miR-99a-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting MTMR3 in Chicken


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Standards
2.2. Animals and Samples
2.3. Cell Culture
2.4. RNA Oligonucleotides, Vectors, and Transfection
2.5. RNA Isolation, cDNA Synthesis, and Quantitative Real-Time PCR (qRT-PCR)
2.6. EdU Assay
2.7. Cell Counting Kit 8 (CCK-8) Assay
2.8. Flow Cytometric Cell Cycle Analysis
2.9. Luciferase Reporter Assay
2.10. Western blot Assay
2.11. Immunofluorescence Assay
2.12. Bioinformatic Analysis
2.13. Statistical Analysis
3. Results
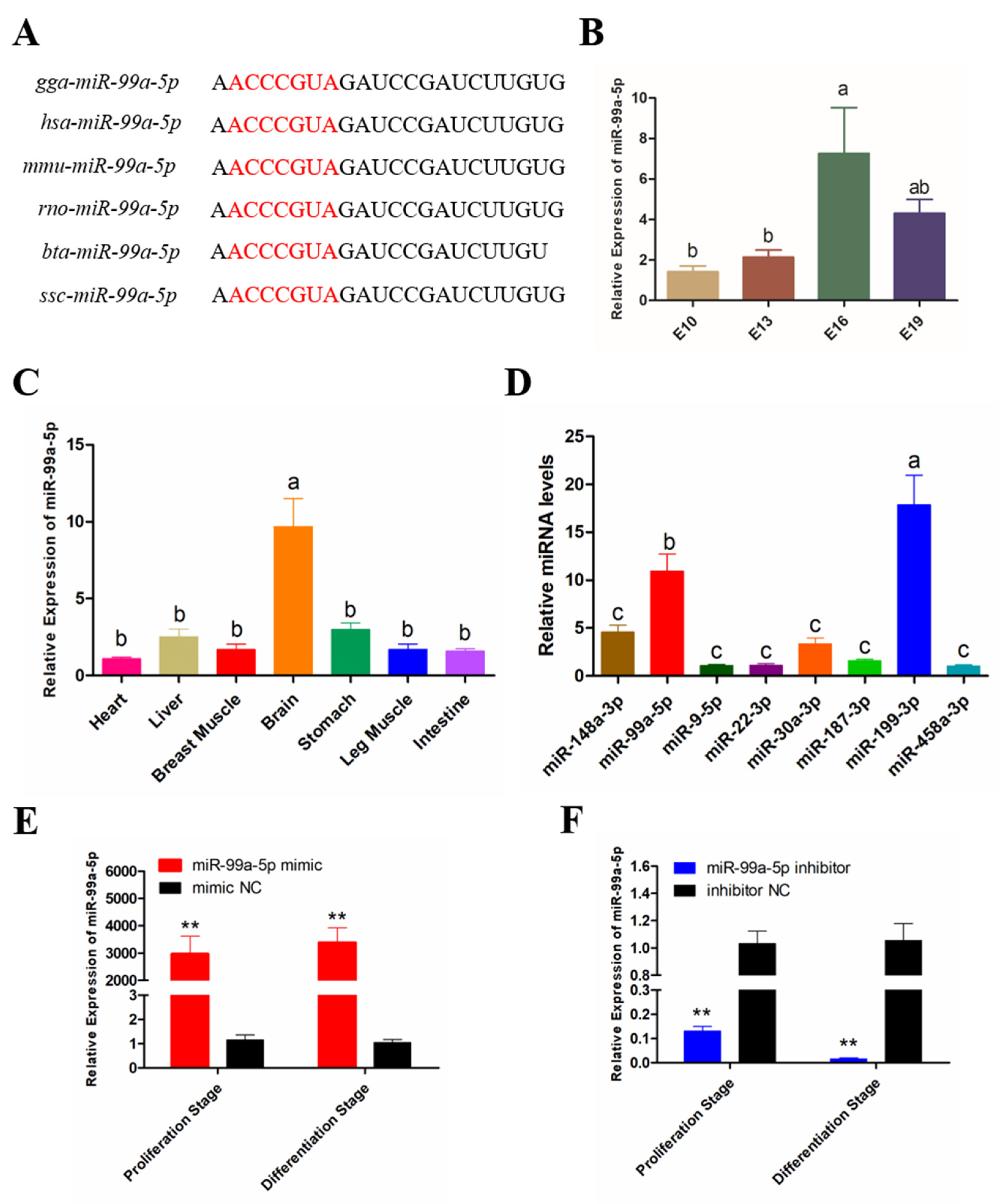
3.1. Expression Pattern of miR-99a-5p in Chicken
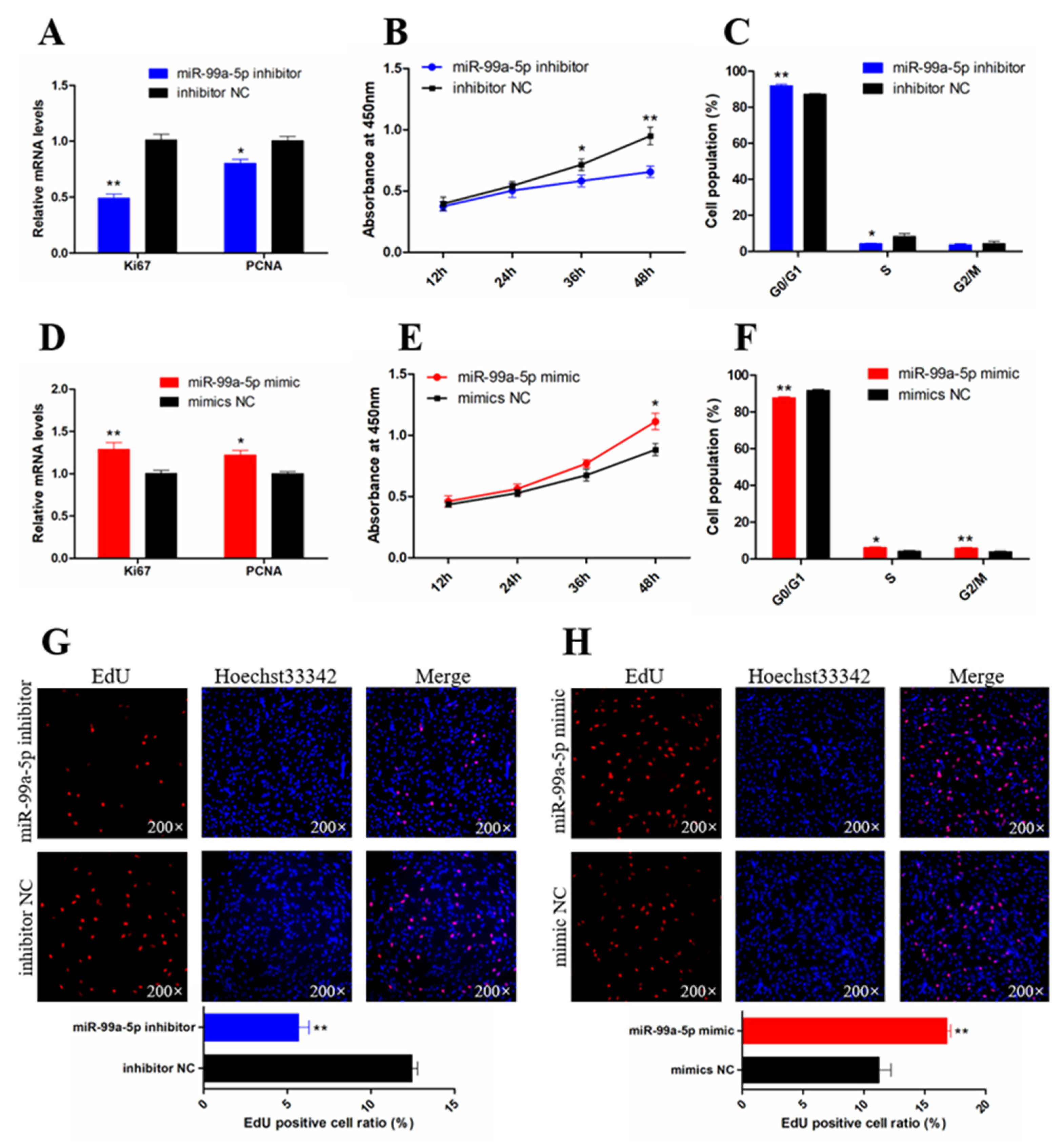
3.2. miR-99a-5p Promotes the Proliferation of Chicken SMSCs
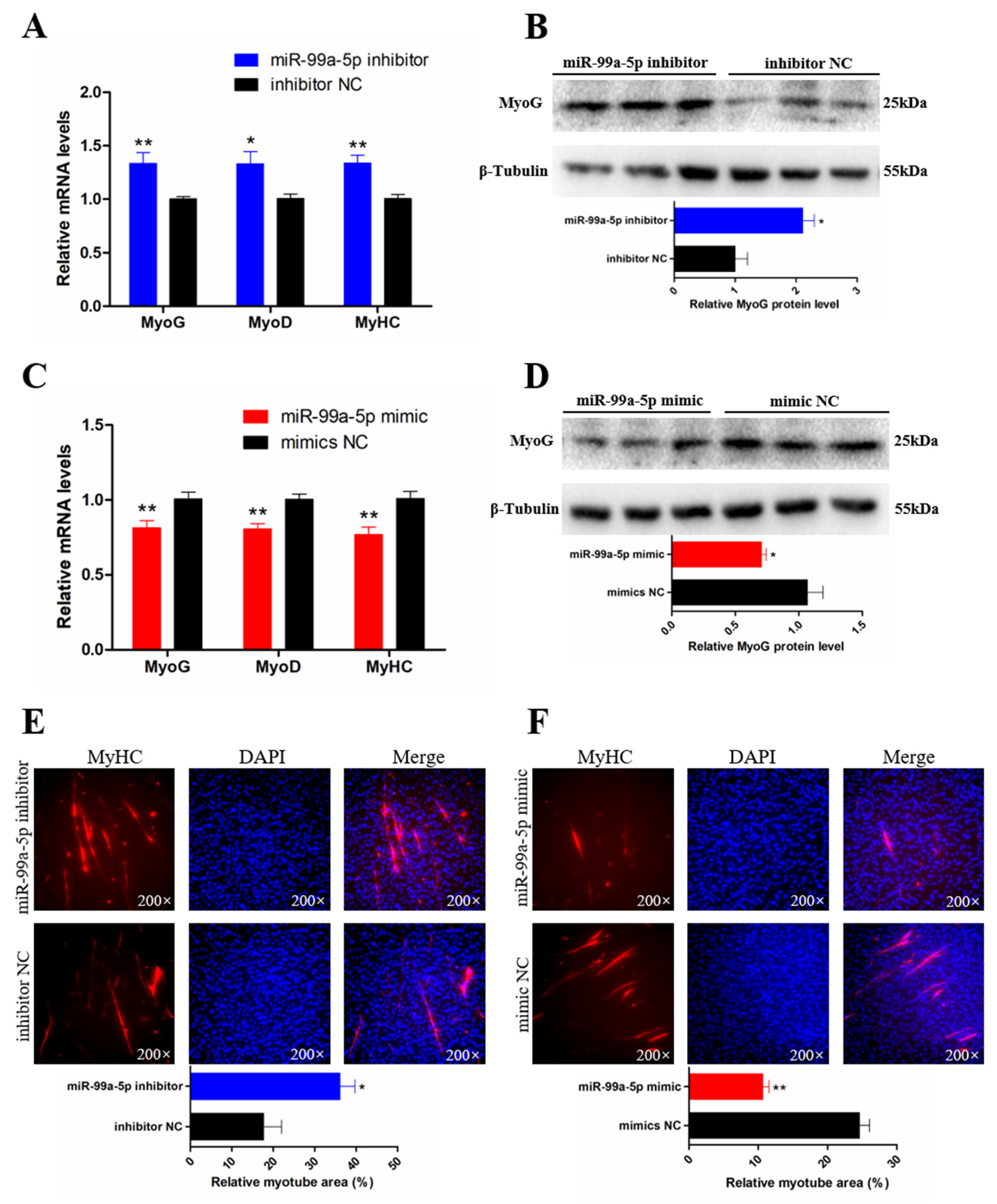
3.3. miR-99a-5p Inhibits the Differentiation of Chicken SMSCs
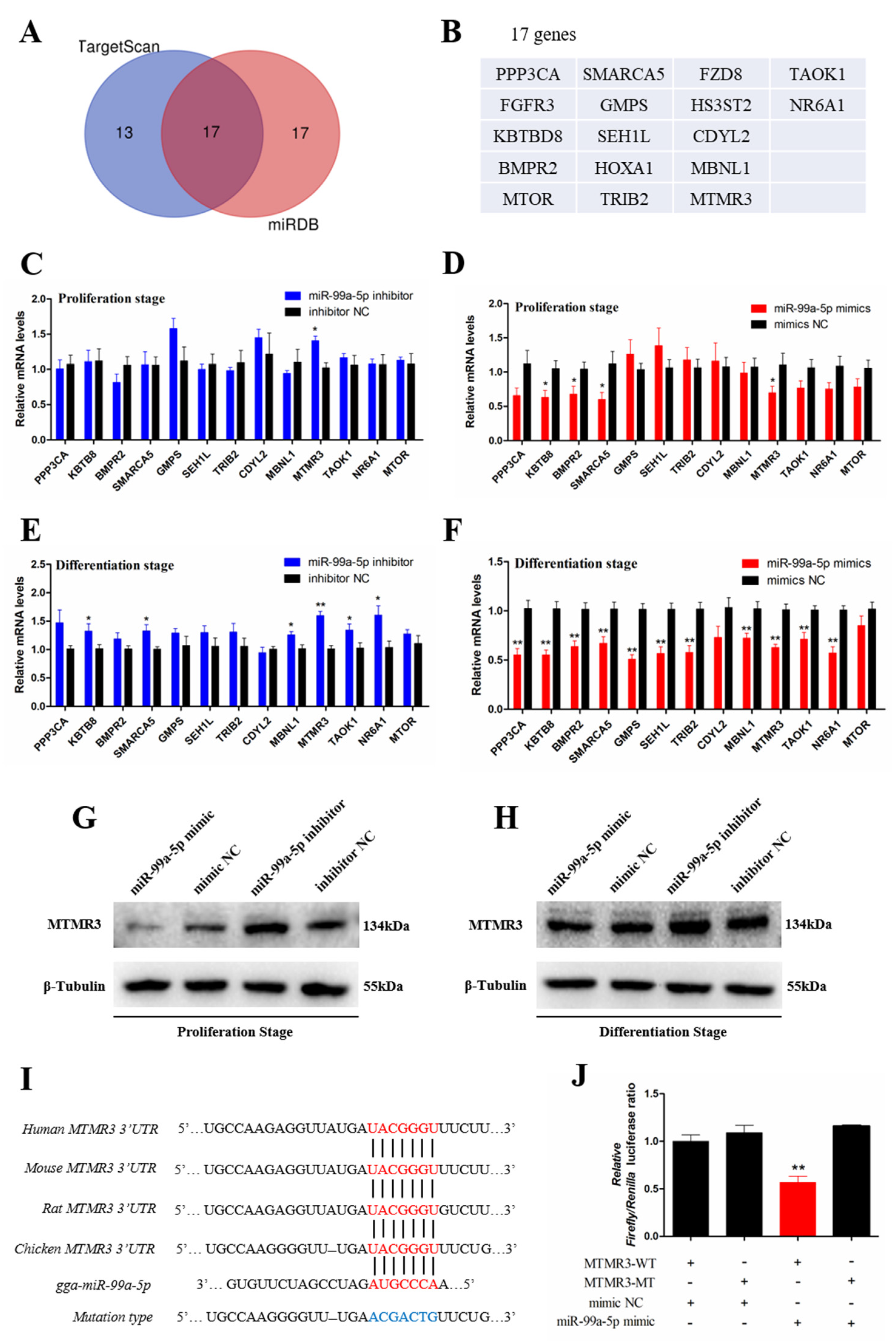
3.4. Target Gene Scanning Revealed that miR-99a-5p Directly Targets the MTMR3 Gene
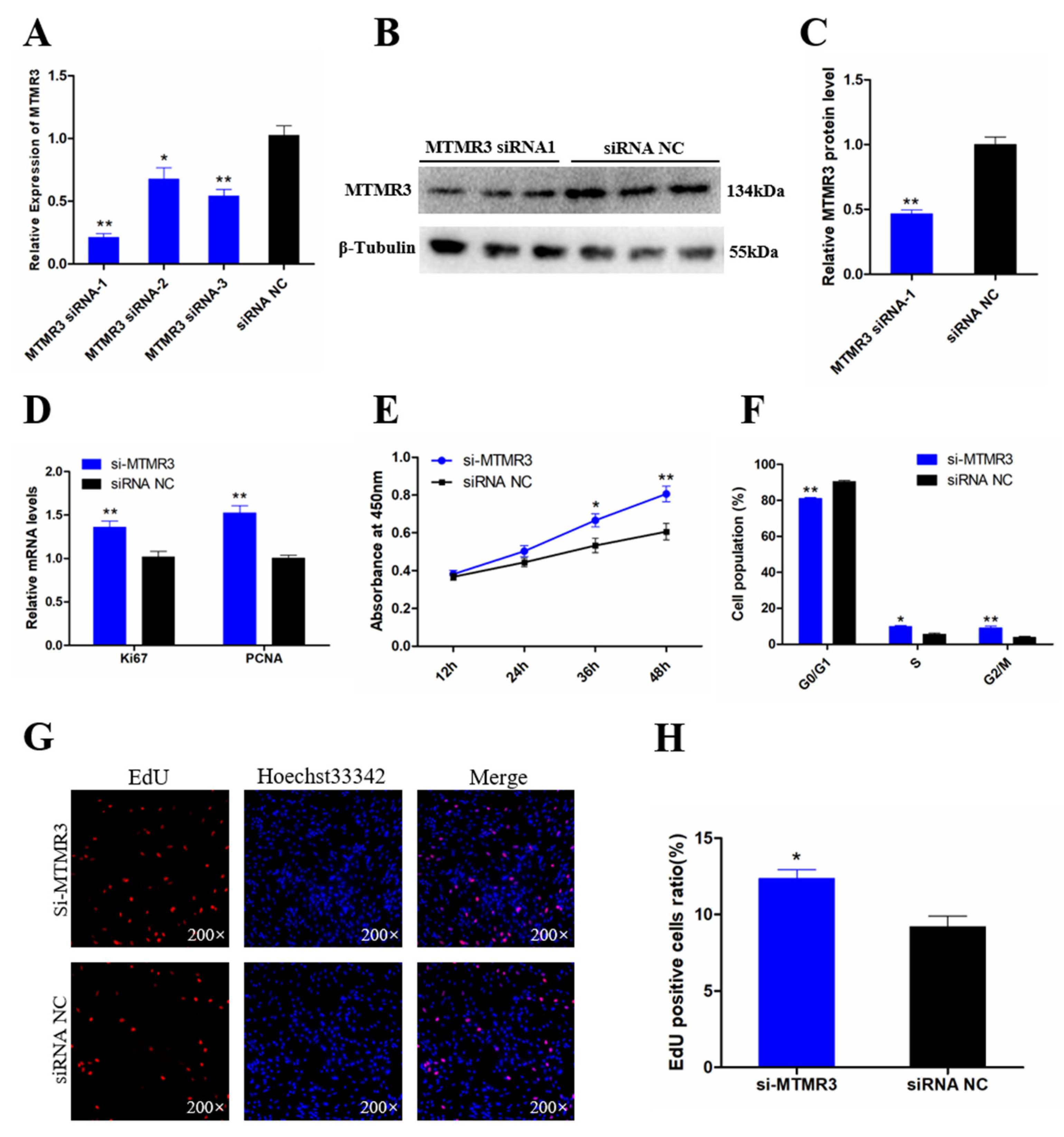
3.5. Knockdown of MTMR3 Facilitates Chicken SMSC Proliferation
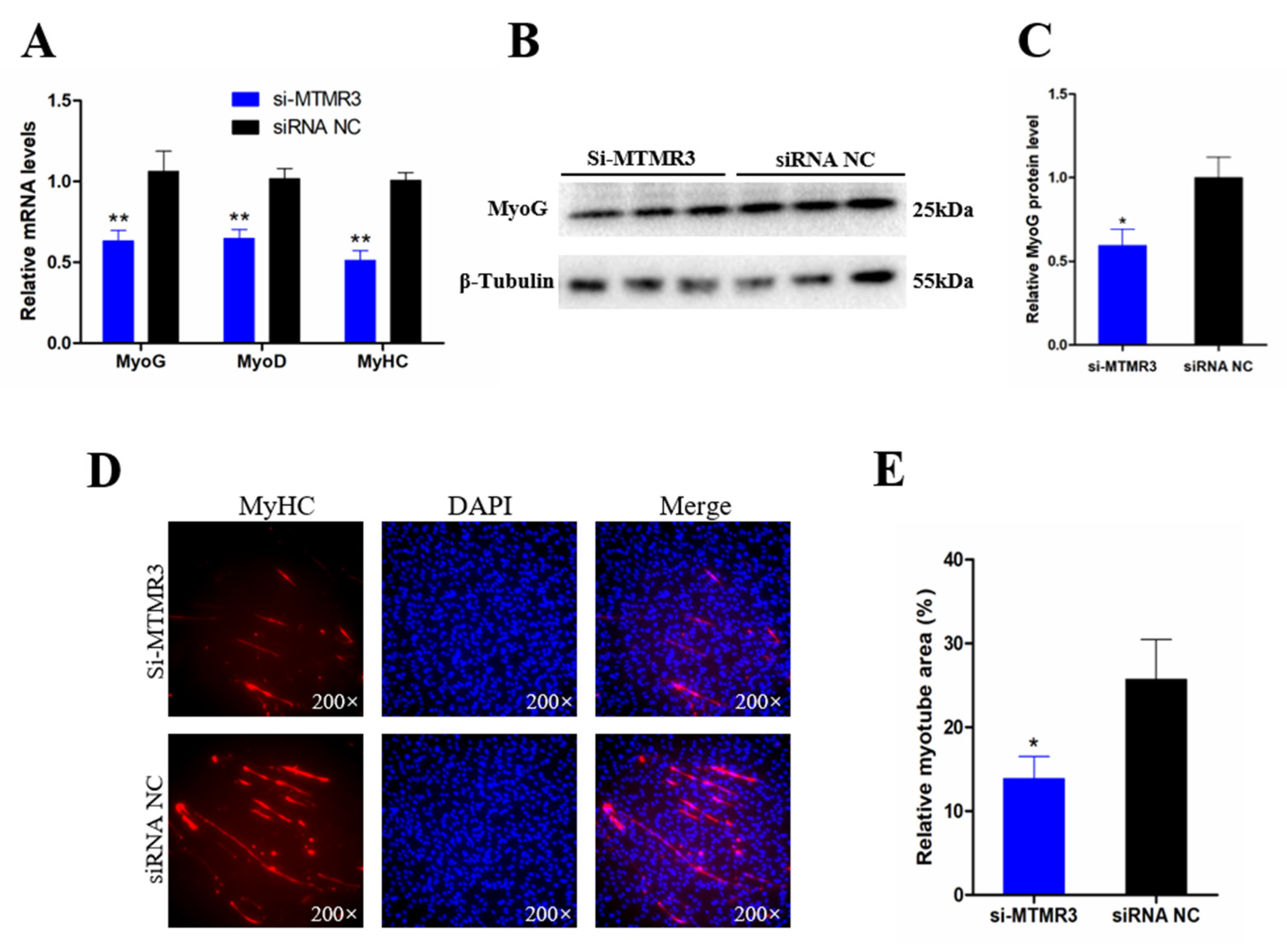
3.6. Knockdown of MTMR3 Represses Chicken SMSC Differentiation
4. Discussion
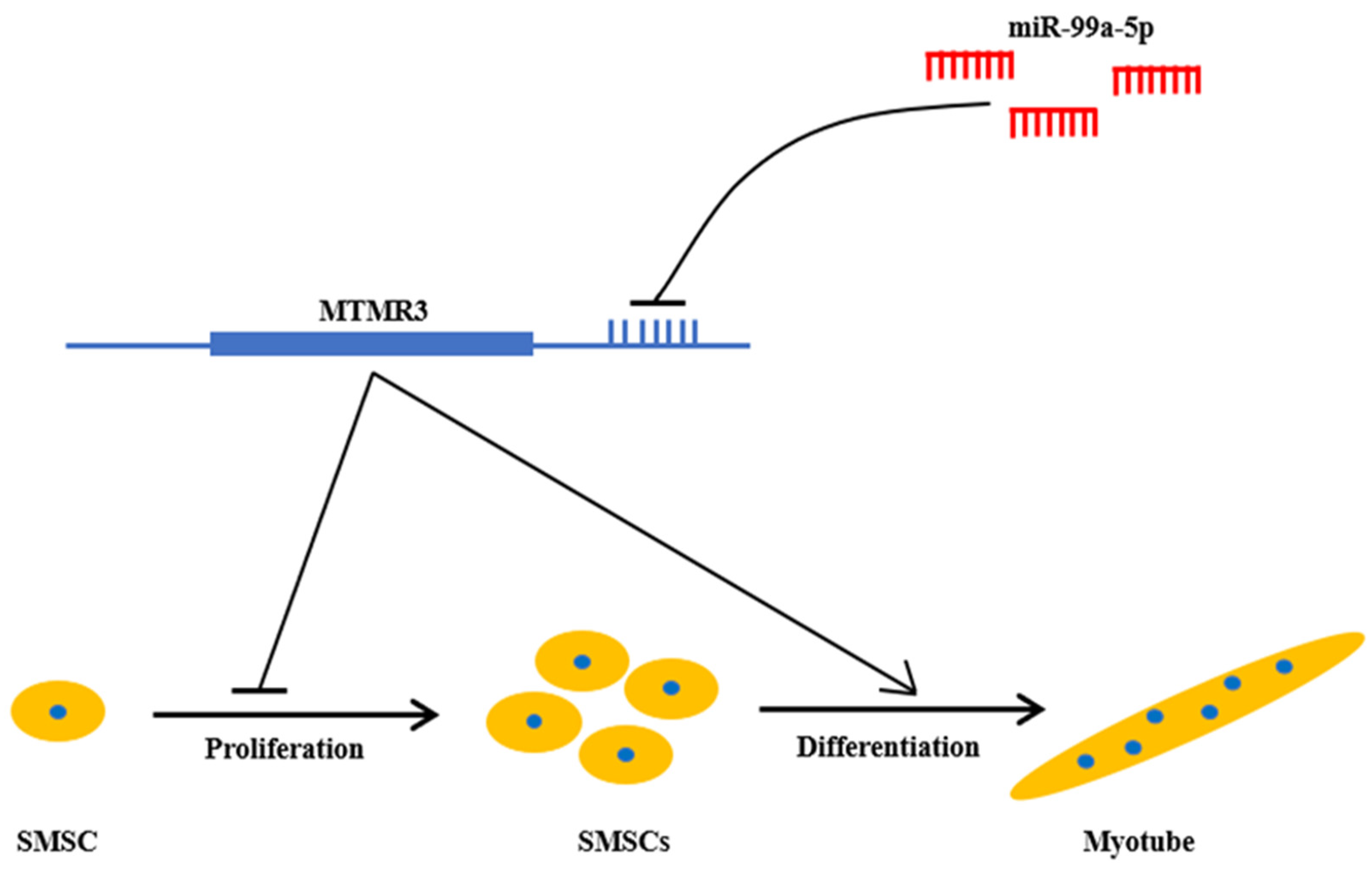
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, J.; Tao, Y.; Li, J.; Deng, Z.; Yan, Z.; Xiao, X.; Wang, D.-Z. microRNA-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7. J. Cell Biol. 2010, 190, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.A. Satellite cell self-renewal. Curr. Opin. Pharmacol. 2006, 6, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Bassel-Duby, R.; Olson, E.N. Signaling pathways in skeletal muscle remodeling. Annu. Rev. Biochem. 2006, 75, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Liu, Z.; Cao, X.; He, H.; Han, S.; Chen, Y.; Cui, C.; Zhao, J.; Li, D.; Wang, Y. Circular RNA profiling identified an abundant circular RNA circTMTC1 that inhibits chicken skeletal muscle satellite cell differentiation by sponging miR-128-3p. Int. J. Biol. Sci. 2019, 15, 2265–2281. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Chekulaeva, M.; Filipowicz, W. Mechanisms of miRNA-mediated post-transcriptional regulation in animal cells. Curr. Opin. Cell Biol. 2009, 21, 452–460. [Google Scholar] [CrossRef]
- Chen, J.-F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.-Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef]
- Kim, H.K.; Lee, Y.S.; Sivaprasad, U.; Malhotra, A.; Dutta, A. Muscle-specific microRNA miR-206 promotes muscle differentiation. J. Cell Biol. 2006, 174, 677–687. [Google Scholar] [CrossRef]
- Xin, J.X.; Yue, Z.; Zhang, S.; Jiang, Z.H.; Wang, P.Y.; Li, Y.J.; Pang, M.; Xie, S.Y. miR-99 inhibits cervical carcinoma cell proliferation by targeting TRIB2. Oncol. Lett. 2013, 6, 1025–1030. [Google Scholar] [CrossRef]
- Sun, D.; Lee, Y.S.; Malhotra, A.; Kim, H.K.; Matecic, M.; Evans, C.; Jensen, R.V.; Moskaluk, C.A.; Dutta, A. miR-99 family of MicroRNAs suppresses the expression of prostate-specific antigen and prostate cancer cell proliferation. Cancer Res. 2011, 71, 1313–1324. [Google Scholar] [CrossRef]
- Chen, D.; Chen, Z.; Jin, Y.; Dragas, D.; Zhang, L.; Adjei, B.S.; Wang, A.; Dai, Y.; Zhou, X. MicroRNA-99 family members suppress Homeobox A1 expression in epithelial cells. PLoS ONE 2013, 8, e80625. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.M.; Urbé, S.; Dove, S.K.; Tenza, D.; Raposo, G.; Clague, M.J. Characterization of MTMR3: An inositol lipid 3-phosphatase with novel substrate specificity. Curr. Biol. 2001, 11, 1600–1605. [Google Scholar] [CrossRef]
- Hao, F.; Itoh, T.; Morita, E.; Shirahama-Noda, K.; Yoshimori, T.; Noda, T. The PtdIns3-phosphatase MTMR3 interacts with mTORC1 and suppresses its activity. FEBS Lett. 2016, 590, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Shen, X.; Cao, X.; He, H.; Han, S.; Chen, Y.; Cui, C.; Wei, Y.; Wang, Y.; Li, D. HDAC4 Regulates the Proliferation, Differentiation and Apoptosis of Chicken Skeletal Muscle Satellite Cells. Animals 2020, 10, 84. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yu, J.; Guo, L.; Byers, M.S.; Wang, Z.; Chen, X.; Xu, H.; Nie, Q. Circular RNA circHIPK3 promotes the proliferation and differentiation of chicken myoblast cells by sponging miR-30a-3p. Cells 2019, 8, 177. [Google Scholar] [CrossRef]
- Jia, L.; Li, Y.-F.; Wu, G.-F.; Song, Z.-Y.; Lu, H.-Z.; Song, C.-C.; Zhang, Q.-L.; Zhu, J.-Y.; Yang, G.-S.; Shi, X.-E. MiRNA-199a-3p regulates C2C12 myoblast differentiation through IGF-1/AKT/mTOR signal pathway. Int. J. Mol. Sci. 2014, 15, 296–308. [Google Scholar] [CrossRef]
- Khalaj, M.; Woolthuis, C.M.; Hu, W.; Durham, B.H.; Chu, S.H.; Qamar, S.; Armstrong, S.A.; Park, C.Y. miR-99 regulates normal and malignant hematopoietic stem cell self-renewal. J. Exp. Med. 2017, 214, 2453–2470. [Google Scholar] [CrossRef]
- Khalaj, M.; Hu, W.; Park, C.Y. Mir-99 Is a Regulator of Hematopoietic and Leukemic Stem Cell Differentiation; American Society of Hematology: Washington, DC, USA, 2015. [Google Scholar]
- Mueller, A.C.; Sun, D.; Dutta, A. The miR-99 family regulates the DNA damage response through its target SNF2H. Oncogene 2013, 32, 1164–1172. [Google Scholar] [CrossRef]
- Gu, S.; Jin, L.; Zhang, F.; Sarnow, P.; Kay, M.A. Biological basis for restriction of microRNA targets to the 3′ untranslated region in mammalian mRNAs. Nat. Struct. Mol. Biol. 2009, 16, 144. [Google Scholar] [CrossRef]
- Kim, D.H.; Sætrom, P.; Snøve, O.; Rossi, J.J. MicroRNA-directed transcriptional gene silencing in mammalian cells. Proc. Natl. Acad. Sci. USA 2008, 105, 16230–16235. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Z.; Hou, Y.; Zhang, K.; Peng, X. gga-miR-99a targets SMARCA5 to regulate Mycoplasma gallisepticum (HS strain) infection by depressing cell proliferation in chicken. Gene 2017, 627, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chang, L.; Li, Z.; Gao, Q.; Cai, D.; Tian, Y.; Zeng, L.; Li, M. miR-99a and-99b inhibit cervical cancer cell proliferation and invasion by targeting mTOR signaling pathway. Med. Oncol. 2014, 31, 934. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Qu, L.; Wang, Y.; Cong, J.; Wang, W.; Yang, X. miR-99a promotes proliferation targeting FGFR3 in human epithelial ovarian cancer cells. Biomed. Pharmacother. 2014, 68, 163–169. [Google Scholar] [CrossRef]
- Friday, B.B.; Mitchell, P.O.; Kegley, K.M.; Pavlath, G.K. Calcineurin initiates skeletal muscle differentiation by activating MEF2 and MyoD. Differ. Orig. Artic. 2003, 71, 217–227. [Google Scholar] [CrossRef]
- Zanchi, N.E.; Lancha, A.H. Mechanical stimuli of skeletal muscle: implications on mTOR/p70s6k and protein synthesis. Eur. J. Appl. Physiol. 2008, 102, 253–263. [Google Scholar] [CrossRef]
- Kuo, Y.Z.; Tai, Y.H.; Lo, H.I.; Chen, Y.L.; Cheng, H.C.; Fang, W.Y.; Lin, S.H.; Yang, C.L.; Tsai, S.T.; Wu, L.W. MiR-99a exerts anti-metastasis through inhibiting myotubularin-related protein 3 expression in oral cancer. Oral Dis. 2014, 20, e65–e75. [Google Scholar] [CrossRef]
- Zou, J.; Chang, S.-C.; Marjanovic, J.; Majerus, P.W. MTMR9 increases MTMR6 enzyme activity, stability, and role in apoptosis. J. Biol. Chem. 2009, 284, 2064–2071. [Google Scholar] [CrossRef]
- Yuan, Z.; Chen, Y.; Zhang, X.; Zhou, X.; Li, M.; Chen, H.; Wu, M.; Zhang, Y.; Mo, D. Silencing myotubularin related protein 7 enhances proliferation and early differentiation of C2C12 myoblast. Biochem. Biophys. Res. Commun. 2017, 484, 592–597. [Google Scholar] [CrossRef]
- Mei, J.; Li, Z.; Gui, J.-F. Cooperation of Mtmr8 with PI3K regulates actin filament modeling and muscle development in zebrafish. PLoS ONE 2009, 4, e4979. [Google Scholar] [CrossRef]
- Shen, J.; Yu, W.-M.; Brotto, M.; Scherman, J.A.; Guo, C.; Stoddard, C.; Nosek, T.M.; Valdivia, H.H.; Qu, C.-K. Deficiency of MIP/MTMR14 phosphatase induces a muscle disorder by disrupting Ca2+ homeostasis. Nat. Cell Biol. 2009, 11, 769–776. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) |
|---|---|
| MTMR3 siRNA-1 | CCAGGAGAACACGUAACUUTT |
| AAGUUACGUGUUCUCCUGGTT | |
| MTMR3 siRNA-2 | CCUAGAGACUGCAGGUAAATT |
| UUUACCUGCAGUCUCUAGGTT | |
| MTMR3 siRNA-3 | GCGUCUCAACGGUGACUAUTT |
| AUAGUCACCGUUGAGACGCTT | |
| siRNA NC | UUCUCCGAACGUGUCACGUTT |
| ACGUGACACGUUCGGAGAATT | |
| miR-99a-5p mimic | AACCCGUAGAUCCGAUCUUGUG |
| Mimic NC | UUGUACUACACAAAAGUACUG |
| miR-99a-5p inhibitor | CACAAGAUCGGAUCUACGGGUU |
| Inhibitor NC | CAGUACUUUUGUGUAGUACAA |
| Gene | Primer Sequence (5′–3′) | Length (bp) |
|---|---|---|
| MyoD1 | F: GCCGCCGATGACTTCTATGA | 66 |
| R: CAGGTCCTCGAAGAAGTGCAT | ||
| MyoG | F: CGTGTGCCACAGCCAATG | 63 |
| R: CCGCCGGAGAGAGACCTT | ||
| MyHC | F: GAAGGAGACCTCAACGAGATGG | 138 |
| R: ATTCAGGTGTCCCAAGTCATCC | ||
| Ki67 | F: GCAACAACAAGGAGGCTTCG | 93 |
| R: TTCAGGTGCCATCCCGTAAC | ||
| PCNA | F: AACACTCAGAGCAGAAGAC | 225 |
| R: GCACAGGAGATGACAACA | ||
| MBNL1 | F: GTGTCATCGCCTGCTT | 133 |
| R: ATGTTCTTCTGCTGGATCA | ||
| PPP3CA | F: CTCATCCTTACTGGCTTCC | 219 |
| R: TCGCTCTTATCTTGTTCCTT | ||
| KBTBD8 | F: CAAGTCTGTGGAGTGTTATG | 237 |
| R: GCTATGTAGTAGATGGAGTCA | ||
| BMPR2 | F: TTAGTCCAACAGTCAATCCA | 195 |
| R: AAGTCAGCGGCGTAGT | ||
| SMARCA5 | F: TTGTCAGAGATGTGTTGTTG | 199 |
| R: GTTAGCAGCAGCCGATT | ||
| GMPS | F: CAAGAAGAGCGTCAGAGAA | 132 |
| R: CCATCAGCAACCTTATCCA | ||
| SEH1L | F: AGCCAATACAAGCCTACAG | 154 |
| R: GTCAGCATCGCAAGAGT | ||
| TRIB2 | F: TGACATTGAGCCTAGTTCC | 206 |
| R: CTTCCTTAGCACCATATCCT | ||
| CDYL2 | F: AGTAATGAATCGCCTGTTGT | 140 |
| R: TGACGCACGCTGTATCT | ||
| TAOK1 | F: GGAGGAGGAAGAGGAACA | 106 |
| R: TTGGCTACTGGCACTGA | ||
| NR6A1 | F: ATGGAGCAACAATGGAGAC | 203 |
| R: CCGAGTGAGCAGAATAGC | ||
| MTOR | F: GGAATGAACCGTGATGAC | 121 |
| R: GCTGCTGCTGAGTGAT | ||
| MTMR3 | F: TGTAGGACGAGCAGAAGA | 227 |
| R: GGCGGATGGCATTGTT | ||
| GAPDH | F: CCAGAACATCATCCCAGCGTC | 136 |
| R: ACGGCAGGTCAGGTCAACAA | ||
| miR-99a-5p | F: AACCCGTAGATCCGATCTTGTG | / |
| R: CAGGTCCAGTTTTTTTTTTTTTT | ||
| U6 | F: GGGCCATGCTAATCTTCTCTGTA | / |
| R: CAGGTCCAGTTTTTTTTTTTTTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, X.; Tang, S.; Du, F.; Li, H.; Shen, X.; Li, D.; Wang, Y.; Zhang, Z.; Xia, L.; Zhu, Q.; et al. miR-99a-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting MTMR3 in Chicken. Genes 2020, 11, 369. https://doi.org/10.3390/genes11040369
Cao X, Tang S, Du F, Li H, Shen X, Li D, Wang Y, Zhang Z, Xia L, Zhu Q, et al. miR-99a-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting MTMR3 in Chicken. Genes. 2020; 11(4):369. https://doi.org/10.3390/genes11040369
Chicago/Turabian StyleCao, Xinao, Shuyue Tang, Fei Du, Hao Li, Xiaoxu Shen, Diyan Li, Yan Wang, Zhichao Zhang, Lu Xia, Qing Zhu, and et al. 2020. "miR-99a-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting MTMR3 in Chicken" Genes 11, no. 4: 369. https://doi.org/10.3390/genes11040369
APA StyleCao, X., Tang, S., Du, F., Li, H., Shen, X., Li, D., Wang, Y., Zhang, Z., Xia, L., Zhu, Q., & Yin, H. (2020). miR-99a-5p Regulates the Proliferation and Differentiation of Skeletal Muscle Satellite Cells by Targeting MTMR3 in Chicken. Genes, 11(4), 369. https://doi.org/10.3390/genes11040369