Gene Expression Profiling of Corpus luteum Reveals Important Insights about Early Pregnancy in Domestic Sheep

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Library Preparation and Sequencing
2.3. Data Preprocessing and Mapping
2.4. Differential Gene Expression Analysis
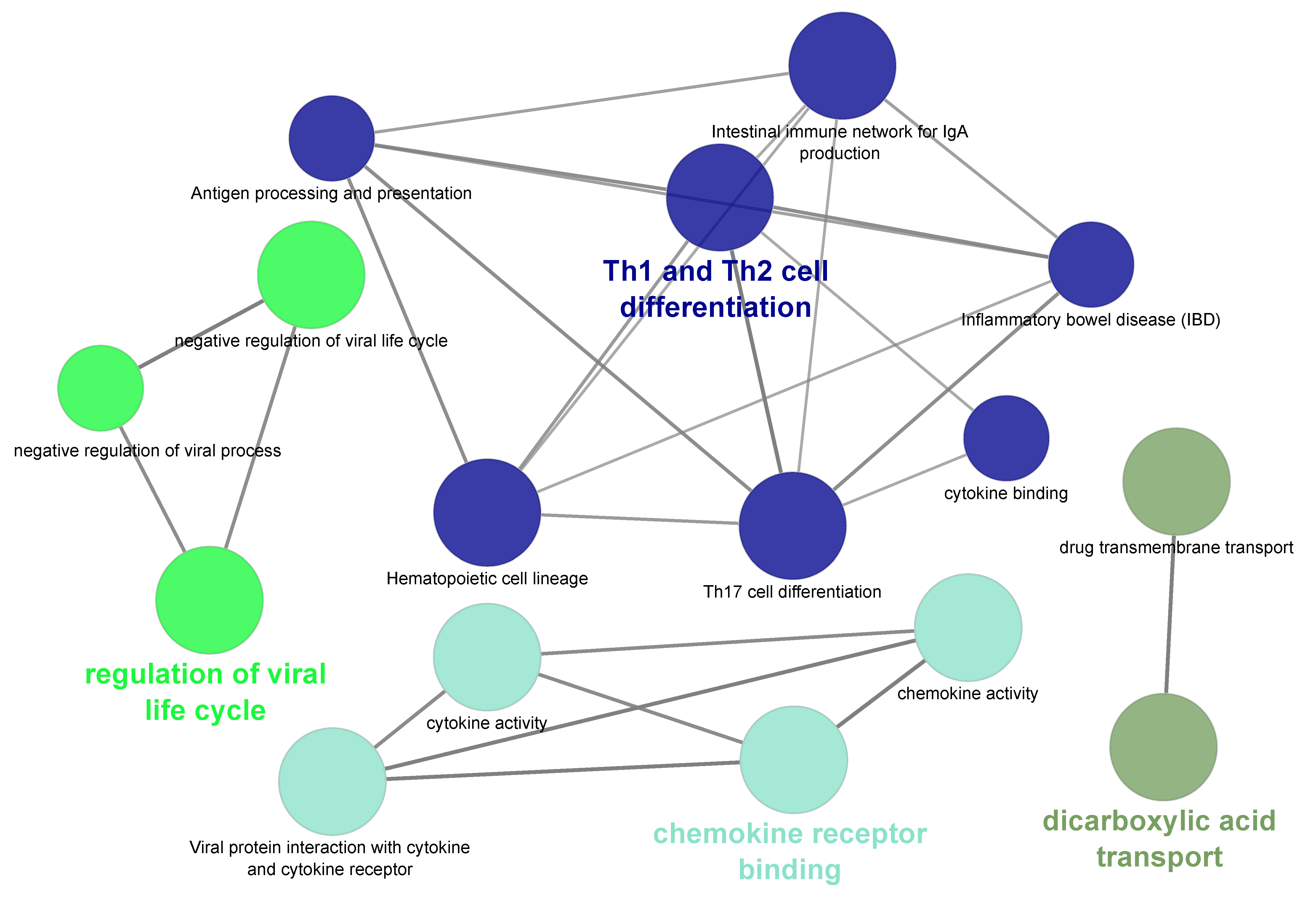
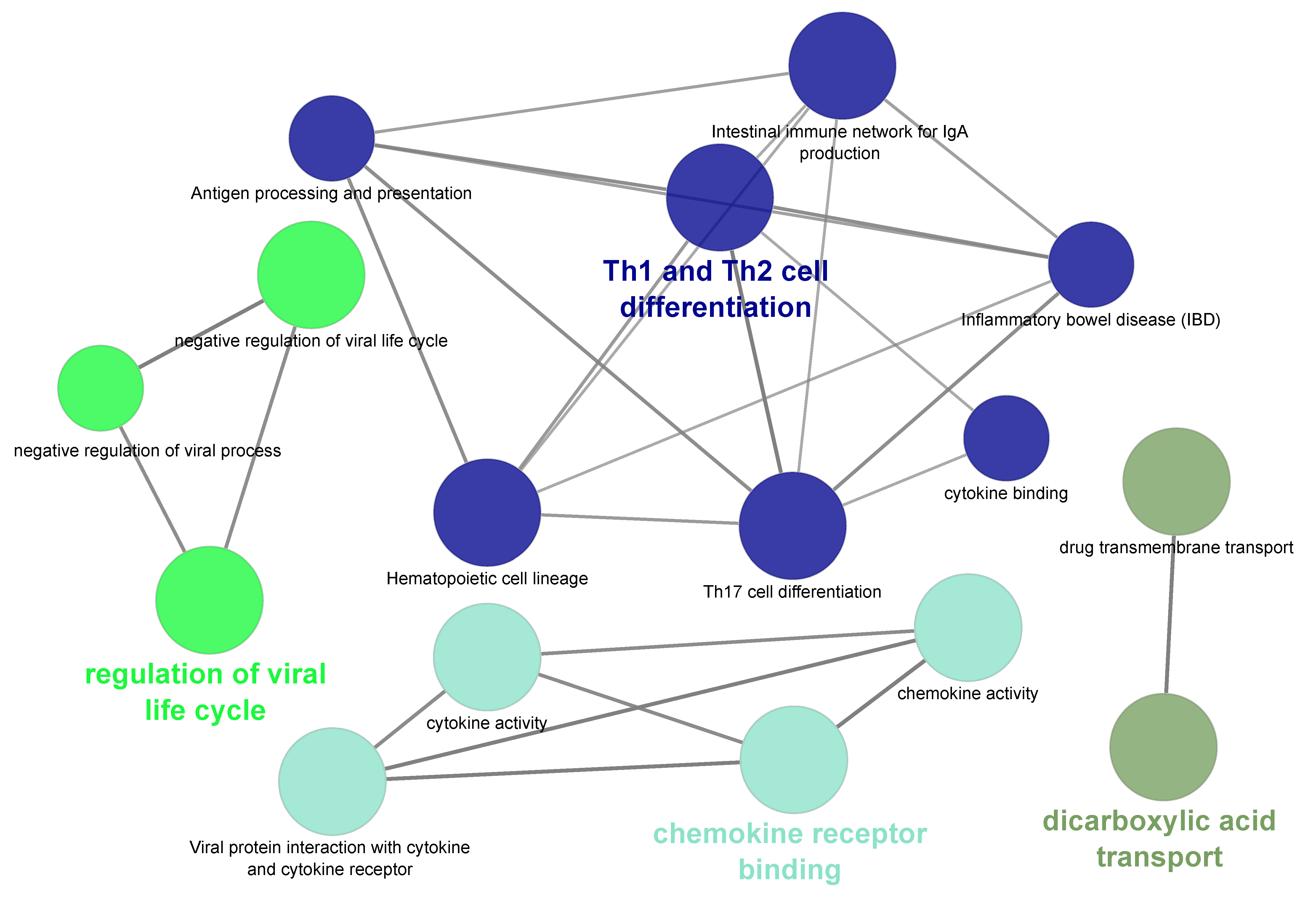
2.5. Functional Analysis of Differentially Expressed Genes
3. Results and Discussion
3.1. Phenotypic Observations
3.2. RNA-Seq Data
3.3. Gene Expression in the CL
3.4. Highly Expressed Genes
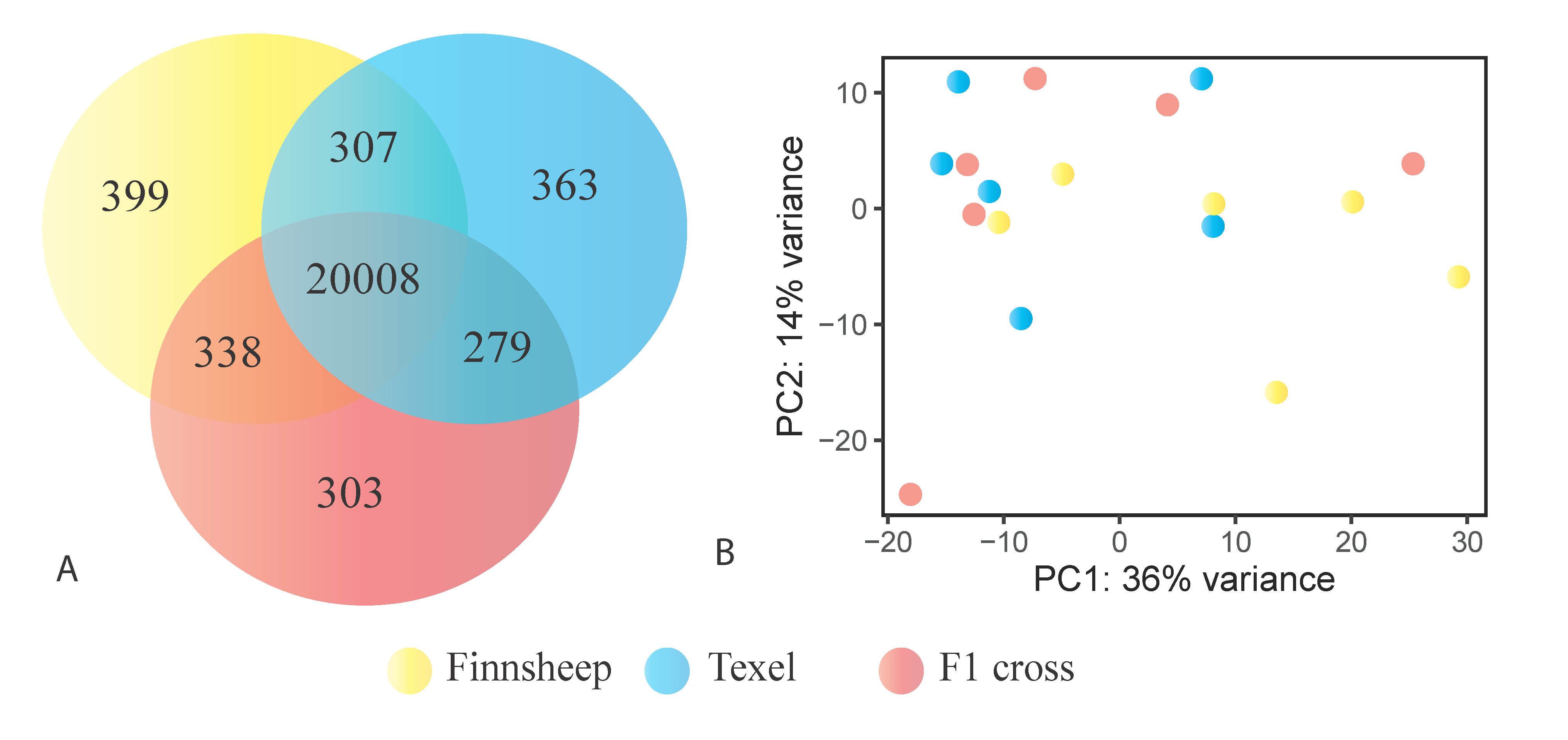
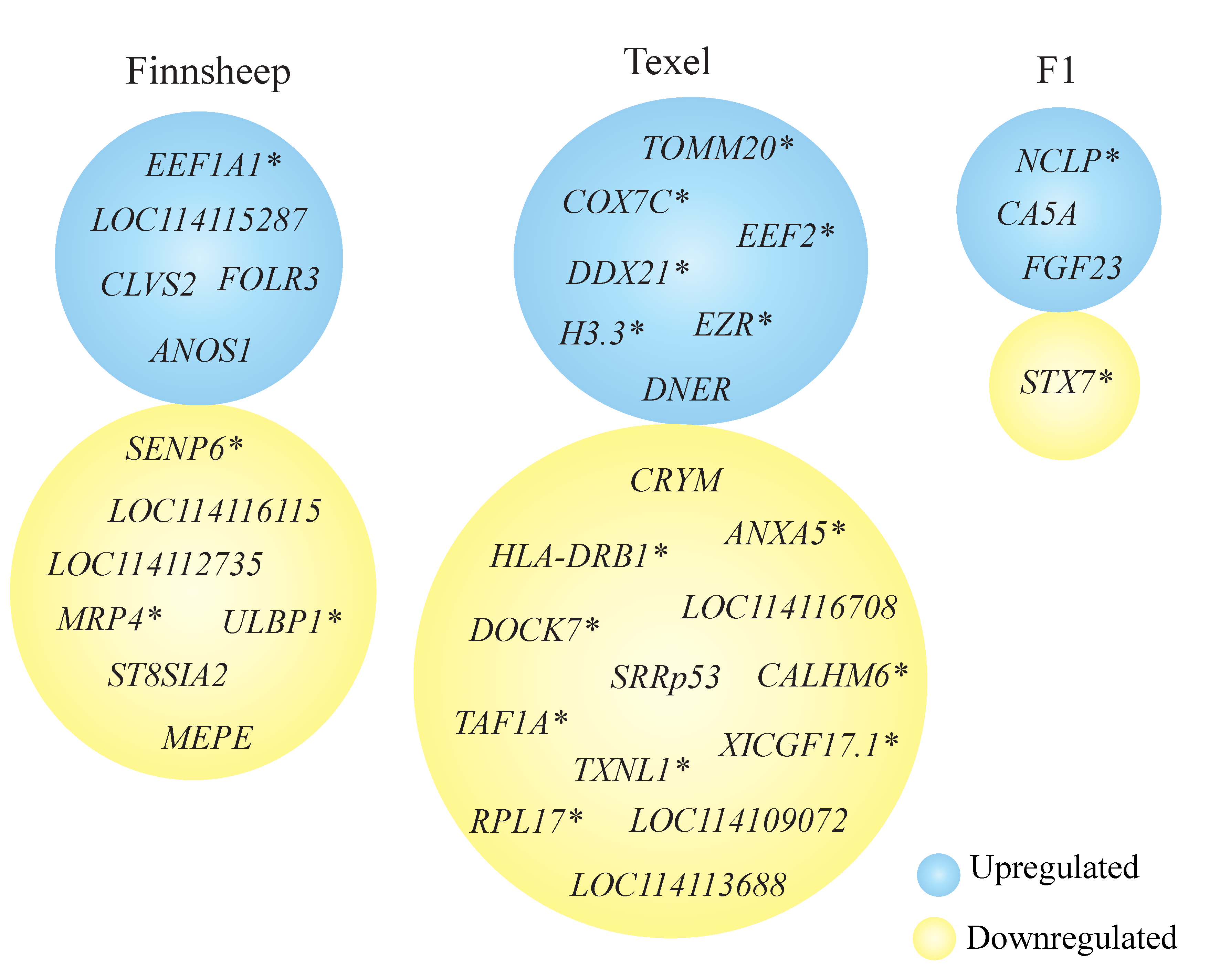
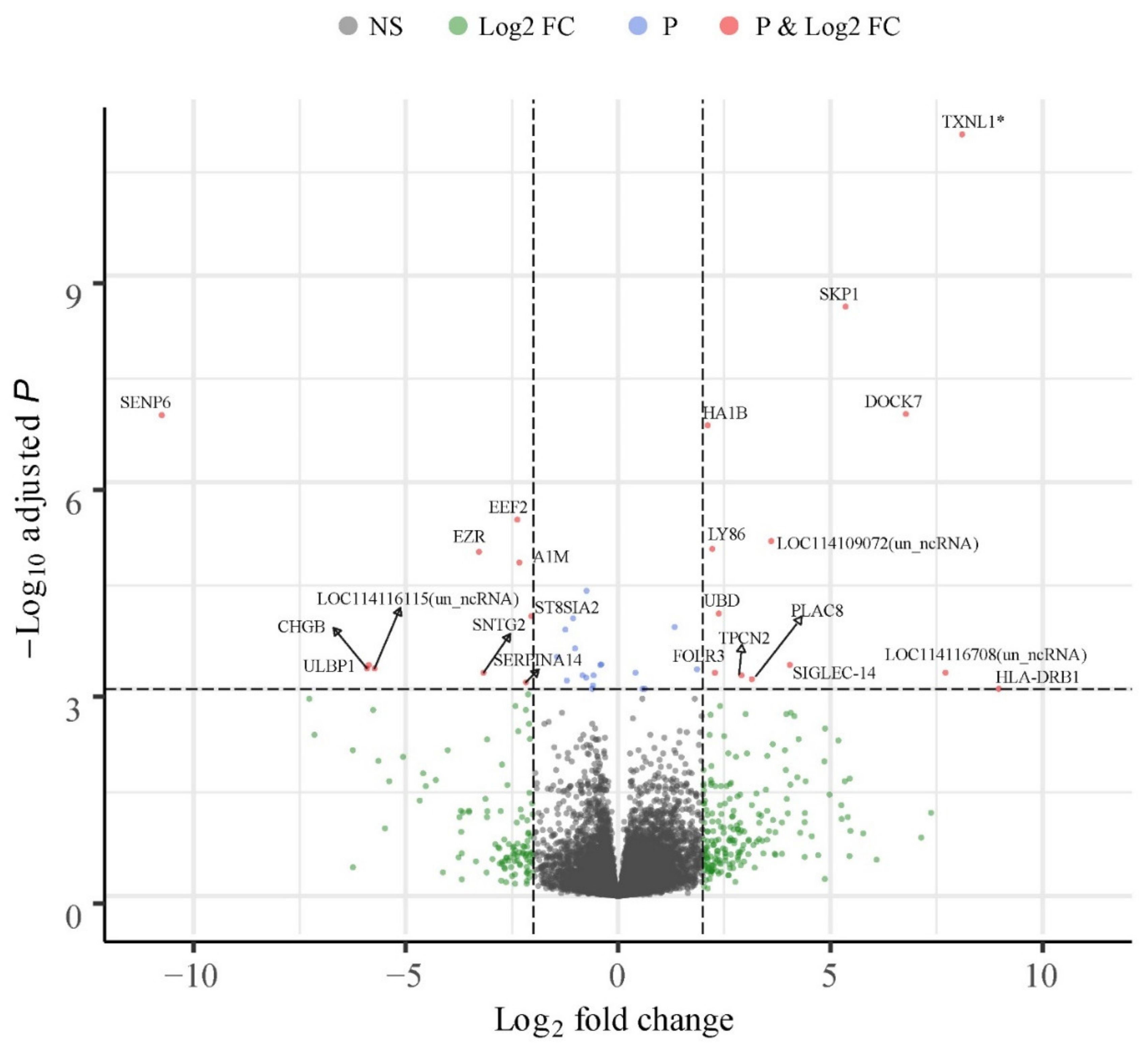
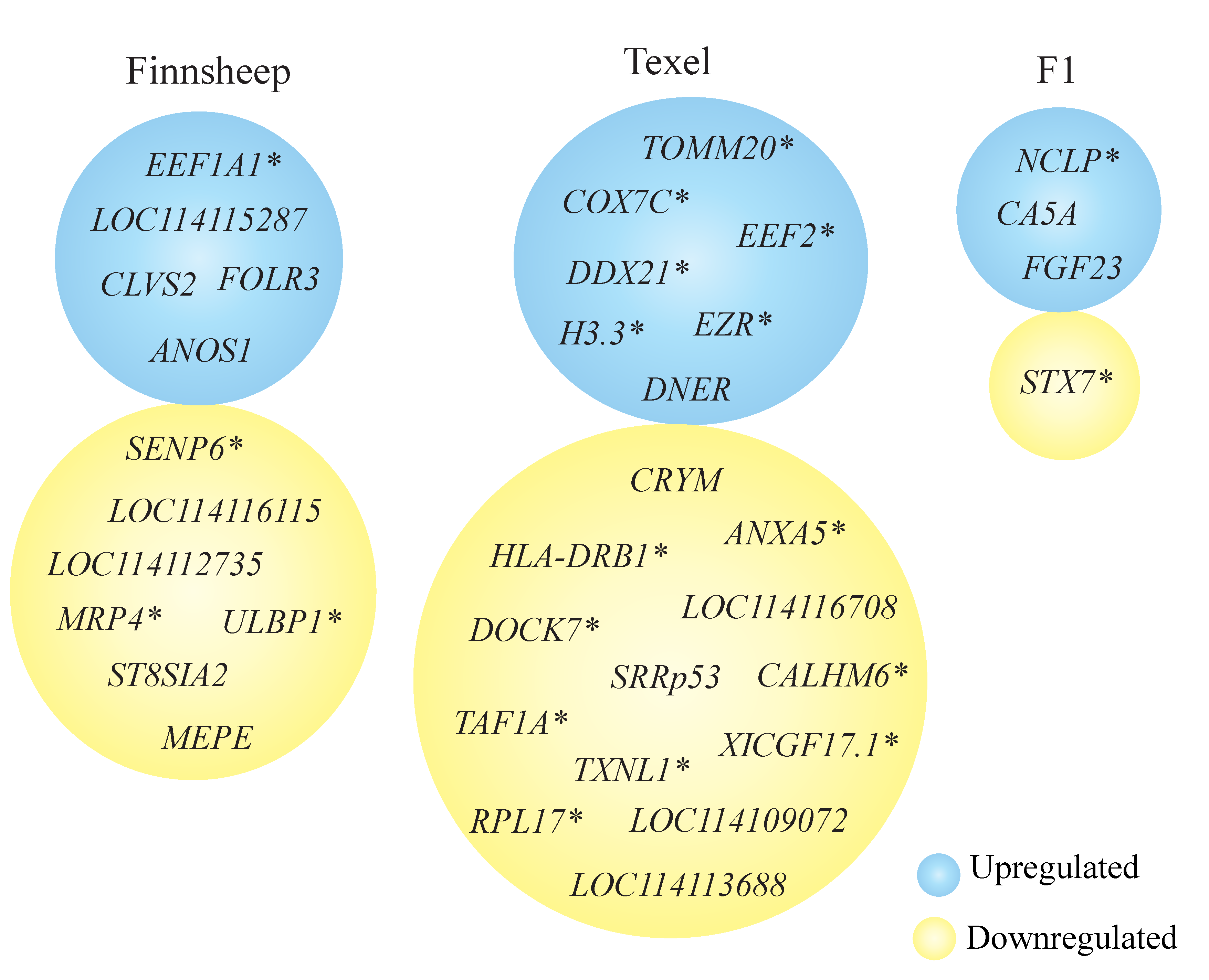
3.5. Breed Wise Gene Expression Differences in the CL
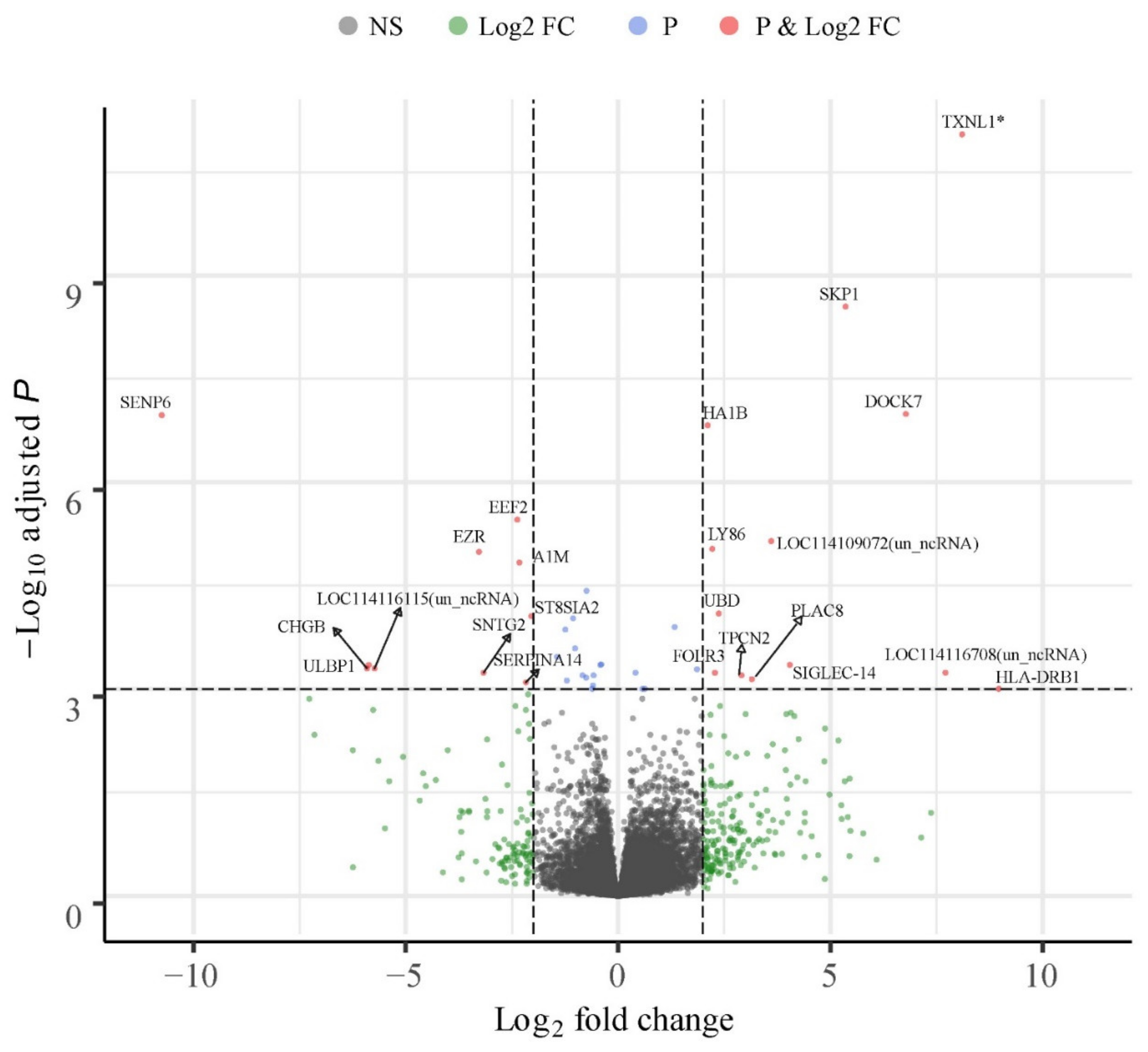
3.6. Uniquely Differentially Expressed Genes
3.7. Influence of Flushing in Gene Expression
3.8. Limitations and Thoughts for Future Studies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davis, G.H.; Farquhar, P.A.; O’Connell, A.R.; Everett-Hincks, J.M.; Wishart, P.J.; Galloway, S.M.; Dodds, K.G. A putative autosomal gene increasing ovulation rate in Romney sheep. Anim. Reprod. Sci. 2006, 92, 65–73. [Google Scholar] [CrossRef]
- Davis, G.H.; Galloway, S.M.; Ross, I.K.; Gregan, S.M.; Ward, J.; Nimbkar, B.V.; Ghalsasi, P.M.; Nimbkar, C.; Gray, G.D.; Inounu, I.; et al. DNA tests in prolific sheep from eight countries provide new evidence on origin of the Booroola (FecB) mutation. Biol. Reprod. 2002, 66, 1869–1874. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.X.; Mu, Y.L.; Fang, L.; Ye, S.C.; Sun, S.H. Prolactin receptor as a candidate gene for prolificacy of small tail Han sheep. Anim. Biotechnol. 2007, 18, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Drouilhet, L.; Mansanet, C.; Sarry, J.; Tabet, K.; Bardou, P.; Woloszyn, F.; Lluch, J.; Harichaux, G.; Viguié, C.; Monniaux, D.; et al. The highly prolific phenotype of Lacaune sheep is associated with an ectopic expression of the B4GALNT2 gene within the ovary. PLoS Genet. 2013, 9, e1003809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulsant, P.; Lecerf, F.; Fabre, S.; Schibler, L.; Monget, P.; Lanneluc, I.; Pisselet, C.; Riquet, J.; Monniaux, D.; Callebaut, I.; et al. Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Mérino ewes. Proc. Natl. Acad. Sci. USA 2001, 98, 5104–5109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, C.J.; MacDougall, C.; MacDougall, C.; Campbell, B.K.; McNeilly, A.S.; Baird, D.T. The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1 B (BMPR1B) gene. J. Endocrinol. 2001, 169, R1–R6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricordeau, G.; Thimonier, J.; Poivey, J.P.; Driancourt, M.A.; Hochereau-De-Reviers, M.T.; Tchamitchian, L. I.N.R.A. research on the Romanov sheep breed in France: A review. Livest. Prod. Sci. 1990, 24, 305–332. [Google Scholar] [CrossRef]
- Xu, S.-S.; Gao, L.; Xie, X.-L.; Ren, Y.-L.; Shen, Z.-Q.; Wang, F.; Shen, M.; Eyϸórsdóttir, E.; Hallsson, J.H.; Kiseleva, T.; et al. Genome-wide association analyses highlight the potential for different genetic mechanisms for litter size among sheep breeds. Front. Genet. 2018, 9, 118. [Google Scholar] [CrossRef]
- Hanrahan, J.P.; Quirke, J.F. Contribution of variation in ovulation rate and embryo survival to within breed variation in litter size. In Genetics of Reproduction in Sheep; Land, R.B., Robinson, D.W., Eds.; Butterworth-Heinemann: Oxford, UK, 1984. [Google Scholar]
- Hanrahan, J.P.; Gregan, S.M.; Mulsant, P.; Mullen, M.; Davis, G.H.; Powell, R.; Galloway, S.M. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol. Reprod. 2004, 70, 900–909. [Google Scholar] [CrossRef]
- Mullen, M.P.; Hanrahan, J.P. Direct evidence on the contribution of a missense mutation in GDF9 to variation in ovulation rate of Finnsheep. PLoS ONE 2014, 9, e95251. [Google Scholar] [CrossRef] [Green Version]
- Våge, D.I.; Husdal, M.; Kent, M.P.; Klemetsdal, G.; Boman, I.A. A missense mutation in growth differentiation factor 9 (GDF9) is strongly associated with litter size in sheep. BMC Genet. 2013, 14, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokharel, K.; Peippo, J.; Honkatukia, M.; Seppälä, A.; Rautiainen, J.; Ghanem, N.; Hamama, T.-M.; Crowe, M.A.; Andersson, M.; Li, M.-H.; et al. Integrated ovarian mRNA and miRNA transcriptome profiling characterizes the genetic basis of prolificacy traits in sheep (Ovis aries). BMC Genomics 2018, 19, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, T.E.; Johnson, G.A.; Bazer, F.W.; Burghardt, R.C.; Palmarini, M. Pregnancy recognition and conceptus implantation in domestic ruminants: Roles of progesterone, interferons and endogenous retroviruses. Reprod. Fertil. Dev. 2007, 19, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Johnson, G.A.; Bazer, F.W.; Burghardt, R.C. Implantation mechanisms: Insights from the sheep. Reprod. Camb. Engl. 2004, 128, 657–668. [Google Scholar] [CrossRef]
- Geisert, R.D.; Morgan, G.L.; Short, E.C.; Zavy, M.T. Endocrine events associated with endometrial function and conceptus development in cattle. Reprod. Fertil. Dev. 1992, 4, 301–305. [Google Scholar] [CrossRef]
- Quinlivan, T.D.; Martin, C.A.; Taylor, W.B.; Cairney, I.M. Estimates of pre- and perinatal mortality in the New Zealand Romney Marsh ewe. I. Pre- and perinatal mortality in those ewes that conceived to one service. J. Reprod. Fertil. 1966, 11, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Bolet, G. Timing and extent of embryonic mortality in pigs sheep and goats: Genetic variability. In Embryonic Mortality in Farm Animals; Springer: Dordrecht, The Netherlands, 1986; pp. 12–43. [Google Scholar]
- Rickard, J.P.; Ryan, G.; Hall, E.; de Graaf, S.P.; Hermes, R. Using transrectal ultrasound to examine the effect of exogenous progesterone on early embryonic loss in sheep. PLoS ONE 2017, 12, e0183659. [Google Scholar] [CrossRef] [Green Version]
- Ashworth, C.J.; Toma, L.M.; Hunter, M.G. Nutritional effects on oocyte and embryo development in mammals: Implications for reproductive efficiency and environmental sustainability. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2009, 364, 3351–3361. [Google Scholar] [CrossRef]
- Jindal, R.; Cosgrove, J.R.; Foxcroft, G.R. Progesterone mediates nutritionally induced effects on embryonic survival in gilts. J. Anim. Sci. 1997, 75, 1063–1070. [Google Scholar] [CrossRef] [Green Version]
- Mamo, S.; Mehta, J.P.; Forde, N.; McGettigan, P.; Lonergan, P. Conceptus-endometrium crosstalk during maternal recognition of pregnancy in cattle. Biol. Reprod. 2012, 87, 1–9. [Google Scholar] [CrossRef]
- Bazer, F.W. Pregnancy recognition signaling mechanisms in ruminants and pigs. J. Anim. Sci. Biotechnol. 2013, 4, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raheem, K.A. An insight into maternal recognition of pregnancy in mammalian species. J. Saudi Soc. Agric. Sci. 2017, 16, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Gray, C.A.; Abbey, C.A.; Beremand, P.D.; Choi, Y.; Farmer, J.L.; Adelson, D.L.; Thomas, T.L.; Bazer, F.W.; Spencer, T.E. Identification of endometrial genes regulated by early pregnancy, progesterone, and interferon tau in the ovine uterus. Biol. Reprod. 2006, 74, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Brooks, K.; Burns, G.W.; Moraes, J.G.N.; Spencer, T.E. Analysis of the uterine epithelial and conceptus transcriptome and luminal fluid proteome during the peri-implantation period of pregnancy in sheep. Biol. Reprod. 2016, 95, 88. [Google Scholar] [CrossRef]
- Moore, S.G.; Pryce, J.E.; Hayes, B.J.; Chamberlain, A.J.; Kemper, K.E.; Berry, D.P.; McCabe, M.; Cormican, P.; Lonergan, P.; Fair, T.; et al. Differentially expressed genes in endometrium and corpus luteum of holstein cows selected for high and low fertility are enriched for sequence variants associated with fertility. Biol. Reprod. 2016, 94, 19. [Google Scholar] [CrossRef] [Green Version]
- Kfir, S.; Basavaraja, R.; Wigoda, N.; Ben-Dor, S.; Orr, I.; Meidan, R. Genomic profiling of bovine corpus luteum maturation. PLoS ONE 2018, 13, e0194456. [Google Scholar] [CrossRef]
- Andrews, S. FastQC 0.11.6; A Quality Control Tool for High Throughput Sequence Data. Babraham Bioinformatics: Cambridge, UK. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 18 January 2013).
- Krueger, F. Trim Galore; Babraham Bioinformatics: Cambridge, UK. Available online: http://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 9 January 2019).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon: Fast and bias-aware quantification of transcript expression using dual-phase inference. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential analyses for RNA-seq: Transcript-level estimates improve gene-level inferences. F1000Research 2016, 4, 1521. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team (2019). Available online: https://www.R-project.org/ (accessed on 8 April 2020).
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Rhind, S.M.; Robinson, J.J.; Fraser, C.; McHattie, I. Ovulation and embryo survival rates and plasma progesterone concentrations of prolific ewes treated with PMSG. J. Reprod. Fertil. 1980, 58, 139–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, C.L.A.D.; van den Brand, H.; Laurenssen, B.F.A.; Broekhuijse, M.L.W.J.; Knol, E.F.; Kemp, B.; Soede, N.M. Relationships between ovulation rate and embryonic and placental characteristics in multiparous sows at 35 days of pregnancy. Animal 2016, 10, 1192–1199. [Google Scholar] [CrossRef] [Green Version]
- Stocco, D.M. The role of the StAR protein in steroidogenesis: Challenges for the future. J. Endocrinol. 2000, 164, 247–253. [Google Scholar] [CrossRef]
- Christenson, L.K.; Devoto, L. Cholesterol transport and steroidogenesis by the corpus luteum. Reprod. Biol. Endocrinol. RBE 2003, 1, 90. [Google Scholar] [CrossRef] [Green Version]
- Juengel, J.L.; Meberg, B.M.; Turzillo, A.M.; Nett, T.M.; Niswender, G.D. Hormonal regulation of messenger ribonucleic acid encoding steroidogenic acute regulatory protein in ovine corpora lutea. Endocrinology 1995, 136, 5423–5429. [Google Scholar] [CrossRef]
- Devoto, L.; Kohen, P.; Gonzalez, R.R.; Castro, O.; Retamales, I.; Vega, M.; Carvallo, P.; Christenson, L.K.; Strauss, J.F. Expression of steroidogenic acute regulatory protein in the human corpus luteum throughout the luteal phase. J. Clin. Endocrinol. Metab. 2001, 86, 5633–5639. [Google Scholar] [CrossRef]
- Davis, J.S.; LaVoie, H.A. Molecular regulation of progesterone production in the corpus luteum. In The Ovary, 3rd ed.; Academic Press: Cambridge, MA, USA, 2019; ISBN 9780128132098. [Google Scholar]
- King, S.R.; LaVoie, H.A. Regulation of the early steps in gonadal steroidogenesis. In Reproductive Endocrinology; Springer: Boston, MA, USA, 2009; pp. 175–193. [Google Scholar]
- Hu, J.; Zhang, Z.; Shen, W.-J.; Azhar, S. Cellular cholesterol delivery, intracellular processing and utilization for biosynthesis of steroid hormones. Nutr. Metab. 2010, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Plant, T.M.; Zeleznik, A.J.; Auchus, R.J. Chapter 8—Human steroid biosynthesis. In Knobil and Neill’s Physiology of Reproduction; Academic Press: Cambridge, MA, USA, 2015; pp. 295–312. ISBN 9780123971753. [Google Scholar]
- Stouffer, R.L.; Hennebold, J.D. Structure, function, and regulation of the corpus luteum. In Knobil and Neill’s Physiology of Reproduction; Academic Press: Cambridge, MA, USA, 2015; pp. 1023–1076. ISBN 9780123971753. [Google Scholar]
- Spencer, T.E.; Burghardt, R.C.; Johnson, G.A.; Bazer, F.W. Conceptus signals for establishment and maintenance of pregnancy. Anim. Reprod. Sci. 2004, 82–83, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Flint, A.P.F.; Sheldrick, E.L. Ovarian oxytocin and the maternal recognition of pregnancy. Reproduction 1986, 76, 831–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendrick, K.M. Oxytocin, motherhood and bonding. Exp. Physiol. 2000, 85, 111s–124s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, C.V. Progesterone inhibition of oxytocin signaling in endometrium. Front. Neurosci. 2013, 7, 138. [Google Scholar] [CrossRef] [Green Version]
- Fleming, J.A.G.W.; Spencer, T.E.; Safe, S.H.; Bazer, F.W. Estrogen regulates transcription of the ovine oxytocin receptor gene through GC-rich SP1 promoter elements. Endocrinology 2006, 147, 899–911. [Google Scholar] [CrossRef] [Green Version]
- Gimpl, G.; Wiegand, V.; Burger, K.; Fahrenholz, F. Cholesterol and steroid hormones: Modulators of oxytocin receptor function. In Vasopressin and Oxytocin; Elsevier: Amsterdam, The Netherlands, 2002; Volume 139, pp. 43–56. [Google Scholar]
- Grazzini, E.; Guillon, G.; Mouillac, B.; Zingg, H.H. Inhibition of oxytocin receptor function by direct binding of progesterone. Nature 1998, 392, 509–512. [Google Scholar] [CrossRef]
- Nissi, R.; Talvensaari-Mattila, A.; Kotila, V.; Niinimäki, M.; Järvelä, I.; Turpeenniemi-Hujanen, T. Circulating matrix metalloproteinase MMP-9 and MMP-2/TIMP-2 complex are associated with spontaneous early pregnancy failure. Reprod. Biol. Endocrinol. RBE 2013, 11, 2. [Google Scholar] [CrossRef] [Green Version]
- Kliem, H.; Welter, H.; Kraetzl, W.D.; Steffl, M.; Meyer, H.H.D.; Schams, D.; Berisha, B. Expression and localisation of extracellular matrix degrading proteases and their inhibitors during the oestrous cycle and after induced luteolysis in the bovine corpus luteum. Reprod. Camb. Engl. 2007, 134, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Curry, T.E.; Osteen, K.G. Cyclic changes in the matrix metalloproteinase system in the ovary and uterus. Biol. Reprod. 2001, 64, 1285–1296. [Google Scholar] [CrossRef] [Green Version]
- Moses, M.A.; Sudhalter, J.; Langer, R. Identification of an inhibitor of neovascularization from cartilage. Science 1990, 248, 1408–1410. [Google Scholar] [CrossRef]
- Docherty, A.J.; Lyons, A.; Smith, B.J.; Wright, E.M.; Stephens, P.E.; Harris, T.J.; Murphy, G.; Reynolds, J.J. Sequence of human tissue inhibitor of metalloproteinases and its identity to erythroid-potentiating activity. Nature 1985, 318, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, Y.; Sugiura, F.; Kishi, J.; Hayakawa, T. Collagenase inhibitor stimulates cleft formation during early morphogenesis of mouse salivary gland. Dev. Biol. 1986, 113, 201–206. [Google Scholar] [CrossRef]
- Hampton, A.L.; Butt, A.R.; Riley, S.C.; Salamonsen, L.A. Tissue inhibitors of metalloproteinases in endometrium of ovariectomized steroid-treated ewes and during the estrous cycle and early pregnancy. Biol. Reprod. 1995, 53, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Librach, C.L.; Werb, Z.; Fitzgerald, M.L.; Chiu, K.; Corwin, N.M.; Esteves, R.A.; Grobelny, D.; Galardy, R.; Damsky, C.H.; Fisher, S.J. 92-kD type IV collagenase mediates invasion of human cytotrophoblasts. J. Cell Biol. 1991, 113, 437–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, S.X.; Wang, Y.L.; Qin, L.; Xiao, Z.J.; Herva, R.; Piao, Y.S. Dynamic expression of matrix metalloproteinases (MMP-2, -9 and -14) and the tissue inhibitors of MMPs (TIMP-1, -2 and -3) at the implantation site during tubal pregnancy. Reproduction 2005, 129, 103–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabler, C.; Killian, G.J.; Einspanier, R. Differential expression of extracellular matrix components in the bovine oviduct during the oestrous cycle. Reprod. Camb. Engl. 2001, 122, 121–130. [Google Scholar] [CrossRef]
- Brioudes, F.; Thierry, A.-M.; Chambrier, P.; Mollereau, B.; Bendahmane, M. Translationally controlled tumor protein is a conserved mitotic growth integrator in animals and plants. Proc. Natl. Acad. Sci. USA 2010, 107, 16384–16389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Chen, X.; Ding, Y.; Liu, X.; Wang, Y.; He, J. Expression of translationally controlled tumor protein (TCTP) in the uterus of mice of early pregnancy and its possible significance during embryo implantation. Hum. Reprod. 2011, 26, 2972–2980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuynder, M.; Susini, L.; Prieur, S.; Besse, S.; Fiucci, G.; Amson, R.; Telerman, A. Biological models and genes of tumor reversion: Cellular reprogramming through tpt1/TCTP and SIAH-1. Proc. Natl. Acad. Sci. USA 2002, 99, 14976–14981. [Google Scholar] [CrossRef] [Green Version]
- Branco, R.; Masle, J. Systemic signalling through translationally controlled tumour protein controls lateral root formation in Arabidopsis. J. Exp. Bot. 2019, 70, 3927–3940. [Google Scholar] [CrossRef]
- Chen, S.H.; Wu, P.-S.; Chou, C.-H.; Yan, Y.-T.; Liu, H.; Weng, S.-Y.; Yang-Yen, H.-F. A knockout mouse approach reveals that TCTP functions as an essential factor for cell proliferation and survival in a tissue- or cell type-specific manner. Mol. Biol. Cell 2007, 18, 2525–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Warburton, D. Matrix Gla protein gene expression is induced by transforming growth factor-beta in embryonic lung culture. Am. J. Physiol. Lung Cell. Mol. Physiol. 1997, 273, L282–L287. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Nishimoto, S.K. Matrix Gla protein gene expression is elevated during postnatal development. Matrix Biol. J. Int. Soc. Matrix Biol. 1996, 15, 131–140. [Google Scholar] [CrossRef]
- Loeser, R.F.; Wallin, R. Cell adhesion to matrix Gla protein and its inhibition by an Arg-Gly-Asp-containing peptide. J. Biol. Chem. 1992, 267, 9459–9462. [Google Scholar]
- Forde, N.; Mehta, J.P.; McGettigan, P.A.; Mamo, S.; Bazer, F.W.; Spencer, T.E.; Lonergan, P. Alterations in expression of endometrial genes coding for proteins secreted into the uterine lumen during conceptus elongation in cattle. BMC Genomics 2013, 14, 321. [Google Scholar] [CrossRef] [Green Version]
- Spencer, T.E.; Stagg, A.G.; Joyce, M.M.; Jenster, G.; Wood, C.G.; Bazer, F.W.; Wiley, A.A.; Bartol, F.F. Discovery and characterization of endometrial epithelial messenger ribonucleic acids using the ovine uterine gland knockout model. Endocrinology 1999, 140, 4070–4080. [Google Scholar] [CrossRef]
- Casey, O.M.; Morris, D.G.; Powell, R.; Sreenan, J.M.; Fitzpatrick, R. Analysis of gene expression in non-regressed and regressed bovine corpus luteum tissue using a customized ovarian cDNA array. Theriogenology 2005, 64, 1963–1976. [Google Scholar] [CrossRef]
- Lacroix-Pépin, N.; Danyod, G.; Krishnaswamy, N.; Mondal, S.; Rong, P.-M.; Chapdelaine, P.; Fortier, M.A. The multidrug resistance-associated protein 4 (MRP4) appears as a functional carrier of prostaglandins regulated by oxytocin in the bovine endometrium. Endocrinology 2011, 152, 4993–5004. [Google Scholar] [CrossRef] [Green Version]
- Varki, A.; Angata, T. Siglecs—The major subfamily of I-type lectins. Glycobiology 2006, 16, 1R–27R. [Google Scholar] [CrossRef] [Green Version]
- Angata, T.; Margulies, E.H.; Green, E.D.; Varki, A. Large-scale sequencing of the CD33-related Siglec gene cluster in five mammalian species reveals rapid evolution by multiple mechanisms. Proc. Natl. Acad. Sci. USA 2004, 101, 13251–13256. [Google Scholar] [CrossRef] [Green Version]
- Pillai, S.; Netravali, I.A.; Cariappa, A.; Mattoo, H. Siglecs and immune regulation. Annu. Rev. Immunol. 2012, 30, 357–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornhöfft, K.F.; Goldammer, T.; Rebl, A.; Galuska, S.P. Siglecs: A journey through the evolution of sialic acid-binding immunoglobulin-type lectins. Dev. Comp. Immunol. 2018, 86, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Crocker, P.R. Evolution of CD33-related siglecs: Regulating host immune functions and escaping pathogen exploitation? Immunology 2011, 132, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Blighe, K.; Rana, S.; Lewis, M. EnhancedVolcano: Publication-Ready Volcano Plots with Enhanced Colouring and Labeling; GitHub: San Francisco, CA, USA, 2019. [Google Scholar]
- Gu, T.; Zhu, M.; Schroyen, M.; Qu, L.; Nettleton, D.; Kuhar, D.; Lunney, J.K.; Ross, J.W.; Zhao, S.; Tuggle, C.K. Endometrial gene expression profiling in pregnant Meishan and Yorkshire pigs on day 12 of gestation. BMC Genomics 2014, 15, 156. [Google Scholar] [CrossRef] [Green Version]
- Tatham, M.H.; Jaffray, E.; Vaughan, O.A.; Desterro, J.M.P.; Botting, C.H.; Naismith, J.H.; Hay, R.T. Polymeric chains of SUMO-2 and SUMO-3 are conjugated to protein substrates by SAE1/SAE2 and Ubc9. J. Biol. Chem. 2001, 276, 35368–35374. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lu, D.; Dou, H.; Liu, H.; Weaver, K.; Wang, W.; Li, J.; Yeh, E.T.H.; Williams, B.O.; Zheng, L.; et al. Desumoylase SENP6 maintains osteochondroprogenitor homeostasis by suppressing the p53 pathway. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Liu, X.; Chen, W.; Wang, Q.; Li, L.; Wang, C. Negative regulation of TLR inflammatory signaling by the SUMO-deconjugating enzyme SENP6. PLOS Pathog. 2013, 9, e1003480. [Google Scholar] [CrossRef] [Green Version]
- Renfree, M.B.; Shaw, G. Diapause. Annu. Rev. Physiol. 2000, 62, 353–375. [Google Scholar] [CrossRef]
- Lopes, F.L.; Desmarais, J.A.; Murphy, B.D. Embryonic diapause and its regulation. Reproduction 2004, 128, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Bartlewski, P.M.; Beard, A.P.; Cook, S.J.; Chandolia, R.K.; Honaramooz, A.; Rawlings, N.C. Ovarian antral follicular dynamics and their relationships with endocrine variables throughout the oestrous cycle in breeds of sheep differing in prolificacy. Reproduction 1999, 115, 111–124. [Google Scholar] [CrossRef]
- Maijala, K. Review of experience about the use of Finnsheep in improving fertility. In Proceedings of the 2nd World Congress on Sheep and Beef Cattle Breeding, Pretoria, South Africa, 16–19 April 1984; pp. 519–531. [Google Scholar]
- Maijala, K.; Kangasniemi, R. Experiences of out-of-season and twice-a-year lambings in Finnsheep. World Rev. Anim. Prod. 1972, 8, 84. [Google Scholar]
- Österberg, S. Breeding season of the Finnsheep ewe. Acta Agric. Scand. 1981, 31, 11–16. [Google Scholar] [CrossRef]
- Hernández-Montiel, W.; Collí-Dula, R.C.; Ramón-Ugalde, J.P.; Martínez-Núñez, M.A.; Zamora-Bustillos, R. RNA-seq transcriptome analysis in ovarian tissue of Pelibuey breed to explore the regulation of prolificacy. Genes 2019, 10, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, C.L.; Chalupny, N.J.; Schooley, K.; VandenBos, T.; Kubin, M.; Cosman, D. UL16-binding proteins, novel MHC class I-related proteins, bind to NKG2D and activate multiple signaling pathways in primary NK cells. J. Immunol. 2002, 168, 671–679. [Google Scholar] [CrossRef] [Green Version]
- Daftary, G.S.; Taylor, H.S. Endocrine regulation of HOX genes. Endocr. Rev. 2006, 27, 331–355. [Google Scholar] [CrossRef] [PubMed]
- Lappin, T.R.; Grier, D.G.; Thompson, A.; Halliday, H.L. HOX genes: Seductive science, mysterious mechanisms. Ulster Med. J. 2006, 75, 23–31. [Google Scholar] [PubMed]
- Mallo, M.; Alonso, C.R. The regulation of Hox gene expression during animal development. Development 2013, 140, 3951–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, K.D.; Wu, Z.; Irizarry, R.A.; Leek, J.T. Sequencing technology does not eliminate biological variability. Nat. Biotechnol. 2011, 29, 572–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | # CL | # Embryos |
|---|---|---|
| Finnsheep | 4.1 | 2.6 |
| Texel | 1.7 | 1.5 |
| F1 | 3.75 | 1.8 |
| Sample | Breed | Diet | Raw Reads (M) | Clean Reads (M) | Uniquely Aligned Reads (STAR, M) | Aligned Reads (Salmon, M) | ENA Accession Code |
|---|---|---|---|---|---|---|---|
| 1033 | TX | C | 56.4 | 55.9 | 48.0 | 50.7 | ERR3349023 |
| 107A | TX | F | 55.0 | 54.5 | 46.5 | 49.3 | ERR3349025 |
| 107B | TX | F | 55.7 | 55.2 | 48.3 | 49.8 | ERR3349027 |
| 251 | TX | F | 41.8 | 41.2 | 35.4 | 36.9 | ERR3349029 |
| 302 | TX | C | 54.5 | 54.1 | 47.1 | 48.5 | ERR3349031 |
| 312A | FS | F | 54.9 | 54.4 | 47.0 | 49.3 | ERR3349033 |
| 312B | FS | F | 79.2 | 78.4 | 69.2 | 71.1 | ERR3349035 |
| 3609 | FS | C | 54.5 | 54.1 | 47.0 | 49.0 | ERR3349037 |
| 379 | TX | F | 43.1 | 42.7 | 36.0 | 38.3 | ERR3349039 |
| 4208 | F1 | C | 54.3 | 53.8 | 44.8 | 49.0 | ERR3349041 |
| 4271A | F1 | F | 47.2 | 46.8 | 40.1 | 42.4 | ERR3349043 |
| 4271B | F1 | F | 46.9 | 46.4 | 40.0 | 42.1 | ERR3349045 |
| 4519 | F1 | F | 51.6 | 51.1 | 44.5 | 46.3 | ERR3349047 |
| 4563 | F1 | F | 54.1 | 53.7 | 45.7 | 48.6 | ERR3349049 |
| 4590 | F1 | F | 48.5 | 48.1 | 40.0 | 43.5 | ERR3349051 |
| 4823 | F1 | C | 52.6 | 52.1 | 42.6 | 47.2 | ERR3349053 |
| 48 | FS | C | 53.4 | 53.0 | 44.7 | 47.9 | ERR3349055 |
| 554 | FS | F | 56.0 | 55.5 | 48.1 | 49.7 | ERR3349057 |
| 73 | TX | C | 46.1 | 45.7 | 39.1 | 41.3 | ERR3349059 |
| 897 | FS | F | 53.3 | 52.9 | 46.1 | 47.9 | ERR3349061 |
| 974 | FS | C | 45.1 | 44.7 | 38.6 | 40.4 | ERR3349063 |
| Gene ID | Gene Description | Chr | Mean TPM |
|---|---|---|---|
| TIMP1 | Tissue inhibitor of metallopeptidase 1 | X | 16705.6 |
| STAR | Steroidogenic acute regulatory protein | 26 | 14226.6 |
| OXT | oxytocin/neurophysin I prepropeptide | 13 | 13113.2 |
| MGP | matrix Gla protein | 3 | 12294.0 |
| LOC114112617 | Sheep mitochondrial genome | MT | 12155.9 |
| LOC114113966 | translationally controlled tumour protein (TCTP) | 3 | 8341.7 |
| HSD3B1 | hydroxy-delta-5-steroid dehydrogenase, 3 beta- and steroid delta-isomerase 1 | 1 | 8249.3 |
| LOC101117785 | Apolipoprotein A1 (APOA1) | 15 | 8213.2 |
| LOC101110773 | elongation factor 1-alpha 1 (EEF1A1) | 10 | 7564.5 |
| LOC101103639 | uncharacterized (protein coding) | 3 | 7272.8 |
| LOC114116158 | thymosin beta-4 pseudogene | 1 | 6689.3 |
| LOC114118103 | 40S ribosomal protein S29 | 14 | 5599.0 |
| CYP11A1 | Cytochrome P450 Family 11 Subfamily A Member 1 | 18 | 5096.8 |
| RPLP1 | ribosomal protein lateral stalk subunit P1 | 7 | 5032.2 |
| LOC114116632 | 60S ribosomal protein L23a (RPL23a) | 10 | 4998.9 |
| FTH1 | Ferritin heavy chain 1 | 21 | 4498.0 |
| RPS8 | ribosomal protein S8 | 1 | 4402.1 |
| LOC105606567 | 60S ribosomal protein L39 | 4 | 4215.9 |
| MSMB | microseminoprotein beta | 25 | 4093.6 |
| RPS24 | ribosomal protein S24 | 13 | 4029.1 |
| RPS17 | ribosomal protein S17 | 18 | 3950.0 |
| LOC114108766 | 60S ribosomal protein L17 (RPL17) | 4 | 3832.5 |
| RPS18 | ribosomal protein S18 | 20 | 3657.2 |
| TIMP2 | Tissue inhibitor of metallopeptidase 2 | 11 | 3587.4 |
| LOC114114908 | 40S ribosomal protein S27 (RPS27) | 5 | 3570.4 |
| Gene | Gene * | Base Mean | LFC | Padj | Condition |
|---|---|---|---|---|---|
| COX7C | 101.753584 | 1.93 | 0.03584019 | FS vs. TX | |
| LOC114116073 | un_ncRNA | 832.4 | −4.60 | 1.26 × 10−28 | FS vs. TX |
| PAPPA2 | 24.4 | −2.27 | 0.00091 | FS vs. TX | |
| LOC114118704 | TVP23(p) | 38.8 | −2.51 | 1.70 × 10−6 | FS vs. TX |
| ADAMTS4 | 892.8 | −1.80 | 0.02409534 | FS vs. F1 | |
| SHOX2 | 74.0 | 1.95 | 0.0200087 | FS vs. F1 | |
| HOXD10 | 99.3 | −2.17 | 0.00279421 | FS vs. F1 | |
| SCTR | 87.9 | 1.70 | 0.0146455 | FS vs. F1 | |
| HOXC11 | 24.1 | −1.81 | 0.01918401 | FS vs. F1 | |
| LOC105611004 | un_ncRNA | 40.9 | −2.06 | 0.00279421 | FS vs. F1 |
| LOC114116073 | un_ncRNA | 832.4 | −4.82 | 5.58 × 10−31 | FS vs. F1 |
| LRP11 | 125.0 | −1.84 | 0.02732988 | FS vs. F1 | |
| LOC114118704 | TVP23(p) | 38.8 | −1.92 | 0.00279421 | FS vs. F1 |
| BTN1A1 | 22.4 | 1.86 | 0.02926403 | FS vs. F1 | |
| KCNQ1 | 857.0 | 1.77 | 0.04977007 | FS vs. F1 | |
| HNR1PA1 | 81.1 | 1.93 | 0.0200087 | FS vs. F1 | |
| COX7C | 101.7 | −1.88 | 0.03094773 | TX vs. F1 | |
| PCSK5 | 469.2 | 1.69 | 0.01712337 | TX vs. F1 | |
| SCTR | 87.9 | 1.79 | 0.00816462 | TX vs. F1 | |
| HOXC11 | 24.1 | −1.92 | 0.00816462 | TX vs. F1 | |
| PPM1H | 575.3 | 1.67 | 0.00684851 | TX vs. F1 | |
| HOXA9 | 74.2 | −1.96 | 0.00816462 | TX vs. F1 | |
| RDH10 | 2603.9 | 1.56 | 0.00816462 | TX vs. F1 | |
| CA5A | 983.4 | −1.50 | 0.01137482 | TX vs. F1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pokharel, K.; Peippo, J.; Weldenegodguad, M.; Honkatukia, M.; Li, M.-H.; Kantanen, J. Gene Expression Profiling of Corpus luteum Reveals Important Insights about Early Pregnancy in Domestic Sheep. Genes 2020, 11, 415. https://doi.org/10.3390/genes11040415
Pokharel K, Peippo J, Weldenegodguad M, Honkatukia M, Li M-H, Kantanen J. Gene Expression Profiling of Corpus luteum Reveals Important Insights about Early Pregnancy in Domestic Sheep. Genes. 2020; 11(4):415. https://doi.org/10.3390/genes11040415
Chicago/Turabian StylePokharel, Kisun, Jaana Peippo, Melak Weldenegodguad, Mervi Honkatukia, Meng-Hua Li, and Juha Kantanen. 2020. "Gene Expression Profiling of Corpus luteum Reveals Important Insights about Early Pregnancy in Domestic Sheep" Genes 11, no. 4: 415. https://doi.org/10.3390/genes11040415
APA StylePokharel, K., Peippo, J., Weldenegodguad, M., Honkatukia, M., Li, M.-H., & Kantanen, J. (2020). Gene Expression Profiling of Corpus luteum Reveals Important Insights about Early Pregnancy in Domestic Sheep. Genes, 11(4), 415. https://doi.org/10.3390/genes11040415