De Novo Transcriptome Identifies Olfactory Genes in Diachasmimorpha longicaudata (Ashmead)

Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. RNA Isolation, Library Construction, and Deep Sequencing
2.3. Transcriptome Assembly
2.4. Annotation for the Transcriptome
2.5. Likely Protein Identification and Annotation
2.6. Noncoding Genes and microRNA Genes
2.7. Differential Expression Analysis
2.8. Function Enrichment Analysis
2.9. Phylogenetic Analysis
2.10. qRT-PCR Verification
3. Results
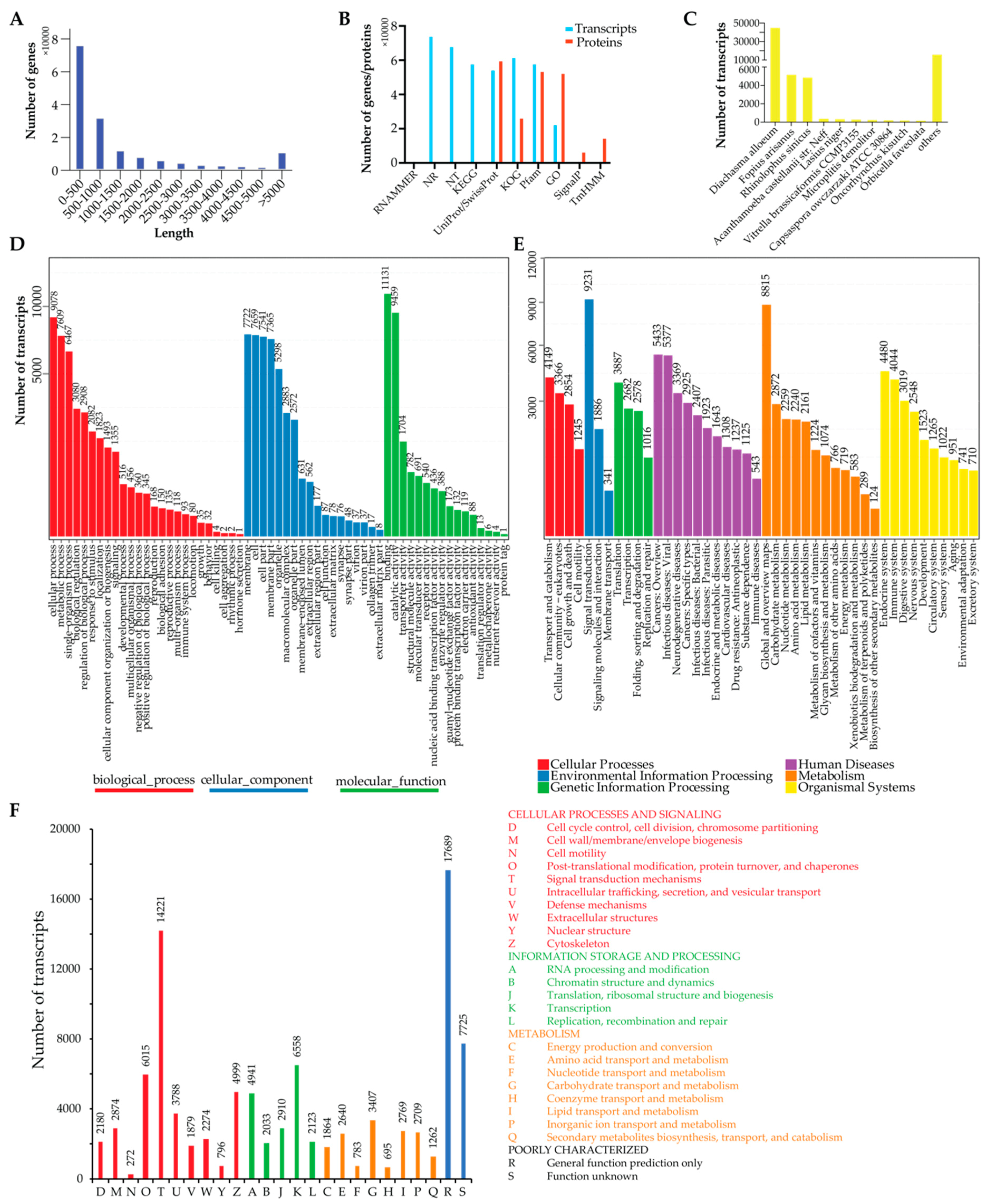
3.1. Animal and Transcriptome Sequencing
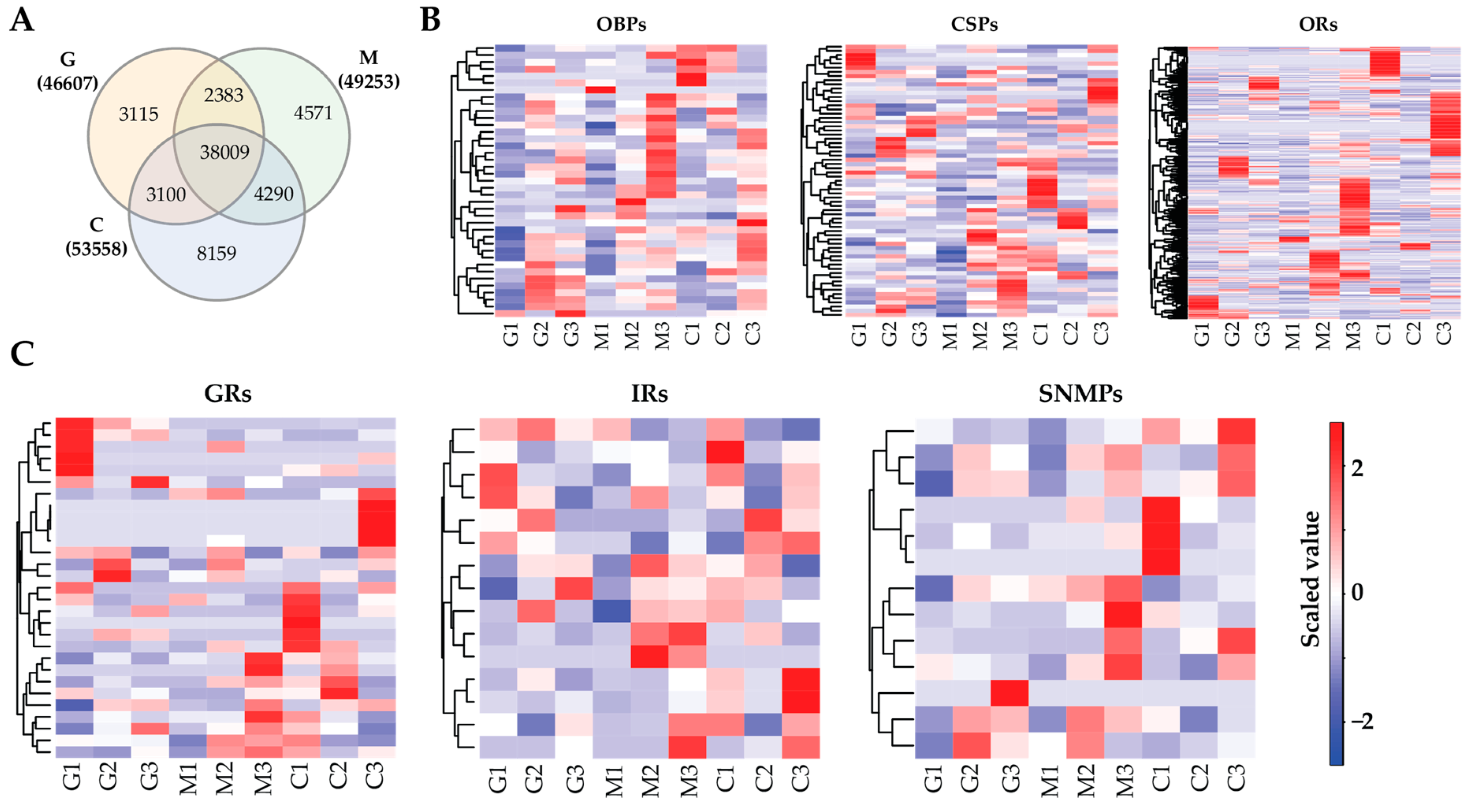
3.2. Transcriptome Annotation
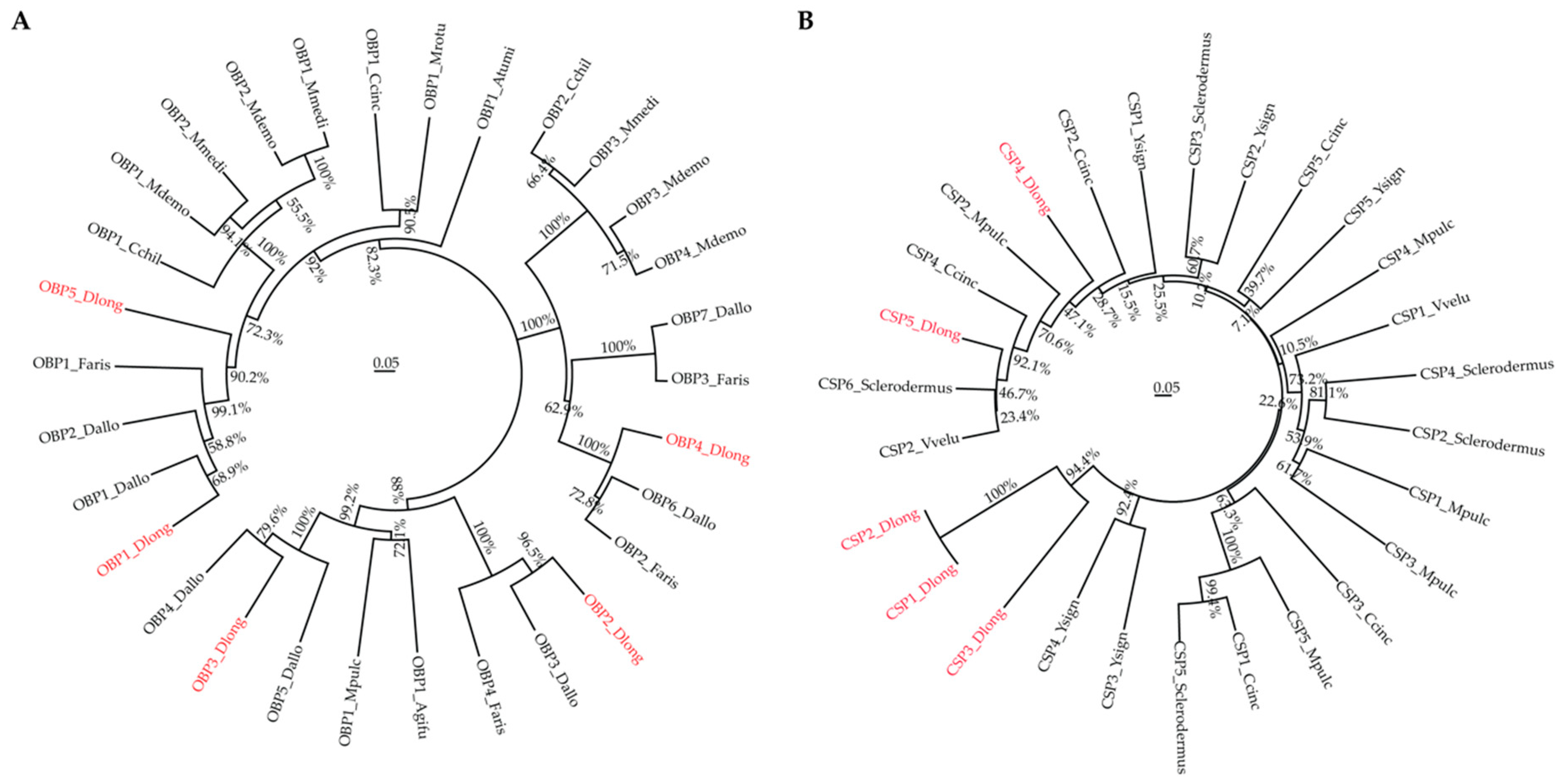
3.3. Olfactory Genes
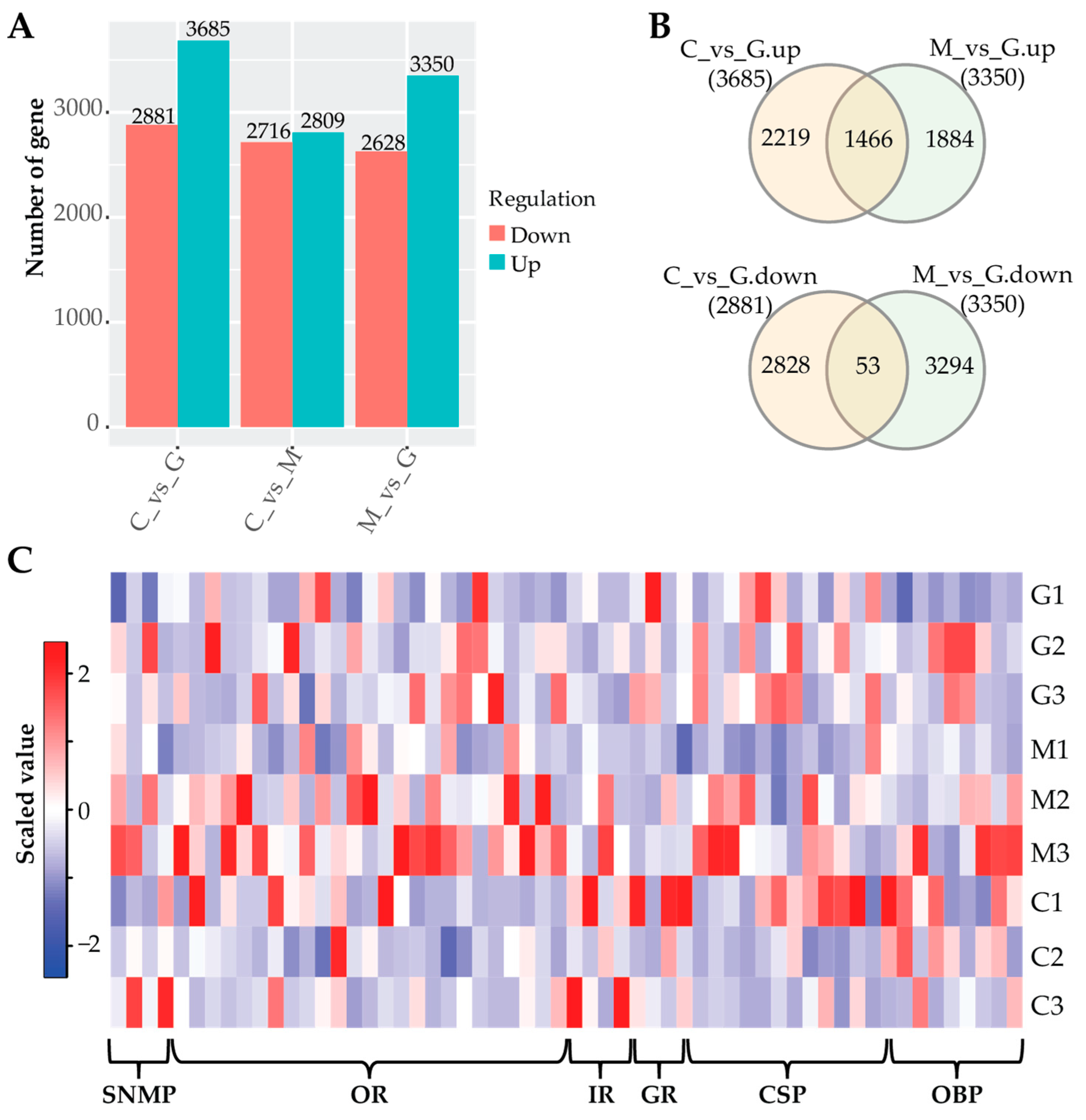
3.4. Gene Expression Profile
3.5. Phylogenetic Analysis
3.6. Differential Expression Analysis
3.7. qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International: Wallingford, UK, 1992. [Google Scholar]
- Lawrence, P.O. Host vibration—A cue to host location by the parasite, Biosteres longicaudatus. Oecologia 1981, 48, 249–251. [Google Scholar] [CrossRef]
- Greany, P.D.; Tumlinson, J.H.; Chambers, D.L.; Boush, G.M. Chemically mediated host finding byBiosteres (Opius) longicaudatus, a parasitoid of tephritid fruit fly larvae. J. Chem. Ecol. 1977, 3, 189–195. [Google Scholar] [CrossRef]
- Carrasco, M.; Montoya, P.; Cruz-lopez, L.; Rojas, J.C. Response of the Fruit Fly Parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae) to Mango Fruit Volatiles. Environ. Entomol. 2005, 34, 576–583. [Google Scholar] [CrossRef]
- Silva, J.W.P.; Bento, J.M.S.; Zucchi, R.A. Olfactory response of three parasitoid species (Hymenoptera: Braconidae) to volatiles of guavas infested or not with fruit fly larvae (Diptera: Tephritidae). Biol. Control. 2007, 41, 304–311. [Google Scholar] [CrossRef]
- Duan, J.J.; Messing, R.H. Effects of host substrate and vibration cues on ovipositor-probing behavior in two larval parasitoids of tephritid fruit flies. J. Insect Behav. 2000, 13, 175–186. [Google Scholar] [CrossRef]
- Stuhl, C.; Sivinski, J.; Teal, P.; Paranhos, B.; Aluja, M. A compound produced by fruigivorous Tephritidae (Diptera) larvae promotes oviposition behavior by the biological control agent Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Environ. Entomol. 2011, 40, 727–736. [Google Scholar] [CrossRef]
- De Bruyne, M.; Warr, C.G. Molecular and cellular organization of insect chemosensory neurons. Bioessays 2006, 28, 23–34. [Google Scholar] [CrossRef]
- Zubkov, S.; Gronenborn, A.M.; Byeon, I.J.; Mohanty, S. Structural consequences of the pH-induced conformational switch in A. polyphemus pheromone-binding protein: Mechanisms of ligand release. J. Mol. Biol. 2005, 354, 1081–1090. [Google Scholar] [CrossRef]
- Vogt, R.G.; Riddiford, L.M.; Prestwich, G.D. Kinetic properties of a sex pheromone-degrading enzyme: the sensillar esterase of Antheraea polyphemus. Proc. Natl. Acad. Sci. USA 1985, 82, 8827–8831. [Google Scholar] [CrossRef]
- Hull, J.J.; Perera, O.P.; Snodgrass, G.L. Cloning and expression profiling of odorant-binding proteins in the tarnished plant bug, Lygus lineolaris. Insect Mol. Biol. 2014, 23, 78–97. [Google Scholar] [CrossRef]
- Sanchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity (Edinb.) 2009, 103, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Dong, X.; Xu, X.; Ren, S. cDNA cloning and recombinant expression of the general odorant binding protein II from Spodoptera litura. Sci. China C Life Sci. 2009, 52, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, Y.; Akami, M.; Niu, C.Y. Identification of olfactory genes and functional analysis of BminCSP and BminOBP21 in Bactrocera minax. PLoS ONE 2019, 14, e0222193. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Yang, P.; Wu, G.; He, Q.; Yin, L.; Xiong, Z. Clone and sequence analysis of the odorant-binding protein genes in Nasonia vitripennis. J. Environ. Entomol. 2010, 32, 476–482. [Google Scholar]
- Zhang, K. Characterization of MmedOBP8 and Its Binding Capacity in Microplitis mediator (Halidag); Huazhong Agricultural University: Wuhan, China, 2011. [Google Scholar]
- Zhao, Y.; Wang, F.; Zhang, X.; Zhang, S.; Guo, S.; Zhu, G.; Liu, Q.; Li, M. Transcriptome and Expression Patterns of Chemosensory Genes in Antennae of the Parasitoid Wasp Chouioia cunea. PLoS ONE 2016, 11, e0148159. [Google Scholar] [CrossRef]
- Harbi, A.; de Pedro, L.; Ferrara, F.A.A.; Tormos, J.; Chermiti, B.; Beitia, F.; Sabater-Munoz, B. Diachasmimorpha longicaudata Parasitism Response to Medfly Host Fruit and Fruit Infestation Age. Insects 2019, 10, 211. [Google Scholar] [CrossRef]
- Chen, M.; Xu, R.; Rai, A.; Suwakulsiri, W.; Izumikawa, K.; Ishikawa, H.; Greening, D.W.; Takahashi, N.; Simpson, R.J. Distinct shed microvesicle and exosome microRNA signatures reveal diagnostic markers for colorectal cancer. PLoS ONE 2019, 14, e0210003. [Google Scholar] [CrossRef]
- Chen, M.; Xu, R.; Ji, H.; Greening, D.W.; Rai, A.; Izumikawa, K.; Ishikawa, H.; Takahashi, N.; Simpson, R.J. Transcriptome and long noncoding RNA sequencing of three extracellular vesicle subtypes released from the human colon cancer LIM1863 cell line. Sci. Rep. 2016, 6, 38397. [Google Scholar] [CrossRef]
- Wei, S.; Ma, X.; Pan, L.; Miao, J.; Fu, J.; Bai, L.; Zhang, Z.; Guan, Y.; Mo, C.; Huang, H.; et al. Transcriptome Analysis of Taxillusi chinensis (DC.) Danser Seeds in Response to Water Loss. PLoS ONE 2017, 12, e0169177. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Lagesen, K.; Hallin, P.; Rodland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.; Forslund, K.; Szklarczyk, D.; Trachana, K.; Roth, A.; Huerta-Cepas, J.; Gabaldon, T.; Rattei, T.; Creevey, C.; Kuhn, M.; et al. eggNOG v4.0: Nested orthology inference across 3686 organisms. Nucleic Acids Res. 2014, 42, D231–D239. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–349. [Google Scholar] [CrossRef]
- Li, R.; Yu, C.; Li, Y.; Lam, T.W.; Yiu, S.M.; Kristiansen, K.; Wang, J. SOAP2: An improved ultrafast tool for short read alignment. Bioinformatics 2009, 25, 1966–1967. [Google Scholar] [CrossRef]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human MicroRNA targets. PLoS Biol 2004, 2, e363. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J. Odorant-binding proteins in insects. Vitam. Horm. 2010, 83, 241–272. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.S.; Sun, Z.; Ma, W.; Chen, W.; Wang, M.Q. De novo analysis of the Nilaparvata lugens (Stal) antenna transcriptome and expression patterns of olfactory genes. Comp. Biochem. Physiol. Part D Genomics Proteomics 2014, 9, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Hekmat-Scafe, D.S.; Scafe, C.R.; McKinney, A.J.; Tanouye, M.A. Genome-wide analysis of the odorant-binding protein gene family in Drosophila melanogaster. Genome Res. 2002, 12, 1357–1369. [Google Scholar] [CrossRef]
- Gong, M.; Jiao, L.; Ma, W. Orthogonal immune algorithm with diversity-based selection for numerical optimization. In Proceedings of the First ACM/SIGEVO Summit on Genetic and Evolutionary Computation, Shanghai, China, 12–14 June 2009; pp. 141–148. [Google Scholar]
- Vogt, R.G.; Prestwich, G.D.; Lerner, M.R. Odorant-binding-protein subfamilies associate with distinct classes of olfactory receptor neurons in insects. J. Neurobiol. 1991, 22, 74–84. [Google Scholar] [CrossRef]
- Vogt, R.G.; Riddiford, L.M. Pheromone binding and inactivation by moth antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef]
- Picimbon, J.F.; Dietrich, K.; Angeli, S.; Scaloni, A.; Krieger, J.; Breer, H.; Pelosi, P. Purification and molecular cloning of chemosensory proteins from Bombyx mori. Arch. Insect. Biochem. Physiol. 2000, 44, 120–129. [Google Scholar] [CrossRef]
- Ma, C.; Cui, S.; Tian, Z.; Zhang, Y.; Chen, G.; Gao, X.; Tian, Z.; Chen, H.; Guo, J.; Zhou, Z. OcomCSP12, a Chemosensory Protein Expressed Specifically by Ovary, Mediates Reproduction in Ophraella communa (Coleoptera: Chrysomelidae). Front. Physiol. 2019, 10, 1290. [Google Scholar] [CrossRef]
- Liu, G.; Yue, S.; Rajashekar, B.; Picimbon, J.-F. Expression of chemosensory protein (CSP) structures in Pediculus humanis corporis and Acinetobacter A. baumannii. SOJ Microbiol. Infect. Dis. 2019, 7, 1–17. [Google Scholar]
- Xuan, N.; Guo, X.; Xie, H.Y.; Lou, Q.N.; Lu, X.B.; Liu, G.X.; Picimbon, J.F. Increased expression of CSP and CYP genes in adult silkworm females exposed to avermectins. Insect. Sci. 2015, 22, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, K.; Utoguchi, A.; Yamada, A.; Yoshikawa, H. Identification and genomic structure of chemosensory proteins (CSP) and odorant binding proteins (OBP) genes expressed in foreleg tarsi of the swallowtail butterfly Papilio xuthus. Insect. Biochem. Mol. Biol. 2008, 38, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Arnaud, P.; Offmann, B.; Picimbon, J.F. Genotyping and Bio-Sensing Chemosensory Proteins in Insects. Sensors (Basel) 2017, 17, 1801. [Google Scholar] [CrossRef]
- Mei, T.; Fu, W.B.; Li, B.; He, Z.B.; Chen, B. Comparative genomics of chemosensory protein genes (CSPs) in twenty-two mosquito species (Diptera: Culicidae): Identification, characterization, and evolution. PLoS ONE 2018, 13, e0190412. [Google Scholar] [CrossRef]
- Liu, G.; Ma, H.; Xie, H.; Xuan, N.; Guo, X.; Fan, Z.; Rajashekar, B.; Arnaud, P.; Offmann, B.; Picimbon, J.F. Biotype Characterization, Developmental Profiling, Insecticide Response and Binding Property of Bemisia tabaci Chemosensory Proteins: Role of CSP in Insect Defense. PLoS ONE 2016, 11, e0154706. [Google Scholar] [CrossRef]
- Ozaki, M.; Wada-Katsumata, A.; Fujikawa, K.; Iwasaki, M.; Yokohari, F.; Satoji, Y.; Nisimura, T.; Yamaoka, R. Ant nestmate and non-nestmate discrimination by a chemosensory sensillum. Science 2005, 309, 311–314. [Google Scholar] [CrossRef]
- Niimura, Y. Evolutionary dynamics of olfactory receptor genes in chordates: Interaction between environments and genomic contents. Hum. Genomics 2009, 4, 107–118. [Google Scholar] [CrossRef]
- Montell, C. Gustatory receptors: Not just for good taste. Curr. Biol. 2013, 23, R929–R932. [Google Scholar] [CrossRef][Green Version]
- Montell, C. A taste of the Drosophila gustatory receptors. Curr. Opin. Neurobiol. 2009, 19, 345–353. [Google Scholar] [CrossRef]
- Ni, L.; Bronk, P.; Chang, E.C.; Lowell, A.M.; Flam, J.O.; Panzano, V.C.; Theobald, D.L.; Griffith, L.C.; Garrity, P.A. A gustatory receptor paralogue controls rapid warmth avoidance in Drosophila. Nature 2013, 500, 580–584. [Google Scholar] [CrossRef]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Rytz, R.; Croset, V.; Benton, R. Ionotropic receptors (IRs): Chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect Biochem. Mol. Biol. 2013, 43, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Vogt, R.G.; Miller, N.E.; Litvack, R.; Fandino, R.A.; Sparks, J.; Staples, J.; Friedman, R.; Dickens, J.C. The insect SNMP gene family. Insect. Biochem. Mol. Biol. 2009, 39, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.E.; Krieger, J.; Vogt, R.G. Antennal SNMPs (sensory neuron membrane proteins) of Lepidoptera define a unique family of invertebrate CD36-like proteins. J. Neurobiol. 2001, 49, 47–61. [Google Scholar] [CrossRef] [PubMed]
- Forstner, M.; Gohl, T.; Gondesen, I.; Raming, K.; Breer, H.; Krieger, J. Differential expression of SNMP-1 and SNMP-2 proteins in pheromone-sensitive hairs of moths. Chem. Senses 2008, 33, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.Q.; Nie, X.P.; Zhang, Z.; Zeng, F.F.; Lv, L.; Zhang, S.; Wang, M.Q. De novo analysis of the oriental armyworm Mythimna separata antennal transcriptome and expression patterns of odorant-binding proteins. Comp. Biochem. Physiol Part. D Genom. Proteom. 2017, 22, 120–130. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| G1 | G2 | G3 | M1 | M2 | M3 | C1 | C2 | C3 | |
|---|---|---|---|---|---|---|---|---|---|
| Clean reads | 67,492,576 | 74,983,476 | 70,212,518 | 78,796,858 | 61,848,876 | 72,961,796 | 71,005,592 | 54,703,286 | 70,316,132 |
| Assembled genes | 34,302 | 26,032 | 25,398 | 24,201 | 33,375 | 33,081 | 33,308 | 24,772 | 24,707 |
| Assembled transcripts | 49,914 | 40,912 | 39,528 | 38,089 | 50,517 | 51,704 | 50,530 | 38,856 | 39,637 |
| Unigenes | 162,621 | ||||||||
| Mean Length (bp) | 1425.14 | ||||||||
| N50 | 3572 | ||||||||
| GC (%) | 41.88 | ||||||||
| Total bases | 231,757,796 | ||||||||
| Expressed transcripts | 10,736 | 10,320 | 10,645 | 9860 | 10,863 | 11,695 | 16,827 | 11,765 | 14,728 |
| Type | Transcripts | Sub-Family | Intact ORF | SignalP | TMHMM |
|---|---|---|---|---|---|
| OBP | 43 | 21 OBP-56, 7 OBP-69, 4 OBP-72, 11 OBP-83 | 35 | 28 | 15 |
| CSP | 69 | 5 CSP-1, 3 CSP-3, 2 CSP-4, 5 CSP-5, 10 CSP-6, 3 CSP-7, 4 CSP-8, 37 CSP * | 63 | 50 | 13 |
| OR | 689 | 45 OR-1, 6 OR-1F12, 21 OR-10, 115 OR-13, 42 OR-2, 46 OR-22, 2 OR-23, 9 OR-24, 2 OR-245, 1 OR-260, 2 OR-266, 1 OR-277, 15 OR-30, 6 OR-33, 31 OR-4, 5 OR-42, 14 OR-43, 2 OR-45, 24 OR-46, 47 OR-47, 26 OR-49, 2 OR-59, 1 OR-5, 1 OR-63, 48 OR-67, 1 OR-69, 6 OR-71, 13 OR-7, 1 OR-81, 62 OR-82, 37 OR-85, 11 OR-92, 11 OR-94, 5 OR-98, 4 OR-9, 9 OR-co, 1 OR-142, 14 OR * | 347 | 25 | 276 |
| IR | 26 | 3 IR-21, 18 IR-25, 4 IR-68, 2 IR-93 | 18 | 4 | 12 |
| GR | 60 | 6 GR-2, 5 GR-107, 3 GR-15, 1 GR-23, 19 GR-28, 4 GR-2, 8 GR-43, 2 GR-64, 3 GR-66, 8 GR * | 26 | 1 | 25 |
| SNMP | 14 | 11 SNMP-1, 3 SNMP-3 | 10 | 1 | 9 |
| Others a | 54 | 16 CR, 38 GOBP | 10 | 12 | 22 |
| TranscriptID | G a | M a | C a | Protein/Gene | Description |
|---|---|---|---|---|---|
| OBP | |||||
| TRINITY_DN22_c0_g3_i1 | 41,518.89 | 55,018.99 | 39,228.41 | OB56D | General odorant-binding protein 56d |
| cluster_contig4670 | 20,459.11 | 27,957.59 | 83,228.40 | OB56H | General odorant-binding protein 56h |
| cluster_contig3738 | 15,552.04 | 13,752.58 | 20,997.13 | OB69A | General odorant-binding protein 69a |
| TRINITY_DN3641_c0_g2_i1 | 5,010.50 | 13,074.19 | 4,838.31 | OB69A | General odorant-binding protein 69a |
| TRINITY_DN23528_c0_g1_i1 | 3853.93 | 5367.95 | 6228.73 | OB56D | General odorant-binding protein 56d |
| CSP | |||||
| TRINITY_DN1018_c0_g1_i5 | 11,065.74 | 15,620.31 | 10,030.62 | CSP | chemosensory protein |
| TRINITY_DN661_c1_g1_i1 | 9447.67 | 10,194.33 | 7724.67 | THK33221.1 | chemosensory protein 4 |
| TRINITY_DN4258_c0_g1_i1 | 4538.67 | 4738.33 | 5976.67 | AZQ24964.1 | chemosensory protein, partial |
| TRINITY_DN1848_c0_g1_i6 | 5121.01 | 3247.27 | 4919.50 | CSP | chemosensory protein |
| cluster_contig14748 | 3679.27 | 3944.22 | 2395.00 | THK33222.1 | chemosensory protein 5 |
| OR | |||||
| cluster_contig6330 | 41,364.85 | 68,243.05 | 24,104.78 | LOC107043577 | Odorant receptor |
| TRINITY_DN1357_c0_g1_i5 | 18,042.97 | 23,565.93 | 11,831.46 | OR43A | Odorant receptor 43a |
| TRINITY_DN1357_c0_g1_i6 | 7765.79 | 27,596.19 | 13,421.25 | OR43A | Odorant receptor 43a |
| cluster_contig12468 | 16,698.80 | 23,528.66 | 8172.54 | LOC107043576 | Odorant receptor |
| cluster_contig9759 | 2835.24 | 2248.79 | 3479.03 | OR43A | Odorant receptor 43a |
| GR | |||||
| TRINITY_DN9119_c0_g1_i2 | 455.68 | 489.30 | 545.92 | GR107 | gustatory receptor Gr107 |
| TRINITY_DN3525_c0_g1_i10 | 288.94 | 333.98 | 230.17 | GR43A | gustatory receptor for sugar taste 43a-like |
| cluster_contig36 | 290.10 | 43.82 | 30.95 | GR43a | gustatory receptor for sugar taste 43a-like |
| TRINITY_DN14061_c0_g1_i3 | 91.79 | 116.05 | 78.74 | GR43a | gustatory receptor for sugar taste 43a-like |
| TRINITY_DN4338_c0_g1_i4 | 90.22 | 77.22 | 64.01 | GR107 | gustatory receptor Gr107 |
| IR | |||||
| TRINITY_DN11328_c0_g1_i1 | 41.09 | 39.27 | 45.12 | IR25A | Ionotropic receptor 25a |
| cluster_contig8505 | 39.77 | 34.65 | 39.58 | IR25A | Ionotropic receptor 25a |
| TRINITY_DN4424_c0_g1_i2 | 16.26 | 12.92 | 47.84 | IR25A | Ionotropic receptor 25a |
| cluster_contig11752 | 19.21 | 25.57 | 18.80 | IR25A | Ionotropic receptor 25a |
| TRINITY_DN13912_c1_g2_i1 | 20.88 | 10.71 | 20.37 | IR25A | Ionotropic receptor 25a |
| SNMP | |||||
| TRINITY_DN1934_c0_g1_i1 | 3567.61 | 3632.31 | 5117.35 | SNMP1 | Sensory neuron membrane protein 1 |
| TRINITY_DN4669_c1_g1_i1 | 176.96 | 204.92 | 206.30 | SNMP1 | Sensory neuron membrane protein 1 |
| TRINITY_DN1867_c0_g2_i1 | 86.72 | 83.98 | 50.41 | XP_015114627.1 | Sensory neuron membrane protein 1-like |
| cluster_contig4264 | 70.20 | 73.23 | 46.97 | SNMP1 | Sensory neuron membrane protein 1 |
| cluster_contig237 | 44.54 | 90.10 | 34.07 | SNMP1 | Sensory neuron membrane protein 1 |
| TRINITY_DN1020_c0_g1_i3 | TRINITY_DN1284_c0_g1_i11 | TRINITY_DN500_c0_g1_i2 | ||
|---|---|---|---|---|
| Primers | Forward | CAACTTCAAGAACAATCCGACAAC | ACTTATAGACGCATGCCAAGACC | GACGTCGCTATGAACGCTTG |
| Reverse | CCACAGCCAGAGACACAGC | GGGCTGGAGAACGGGGATG | GATTCTGATTTCCAGTACGAATACG | |
| C_vs_G | Log2 FC | 10.00 | 9.84 | 0 |
| p-value | 4.12 × 10−28 | 6.02 × 10−27 | 1 | |
| RNE | 1.13 | 2.32 | 0.42 | |
| C_vs_M | Log2 FC | −9.47 | 1.43 | 6.43 |
| p-value | 2.28 × 10−23 | 0.0013 | 1.98 × 10−33 | |
| RNE | 1.68 | 0.66 | 0.00 | |
| M_vs_G | Log2 FC | −0.53 | 8.32 | 6.66 |
| p-value | 0.3856 | 1.13 × 10−15 | 7.18 × 10−34 | |
| RNE | 0.67 | 3.52 | 79.36 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, L.; Liu, J.; Liu, L.; Yu, Y.; Zhao, H.; Lu, W. De Novo Transcriptome Identifies Olfactory Genes in Diachasmimorpha longicaudata (Ashmead). Genes 2020, 11, 144. https://doi.org/10.3390/genes11020144
Tang L, Liu J, Liu L, Yu Y, Zhao H, Lu W. De Novo Transcriptome Identifies Olfactory Genes in Diachasmimorpha longicaudata (Ashmead). Genes. 2020; 11(2):144. https://doi.org/10.3390/genes11020144
Chicago/Turabian StyleTang, Liangde, Jimin Liu, Lihui Liu, Yonghao Yu, Haiyan Zhao, and Wen Lu. 2020. "De Novo Transcriptome Identifies Olfactory Genes in Diachasmimorpha longicaudata (Ashmead)" Genes 11, no. 2: 144. https://doi.org/10.3390/genes11020144
APA StyleTang, L., Liu, J., Liu, L., Yu, Y., Zhao, H., & Lu, W. (2020). De Novo Transcriptome Identifies Olfactory Genes in Diachasmimorpha longicaudata (Ashmead). Genes, 11(2), 144. https://doi.org/10.3390/genes11020144