Cryptococcus neoformans. Front. Microbiol. 2019, 10, 2690. [Google Scholar] [CrossRef] [PubMed]
- Demidenko, Z.N.; Blagosklonny, M.V. Growth stimulation leads to cellular senescence when the cell cycle is blocked. Cell Cycle 2008, 7, 3355–3361. [Google Scholar] [CrossRef] [PubMed]
- Mollinedo, F.; Gajate, C. Microtubules, microtubule-interfering agents and apoptosis. Apoptosis 2003, 8, 413–450. [Google Scholar] [CrossRef]
- Kelliher, C.M.; Leman, A.R.; Sierra, C.S.; Haase, S.B.; Geraldine, B. Investigating conservation of the cell-cycle-regulated transcriptional program in the fungal pathogen, Cryptococcus neoformans. PLoS Genet. 2016, 12, e1006453. [Google Scholar] [CrossRef]
- Tonini, T.; Hillson, C.; Claudio, P.P. Cell separation and the maintenance of cell integrity during cytokinesis in yeast: The assembly of a septum. Yeast 2010, 27, 521–530. [Google Scholar]
- Shah, P.; Wolf, K.; Lammerding, J. Bursting the bubble – nuclear envelope rupture as a path to genomic instability? Trends Cell Biol. 2017, 27, 546–555. [Google Scholar] [CrossRef]
- Haimovich, G.; Medina, D.A.; Causse, S.Z.; Garber, M.; Millán-Zambrano, G.; Barkai, O.; Chávez, S.; Pérez-Ortín, J.E.; Darzacq, X.; Choder, M. Gene expression is circular: Factors for mRNA degradation also foster mRNA synthesis. Cell 2013, 153, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Goebels, C.; Thonn, A.; Gonzalez-Hilarion, S.; Rolland, O.; Moyrand, F.; Beilharz, T.H.; Janbon, G.; Madhani, H.D. Introns regulate gene expression in Cryptococcus neoformans in a Pab2p dependent pathway. PLoS Genet. 2013, 9, e1003686. [Google Scholar] [CrossRef] [PubMed]
- Kozubowski, L.; Lee, S.C.; Heitman, J. Signalling pathways in the pathogenesis of Cryptococcus. Cell Microbiol. 2009, 11, 370–380. [Google Scholar] [CrossRef]
- Milewski, S.; Gabriel, I.; Olchowy, J. Enzymes of UDP-GlcNAc biosynthesis in yeast. Yeast 2006, 23, 1–14. [Google Scholar] [CrossRef]
- Fernandes, J.D.; Martho, K.; Tofik, V.; Vallim, M.A.; Pascon, R.C. The role of amino acid permeases and tryptophan biosynthesis in Cryptococcus neoformans survival. PLoS ONE 2015, 10, e0132369. [Google Scholar] [CrossRef] [PubMed]
- Choera, T.; Zelante, T.; Romani, L.; Keller, N.P. A multifaceted role of tryptophan metabolism and indoleamine 2,3-dioxygenase activity in Aspergillus fumigatus-host interactions. Front. Immunol. 2017, 8, 1996. [Google Scholar] [CrossRef] [PubMed]
- Brace, J.L.; Doerfler, M.D.; Weiss, E.L. A cell separation checkpoint that enforces the proper order of late cytokinetic events. J. Cell Biol. 2019, 218, 150–170. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Kanki, T. Mechanisms and physiological roles of mitophagy in yeast. Mol. Cells 2018, 41, 35–44. [Google Scholar]
- Baudrimont, A.; Voegeli, S.; Viloria, E.C.; Stritt, F.; Lenon, M.; Wada, T.; Jaquet, V.; Becskei, A. Multiplexed gene control reveals rapid mRNA turnover. Sci. Adv. 2017, 3, e1700006. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway ID 1 | Pathway Name | Level | Q Value |
|---|---|---|---|
| ko04113 | Meiosis-yeast | Cell growth and death | 0.000301 |
| ko04011 | MAPK signaling pathway-yeast | Signal transduction | 0.000342 |
| ko00380 | Tryptophan metabolism | Amino acid metabolism | 0.003806 |
| ko00520 | Amino sugar and nucleotide sugar metabolism | Carbohydrate metabolism | 0.006373 |
| ko04111 | Cell cycle-yeast | Cell growth and death | 0.020641 |
| ko00360 | Phenylalanine metabolism | Amino acid metabolism | 0.042172 |
| ko04392 | Hippo signaling pathway-multiple species | Signal transduction | 0.042172 |
| ko04139 | Mitophagy-yeast | Transport and catabolism | 0.045003 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Li, X.; Zhang, P.; Li, C.; Feng, W.; Zhu, X.; Wei, D. Effects of 5′-3′ Exonuclease Xrn1 on Cell Size, Proliferation and Division, and mRNA Levels of Periodic Genes in Cryptococcus neoformans. Genes 2020, 11, 430. https://doi.org/10.3390/genes11040430
Zhao X, Li X, Zhang P, Li C, Feng W, Zhu X, Wei D. Effects of 5′-3′ Exonuclease Xrn1 on Cell Size, Proliferation and Division, and mRNA Levels of Periodic Genes in Cryptococcus neoformans. Genes. 2020; 11(4):430. https://doi.org/10.3390/genes11040430
Chicago/Turabian StyleZhao, Xueru, Xin Li, Ping Zhang, Chenxi Li, Weijia Feng, Xudong Zhu, and Dongsheng Wei. 2020. "Effects of 5′-3′ Exonuclease Xrn1 on Cell Size, Proliferation and Division, and mRNA Levels of Periodic Genes in Cryptococcus neoformans" Genes 11, no. 4: 430. https://doi.org/10.3390/genes11040430
APA StyleZhao, X., Li, X., Zhang, P., Li, C., Feng, W., Zhu, X., & Wei, D. (2020). Effects of 5′-3′ Exonuclease Xrn1 on Cell Size, Proliferation and Division, and mRNA Levels of Periodic Genes in Cryptococcus neoformans. Genes, 11(4), 430. https://doi.org/10.3390/genes11040430