A Comprehensive Transcriptional Profiling of Pepper Responses to Root-Knot Nematode

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant and Nematode Cultures
2.2. Experimental Design and Sample Collection
2.3. Sequencing Processing and Gene Expression Analysis
2.4. Availability of Data
3. Results
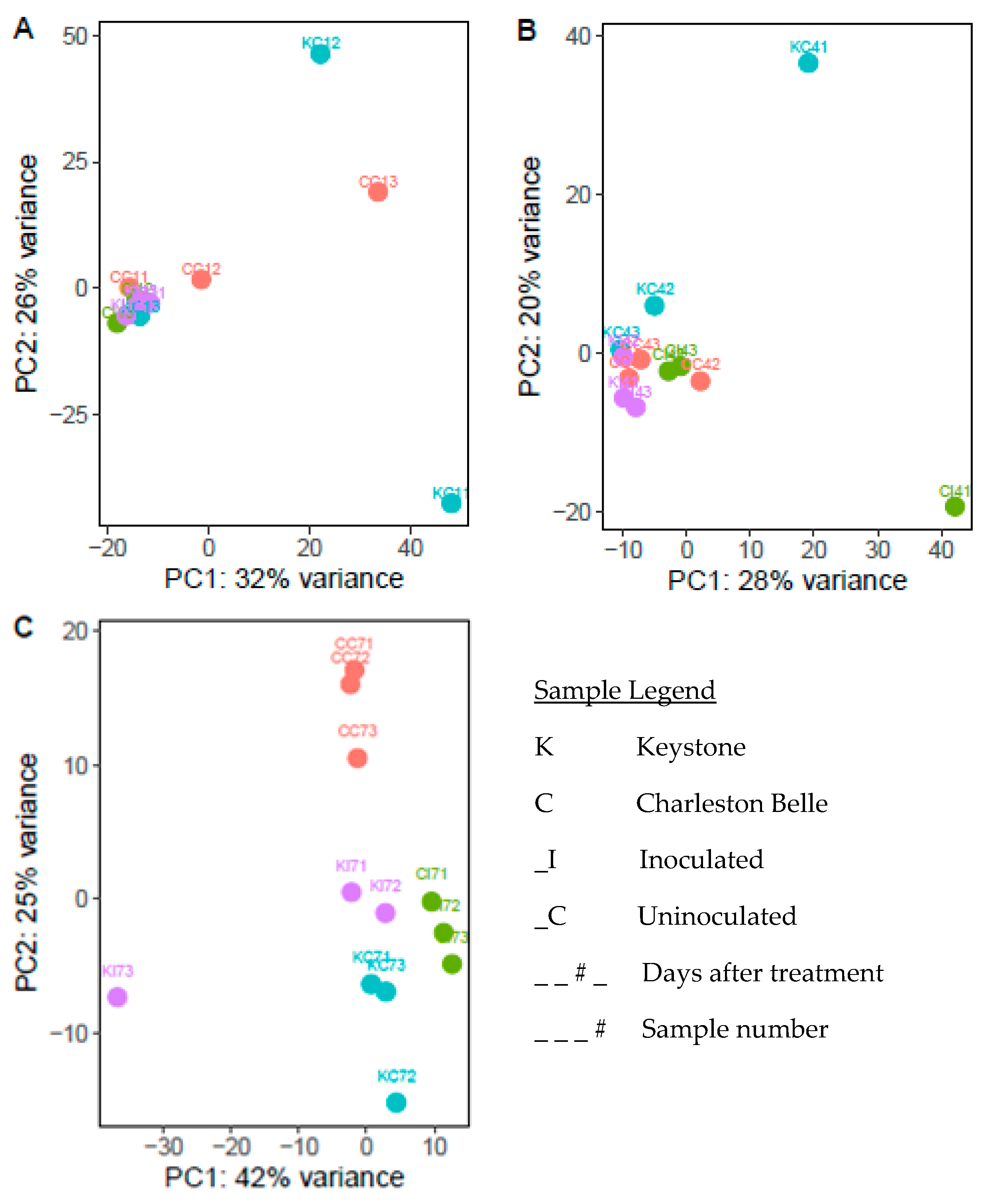
3.1. Sequencing Data Quality Control
3.2. Sequencing and Gene Expression Summary
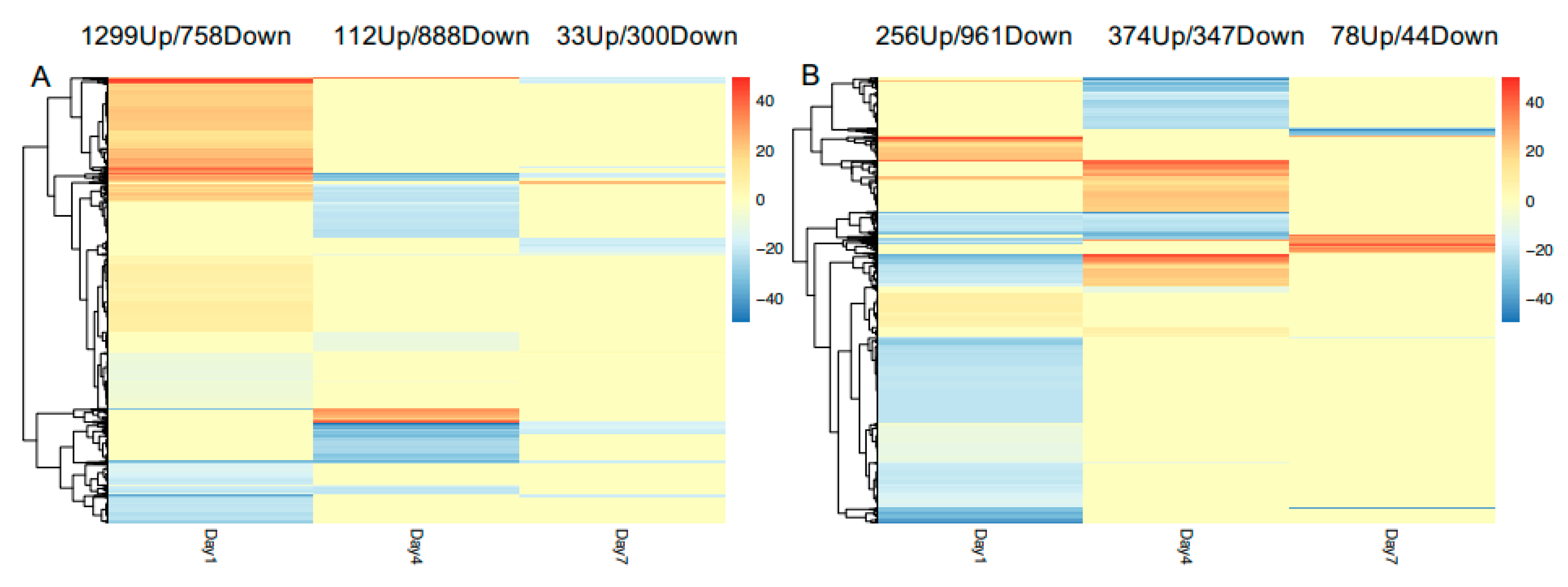
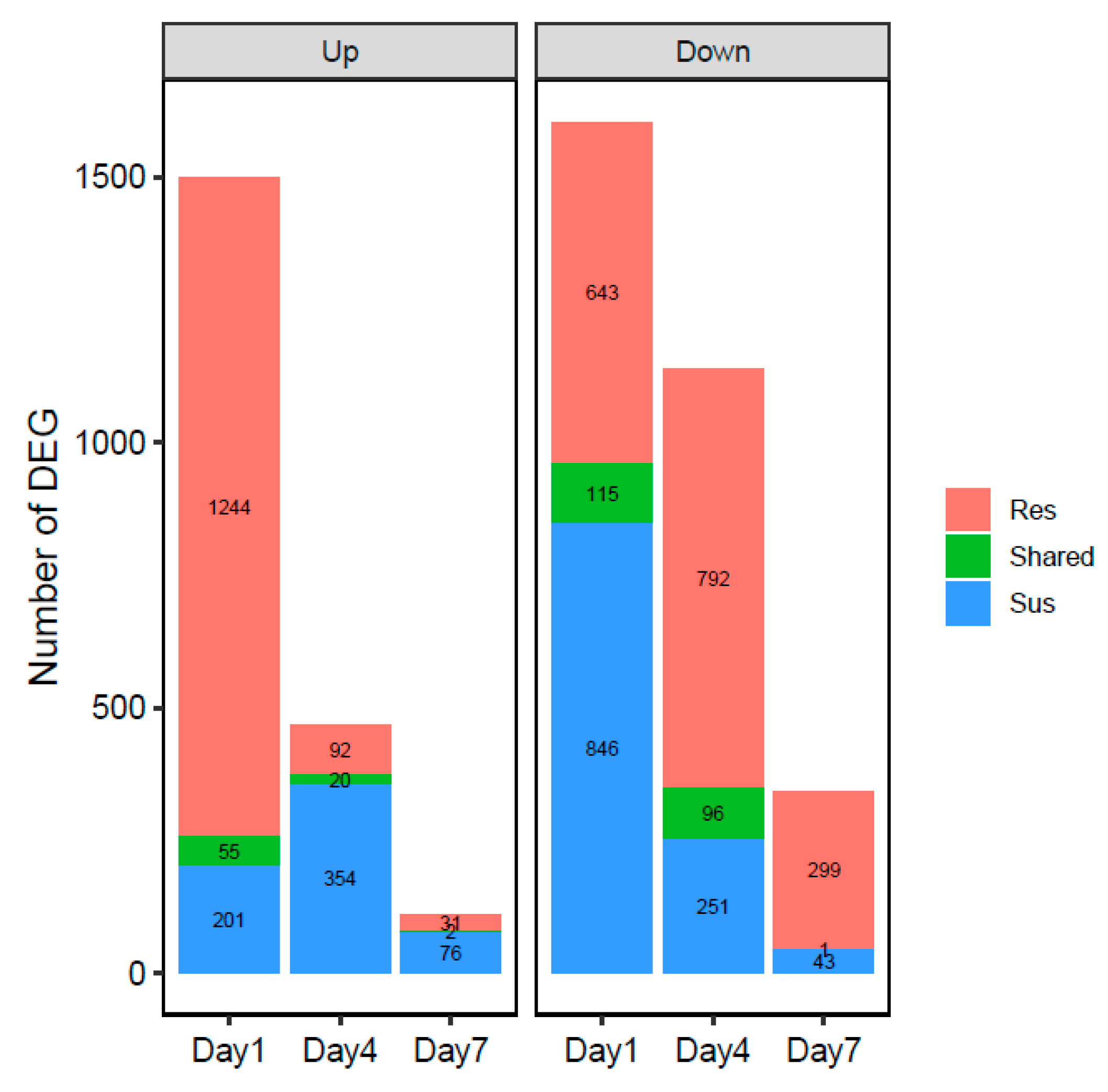
3.3. General Pepper Responses to Nematode Infection
3.4. Unique Cultivar Responses to Nematode Infection
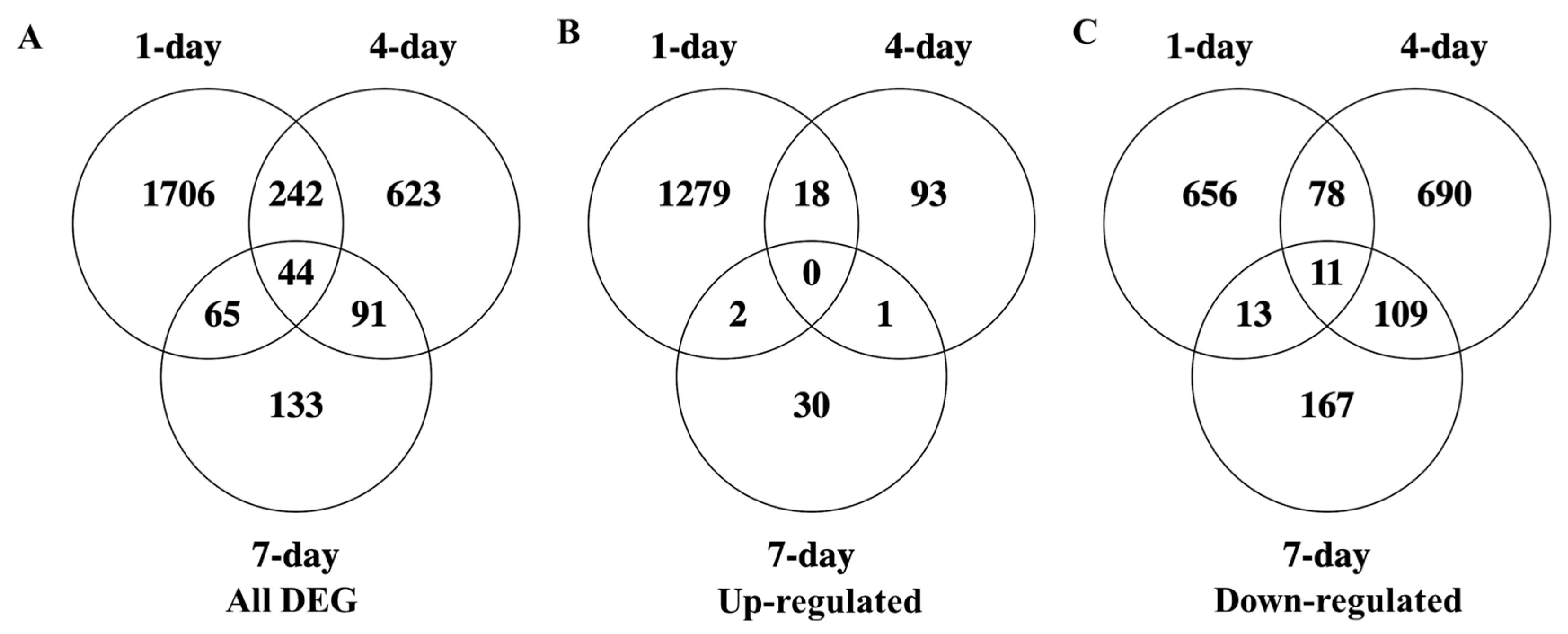
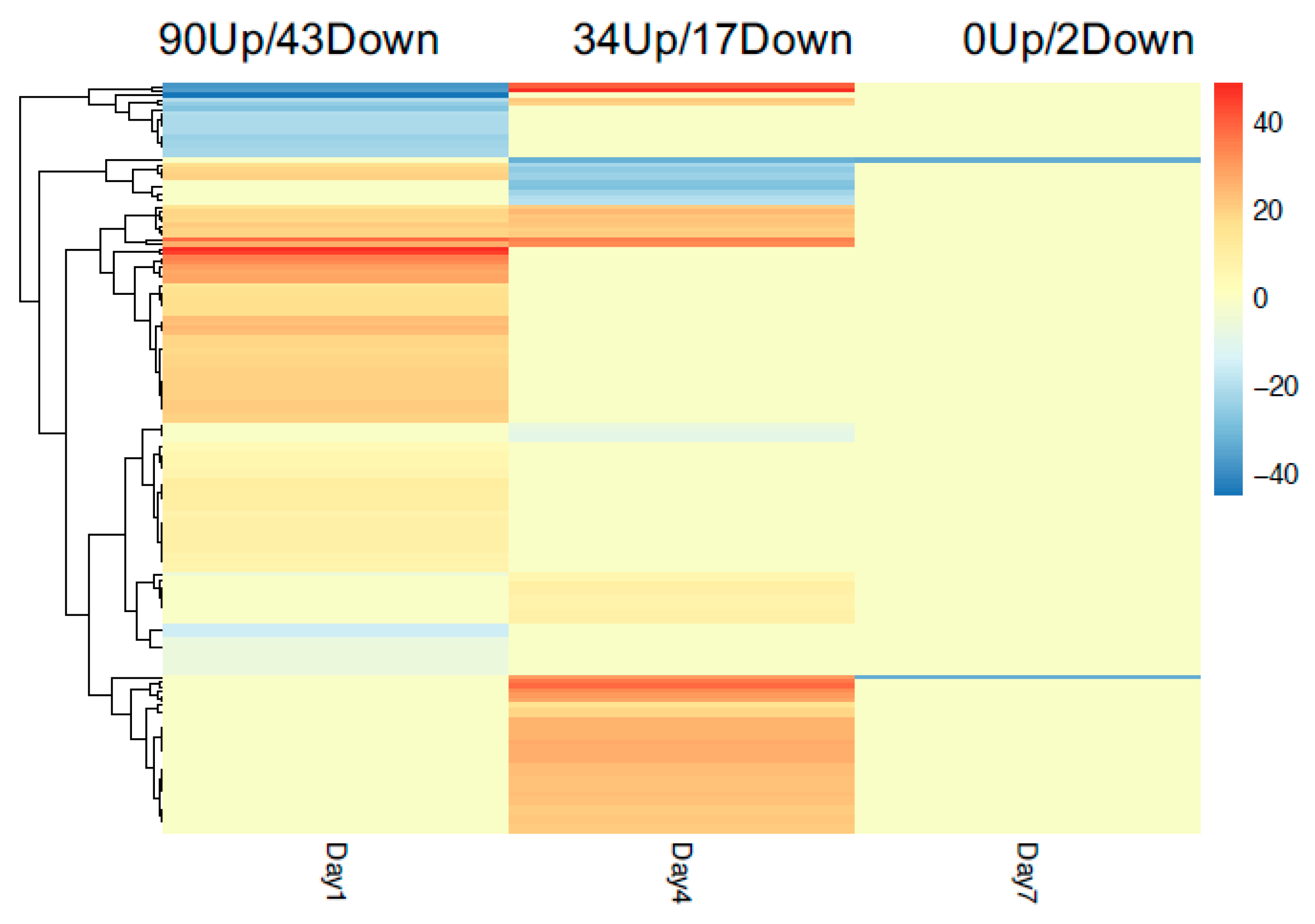
3.5. Temporal Variations in Cultivar Responses to Nematode Infection
3.6. Chromosome P9 and Putative RKN Resistance Loci
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chitwood, D.J. Research on plant-parasitic nematode biology conducted by the United States Department of Agriculture—Agricultural Research Service. Pest Manag. Sci. 2003, 59, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Thies, J.A.; Fery, R.L. Characterization of Resistance Conferred by the N gene to Meloidogyne arenaria Races 1 and 2, M. hapla, and M. javanica in Two Sets of Isogenic Lines of Capsicum annuum L. J. Am. Soc. Hortic. Sci. 2000, 125, 71–75. [Google Scholar] [CrossRef]
- Taylor, A.L.; Sasser, J.N. Biology, Identification and Control of Root-Knot Nematodes (Meloidogyne species); Department of Plant Pathology, North Carolina State University, United States Agency for International Development: Raleigh, NC, USA, 1978. [Google Scholar]
- Martin, J.A.; Crawford, J.H. Carolina hot pepper. In 1958: South Carolina Agricultural Experiment Station; Clemson Agricultural College: Clemson, SC, USA, 1958. [Google Scholar]
- Hare, W. Inheritance of resistance to root-knot nematodes in pepper. Phytopathology 1957, 47, 455–459. [Google Scholar]
- Hare, W. New pimiento is resistant to nematodes. Miss. Farm Res 1966, 29, 1–8. [Google Scholar]
- Fery, R.L.; Dukes, P.D., Sr.; Thies, J.A. ‘Carolina Wonder’ and ‘Charleston Belle’: Southern root-knot nematode-resistant bell peppers. HortScience 1998. [Google Scholar] [CrossRef]
- Djian-Caporalino, C.; Fazari, A.; Arguel, M.J.; Vernie, T.; VandeCasteele, C.; Faure, I.; Brunond, G.; Pijarowski, L.; Palloix, A.; Lefebvre, V.; et al. Root-knot nematode (Meloidogyne spp.) Me resistance genes in pepper (Capsicum annuum L.) are clustered on the P9 chromosome. Theor. Appl. Genet. 2007, 114, 473–486. [Google Scholar] [CrossRef]
- Fazari, A.; Palloix, A.; Wang, L.; Yan Hua, M.; Sage-Palloix, A.M.; Zhang, B.X.; Djian-Caporalino, C. The root-knot nematode resistance N-gene co-localizes in the Me-genes cluster on the pepper (Capsicum annuum L.) P9 chromosome. Plant Breed. 2012, 131, 665–673. [Google Scholar] [CrossRef]
- Djian-Caporalino, C.; Pijarowski, L.; Januel, A.; Lefebvre, V.; Daubeze, A.; Palloix, A.; Dalmasso, A.; Abad, P. Spectrum of resistance to root-knot nematodes and inheritance of heat-stable resistance in in pepper (Capsicum annuum L.). Theor. Appl. Genet. 1999, 99, 496–502. [Google Scholar] [CrossRef]
- Grube, R.C.; Radwanski, E.R.; Jahn, M. Comparative genetics of disease resistance within the Solanaceae. Genetics 2000, 155, 873–887. [Google Scholar]
- Mazourek, M.; Cirulli, E.T.; Collier, S.M.; Landry, L.G.; Kang, B.C.; Quirin, E.A.; Bradeen, J.M.; Moffett, P.; Jahn, M.M. The fractionated orthology of Bs2 and Rx/Gpa2 supports shared synteny of disease resistance in the Solanaceae. Genetics 2009, 182, 1351–1364. [Google Scholar] [CrossRef][Green Version]
- Xiao, G.; Ang, X.H.; Yu, Y.A.N.G.; Mao, Z.C.; Feng, L.I.U.; Xie, B.Y.; Li, G.C. Bacterial artificial chromosome library construction of root-knot nematode resistant pepper genotype HDA149 and identification of clones linked to Me3 resistant locus. J. Integr. Agric. 2017, 16, 57–64. [Google Scholar]
- El-Sappah, A.H.; Islam, M.M.; El-Awady, H.H.; Yan, S.; Qi, S.; Liu, J.; Cheng, G.T.; Liang, Y. Tomato Natural Resistance Genes in Controlling the Root-Knot Nematode. Genes 2019, 10, 925. [Google Scholar] [CrossRef] [PubMed]
- Milligan, S.B.; Bodeau, J.; Yaghoobi, J.; Kaloshian, I.; Zabel, P.; Williamson, V.M. The root knot nematode resistance gene Mi from tomato is a member of the leucine zipper, nucleotide binding, leucine-rich repeat family of plant genes. Plant Cell 1998, 10, 1307–1319. [Google Scholar] [CrossRef] [PubMed]
- Nombela, G.; Williamson, V.M.; Muñiz, M. The root-knot nematode resistance gene Mi-1.2 of tomato is responsible for resistance against the whitefly Bemisia tabaci. Mol. Plant-Microbe Interact. 2003, 16, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.-F.; Bhakta, A.V.; Truesdell, G.M.; Pudlo, W.M.; Williamson, V.M. Evidence for a role of the N terminus and leucine-rich repeat region of the Mi gene product in regulation of localized cell death. Plant Cell 2000, 12, 1319–1329. [Google Scholar] [CrossRef] [PubMed]
- Gleason, C.A.; Liu, Q.L.; Williamson, V.M. Silencing a candidate nematode effector gene corresponding to the tomato resistance gene Mi-1 leads to acquisition of virulence. Mol. Plant-Microbe Interact. 2008, 21, 576–585. [Google Scholar] [CrossRef]
- Rossi, M.; Goggin, F.L.; Milligan, S.B.; Kaloshian, I.; Ullman, D.E.; Williamson, V.M. The nematode resistance gene Mi of tomato confers resistance against the potato aphid. Proc. Natl. Acad. Sci. USA 1998, 95, 9750–9754. [Google Scholar] [CrossRef]
- de Ilarduya, O.M.; Xie, Q.G.; Kaloshian, I. Aphid-induced defense responses in Mi-1-mediated compatible and incompatible tomato interactions. Mol. Plant-Microbe Interact. 2003, 16, 699–708. [Google Scholar] [CrossRef]
- Schneider, D.S. Plant immunity and film noir: What gumshoe detectives can teach us about plant-pathogen interactions. Cell 2002, 109, 537–540. [Google Scholar] [CrossRef][Green Version]
- Jarquin-Barberena, H.; Castagnone-Sereno, P.; Bongiovanni, M.; De Jong, A. Acquired virulence in the plant parasitic nematode Meloidogyne incognita. I, Biological analysis of the phenomenon. Rev. Nematol. 1991, 14, 299–303. [Google Scholar]
- Verdejo-Lucas, S.; Cortada, L.; Sorribas, F.J.; Ornat, C. Selection of virulent populations of Meloidogyne javanica by repeated cultivation of Mi resistance gene tomato rootstocks under field conditions. Plant Pathol. 2009, 58, 990–998. [Google Scholar] [CrossRef]
- Molinari, S.; Miacola, C. Interactions between resistant tomato cys and Meloidogyne spp. in vitro. Nematol. Mediterr. 1997, 25, 63–72. [Google Scholar]
- Roberts, P.; Thomason, J. Variability in reproduction of isolates of Meloidogyne incognita and M. javanica on resistant tomato genotypes. Plant Disease 1986, 70, 547–551. [Google Scholar] [CrossRef]
- Ogallo, J.; McClure, M. Systemic acquired resistance and susceptibility to root-knot nematodes in tomato. Phytopathology 1996, 5, 498–501. [Google Scholar] [CrossRef]
- Blanc-Mathieu, R.; Perfus-Barbeoch, L.; Aury, J.M.; Da Rocha, M.; Gouzy, J.; Sallet, E.; Martin-Jimenez, C.; Bailly-Bechet, M.; Castagnone-Sereno, P.; Flot, J.F.; et al. Hybridization and polyploidy enable genomic plasticity without sex in the most devastating plant-parasitic nematodes. PLoS Genet. 2017, 13, e1006777. [Google Scholar] [CrossRef] [PubMed]
- Changkwian, A.; Venkatesh, J.; Lee, J.H.; Han, J.W.; Kwon, J.K.; Siddique, M.I.; Abate, S.; Choi, G.J.; Kim, E.; Seo, Y.; et al. Physical Localization of the Root-Knot Nematode (Meloidogyne incognita) Resistance Locus Me7 in Pepper (Capsicum annuum). Front. Plant Sci. 2019, 10, 886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kjemtrup-Lovelace, S.; Li, C.; Luo, Y.; Chen, L.P.; Song, B.H. Comparative RNA-seq analysis uncovers a complex regulatory network for soybean cyst nematode resistance in wild soybean (Glycine soja). Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Vieira, P.; Gleason, C. Plant-parasitic nematode effectors—Insights into their diversity and new tools for their identification. Curr. Opin. Plant Biol. 2019, 50, 37–43. [Google Scholar] [CrossRef]
- Bali, S.; Vining, K.; Gleason, C.; Majtahedi, H.; Brown, C.R.; Sathuvalli, V. Transcriptome profiling of resistance response to Meloidogyne chitwoodi introgressed from wild species Solanum bulbocastanum into cultivated potato. BMC Genom. 2019, 20, 907. [Google Scholar] [CrossRef]
- Petre, B.; Lorrain, C.; Stukenbrock, E.H.; Duplessis, S. Host-specialized transcriptome of plant-associated organisms. Curr. Opin. Plant Biol. 2020, 56, 81–88. [Google Scholar] [CrossRef]
- Maquilan, M.A.D.; Padilla, D.C.; Dickson, D.W.; Rathinasabapathi, B. Improved Resistance to Root-knot Nematode Species in an Advanced Inbred Line of Specialty Pepper (Capsicum annuum). HortScience 2020, 1, 1–6. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A quality control tool for high throughput sequence data. Babraham Inst. 2010. Available online: https://www.scienceopen.com/document?vid=a2919c6f-fc3a-420e-8b34-e7024127c2ef (accessed on 14 October 2020).
- Kim, S.; Park, M.; Yeom, S.I.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Oliveros, J.C. VENNY. An Interactive Tool for Comparing Lists with Venn’s Diagrams. 2007. Available online: http//bioinfogp.cnb. csic.es/tools/venny/index.html (accessed on 14 October 2020).
- Guo, A.-Y.; Chen, X.; Gao, G.; Zhang, H.; Zhu, Q.H.; Liu, X.C.; Zhong, Y.F.; Gu, X.; He, K.; Luo, J. PlantTFDB: A comprehensive plant transcription factor database. Nucleic Acids Res. 2007, 36 (Suppl. 1), D966–D969. [Google Scholar] [CrossRef]
- Berardini, T.Z.; Reiser, L.; Li, D.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The Arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef]
- UniProt Consortium, UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [CrossRef]
- Alexa, A.; Rahnenfuhrer, J. topGO: Enrichment analysis for gene ontology. R Package Version 2010, 2, 2010. [Google Scholar]
- Kushalappa, A.C.; Yogendra, K.N.; Karre, S. Plant innate immune response: Qualitative and quantitative resistance. Crit. Rev. Plant Sci. 2016, 35, 38–55. [Google Scholar] [CrossRef]
- Celik, I.; Sogut, M.A.; Ozkaynak, E.; Doganlar, S.; Frary, A. Physical mapping of NBS-coding resistance genes to the Me-gene cluster on chromosome P9 reveals markers tightly linked to the N gene for root-knot nematode resistance in pepper. Mol. Breed. 2016, 36, 137. [Google Scholar] [CrossRef]
- Thies, J.A.; Fery, R.L. Modified expression of the N gene for southern root-knot nematode resistance in pepper at high soil temperatures. J. Am. Soc. Horticul. Sci. 1998, 123, 1012–1015. [Google Scholar] [CrossRef]
- Kaloshian, I.; Williamson, V.; Miyao, G.; Lawn, D.; Westerdahl, B. “Resistance-breaking” nematodes identified in California tomatoes. Calif. Agric. 1996, 50, 18–19. [Google Scholar] [CrossRef]
- Castagnone-Sereno, P.; Wajnberg, E.; Bongiovanni, M.; Leroy, F.; Dalmasso, A. Genetic variation in Meloidogyne incognita virulence against the tomato Mi resistance gene: Evidence from isofemale line selection studies. Theor. Appl. Genet. 1994, 88, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Tzortzakakis, E.A.; Adam, M.A.; Blok, V.C.; Paraskevopoulos, C.; Bourtzis, K. Occurrence of resistance-breaking populations of root-knot nematodes on tomato in Greece. Eur. J. Plant Pathol. 2005, 113, 101–105. [Google Scholar] [CrossRef]
- Balestrini, R.; Rosso, L.C.; Veronico, P.; Melillo, M.T.; De Luca, F.; Fanelli, E.; Colagiero, M.; Salvioli, A.; Ciancio, A.; Pentimone, I. Transcriptomic responses to water deficit and nematode infection in mycorrhizal tomato roots. Front. Microbiol. 2019, 10, 1807. [Google Scholar] [CrossRef]
- Kaloshian, I.; Teixeira, M. Advances in Plant—Nematode Interactions with Emphasis on the Notorious Nematode Genus Meloidogyne. Phytopathology 2019, 109, 1988–1996. [Google Scholar] [CrossRef]
- Bleve-Zacheo, T.; Bongiovanni, M.; Melillo, M.T.; Castagnone-Sereno, P. The pepper resistance genes Me1 and Me3 induce differential penetration rates and temporal sequences of root cell ultrastructural changes upon nematode infection. Plant Sci. 1998, 133, 79–90. [Google Scholar] [CrossRef]
- Peng, H.-C.; Kaloshian, I. The tomato leucine-rich repeat receptor-like kinases SlSERK3A and SlSERK3B have overlapping functions in bacterial and nematode innate immunity. PLoS ONE 2014, 9, e93302. [Google Scholar] [CrossRef]
- Teixeira, M.A.; Wei, L.; Kaloshian, I. Root-knot nematodes induce pattern-triggered immunity in Arabidopsis thaliana roots. New Phytol. 2016, 211, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Mendy, B.; Wang’ombe, M.W.; Radakovic, Z.S.; Holbein, J.; Ilyas, M.; Chopra, D.; Holton, N.; Zipfel, C.; Grundler, F.M.; Siddique, S. Arabidopsis leucine-rich repeat receptor–like kinase NILR1 is required for induction of innate immunity to parasitic nematodes. PLoS Pathog. 2017, 13, e1006284. [Google Scholar] [CrossRef] [PubMed]
- Siddique, S.; Sobczak, M.; Tenhaken, R.; Grundler, F.M.; Bohlmann, H. Cell wall ingrowths in nematode induced syncytia require UGD2 and UGD3. PLoS ONE 2012, 7, e41515. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Teng, Q.; Zhong, R.; Yuan, Y.; Haghighat, M.; Ye, Z.H. Three Arabidopsis DUF579 Domain-Containing GXM Proteins are Methyltransferases Catalyzing 4-O-Methylation of Glucuronic Acid on Xylan. Plant Cell Physiol. 2012, 53, 1934–1949. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, P.; Wang, Y.; Cheng, Q.; Lu, Q.; Liu, J.; Li, T.; Ai, Y.; Yang, W.; Sun, L.; et al. Genome-Wide Correlation of 36 Agronomic Traits in the 287 Pepper (Capsicum) Accessions Obtained from the SLAF-seq-Based GWAS. Int. J. Mol. Sci. 2019, 20, 5675. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.; Simons, G.; Jesse, T.; Wijbrandi, J.; Heinen, L.; Hogers, R.; Frijters, A.; Groenendijk, J.; Diergaarde, P.; Reijans, M.; et al. The tomato Mi-1 gene confers resistance to both root-knot nematodes and potato aphids. Nat. Biotechnol. 1998, 16, 1365–1369. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GeneID | Resistant 1 | Susceptible 1 | Time Point | Function |
|---|---|---|---|---|
| CA09g00350 | 19.97 | −20 | day 1 | Rac GTPase activating protein 1 |
| CA09g07060 | −13.44 | 23.41 | day 1 | Unknown protein |
| CA09g08070 | 17 | −22.02 | day 1 | subtilisin-like protease-like |
| CA09g08670 | 37.37 | −31.48 | day 1 | Acetyl-CoA carboxylase, putative |
| CA09g08790 | 20.61 | −20.69 | day 1 | Protein binding protein, putative |
| CA09g09700 | 20.99 | −21.9 | day 1 | Multicopper oxidase, putative |
| CA09g09910 | −16.65 | 19.09 | day 1 | Tetratricopeptide repeat-containing protein |
| CA09g11690 | 21.56 | −19.29 | day 1 | STY-L protein |
| CA09g11780 | 21.24 | −20.79 | day 1 | 3-ketoacyl-CoA reductase 1 |
| CA09g14450 | 20.12 | −20.88 | day 1 | WD repeat-containing protein 26-like isoform X1 |
| CA09g15640 | 16.25 | −13.7 | day 1 | ATP synthase subunit α |
| CA09g16660 | 15.2 | −20.24 | day 1 | Unknown protein |
| CA09g16770 | 7.72 | −9.87 | day 1 | Unknown protein |
| CA09g16890 | 20.97 | −19.79 | day 1 | Unknown protein |
| CA09g17010 | 48.07 | −39.53 | day 1 | BED finger-nbs-lrr resistance protein |
| CA09g04320 | 28.3 | −27.12 | day 4 | F-box/kelch-repeat protein At3g23880-like |
| CA09g05170 | −21.52 | 19.29 | day 4 | GRAS family transcription factor |
| CA09g12300 | −36.36 | 33.82 | day 4 | Purple acid phosphatase |
| CA09g15320 | −33.87 | 37.99 | day 4 | Putative serine/threonine protein kinase |
| CA09g16890 | −21.29 | 21.94 | day 4 | Unknown protein |
| CA09g18730 | −21.59 | 22.2 | day 4 | Senescence-associated protein |
| CA09g17960 | −18.13 | 33 | day 7 | glucuronoxylan 4-O-methyltransferase 2-like |
| Cultivar and Resistance Type | Time and Number of Genes | ||
|---|---|---|---|
| Resistant | day 1 | day 4 | day 7 |
| NBS-LRR resistance genes | 26 | 9 | 6 |
| Nematode resistance genes | 4 | 3 | 0 |
| Susceptible | day 1 | day 4 | day 7 |
| NBS-LRR resistance genes | 2 | 1 | 0 |
| Nematode resistance genes | 7 | 7 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, W.; Kingsbury, K.; Mishra, S.; DiGennaro, P. A Comprehensive Transcriptional Profiling of Pepper Responses to Root-Knot Nematode. Genes 2020, 11, 1507. https://doi.org/10.3390/genes11121507
Hu W, Kingsbury K, Mishra S, DiGennaro P. A Comprehensive Transcriptional Profiling of Pepper Responses to Root-Knot Nematode. Genes. 2020; 11(12):1507. https://doi.org/10.3390/genes11121507
Chicago/Turabian StyleHu, Weiming, Krista Kingsbury, Shova Mishra, and Peter DiGennaro. 2020. "A Comprehensive Transcriptional Profiling of Pepper Responses to Root-Knot Nematode" Genes 11, no. 12: 1507. https://doi.org/10.3390/genes11121507
APA StyleHu, W., Kingsbury, K., Mishra, S., & DiGennaro, P. (2020). A Comprehensive Transcriptional Profiling of Pepper Responses to Root-Knot Nematode. Genes, 11(12), 1507. https://doi.org/10.3390/genes11121507