Single Nucleotide Polymorphisms of NUCB2 and Their Genetic Associations with Milk Production Traits in Dairy Cows

1
Department of Animal Genetics, Breeding and Reproduction, College of Animal Science and Technology, Key Laboratory of Animal Genetics, Breeding and Reproduction of Ministry of Agriculture and Rural Affairs, National Engineering Laboratory for Animal Breeding, China Agricultural University, Beijing 100193, China
2
Beijing Key Laboratory of Dairy Cattle Genetic, Breeding and Reproduction, Beijing Dairy Cattle Center, Beijing 100192, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Genes 2019, 10(6), 449; https://doi.org/10.3390/genes10060449
Submission received: 6 May 2019
/
Revised: 4 June 2019
/
Accepted: 12 June 2019
/
Published: 13 June 2019
(This article belongs to the Section Animal Genetics and Genomics)
Abstract
:We previously used the RNA sequencing technique to detect the hepatic transcriptome of Chinese Holstein cows among the dry period, early lactation, and peak of lactation, and implied that the nucleobindin 2 (NUCB2) gene might be associated with milk production traits due to its expression being significantly increased in early lactation or peak of lactation as compared to dry period (q value < 0.05). Hence, in this study, we detected the single nucleotide polymorphisms (SNPs) of NUCB2 and analyzed their genetic associations with milk yield, fat yield, fat percentage, protein yield, and protein percentage. We re-sequenced the entire coding and 2000 bp of 5′ and 3′ flanking regions of NUCB2 by pooled sequencing, and identified ten SNPs, including one in 5′ flanking region, two in 3′ untranslated region (UTR), and seven in 3′ flanking region. The single-SNP association analysis results showed that the ten SNPs were significantly associated with milk yield, fat yield, fat percentage, protein yield, or protein percentage in the first or second lactation (p values <= 1 × 10−4 and 0.05). In addition, we estimated the linkage disequilibrium (LD) of the ten SNPs by Haploview 4.2, and found that the SNPs were highly linked in one haplotype block (D′ = 0.98–1.00), and the block was also significantly associated with at least one milk traits in the two lactations (p values: 0.0002–0.047). Further, we predicted the changes of transcription factor binding sites (TFBSs) that are caused by the SNPs in the 5′ flanking region of NUCB2, and considered that g.35735477C>T might affect the expression of NUCB2 by changing the TFBSs for ETS transcription factor 3 (ELF3), caudal type homeobox 2 (CDX2), mammalian C-type LTR TATA box (VTATA), nuclear factor of activated T-cells (NFAT), and v-ets erythroblastosis virus E26 oncogene homolog (ERG) (matrix similarity threshold, MST > 0.85). However, the further study should be performed to verify the regulatory mechanisms of NUCB2 and its polymorphisms on milk traits. Our findings first revealed the genetic effects of NUCB2 on the milk traits in dairy cows, and suggested that the significant SNPs could be used in genomic selection to improve the accuracy of selection for dairy cattle breeding.
1. Introduction
Milk provides rich nutrients, protein, fatty acids, and vitamins, etc., for human [1], so the milk production traits are considered as the most important economic traits in the dairy industry, including milk yield, fat yield, fat percentage, protein yield, and protein percentage [2]. Researchers have attempted to identify the functional genes that have large effects on milk production traits to improve the accuracy of selection since the implementation of whole genomic selection. At the present, the RNA sequencing has been widely applied to study the specific gene expression patterns at different developmental stages or in different tissues [3]. We previously used this technique to obtain hepatic transcriptome of Chinese Holstein cows among dry period, early lactation, and peak of lactation, and found that the expression of the nucleobindin 2 (NUCB2) gene was significantly increased in early lactation (q value = 0.00811) and the peak of lactation (q value = 0.049246) as compared to dry period, respectively [4], so we considered that the NUCB2 might be associated with milk production traits.
Nucleobindin 2 (NUCB2) is a Ca2+ binding protein, and it processes to generate a terminal peptide, termed nesfatin-1. Nesfatin-1 regulates the appetite and glucose metabolisms in humans and domestic animals [5,6,7]. NUCB2 exhibits a high conservation among species, and its mRNA is ubiquitously expressed [8]. Sun et al. first demonstrated that NUCB2 mRNA and nesfatin-1 protein were highly expressed in the liver of mouse fetus at embryonic day (E) 10.5, and their expression were extensively decreased at E17.5, which suggests that NUCB2 might play a critical role in liver development and physiological functions in the developmental process of mouse fetus [9]. NUCB2 is involved in human mammary epithelial cell proliferation and migration [10], and its expression is down-regulated when the milk production is decreased [11], which implies that the expression of NUCB2 might be beneficial to the increase of milk production. In addition, the NUCB2 gene was located on chr.15:43.5418 and it had a distance of 0.86 and 2.54 cM to the peak of reported quantitative trait locus (QTL) regions that have large effect on protein percentage [12] and protein yield [13], respectively. The NUCB2 was also near to three significant SNPs for milk traits, BFGL-NGS-116109, BTB-00590405, and BTB-00590603, with the distance of 0.93–5.99 Mb [14]. These data indicate that NUCB2 may be involved in milk production traits.
To date, rare studies have been reported to uncover the genetic association between the single nucleotide polymorphisms (SNPs) of NUCB2 and milk production traits in dairy cattle. In addition, the studies have revealed that haplotype blocks that are formed by the SNPs have important implications for identifying associations with complex traits [15,16,17]. Hence, in this study, we detected the polymorphisms of the NUCB2 gene and estimated the haplotype block that was formed by the identified SNPs, and then analyzed their associations with five milk traits, including milk yield, fat yield, fat percentage, protein yield, and protein percentage.
2. Materials and Methods
2.1. Animals
We selected a total of 1067 Chinese Holstein cows of 40 sire families from 22 dairy farms of the Sanyuan Lvhe Dairy Farming Centre (Beijing, China) on the same feeding conditions. The cows in each sire family were distributed in different dairy farms. These 1067 cows have three-generation pedigree information at least and Dairy Herd Improvement (DHI) records for 305-day milk yield, protein yield, protein percentage, fat yield, and fat percentage. Ethics Approval: The Institutional Animal Care and Use Committee (IACUC) of China Agricultural University approved all of the protocols for sample collection (permit number: DK996).
2.2. Phenotypic Data Collection
The Beijing Dairy Cattle Centre provided the phenotypic data of 305-day milk yield, fat yield, fat percentage, protein yield, and protein percentage in the first and second lactations (http://www.bdcc.com.cn/), and the descriptive statistics of them are shown in Table S1.
2.3. DNA Extraction
The semen DNAs of 40 sires were extracted using the salt-out procedures, and the blood DNAs of 1067 daughters were extracted while using the TIANamp Blood DNA Kit (Tiangen, Beijing, China), according to the manufacturer’s instructions. Subsequently, we measured the quantity and quality of the extracted DNAs by the NanoDrop 2000 spectrophotometer (Thermo Scientific, Hudson, NH, USA) and gel electrophoresis, respectively.
2.4. SNP Identification and Genotyping
We designed the primers (Table S2) to amplify the entire coding region and 2000 bp of the 5′ and 3′ flanking regions based on the bovine reference genome sequences of NUCB2 by Primer3 (version 0.4.0, Whitehead Institute for Biomedical Research, Cambridge, MA, USA) (http://bioinfo.ut.ee/primer3-0.4.0/), and then synthesized them in the Beijing Genomics Institute (BGI, Beijing, China). We randomly mixed 40 semen DNAs into two pools (20 sires per pool) with equal concentrations (50 ng/µL) for each DNA. The reaction conditions and amplification procedures for PCR are provided in the Table S2. Afterwards, the PCR products were purified and sequenced by the ABI3730XL DNA analyser (Applied Biosystems, Foster City, CA, USA). Subsequently, we analyzed the sequencing data while using CHROMAS (version 2.23, Technelysium, Tewantin, Queensland, Australia) to discovery the potential SNPs. The identified SNPs were individually genotyped using Sequenom MassArray (Agena Bioscience, San Diego, CA, USA) for all the 1067 cows by matrix-assisted laser desorption/ionization time of flight mass spectrometry (MALDI-TOF MS, Bioyong Technologies Inc., HK).
2.5. Estimation of Linkage Disequilibrium (LD) and Association Analyses
The extent of LD between the identified SNPs was estimated while using Haploview 4.2 (Broad Institute of MIT and Harvard, Cambridge, MA, USA). Subsequently, we used the Statistical Analysis System (SAS) 9.13 software (SAS Institute, Cary, NC, USA) to analyze the associations of the SNPs and haplotype blocks with milk production traits with the following animal model: , in which, is the phenotypic value of each trait for each cow; μ is the overall mean; hys is the fixed effect of farm, year, and season; is the regression coefficient of covariant ; is the fixed effect of calving month; is the genotype or haplotype combination effect; is the individual random additive genetic effect, being distributed as , with the additive genetic variance ; and, e is the random residual, which is distributed as , with identity matrix I and residual error variance . Bonferroni correction was performed for multiple testing, and the significant level was equal to the raw P value, divided by number of genotypes or haplotype combinations. In addition, the additive (a), dominant (d), and substitution (α) effects were calculated, as follows: , where, AA, BB, and AB are the least square means of the milk production traits in the corresponding genotypes, p is the frequency of allele A, and q is the frequency of allele B.
2.6. Transcription Factor Binding Sites (TFBSs) Prediction
We used the MatInspector (version 3.11, Genomatix, Munich, Germany) (http://www.genomatix.de/cgi-bin/welcome/welcome.pl?s=d1b5c9a9015b02bb3b1a806f9c03293f) to predict the changes of the TFBSs that are caused by the identified SNPs in the 5′ flanking region of the NUCB2 (matrix similarity threshold, MST > 0.85).
2.7. NUCB2 Gene Expression Analysis
Three healthy lactating Chinese Holstein cows were selected from the Sanyuanlvhe Dairy Farming Center (Beijing, China), and the heart, liver, spleen, lung, kidney, ovary, mammary, and uterus from each cow were collected. Subsequently, the expression of NUCB2 in eight tissues was detected to further reveal its potential function. The total RNAs of the tissues were extracted while using a Trizol reagent (Invitrogen, Carlsbad, CA, USA), and the quantity and quality of the RNA were measured with a NanoDrop 2000 spectrophotometer (Thermo Scientific, Hudson, DE, USA) and gel electrophoresis, respectively. We used a PrimerScriptH RT reagent Kit (TaKaRa Biotechnology Co., Ltd., Dalian, China) for the reverse transcription. The qPCR primers of NUCB2 and two reference genes (Ribosomal Protein S9, RPS9, and Ubiquitously Expressed Prefoldin Like Chaperone, UXT) were presented in Table S2. We conducted the qPCR while using a LightCycler® 480 II (Roche, Penzberg, Germany) with the procedures that are shown in Table S2. All of the measurements were performed in triplicate and the relative gene expression was normalized by the RPS9 and UXT with 2-ΔΔCt method.
3. Results
3.1. SNP Identification in NUCB2 Gene
By re-sequencing, we identified ten SNPs in the entire coding region and 2000 bp of 5′ and 3′ flanking regions of NUCB2, including one in the 5′ flanking region, two in 3′ UTR, and seven in the 3′ flanking region (Table 1). Further, we individually genotyped these ten SNPs while using Sequenom MassArray for all the 1067 cows, and their allelic and genotypic frequencies are shown in Figure 1.
3.2. Associations between Ten SNPs and Five Milk Production Traits
We analyzed the genetic associations between the ten SNPs of NUCB2 and milk yield, fat yield, fat percentage, protein yield, and protein percentage, respectively, by SAS, and the results showed that these ten SNPs were significantly associated with at least one milk traits in the first or second lactation (Table 2). In the first lactation, there were five SNPs that were associated with fat yield (p values: 0.0012–0.0178), nine with fat percentage (p values: 0.0035–0.0388), and eight with protein percentage (p values: 0.0075–0.0334), and no association was found between the SNPs and milk and protein yields (p values > 0.05). In the second lactation, ten SNPs had strong associations with milk yield (p values <= 1 × 10−4 and 0.0089), five with fat yield (p values: 0.0073–0.0466), one with fat percentage (p value = 0.0257), ten with protein yield (p values <= 1 × 10−4 and 0.0347), and six with protein percentage (p values: 0.0107–0.0467), respectively. Two SNPs, c.*1253A>G and g.35692674A>G, were strongly associated with fat yield (p values: 0.0073–0.0383) in both of the two lactations. The g.35735477C>T had significant association with fat percentage in the first (p value = 0.0122) and second lactation (p value = 0.0257). There were four SNPs, g.35735477C>T, c.*1276C>T, g.35692145C>T, and g.35691328T>C, which were associated with protein percentage ((p values: 0.0075–0.0357) in both lactations. In addition, as the results show in Table S3, the additive, dominant, and substitution effects of the ten SNPs were found to be significantly associated with milk traits (p values < 0.05).
3.3. Associations between Haplotype Combinations with Five Milk Traits
We estimated the LD among the identified SNPs of NUCB2 and found a haplotype block (D′ =0.98–1.00; Figure 2) that was included all ten SNPs. The block consisted of three haplotypes, H1 (TACTTAACAC), H2 (TACTTAACAT), and H3 (CGTCCGGTGT), with the frequency of 75.3%, 1.3%, and 22.6%, respectively. By haplotype-based association analyses, we found that the haplotype block was significantly associated with protein percentage (p value = 0.0393) in the first lactation, milk yield (p value = 0.0002), fat percentage (p value = 0.047), protein yield (p value = 0.0015), and protein percentage (p value = 0.0386) in the second lactation, respectively (Table 3).
3.4. TFBSs Changed by g.35735477C>T of NUCB2
For the SNP, g.35735477C>T, in the 5′ flanking region of the NUCB2 gene, we predicted the changes of TFBSs caused by them while using Matinspector. The allele C in g.35735477C>T was predicted to create the binding sites for the transcription factors E74, like ETS transcription factor 3 (ELF3; MST = 0.89), caudal type homeobox 2 (CDX2; MST = 0.93), mammalian C-type LTR TATA box (VTATA; MST = 0.92), and the allele T in this SNP could create the TFBSs for nuclear factor of activated T-cells (NFAT; MST = 0.96) and v-ets erythroblastosis virus E26 oncogene homolog (ERG; MST = 0.94; Figure 3).
3.5. Expression of NUCB2 in Tissues
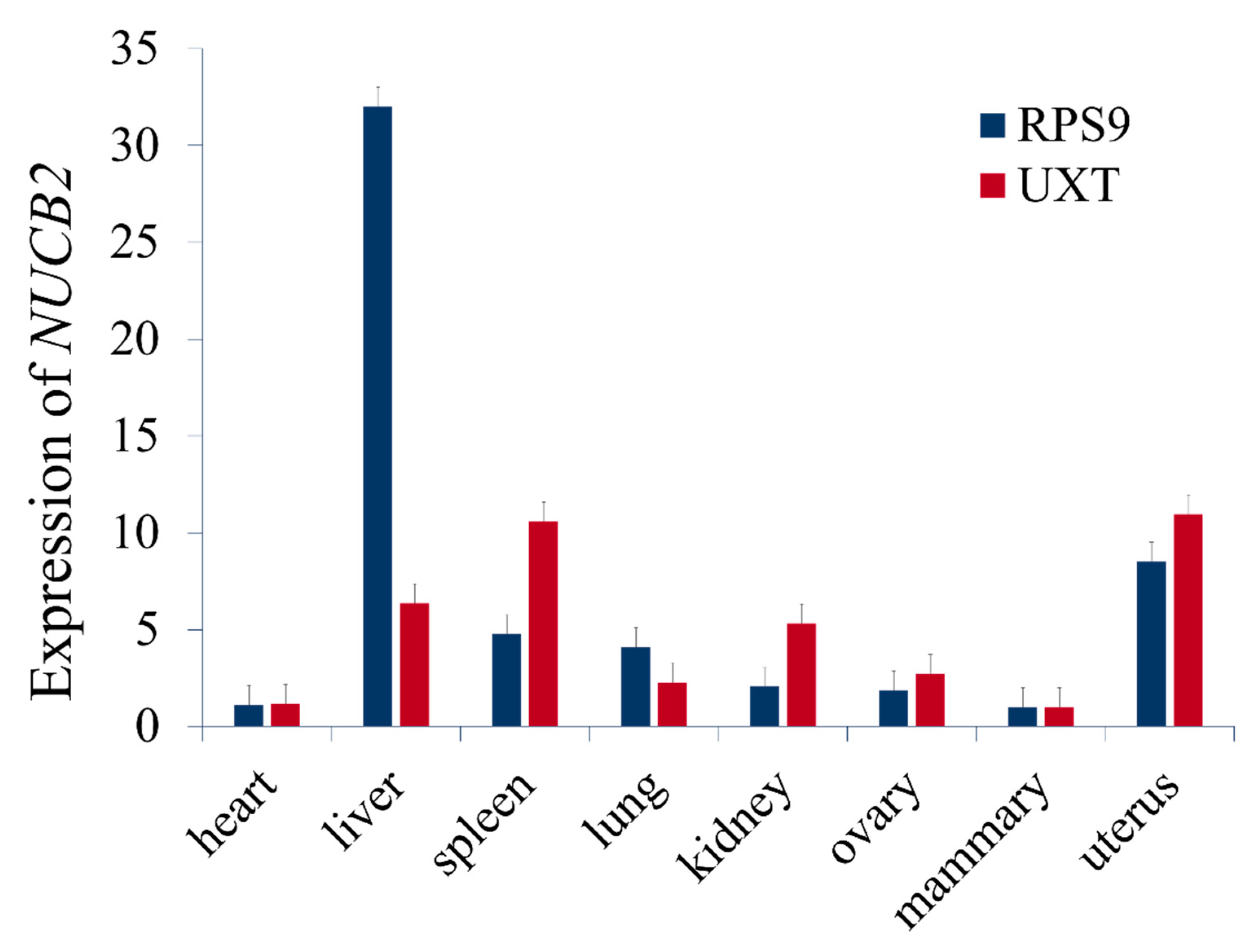
We found that NUCB2 gene was expressed in heart, liver, spleen, lung, kidney, ovary, mammary gland, and uterus of the lactating Chinese Holstein and highly expressed in liver tissue while using the qPCR, (Figure 4), which suggested that it might be involved with substance metabolisms for milk synthesis in the liver.
4. Discussion
NUCB2 was involved with milk production traits in dairy cows n our previous hepatic transcriptome study, and this follow-up investigation underlined that the SNPs of NUCB2 have significant genetic effects on milk yield and composition.
Genomic selection has revolutionized dairy cattle breeding, and the evaluation system with DNA marker technology and genomics has doubled the rate of genetic progress for economic traits [18]. A study has revealed that the use of available loci improved the accuracy of genomic prediction in dairy cattle [19], which suggests that loci that are associated with milk production traits could be used in genomic selection programs that are aimed at accelerating genetic progress of dairy cattle. Studies have shown that the SNPs in the candidate genes significantly influenced the milk yield and composition in Holsteins [20,21,22,23,24,25]. In this study, we identified ten SNPs in NUCB2 and found that the SNPs and haplotypes of this gene were significantly associated with milk production traits. Currently, the Illumina 50K/770K chip and the GeneSeek 90K/150K chip are commonly used, and most of the SNP markers in chips were from the SNP database with even distribution across the whole genome. Thus, the certain SNPs of NUCB2 gene with significant genetic effects on milk traits could be put into the SNP chip to increase the selection efficiency for the milk production in dairy cattle.
In addition, for each identified SNP of NUCB2 in the current study, we observed that the genotypic frequency of the cows with significantly high phenotypic values of 305-day milk yield, fat yield, fat percentage, protein yield, or protein percentage were higher. This implied the effectiveness of dairy cattle breeding in recent years, and also reflected that these loci might be closely associated with the milk production traits. However, we found that the ten SNPs of NUCB2 showed different associations between the first and second lactations, and considered that the possible reason might be the different number of cows that are used for genetic association analysis, that is, we used 1067 cows in the first lactation and 740 in the second lactation (327 cows merely completed the milking of first lactation), which might impact the statistical significance. Generally, cows have higher milk production in the second lactation, and the different physiologic status of cows between the two lactations was also one possibility for the different genetic effects across lactations. In addition, the unstable association of the SNPs in two lactations was probably due to the fact that the causal polymorphisms are not very far, but not for these ones. Besides, we observed that the cows with heterozygous genotype showed the dominance for milk traits, ten SNPs for milk yield in the second lactation, for instance, which were caused by the dominance effects of the heterozygotes. We usually do not consider it in breeding, since the dominant effect is not heritable across generations. Hence, we should choose SNPs with significant additive effects on milk traits in all lactations for dairy cattle breeding.
In this study, we used the qPCR and observed that NUCB2 was expressed in heart, liver, spleen, lung, kidney, ovary, mammary, and uterus tissues of dairy cows, and relatively highly expressed in liver. In dairy cows, the liver participates in carbohydrate, fat, protein, and other substance metabolisms, through blood circulation, it provides cholesterol, triglyceride, and other ingredients to mammary gland to synthesize milk protein and fat [26,27,28]. Our data suggest that NUCB2 may play an important role in milk synthesis metabolism in dairy cows.
The specific binding of transcription factors to the regulatory regions of the DNAs is a important regulatory mechanism that affects gene expression [29,30]. In the current study, the allele C of g.35735477C>T was predicted to create the binding sites for ELF3, CDX2, and VTATA, and the allele T to invent the binding sites for NFAT and ERG. Oettgen et al. revealed that ELF3 directly drives SPRR2A mRNA expression through binding to the sites in its promoter during terminal differentiation of the epidermis [31]. The overexpression of ELF3 transactivates the promoter of KRT12 in the differentiated mouse corneal epithelium [32]. ELF3 binds to the miR-141-3p promoter and it suppresses its expression, thereby promoting the epithelial-mesenchymal transition in human hepatocellular carcinoma [33]. ELF3 directly binds Estrogen Receptor α (ERα) and it represses its transcriptional activity in ERα positive breast cancer, showing a tumour suppressive role [34]. Zhu et al. reported that CDX2 directly transactivated the oncogene CDH-17 and promoted hepatocellular carcinoma cell proliferation [35]. CDX2 transactivates the expression of GSK-3β and Axin2 to inhibit the cell proliferation and tumor formation in colon cancer [36]. CDX2 binds to the distinct target genes and it activates their expression in developing versus human and adult mouse intestinal cells [37]. Ross et al. identified a large number of potential TFBSs in the 5′ flanking sequences of SRY gene in human and bovine, including the TFBS for VTATA in the TATA-binding protein factors family with the matrix similarity score of 0.923 in bovine [38]. The NFAT family consists of five members, NFAT1–NFAT5, first found in T lymphocytes as an inducible nuclear factor that could bind to the IL-2 promoter and transactivate its expression [39]. NFAT proteins frequently cooperate with other transcriptional partners due to its weak DNA-binding capacity, and reports have shown the specific molecules that are up-/down-regulated by the different NFATs [40]. ERG regulates the expression of SMAD2/3 to cause endothelial-dependent liver fibrogenesis [41], and it promotes endothelial homeostasis via the regulation of lineage-specific enhancers and super-enhancers [42]. These data indicate that these five transcription factors can regulate the expression of target genes by binding the TFBSs. Therefore, we speculated that the g.35735477C>T could affect the expression of NUCB2 by the coactions of transcription factors ELF3, CDX2, VTATA, NFAT, and ERG. However, the further functional validation should be performed to verify the regulatory roles of NUCB2 and its polymorphisms on milk traits in dairy cattle.
5. Conclusions
This is the first study to demonstrate the genetic associations of SNPs of NUCB2 gene with milk production traits in dairy cows, and to show that g.35735477C>T might change the TFBSs of NUCB2. Our findings provide valuable molecular information for dairy cattle breeding, and the significant SNPs could be put into the SNP chip for genomic selection to improve the milk production in dairy cows.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4425/10/6/449/s1, Table S1: Descriptive statistics of the phenotypic values for milk production traits. Table S2: Primers and procedures for PCR and qPCR used in SNP identification. Table S3: Additive, dominant and allele substitution effects of SNPs on milk production traits in Chinese Holstein.
Author Contributions
Data curation, B.H., Y.Y., Y.L., L.L. and D.S.; Formal analysis, B.H. and Y.Y.; Funding acquisition, B.H. and D.S.; Investigation, B.H. and Y.Y.; Methodology, B.H., Y.Y., and D.S.; Project administration, D.S.; Resources, Y.L., L.L. and D.S.; Supervision, D.S.; Writing—original draft, B.H; Writing—review and editing, B.H. and D.S.
Funding
This research was funded by National Natural Science Foundation of China, grant number 31802041 and 31872330; Beijing Municipal Bureau of Agriculture, grant number BAIC06-2018/2019; Beijing Municipal Science and Technology Commission, grant number D171100002417001; Chinese Ministry of Science and Technology, grant number 2013AA102504; Chinese Ministry of Agriculture and Rural Affairs, grant number CARS-36; Chinese Ministry of Education, grant number IRT_15R62.
Acknowledgments
This work was financially supported by the National Natural Science Foundation of China (31802041, 31872330), Beijing Dairy Industry Innovation Team (BAIC06-2018/2019), Beijing Science and Technology Program (D171100002417001), National Science and Technology Programs of China (2013AA102504), earmarked fund for Modern Agro-industry Technology Research System (CARS-36), and the Program for Changjiang Scholar and Innovation Research Team in University (IRT_15R62).
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Tunick, M.H.; Van Hekken, D.L. Dairy Products and Health: Recent Insights. J. Agric. Food Chem. 2015, 63, 9381–9388. [Google Scholar] [CrossRef]
- Spelman, R.J.; Coppieters, W.; Karim, L.; van Arendonk, J.A.; Bovenhuis, H. Quantitative trait loci analysis for five milk production traits on chromosome six in the Dutch Holstein-Friesian population. Genetics 1996, 144, 1799–1808. [Google Scholar]
- Denoeud, F.; Aury, J.M.; Da Silva, C.; Noel, B.; Rogier, O.; Delledonne, M.; Morgante, M.; Valle, G.; Wincker, P.; Scarpelli, C.; et al. Annotating genomes with massive-scale RNA sequencing. Genome Biol. 2008, 9, R175. [Google Scholar] [CrossRef]
- Liang, R.; Han, B.; Li, Q.; Yuan, Y.; Li, J.; Sun, D. Using RNA sequencing to identify putative competing endogenous RNAs (ceRNAs) potentially regulating fat metabolism in bovine liver. Sci. Rep. 2017, 7, 6396. [Google Scholar] [CrossRef]
- Oh, S.; Shimizu, H.; Satoh, T.; Okada, S.; Adachi, S.; Inoue, K.; Eguchi, H.; Yamamoto, M.; Imaki, T.; Hashimoto, K.; et al. Identification of nesfatin-1 as a satiety molecule in the hypothalamus. Nature 2006, 443, 709–712. [Google Scholar] [CrossRef]
- Stengel, A.; Tache, Y. Minireview: Nesfatin-1-An Emerging New Player in the Brain-Gut, Endocrine, and Metabolic Axis. Endocrinology 2011, 152, 4033–4038. [Google Scholar] [CrossRef]
- Ramesh, N.; Gawli, K.; Pasupulleti, V.K.; Unniappan, S. Metabolic and Cardiovascular Actions of Nesfatin-1: Implications in Health and Disease. Curr. Pharm. Des. 2017, 23, 1453–1464. [Google Scholar] [CrossRef]
- Mohan, H.; Unniappan, S. Phylogenetic aspects of nucleobindin-2/nesfatin-1. Curr. Pharm. Des. 2013, 19, 6929–6934. [Google Scholar] [CrossRef]
- Sun, S.; Yang, H. Tissue-Specific Localization NUCB2/nesfatin-1 in the Liver and Heart of Mouse Fetus. Dev. Reprod. 2018, 22, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Takagi, K.; Miki, Y.; Onodera, Y.; Akahira, J.; Ebata, A.; Ishida, T.; Watanabe, M.; Sasano, H.; Suzuki, T. Nucleobindin 2 in human breast carcinoma as a potent prognostic factor. Cancer Sci. 2012, 103, 136–143. [Google Scholar] [CrossRef]
- Boutinaud, M.; Galio, L.; Lollivier, V.; Finot, L.; Wiart, S.; Esquerre, D.; Devinoy, E. Unilateral once daily milking locally induces differential gene expression in both mammary tissue and milk epithelial cells revealing mammary remodeling. Physiol. Genom. 2013, 45, 973–985. [Google Scholar] [CrossRef] [Green Version]
- Schopen, G.C.; Koks, P.D.; van Arendonk, J.A.; Bovenhuis, H.; Visker, M.H. Whole genome scan to detect quantitative trait loci for bovine milk protein composition. Anim. Genet. 2009, 40, 524–537. [Google Scholar] [CrossRef]
- Daetwyler, H.D.; Schenkel, F.S.; Sargolzaei, M.; Robinson, J.A.B. A genome scan to detect quantitative trait loci for economically important traits in Holstein cattle using two methods and a dense single nucleotide polymorphism map. J. Dairy Sci. 2008, 91, 3225–3236. [Google Scholar] [CrossRef]
- Cole, J.B.; Wiggans, G.R.; Ma, L.; Sonstegard, T.S.; Lawlor, T.J., Jr.; Crooker, B.A.; Van Tassell, C.P.; Yang, J.; Wang, S.; Matukumalli, L.K.; et al. Genome-wide association analysis of thirty one production, health, reproduction and body conformation traits in contemporary U.S. Holstein cows. BMC Genom. 2011, 12, 408. [Google Scholar] [CrossRef]
- Hagenblad, J.; Tang, C.; Molitor, J.; Werner, J.; Zhao, K.; Zheng, H.; Marjoram, P.; Weigel, D.; Nordborg, M. Haplotype structure and phenotypic associations in the chromosomal regions surrounding two Arabidopsis thaliana flowering time loci. Genetics 2004, 168, 1627–1638. [Google Scholar] [CrossRef]
- Nothnagel, M.; Rohde, K. The effect of single-nucleotide polymorphism marker selection on patterns of haplotype blocks and haplotype frequency estimates. Am. J. Hum. Genet. 2005, 77, 988–998. [Google Scholar] [CrossRef]
- Patil, N.; Berno, A.J.; Hinds, D.A.; Barrett, W.A.; Doshi, J.M.; Hacker, C.R.; Kautzer, C.R.; Lee, D.H.; Marjoribanks, C.; McDonough, D.P.; et al. Blocks of limited haplotype diversity revealed by high-resolution scanning of human chromosome 21. Science 2001, 294, 1719–1723. [Google Scholar] [CrossRef]
- Wiggans, G.R.; Cole, J.B.; Hubbard, S.M.; Sonstegard, T.S. Genomic Selection in Dairy Cattle: The USDA Experience. Annu. Rev. Anim. Biosci. 2017, 5, 309–327. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Ober, U.; Erbe, M.; Zhang, H.; Gao, N.; He, J.; Li, J.; Simianer, H. Improving the accuracy of whole genome prediction for complex traits using the results of genome wide association studies. PLoS ONE 2014, 9, e93017. [Google Scholar] [CrossRef]
- Bouwman, A.C.; Bovenhuis, H.; Visker, M.H.; van Arendonk, J.A. Genome-wide association of milk fatty acids in Dutch dairy cattle. BMC Genet. 2011, 12, 43. [Google Scholar] [CrossRef]
- Matsumoto, H.; Sasaki, K.; Bessho, T.; Kobayashi, E.; Abe, T.; Sasazaki, S.; Oyama, K.; Mannen, H. The SNPs in the ACACA gene are effective on fatty acid composition in Holstein milk. Mol. Biol. Rep. 2012, 39, 8637–8644. [Google Scholar] [CrossRef]
- Bhattarai, D.; Chen, X.; Ur Rehman, Z.; Hao, X.; Ullah, F.; Dad, R.; Talpur, H.S.; Kadariya, I.; Cui, L.; Fan, M.; et al. Association of MAP4K4 gene single nucleotide polymorphism with mastitis and milk traits in Chinese Holstein cattle. J. Dairy. Res. 2017, 84, 76–79. [Google Scholar] [CrossRef]
- Han, B.; Liang, W.; Liu, L.; Li, Y.; Sun, D. Determination of genetic effects of ATF3 and CDKN1A genes on milk yield and compositions in Chinese Holstein population. BMC Genet. 2017, 18, 47. [Google Scholar] [CrossRef]
- Han, B.; Liang, W.; Liu, L.; Li, Y.; Sun, D. Genetic association of the ACACB gene with milk yield and composition traits in dairy cattle. Anim. Genet. 2018. [Google Scholar] [CrossRef]
- Han, B.; Yuan, Y.; Liang, R.; Li, Y.; Liu, L.; Sun, D. Genetic Effects of LPIN1 Polymorphisms on Milk Production Traits in Dairy Cattle. Genes (Basel) 2019, 10, 265. [Google Scholar] [CrossRef]
- Bell, A.W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci. 1995, 73, 2804–2819. [Google Scholar] [CrossRef]
- Rawson, P.; Stockum, C.; Peng, L.F.; Manivannan, B.; Lehnert, K.; Ward, H.E.; Berry, S.D.; Davis, S.R.; Snell, R.G.; McLauchlan, D.; et al. Metabolic proteomics of the liver and mammary gland during lactation. J. Proteom. 2012, 75, 4429–4435. [Google Scholar] [CrossRef]
- Moyes, K.M.; Bendixen, E.; Codrea, M.C.; Ingvartsen, K.L. Identification of hepatic biomarkers for physiological imbalance of dairy cows in early and mid lactation using proteomic technology. J. Dairy Sci. 2013, 96, 3599–3610. [Google Scholar] [CrossRef]
- Tugrul, M.; Paixao, T.; Barton, N.H.; Tkacik, G. Dynamics of Transcription Factor Binding Site Evolution. PLoS Genet. 2015, 11, e1005639. [Google Scholar] [CrossRef]
- Talebzadeh, M.; Zare-Mirakabad, F. Transcription factor binding sites prediction based on modified nucleosomes. PLoS ONE 2014, 9, e89226. [Google Scholar] [CrossRef]
- Oettgen, P.; Alani, R.M.; Barcinski, M.A.; Brown, L.; Akbarali, Y.; Boltax, J.; Kunsch, C.; Munger, K.; Libermann, T.A. Isolation and characterization of a novel epithelium-specific transcription factor, ESE-1, a member of the ETS family. Mol. Cell Biol. 1997, 17, 4419–4433. [Google Scholar] [CrossRef]
- Yoshida, N.; Yoshida, S.; Araie, M.; Handa, H.; Nabeshima, Y. Ets family transcription factor ESE-1 is expressed in corneal epithelial cells and is involved in their differentiation. Mech. Deve. 2000, 97, 27–34. [Google Scholar] [CrossRef]
- Zheng, L.; Xu, M.; Xu, J.; Wu, K.; Fang, Q.; Liang, Y.; Zhou, S.; Cen, D.; Ji, L.; Han, W.; et al. ELF3 promotes epithelial-mesenchymal transition by protecting ZEB1 from miR-141-3p-mediated silencing in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 387. [Google Scholar] [CrossRef]
- Gajulapalli, V.N.R.; Samanthapudi, V.S.K.; Pulaganti, M.; Khumukcham, S.S.; Malisetty, V.L.; Guruprasad, L.; Chitta, S.K.; Manavathi, B. A transcriptional repressive role for epithelial-specific ETS factor ELF3 on oestrogen receptor α in breast cancer cells. Biochem. J. 2016, 473, 1047–1061. [Google Scholar] [CrossRef]
- Zhu, R.; Wong, K.F.; Lee, N.P.; Lee, K.F.; Luk, J.M. HNF1alpha and CDX2 transcriptional factors bind to cadherin-17 (CDH17) gene promoter and modulate its expression in hepatocellular carcinoma. J. Cell Biochem. 2010, 111, 618–626. [Google Scholar] [CrossRef]
- Yu, J.H.; Liu, D.; Sun, X.J.; Yang, K.; Yao, J.F.; Cheng, C.; Wang, C.B.; Zheng, J.B. CDX2 inhibits the proliferation and tumor formation of colon cancer cells by suppressing Wnt/β-catenin signaling via transactivation of GSK-3 β and Axin2 expression. Cell Death Dis. 2019, 10, 26. [Google Scholar] [CrossRef]
- Kumar, N.; Tsai, Y.H.; Chen, L.; Zhou, A.; Banerjee, K.K.; Saxena, M.; Huang, S.; Toke, N.H.; Xing, J.; Shivdasani, R.A.; et al. The lineage-specific transcription factor CDX2 navigates dynamic chromatin to control distinct stages of intestine development. Development 2019, 146. [Google Scholar] [CrossRef]
- Ross, D.G.F.; Bowles, J.; Koopman, P.; Lehnert, S. New insights into SRY regulation through identification of 5 conserved sequences. BMC Mol. Biol. 2008, 9. [Google Scholar] [CrossRef]
- Shaw, J.P.; Utz, P.J.; Durand, D.B.; Toole, J.J.; Emmel, E.A.; Crabtree, G.R. Identification of a putative regulator of early T cell activation genes. Science 1988, 241, 202–205. [Google Scholar] [CrossRef]
- Mognol, G.P.; Carneiro, F.R.; Robbs, B.K.; Faget, D.V.; Viola, J.P. Cell cycle and apoptosis regulation by NFAT transcription factors: New roles for an old player. Cell Death Dis. 2016, 7, e2199. [Google Scholar] [CrossRef]
- Dufton, N.P.; Peghaire, C.R.; Osuna-Almagro, L.; Raimondi, C.; Kalna, V.; Chuahan, A.; Webb, G.; Yang, Y.W.; Birdsey, G.M.; Lalor, P.; et al. Dynamic regulation of canonical TGF β signalling by endothelial transcription factor ERG protects from liver fibrogenesis. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Kalna, V.; Yang, Y.; Peghaire, C.; Frudd, K.; Hannah, R.; Shah, A.V.; Osuna Almagro, L.; Boyle, J.J.; Gottgens, B.; Ferrer, J.; et al. The Transcription Factor ERG Regulates Super-Enhancers Associated with an Endothelial-Specific Gene Expression Program. Circ. Res. 2019. [Google Scholar] [CrossRef]
Figure 1.
Allelic and genotypic frequencies of the ten single nucleotide polymorphisms (SNPs) in nucleobindin 2 (NUCB2). The “*” shows the SNPs located in the 3’untranslated region (UTR).
Figure 1.
Allelic and genotypic frequencies of the ten single nucleotide polymorphisms (SNPs) in nucleobindin 2 (NUCB2). The “*” shows the SNPs located in the 3’untranslated region (UTR).
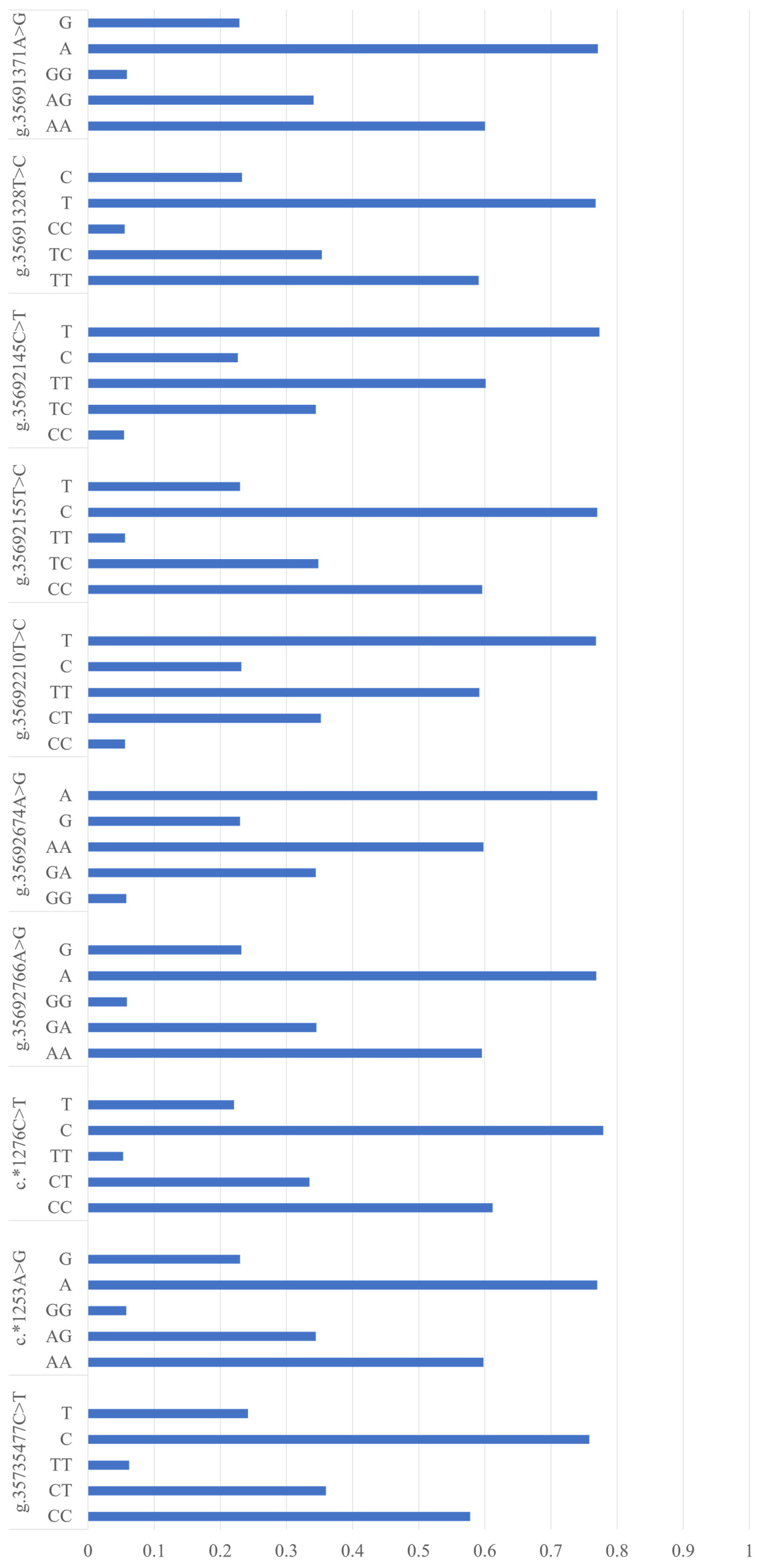
Figure 2.
Linkage disequilibrium estimated among the SNPs of NUCB2 (D′ = 0.98–1.00). The blocks indicate haplotype blocks and the text above the horizontal numbers is the SNP names. The values in boxes are pairwise SNP correlations (D′), while the boxes without numbers indicate complete linkage disequilibrium (LD) (D′ = 1).
Figure 2.
Linkage disequilibrium estimated among the SNPs of NUCB2 (D′ = 0.98–1.00). The blocks indicate haplotype blocks and the text above the horizontal numbers is the SNP names. The values in boxes are pairwise SNP correlations (D′), while the boxes without numbers indicate complete linkage disequilibrium (LD) (D′ = 1).
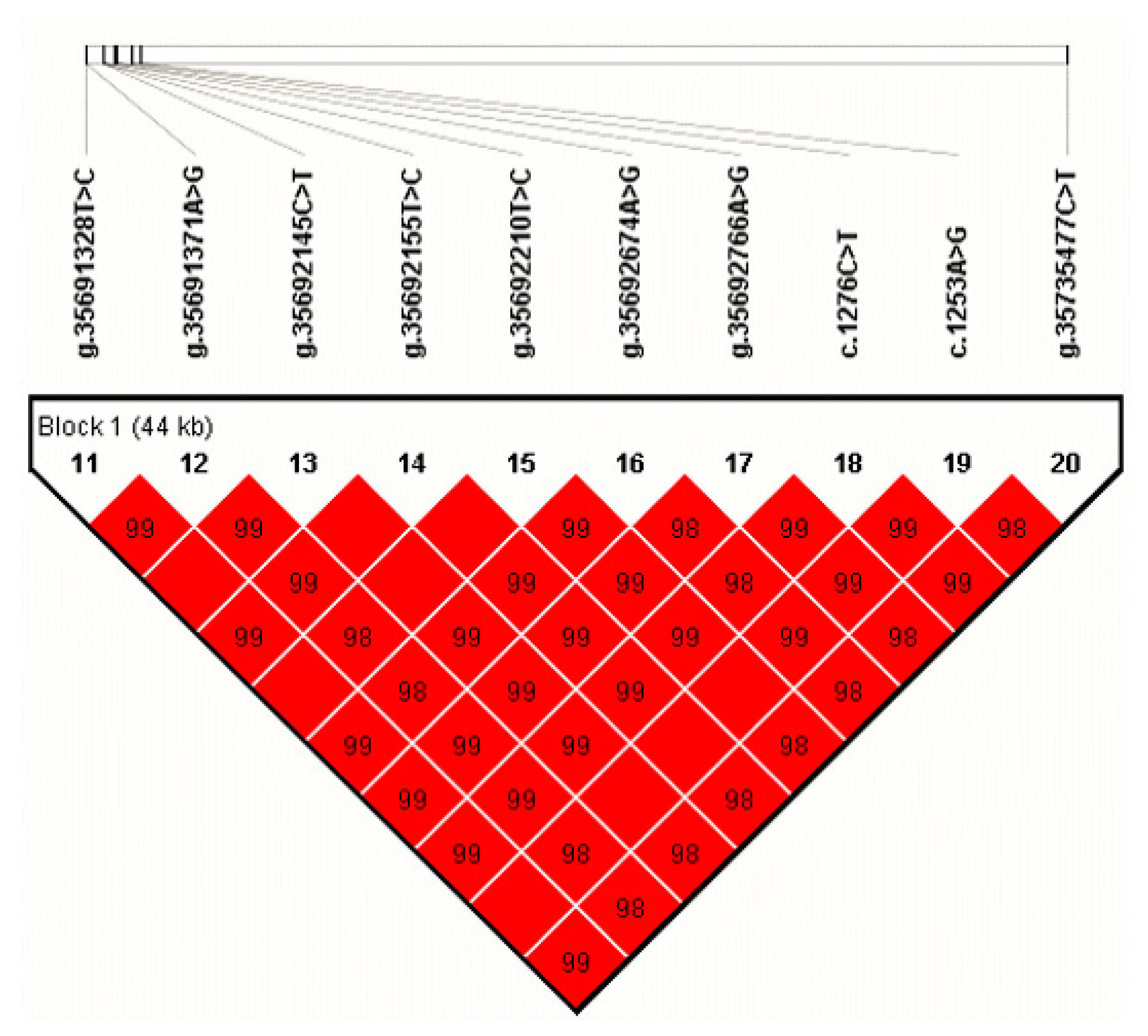
Figure 3.
Transcription factor binding sites (TFBSs) in the 5′ flanking region of NUCB2 gene. The nucleotides represent the transcription factor binding site sequences of ETS transcription factor 3 (ELF3), caudal type homeobox 2 (CDX2), mammalian C-type LTR TATA box (VTATA), nuclear factor of activated T-cells (NFAT), and v-ets erythroblastosis virus E26 oncogene homolog (ERG) with their matrix similarity over 0.85 (in blue), and the underlined nucleotide in red was the SNP.
Figure 3.
Transcription factor binding sites (TFBSs) in the 5′ flanking region of NUCB2 gene. The nucleotides represent the transcription factor binding site sequences of ETS transcription factor 3 (ELF3), caudal type homeobox 2 (CDX2), mammalian C-type LTR TATA box (VTATA), nuclear factor of activated T-cells (NFAT), and v-ets erythroblastosis virus E26 oncogene homolog (ERG) with their matrix similarity over 0.85 (in blue), and the underlined nucleotide in red was the SNP.
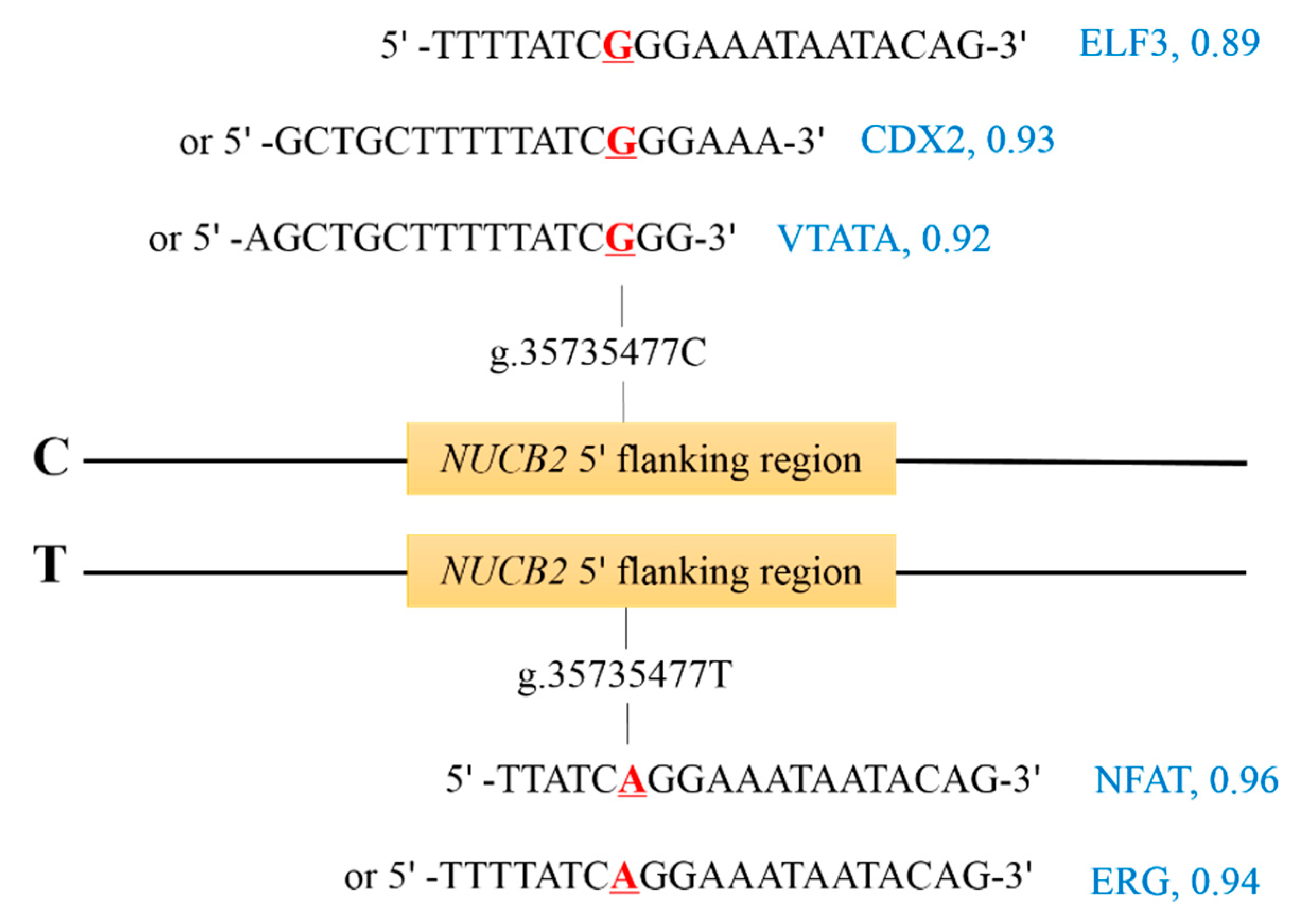
Figure 4.
Expression of NUCB2 gene in bovine tissues. Relative mRNA expression of NUCB2 in eight tissues of lactating Holstein cows normalized by two reference genes, RPS9 and UXT.
Figure 4.
Expression of NUCB2 gene in bovine tissues. Relative mRNA expression of NUCB2 in eight tissues of lactating Holstein cows normalized by two reference genes, RPS9 and UXT.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Details about the identified SNPs of NUCB2.
| SNP Name | GenBank No. | Location | Position (UMD3.1) | Mutation |
|---|---|---|---|---|
| g.35735477C>T | rs135882628 | 5′flanking region | chr15:35735477 | C/T |
| c.*1253A>G | rs207689152 | 3′UTR | chr15:35693831 | A/G |
| c.*1276C>T | rs208861167 | 3′UTR | chr15:35693399 | C/T |
| g.35692766A>G | rs109930800 | 3′flanking region | chr15:35692766 | A/G |
| g.35692674A>G | rs109930800 | 3′flanking region | chr15:35692674 | A/G |
| g.35692210T>C | rs208420141 | 3′flanking region | chr15:35692210 | T/C |
| g.35692155T>C | rs210564196 | 3′flanking region | chr15:35692155 | T/C |
| g.35692145C>T | rs209421884 | 3′flanking region | chr15:35692145 | C/T |
| g.35691328T>C | rs210746263 | 3′flanking region | chr15:35691328 | T/C |
| g.35691371A>G | rs207595426 | 3′flanking region | chr15:35691371 | A/G |
Note: UTR: untranslated region. The “*” shows the SNPs located in the 3’ UTR.
Table 2.
Associations of ten SNPs of NUCB2 gene with milk production traits in two lactations in Chinese Holstein (LSM ± SE).
Table 2.
Associations of ten SNPs of NUCB2 gene with milk production traits in two lactations in Chinese Holstein (LSM ± SE).
| SNP | Lactation | Genotype (No.) | Milk Yield (kg) | Fat Yield (kg) | Fat Percentage (%) | Protein Yield (kg) | Protein Percentage (%) | No. of Significant SNPs |
|---|---|---|---|---|---|---|---|---|
| g.35735477C>T | 1 | CC (604) | 10326 ± 63.16 | 343.09 ± 2.81 | 3.33 ± 0.03 Aa | 306 ± 2.05 | 2.96 ± 0.01 a | 2 |
| CT (376) | 10309 ± 68.49 | 340.27 ± 3.02 | 3.31 ± 0.03 ab | 305.52 ± 2.2 | 2.96 ± 0.01 a | |||
| TT (65) | 10456 ± 111.13 | 338.57 ± 4.65 | 3.21 ± 0.05 Bb | 306.19 ± 3.39 | 2.92 ± 0.02 b | |||
| p value | 0.3565 | 0.285 | 0.0122 | 0.9428 | 0.0327 | |||
| 2 | CC (415) | 10617 ± 67.14 Aa | 385.18 ± 2.99 | 3.65 ± 0.03 a | 315.66 ± 2.18 b | 2.99 ± 0.01 Aa | 4 | |
| CT (258) | 10871 ± 72.95 B | 387.94 ± 3.21 | 3.59 ± 0.03 b | 320.54 ± 2.33 a | 2.96 ± 0.01 Bb | |||
| TT (49) | 10475 ± 126.69 Aa | 383.05 ± 5.25 | 3.67 ± 0.05 ab | 309.98 ± 3.83b | 2.97 ± 0.02 ab | |||
| p value | <0.0001 | 0.4542 | 0.0257 | 0.0036 | 0.0107 | |||
| c.*1253A>G | 1 | AA (622) | 10377 ± 62.96 | 346.91 ± 2.81 a | 3.35 ± 0.03 a | 308.01 ± 2.05 | 2.96 ± 0.01 a | 3 |
| AG (358) | 10330 ± 67.85 | 343.52 ± 2.99 ab | 3.33 ± 0.03 ab | 306.64 ± 2.17 | 2.96 ± 0.01 a | |||
| GG (60) | 10411 ± 113.36 | 335.92 ± 4.73 b | 3.23 ± 0.05 b | 304.51 ± 3.44 | 2.92 ± 0.02 b | |||
| p value | 0.5823 | 0.0178 | 0.0167 | 0.4162 | 0.0334 | |||
| 2 | AA (434) | 10739 ± 65.59 Aa | 389.07 ± 3.02 a | 3.63 ± 0.03 | 319.03 ± 2.12 a | 2.98 ± 0.01 | 3 | |
| AG (248) | 10934 ± 73.57 B | 396.99 ± 3.26 b | 3.6 ± 0.03 | 322.97 ± 2.34 Aa | 2.96 ± 0.01 | |||
| GG (45) | 10468 ± 130.21 Aa | 386.53 ± 5.54 ab | 3.66 ± 0.05 | 308.67 ± 3.93 Bb | 2.96 ± 0.02 | |||
| p value | 0.0002 | 0.0073 | 0.3202 | 0.0006 | 0.089 | |||
| c.*1276C>T | 1 | CC (632) | 10332 ± 62.27 | 346.1 ± 2.78 | 3.36 ± 0.03 a | 305.38 ± 2.03 | 2.96 ± 0.01 a | 3 |
| CT (346) | 10320 ± 68.73 | 343.57 ± 3.02 | 3.34 ± 0.03 ab | 305.67 ± 2.2 | 2.96 ± 0.01 a | |||
| TT (55) | 10392 ± 113.98 | 336.79 ± 4.74 | 3.25 ± 0.05 b | 302.85 ± 3.46 | 2.92 ± 0.02 b | |||
| p value | 0.7939 | 0.0661 | 0.0278 | 0.6665 | 0.0217 | |||
| 2 | CC (442) | 10687 ± 65.62 a | 386.11 ± 2.93 | 3.63 ± 0.03 | 316.61 ± 2.13 ab | 2.98 ± 0.01 a | 3 | |
| CT (238) | 10850 ± 74 b | 388.08 ± 3.23 | 3.59 ± 0.03 | 319.51 ± 2.36 a | 2.96 ± 0.01 b | |||
| TT (42) | 10521 ± 130.66 a | 384.93±5.4 | 3.67 ± 0.05 | 309.97 ± 3.94 b | 2.95 ± 0.02 ab | |||
| p value | 0.0089 | 0.7061 | 0.1478 | 0.0347 | 0.0357 | |||
| g.35692766A>G | 1 | AA (626) | 10418 ± 63.52 | 347.12±2.84 | 3.35 ± 0.03a | 307.67 ± 2.07 | 2.96 ± 0.01 | 1 |
| GA (363) | 10345 ± 68.1 | 343.28±3 | 3.33 ± 0.03ab | 305.79 ± 2.19 | 2.96 ± 0.01 | |||
| GG (62) | 10510 ± 112.05 | 339.43±4.68 | 3.24 ± 0.05b | 306.98 ± 3.41 | 2.92 ± 0.02 | |||
| p value | 0.1772 | 0.0623 | 0.0382 | 0.4738 | 0.0638 | |||
| 2 | AA (434) | 10713 ± 65.44 Aa | 386.64±2.91 | 3.63 ± 0.03 | 318.28 ± 2.12 b | 2.98 ± 0.01 a | 4 | |
| GA (249) | 10930 ± 72.31 B | 392.91±3.16 | 3.61 ± 0.03 | 323.49 ± 2.3 Aa | 2.96 ± 0.01 ab | |||
| GG (46) | 10498 ± 130.83 Aa | 381.61±5.43 | 3.66 ± 0.05 | 308.07 ± 3.96 Ba | 2.94 ± 0.02 b | |||
| p value | 0.0002 | 0.018 | 0.4651 | <0.0001 | 0.0467 | |||
| g.35692674A>G | 1 | CC (60) | 10345 ± 113.56 | 332.97±4.74Aa | 3.23 ± 0.05a | 301.01 ± 3.45 | 2.92 ± 0.02 a | 3 |
| CT (358) | 10330 ± 67.9 | 343.02±2.99ab | 3.33 ± 0.03ab | 305.25 ± 2.18 | 2.96 ± 0.01 b | |||
| TT (622) | 10348 ± 62.26 | 345.78±2.78Bb | 3.36 ± 0.03b | 305.7 ± 2.02 | 2.96 ± 0.01 b | |||
| p value | 0.9433 | 0.0081 | 0.0116 | 0.3135 | 0.0273 | |||
| 2 | CC (45) | 10599 ± 129.91 b | 389.01±5.38ab | 3.68 ± 0.05 | 313.59 ± 3.92 Aa | 2.96 ± 0.02 | 3 | |
| CT (248) | 10973 ± 73.35 Aa | 395.5±3.21a | 3.61 ± 0.03 | 324.92 ± 2.34 B | 2.96 ± 0.01 | |||
| TT (434) | 10733 ± 65.85 Bb | 388.86±2.93b | 3.63 ± 0.03 | 318.92 ± 2.13 Aa | 2.98 ± 0.01 | |||
| p value | 0.0002 | 0.0383 | 0.3137 | 0.0009 | 0.1688 | |||
| g.35692210T>C | 1 | CC (59) | 10441 ± 113.16 | 337.01 ± 4.71 | 3.24 ± 0.05 | 305.33 ± 3.43 | 2.93 ± 0.02 | 0 |
| CT (372) | 10325 ± 68.18 | 340.45 ± 3.01 | 3.31 ± 0.03 | 305.34 ± 2.19 | 2.96 ± 0.01 | |||
| TT (625) | 10373 ± 63.73 | 344.35 ± 2.85 | 3.34 ± 0.03 | 306.62 ± 2.08 | 2.96 ± 0.01 | |||
| p value | 0.4509 | 0.0683 | 0.0533 | 0.6822 | 0.0824 | |||
| 2 | CC (44) | 10443 ± 132.87 Aa | 377.54 ± 5.51 a | 3.64 ± 0.05 | 307.55 ± 4.02 Aa | 2.95 ± 0.02 ab | 4 | |
| CT (254) | 10916 ± 73.07 B | 388.6 ± 3.21 b | 3.58 ± 0.03 | 321.67 ± 2.33 Bb | 2.96 ± 0.01 a | |||
| TT (434) | 10688 ± 66.14 Aa | 383.07 ± 2.95 a | 3.61 ± 0.03 | 316.51 ± 2.15 a | 2.98 ± 0.01b | |||
| p value | <0.0001 | 0.0331 | 0.3355 | 0.0003 | 0.0143 | |||
| g.35692155T>C | 1 | CC (620) | 10297 ± 63.03 | 344.52 ± 2.83 Aa | 3.35 ± 0.03 a | 304.35 ± 2.05 | 2.96 ± 0.01 a | 3 |
| TC (362) | 10284 ± 67.63 | 341.51 ± 3 a | 3.32 ± 0.03 ab | 304.36 ± 2.17 | 2.96 ± 0.01 a | |||
| TT (58) | 10378 ± 114.2 | 328.5 ± 4.92 Bb | 3.24 ± 0.05 b | 302.2 ± 3.46 | 2.92 ± 0.02 B | |||
| p value | 0.6782 | 0.0012 | 0.0144 | 0.7741 | 0.0138 | |||
| 2 | CC (432) | 10690 ± 65.94 Aa | 386.06 ± 2.94 | 3.63 ± 0.03 | 317.22 ± 2.14 ab | 2.98 ± 0.01 | 2 | |
| TC (249) | 10889 ± 73.19 B | 390.76 ± 3.2 | 3.6 ± 0.03 | 321.46 ± 2.33 Aa | 2.96 ± 0.01 | |||
| TT (44) | 10474 ± 131.19 Aa | 382.27 ± 5.43 | 3.66 ± 0.05 | 308.69 ± 3.96 Bb | 2.95 ± 0.02 | |||
| p value | 0.0006 | 0.109 | 0.3315 | 0.0018 | 0.0989 | |||
| g.35692145C>T | 1 | CC (57) | 10447 ± 113.95 | 338.63 ± 4.74 | 3.26 ± 0.05 a | 303.9 ± 3.45 | 2.91 ± 0.02 a | 2 |
| TC (359) | 10342 ± 68.09 | 343.23 ± 3 | 3.33 ± 0.03 ab | 305.98 ± 2.18 | 2.96 ± 0.01 b | |||
| TT (627) | 10366 ± 63.35 | 346.42 ± 2.83 | 3.36 ± 0.03 b | 306.41 ± 2.06 | 2.96 ± 0.01 b | |||
| p value | 0.6061 | 0.0937 | 0.0388 | 0.7116 | 0.0133 | |||
| 2 | CC (43) | 10461 ± 131.5 Aa | 383.01 ± 5.44ab | 3.67 ± 0.05 | 309.35 ± 3.97 Aa | 2.95 ± 0.02 ab | 3 | |
| TC (247) | 10948 ± 72.69 B | 392.58 ± 3.18a | 3.59 ± 0.03 | 323.82 ± 2.31 Bb | 2.96 ± 0.01 a | |||
| TT (439) | 10692 ± 65.5 Aa | 386.85 ± 2.92b | 3.64 ± 0.03 | 318.16 ± 2.13 a | 2.99 ± 0.01 b | |||
| p value | <0.0001 | 0.0466 | 0.1716 | 0.0002 | 0.0185 | |||
| g.35691328T>C | 1 | AA (626) | 10310 ± 63.52 | 347.08 ± 2.81Aa | 3.36 ± 0.03 a | 304.24 ± 2.07 | 2.96 ± 0.01 a | 3 |
| AG (375) | 10289 ± 68.03 | 344.2 ± 2.97a | 3.33 ± 0.03 ab | 303.88 ± 2.18 | 2.96 ± 0.01 a | |||
| GG (59) | 10389 ± 112.86 | 331.3 ± 4.95Bb | 3.25 ± 0.05 b | 302.57 ± 3.42 | 2.92 ± 0.02 b | |||
| p value | 0.6345 | 0.0014 | 0.0248 | 0.8566 | 0.0302 | |||
| 2 | AA (436) | 10631 ± 65.52 Aa | 384.07 ± 2.92 | 3.64 ± 0.03 | 316.66 ± 2.12 Aa | 2.99 ± 0.01 a | 3 | |
| AG (254) | 10910 ± 72.91 B | 388.98 ± 3.2 | 3.58 ± 0.03 | 322.6 ± 2.33 B | 2.96 ± 0.01 b | |||
| GG (44) | 10395 ± 131.93 Aa | 378.08 ± 5.47 | 3.66 ± 0.05 | 307.3 ± 3.99 Ab | 2.96 ± 0.02 ab | |||
| p value | <.0001 | 0.0505 | 0.0698 | <.0001 | 0.0175 | |||
| g.35691371A>G | 1 | AA (604) | 10378 ± 62.51 | 345.22 ± 2.79 Aa | 3.34 ± 0.03 Aa | 306.3 ± 2.03 | 2.95 ± 0.01 Aa | 3 |
| AG (343) | 10336 ± 68.31 | 341.23 ± 3.01 ab | 3.31 ± 0.03 a | 305.12 ± 2.19 | 2.95 ± 0.01 a | |||
| GG (59) | 10417 ± 112.77 | 332.6 ± 4.7 Bb | 3.2 ± 0.05B b | 302.27 ± 3.43 | 2.91 ± 0.02 Bb | |||
| p value | 0.6244 | 0.0048 | 0.0035 | 0.3689 | 0.0075 | |||
| 2 | AA (435) | 10755 ± 66.43 b | 388.73 ± 2.96 | 3.63 ± 0.03 | 320.29 ± 2.16 ab | 2.98 ± 0.01 | 2 | |
| AG (246) | 10938 ± 73.48 Aa | 391.8 ± 3.22 | 3.59 ± 0.03 | 324.21 ± 2.34 Aa | 2.96 ± 0.01 | |||
| GG (45) | 10506 ± 130.52 Bb | 385.76 ± 5.41 | 3.68 ± 0.05 | 311.74 ± 3.94Bb | 2.96 ± 0.02 | |||
| p value | 0.0007 | 0.359 | 0.1113 | 0.0026 | 0.1356 | |||
| No. of significant SNPs | 1 | 0 | 5 | 9 | 0 | 8 | ||
| 2 | 10 | 5 | 1 | 10 | 6 |
Note: The number in the bracket represents the number of cows for the corresponding genotype; p value shows the significance for the genetic effects of SNPs; a, b within the same column with different superscripts means p value < 0.05; A, B within the same column with different superscripts means p value < 0.01.
Table 3.
Associations of haplotype blocks with milk production traits in two lactations in Chinese Holstein (LSM ± SE).
Table 3.
Associations of haplotype blocks with milk production traits in two lactations in Chinese Holstein (LSM ± SE).
| Lactation | Haplotype Combination (No.) | Milk Yield (kg) | Fat Yield (kg) | Fat Percentage (%) | Protein Yield (kg) | Protein Percentage (%) |
|---|---|---|---|---|---|---|
| 1 | H1H1(603) | 10420 ± 65.07 | 347.91 ± 2.9 | 3.35 ± 0.03 | 309.3 ± 2.11 | 2.96 ± 0.01 a |
| H1H3(350) | 10363 ± 70.17 | 343.55 ± 3.09 | 3.33 ± 0.03 | 307.37 ± 2.25 | 2.96 ± 0.01 a | |
| H3H3(59) | 10495 ± 114.84 | 341.13 ± 4.79 | 3.26 ± 0.05 | 307.19 ± 3.49 | 2.93 ± 0.02 b | |
| p value | 0.358 | 0.066 | 0.0819 | 0.4332 | 0.0393 | |
| 2 | H1H1(416) | 10654 ± 67.2 A | 384.36 ± 3 | 3.63 ± 0.03a | 316.81 ± 2.18 Aa | 2.98 ± 0.01 a |
| H1H3(238) | 10875 ± 74.21 B | 386.56 ± 3.24 | 3.57 ± 0.03b | 321.19 ± 2.36 ab | 2.96 ± 0.01 b | |
| H3H3(44) | 10416 ± 133.65 A | 379.5 ± 5.54 | 3.66 ± 0.05ab | 307.95 ± 4.04 Bb | 2.96 ± 0.02 ab | |
| p value | 0.0002 | 0.3786 | 0.047 | 0.0015 | 0.0386 |
Note: H means haplotype; the number in the bracket represents the number of cows for the corresponding haplotype combination; H1 (TACTTAACAC), H2 (TACTTAACAT), H3 (CGTCCGGTGT); p value shows the significance for genetic effects among the haplotype blocks; a, b within the same column with different superscripts means p value < 0.05; A, B within the same column with different superscripts means p value < 0.01.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Han, B.; Yuan, Y.; Li, Y.; Liu, L.; Sun, D. Single Nucleotide Polymorphisms of NUCB2 and Their Genetic Associations with Milk Production Traits in Dairy Cows. Genes 2019, 10, 449. https://doi.org/10.3390/genes10060449
AMA Style
Han B, Yuan Y, Li Y, Liu L, Sun D. Single Nucleotide Polymorphisms of NUCB2 and Their Genetic Associations with Milk Production Traits in Dairy Cows. Genes. 2019; 10(6):449. https://doi.org/10.3390/genes10060449
Chicago/Turabian StyleHan, Bo, Yuwei Yuan, Yanhua Li, Lin Liu, and Dongxiao Sun. 2019. "Single Nucleotide Polymorphisms of NUCB2 and Their Genetic Associations with Milk Production Traits in Dairy Cows" Genes 10, no. 6: 449. https://doi.org/10.3390/genes10060449
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.