MicroRNA-214 Inhibits Chicken Myoblasts Proliferation, Promotes Their Differentiation, and Targets the TRMT61A Gene

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals and RNA Extraction
2.3. cDNA Synthesis and Design of the Primers for Quantitative Real-Time PCR (qPCR)
2.4. Isolation and Culture of CPMs
2.5. CCK-8 Assay
2.6. Cell Cycle Analysis
2.7. EdU Assay
2.8. Construction and Transfection of Plasmid
2.9. Immunofluorescence
2.10. Western Blotting Assay
2.11. Statistical Analysis
3. Results
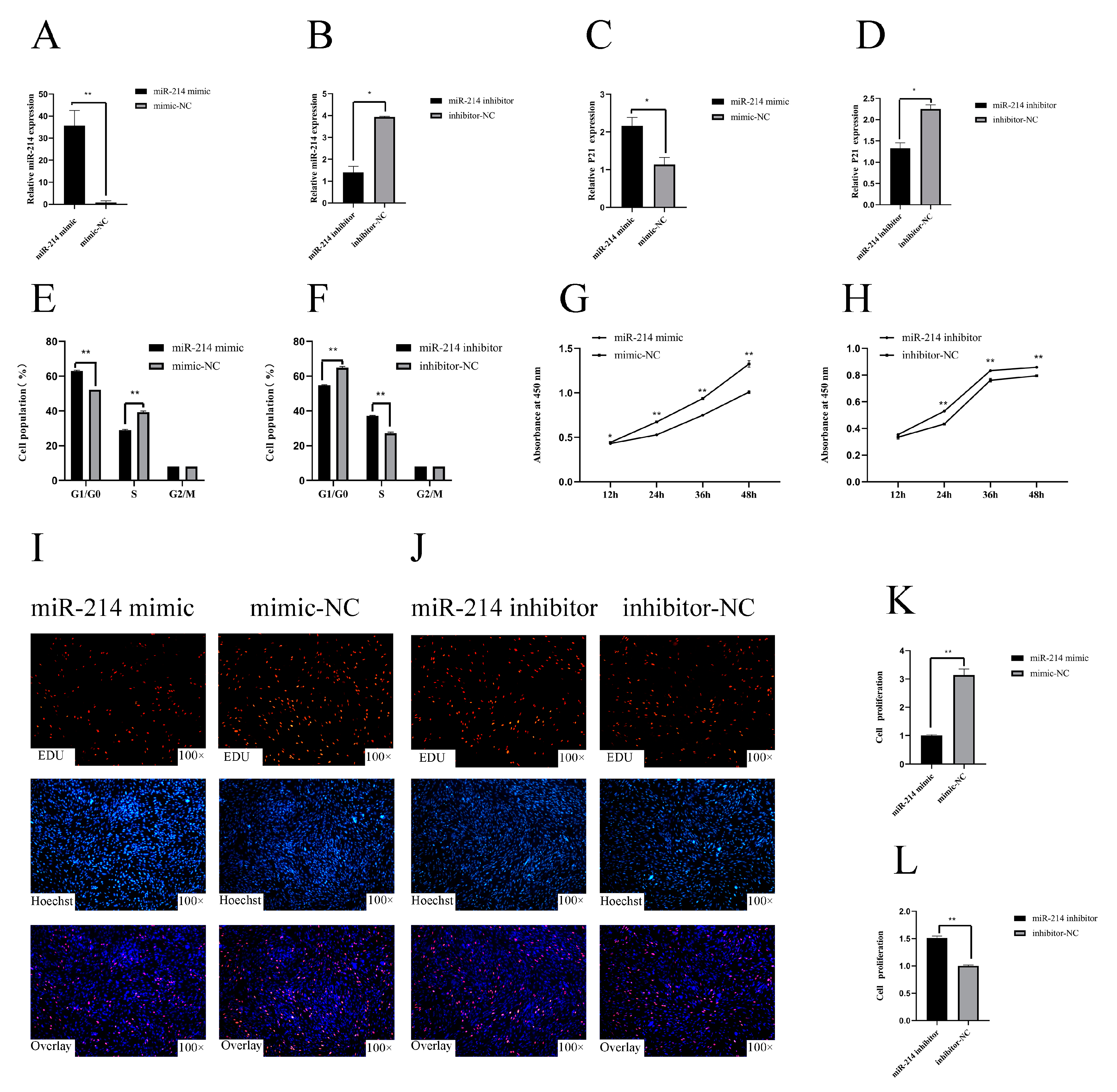
3.1. miR-214 Inhibits the Proliferation of Chicken Myoblasts
3.2. miR-214 Promotes the Differentiation of Chicken Myoblasts
3.3. miR-214 Targets TRMT61A and Negatively Regulates TRMT61A
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pavlath, G.K.; Horsley, V. Cell fusion in skeletal muscle—Central role of NFATC2 in regulating muscle cell size. Cell Cycle 2003, 2, 419–422. [Google Scholar] [CrossRef]
- Koomkrong, N.; Theerawatanasirikul, S.; Boonkaewwan, C.; Jaturasitha, S.; Kayan, A. Breed-Related Number and Size of Muscle Fibres and Their Response to Carcass Quality in Chickens. Ital. J. Anim. Sci. 2015, 14, 4145. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Wu, H.; Ye, Y.; Li, Z.; Hao, S.; Kong, L.; Zheng, X.; Lin, S.; Nie, Q.; Zhang, X. The transient expression of miR-203 and its inhibiting effects on skeletal muscle cell proliferation and differentiation. Cell Death Dis. 2014, 5, e1347. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Luo, W.; Abdalla, B.A.; Ouyang, H.; Yu, J.; Hu, F.; Nie, Q.; Zhang, X. miRNA-223 upregulated by MYOD inhibits myoblast proliferation by repressing IGF2 and facilitates myoblast differentiation by inhibiting ZEB1. Cell Death Dis. 2017, 8, e3094. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.S.; Yang, X.H.; Wang, X.D.; Wang, Y.L.; Zhou, B.; Song, Z.S. MiR-214 regulate gastric cancer cell proliferation, migration and invasion by targeting PTEN. Cancer Cell Int. 2013, 13, 68. [Google Scholar] [CrossRef]
- Xia, H.; Ooi, L.L.P.J.; Hui, K.M. MiR-214 Targets β-Catenin Pathway to Suppress Invasion, Stem-Like Traits and Recurrence of Human Hepatocellular Carcinoma. PLoS ONE 2012, 7, e44206. [Google Scholar] [CrossRef]
- Flynt, A.S.; Li, N.; Thatcher, E.J.; Solnica-Krezel, L.; Patton, J.G. Zebrafish miR-214 modulates Hedgehog signaling to specify muscle cell fate. Nat. Genet. 2007, 39, 259–263. [Google Scholar] [CrossRef]
- Liu, J.; Luo, X.J.; Xiong, A.W.; Zhang, Z.D.; Yue, S.; Zhu, M.S.; Cheng, S.Y. MicroRNA-214 Promotes Myogenic Differentiation by Facilitating Exit from Mitosis via Down-regulation of Proto-oncogene N-ras. J. Biol. Chem. 2010, 285, 26599–26607. [Google Scholar] [CrossRef]
- Li, T.; Zhang, G.; Wu, P.; Duan, L.; Li, G.; Liu, Q.; Wang, J. Dissection of Myogenic Differentiation Signatures in Chickens by RNA-Seq Analysis. Genes 2018, 9, 34. [Google Scholar] [CrossRef]
- Schwartz, S. M1A within cytoplasmic mRNAs at single nucleotide resolution: A reconciled transcriptome-wide map. RNA 2018, 24, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Ma, M.; Chen, B.; Li, Z.; Abdalla, B.A.; Nie, Q.; Zhang, X. MiR-16-5p targets SESN1 to regulate the p53 signaling pathway, affecting myoblast proliferation and apoptosis, and is involved in myoblast differentiation. Cell Death Dis. 2018, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Tan, W.-H.; Liu, W.; Jin, Y.-X.; Liu, Q. Effects of miR-214 on cervical cancer cell proliferation, apoptosis and invasion via modulating PI3K/AKT/mTOR signal pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Li, L.; Wang, J.; Liu, Z. The effect of miR-214 on HepG2 cell proliferation and cell cycle. Chin. Public Health 2017, 33, 614–617. [Google Scholar]
- El-Deiry, W.S.; Tokino, T.; Velculescu, V.E.; Levy, D.B.; Parsons, R.; Trent, J.M.; Lin, D.; Mercer, W.E.; Kinzler, K.W.; Vogelstein, B.E.; et al. WAF1, a potential mediator of p53 tumor suppression. Cell 1993, 75, 817–825. [Google Scholar] [CrossRef]
- Liu, L.; Cai, X.; Liu, E.; Tian, X.; Tian, C. MicroRNA-18a promotes proliferation and metastasis in hepatocellular carcinoma via targeting KLF4. Oncotarget 2017, 8, 68263. [Google Scholar] [CrossRef]
- Speidel, D. Transcription-independent p53 apoptosis: An alternative route to death. Trends Cell Biol. 2010, 20, 14–24. [Google Scholar] [CrossRef]
- Juan, A.H.; Kumar, R.M.; Marx, J.G.; Young, R.A.; Sartorelli, V. Mir-214-Dependent Regulation of the Polycomb Protein Ezh2 in Skeletal Muscle and Embryonic Stem Cells. Mol. Cell 2009, 36, 61–74. [Google Scholar] [CrossRef]
- Lee, T.I.; Jenner, R.G.; Boyer, L.A.; Guenther, M.G.; Levine, S.S.; Kumar, R.M.; Chevalier, B.; Johnstone, S.E.; Cole, M.F.; Isono, K.; et al. Control of developmental regulators by Polycomb in human embryonic stem cells. Cell 2006, 125, 301–313. [Google Scholar] [CrossRef]
- Caretti, G.; Di Padova, M.; Micales, B.; Lyons, G.E.; Sartorelli, V. The Polycomb Ezh2 methyltransferase regulates muscle gene expression and skeletal muscle differentiation. Genes Dev. 2004, 18, 2627–2638. [Google Scholar] [CrossRef]
- Huang, W.; Guo, L.; Zhao, M.; Zhang, D.; Xu, H.; Nie, Q. The Inhibition on MDFIC and PI3K/AKT Pathway Caused by miR-146b-3p Triggers Suppression of Myoblast Proliferation and Differentiation and Promotion of Apoptosis. Cells 2019, 8, 656. [Google Scholar] [CrossRef] [PubMed]
- Safra, M.; Sas-Chen, A.; Nir, R.; Winkler, R.; Nachshon, A.; Bar-Yaacov, D.; Erlacher, M.; Rossmanith, W.; Stern-Ginossar, N.; Schwartz, S. The m1A landscape on cytosolic and mitochondrial mRNA at single-base resolution. Nature 2017, 551, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, Q.; Kaboli, P.J.; Shen, J.; Li, M.; Wu, X.; Yin, J.; Zhang, H.; Wu, Y.; Lin, L.; et al. m1A Regulated Genes Modulate PI3K/AKT/mTOR and ErbB Pathways in Gastrointestinal Cancer. Transl. Oncol. 2019, 12, 1323–1333. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequences (5′ to 3′) | Product Size (bp) | Annealing Temperature (°C) | Accession Number |
|---|---|---|---|---|
| TRMT61A | F: GTGGGAAGCCATTGGACAT | 266 | 57 | XM_421386.6 |
| R: CTGGGCTACCTTGGTTTGAT | ||||
| p21 | F: CCCGTAGACCACGAGCAGAT | 102 | 61 | NM_204396.1 |
| R: CGTCTCGGTCTCGAAGTTGA | ||||
| MYOD | GCTACTACACGGAATCACCAAAT | 200 | 53 | NM_204214.2 |
| R: CTGGGCTCCACTGTCACTCA | ||||
| MYHC | F: CTCCTCACGCTTTGGTAA | 213 | 53 | NM_001319304.1 |
| R: TGATAGTCGTATGGGTTGGT | ||||
| MYOG | F: CGGAGGCTGAAGAAGGTGAA | 320 | 53 | NM_204184.1 |
| R: CGGTCCTCTGCCTGGTCAT | ||||
| β-actin | F: CAGCCATCTTTCTTGGGTAT | 169 | 60 | NM_205518.1 |
| R: CTGTGATCTCCTTCTGCATCC |
| Gene Name | Primer Sequences (5′ to 3′) | Annealing Temperature (°C) |
|---|---|---|
| gga-miR-214 | F: CGCGACAGCAGGCACAGAC | 60 |
| R: AGTGCAGGGTCCGAGGTATT | ||
| U6 | F: GTCACTTCTGGTGGCGGTAA | 60 |
| R: GTTCAGTAGAGGGTCAAA | ||
| Stem loop primer | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCTGCCT |
| Sequence Name | Sequence Information |
|---|---|
| miR-214 mimic | ACAGCAGGCACAGACAGGCAG |
| GCCUGUCUGUGCCUGCUGUUU | |
| Mimic NC | UUCUCCGAACGUGUCACGUTT |
| ACGUGACACGUUCGGAGAATT | |
| miR-214 inhibitor | CUGCCUGUCUGUGCCUGCUGU |
| Inhibitor NC | CAGUACUUUUGUGUAGUACAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, Y.; Wu, Y.; Yin, X.; Li, T.; Chen, F.; Wu, P.; Zhang, S.; Wang, J.; Zhang, G. MicroRNA-214 Inhibits Chicken Myoblasts Proliferation, Promotes Their Differentiation, and Targets the TRMT61A Gene. Genes 2020, 11, 1400. https://doi.org/10.3390/genes11121400
Duan Y, Wu Y, Yin X, Li T, Chen F, Wu P, Zhang S, Wang J, Zhang G. MicroRNA-214 Inhibits Chicken Myoblasts Proliferation, Promotes Their Differentiation, and Targets the TRMT61A Gene. Genes. 2020; 11(12):1400. https://doi.org/10.3390/genes11121400
Chicago/Turabian StyleDuan, Yanjun, Yulin Wu, Xuemei Yin, Tingting Li, Fuxiang Chen, Pengfei Wu, Shanshan Zhang, Jinyu Wang, and Genxi Zhang. 2020. "MicroRNA-214 Inhibits Chicken Myoblasts Proliferation, Promotes Their Differentiation, and Targets the TRMT61A Gene" Genes 11, no. 12: 1400. https://doi.org/10.3390/genes11121400
APA StyleDuan, Y., Wu, Y., Yin, X., Li, T., Chen, F., Wu, P., Zhang, S., Wang, J., & Zhang, G. (2020). MicroRNA-214 Inhibits Chicken Myoblasts Proliferation, Promotes Their Differentiation, and Targets the TRMT61A Gene. Genes, 11(12), 1400. https://doi.org/10.3390/genes11121400