Reference Expression Profile of Three FBN1 Transcript Isoforms and Their Association with Clinical Variability in Marfan Syndrome

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient and Control Samples
2.2. Cell Culture and RNA Purification
2.3. Real-Time Quantitative PCR (RT-qPCR)
2.4. Statistical Analysis
3. Results
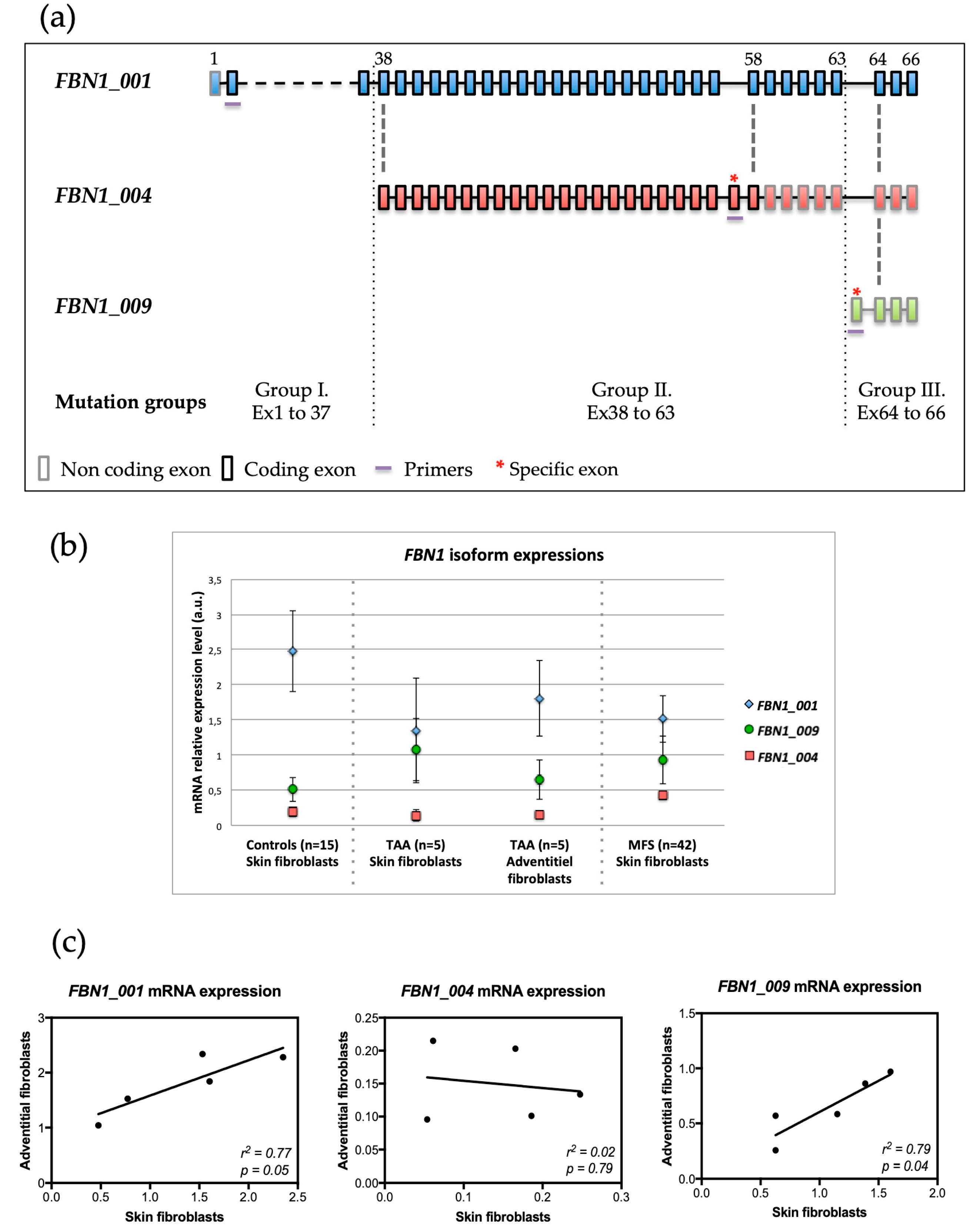
3.1. Expression of FBN1 Isoforms in Skin Fibroblasts from Controls
3.2. Well-Correlated Expression of FBN1 between Skin and Aortic Adventitial Fibroblasts
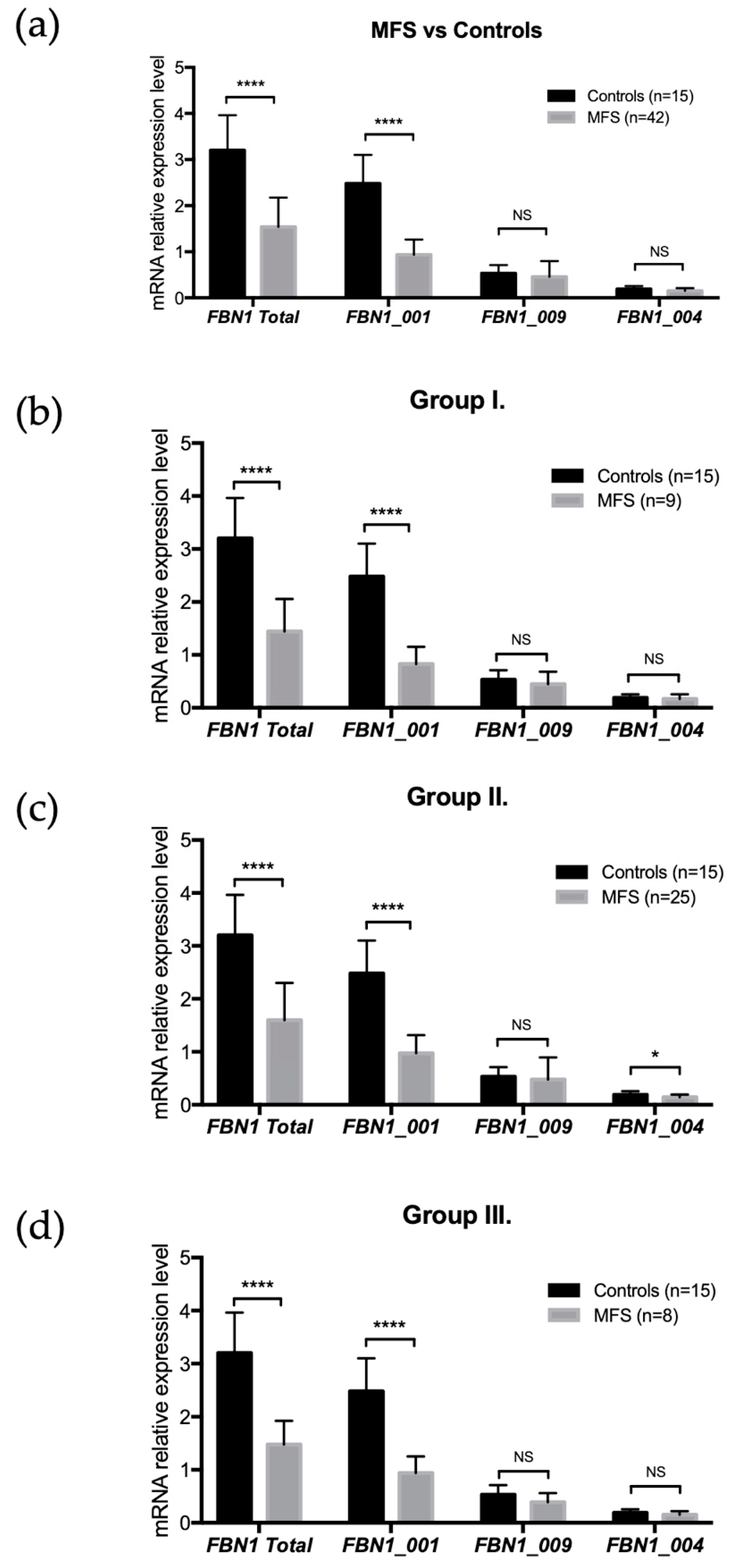
3.3. Expression of FBN1 Isoforms in Skin Fibroblasts from MFS Patients
3.4. FBN1 Isoform Expressions and FBN1 Mutation Locations
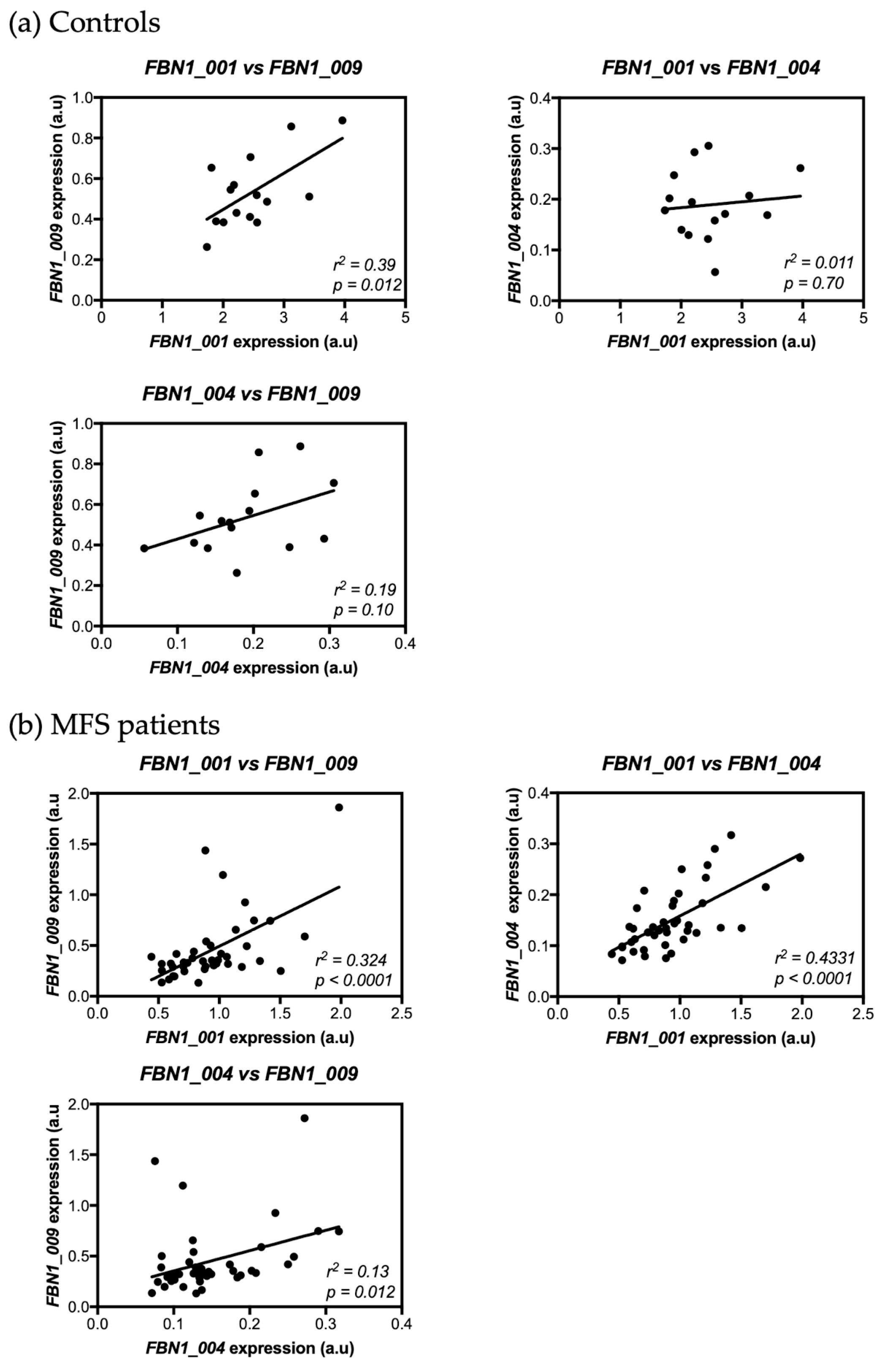
3.5. Correlation between FBN1 Isoform Expressions in Skin Fibroblasts from Controls and MFS Patients
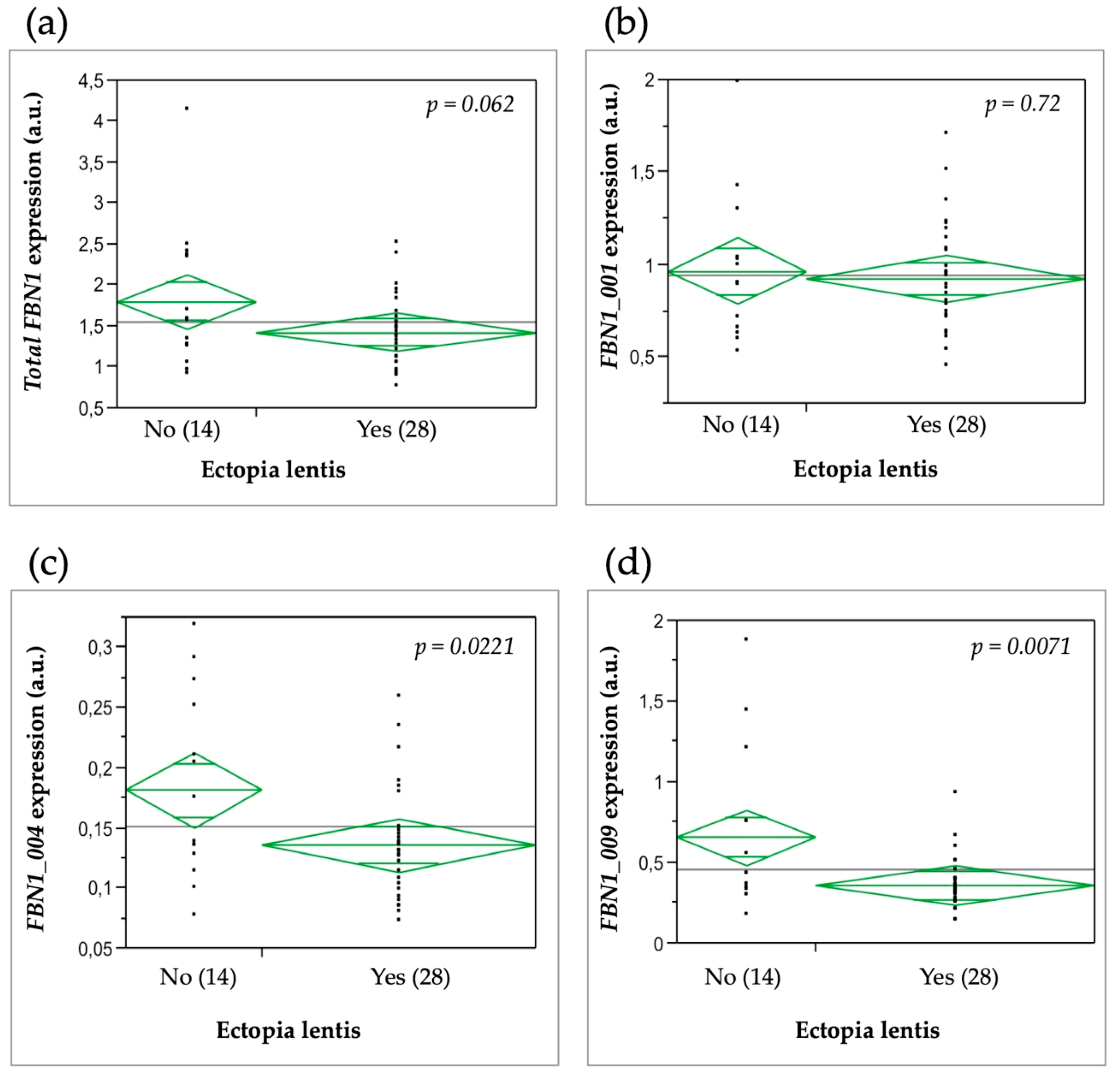
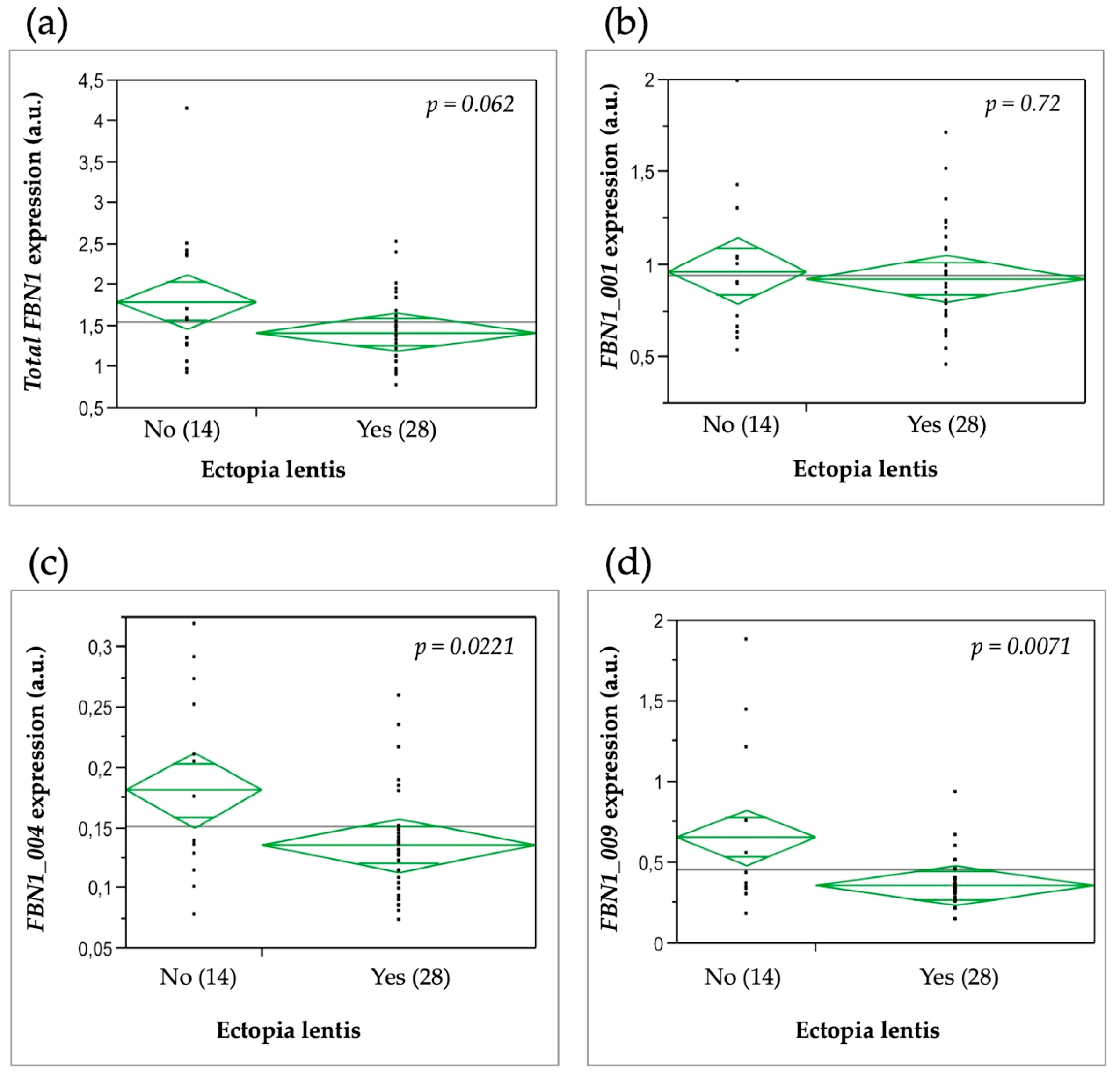
3.6. Relationship between FBN1 Isoform Expression and the Clinical Severity of MFS Features
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Collod-Béroud, G.; Béroud, C.; Adès, L.; Black, C.; Boxer, M.; Brock, D.J.; Godfrey, M.; Hayward, C.; Karttunen, L.; Milewicz, D.; et al. Marfan Database (second edition): software and database for the analysis of mutations in the human FBN1 gene. Nucleic Acids Res. 1997, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Loeys, B.L.; Dietz, H.C.; Braverman, A.C.; Callewaert, B.L.; De Backer, J.; Devereux, R.B.; Hilhorst-Hofstee, Y.; Jondeau, G.; Faivre, L.; Milewicz, D.M.; et al. The revised Ghent nosology for the Marfan syndrome. J. Med. Genet. 2010, 47, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Baudhuin, L.M.; Kotzer, K.E.; Lagerstedt, S.A. Increased frequency of FBN1 truncating and splicing variants in Marfan syndrome patients with aortic events. Genet. Med. 2015, 17, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Muñoz, V.M.; Gómez-Doblas, J.J.; Porras-Martín, C.; Such-Martínez, M.; Crespo-Leiro, M.G.; Barriales-Villa, R.; de Teresa-Galván, E.; Jiménez-Navarro, M.; Cabrera-Bueno, F. The importance of genotype-phenotype correlation in the clinical management of Marfan syndrome. Orphanet J. Rare Dis. 2018, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- Faivre, L.; Collod-Beroud, G.; Loeys, B.L.; Child, A.; Binquet, C.; Gautier, E.; Callewaert, B.; Arbustini, E.; Mayer, K.; Arslan-Kirchner, M.; et al. Effect of mutation type and location on clinical outcome in 1,013 probands with Marfan syndrome or related phenotypes and FBN1 mutations: An international study. Am. J. Hum. Genet. 2007, 81, 454–466. [Google Scholar] [CrossRef] [PubMed]
- Franken, R.; Groenink, M.; de Waard, V.; Feenstra, H.M.A.; Scholte, A.J.; van den Berg, M.P.; Pals, G.; Zwinderman, A.H.; Timmermans, J.; Mulder, B.J.M. Genotype impacts survival in Marfan syndrome. Eur. Heart J. 2016, 37, 3285–3290. [Google Scholar] [CrossRef] [PubMed]
- Franken, R.; Teixido-Tura, G.; Brion, M.; Forteza, A.; Rodriguez-Palomares, J.; Gutierrez, L.; Garcia Dorado, D.; Pals, G.; Mulder, B.J.; Evangelista, A. Relationship between fibrillin-1 genotype and severity of cardiovascular involvement in Marfan syndrome. Heart 2017, 103, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- Rommel, K.; Karck, M.; Haverich, A.; von Kodolitsch, Y.; Rybczynski, M.; Müller, G.; Singh, K.K.; Schmidtke, J.; Arslan-Kirchner, M. Identification of 29 novel and nine recurrent fibrillin-1 (FBN1) mutations and genotype-phenotype correlations in 76 patients with Marfan syndrome. Hum. Mutat. 2005, 26, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Schrijver, I.; Liu, W.; Odom, R.; Brenn, T.; Oefner, P.; Furthmayr, H.; Francke, U. Premature termination mutations in FBN1: Distinct effects on differential allelic expression and on protein and clinical phenotypes. Am. J. Hum. Genet. 2002, 71, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Takeda, N.; Inuzuka, R.; Maemura, S.; Morita, H.; Nawata, K.; Fujita, D.; Taniguchi, Y.; Yamauchi, H.; Yagi, H.; Kato, M.; et al. Impact of pathogenic FBN1 variant types on the progression of aortic disease in patients with Marfan syndrome. Circ. Genom. Precis. Med. 2018, 11, e002058. [Google Scholar] [CrossRef] [PubMed]
- Aubart, M.; Gazal, S.; Arnaud, P.; Benarroch, L.; Gross, M.-S.; Buratti, J.; Boland, A.; Meyer, V.; Zouali, H.; Hanna, N.; et al. Association of modifiers and other genetic factors explain Marfan syndrome clinical variability. Eur. J. Hum. Genet. 2018. [Google Scholar] [CrossRef] [PubMed]
- Aubart, M.; Gross, M.-S.; Hanna, N.; Zabot, M.-T.; Sznajder, M.; Detaint, D.; Gouya, L.; Jondeau, G.; Boileau, C.; Stheneur, C. The clinical presentation of Marfan syndrome is modulated by expression of wild-type FBN1 allele. Hum. Mol. Genet. 2015, 24, 2764–2770. [Google Scholar] [CrossRef] [PubMed]
- Burchett, M.E.; Ling, I.-F.; Estus, S. FBN1 isoform expression varies in a tissue and development-specific fashion. Biochem. Biophys. Res. Commun. 2011, 411, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, L.; Aubart, M.; Gross, M.-S.; Jacob, M.-P.; Arnaud, P.; Hanna, N.; Jondeau, G.; Boileau, C. Marfan syndrome variability: Investigation of the roles of sarcolipin and calcium as potential transregulator of FBN1 expression. Genes 2018, 9, 421. [Google Scholar] [CrossRef] [PubMed]
- Ensembl Genome Browser 94. Available online: https://www.ensembl.org/index.html (accessed on 7 January 2019).
- GTEx Portal. Available online: https://gtexportal.org/home/ (accessed on 7 January 2019).
- Corson, G.M.; Chalberg, S.C.; Dietz, H.C.; Charbonneau, N.L.; Sakai, L.Y. Fibrillin binds calcium and is coded by cDNAs that reveal a multidomain structure and alternatively spliced exons at the 5′ end. Genomics 1993, 17, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Bauer, S.; Hecht, J.; Schulz, M.H.; Busche, A.; Robinson, P.N. A short ultraconserved sequence drives transcription from an alternate FBN1 promoter. Int. J. Biochem. Cell Biol. 2008, 40, 638–650. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Rödelsperger, C.; Digweed, M.; Robinson, P.N. Regulation of fibrillin-1 gene expression by Sp1. Gene 2013, 527, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Biery, N.J.; Eldadah, Z.A.; Moore, C.S.; Stetten, G.; Spencer, F.; Dietz, H.C. Revised genomic organization of FBN1 and significance for regulated gene expression. Genomics 1999, 56, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Summers, K.M.; Bokil, N.J.; Baisden, J.M.; West, M.J.; Sweet, M.J.; Raggatt, L.J.; Hume, D.A. Experimental and bioinformatic characterisation of the promoter region of the Marfan syndrome gene, FBN1. Genomics 2009, 94, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Lykke-Andersen, S.; Jensen, T.H. Nonsense-mediated mRNA decay: An intricate machinery that shapes transcriptomes. Nat. Rev. Mol. Cell Biol. 2015, 16, 665–677. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Forward | Reverse |
|---|---|---|
| FBN1_001 | AGTCGGGCCAAGAGAAGAGGCG | TCCATCCAGGGCAACAGTAAGCAT |
| FBN1_004 | TTTTACTGCTGTCTCCAGCTTTCC | ACAGCAGCATTCCGATTTGGTG |
| FBN1_009 | AAACTCATGGTTTTCCCCCTTCT | TGATGTCTTGGCATCCTCCAC |
| GAPDH | GTCGCCAGCCGAGCCACATC | CCAGGCGCCCAATACGACCA |
| SDHA | AAGGGCTCCGACTGGCTGGGG | TTTCTAGCTCGACCACGGCGGC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benarroch, L.; Aubart, M.; Gross, M.-S.; Arnaud, P.; Hanna, N.; Jondeau, G.; Boileau, C. Reference Expression Profile of Three FBN1 Transcript Isoforms and Their Association with Clinical Variability in Marfan Syndrome. Genes 2019, 10, 128. https://doi.org/10.3390/genes10020128
Benarroch L, Aubart M, Gross M-S, Arnaud P, Hanna N, Jondeau G, Boileau C. Reference Expression Profile of Three FBN1 Transcript Isoforms and Their Association with Clinical Variability in Marfan Syndrome. Genes. 2019; 10(2):128. https://doi.org/10.3390/genes10020128
Chicago/Turabian StyleBenarroch, Louise, Mélodie Aubart, Marie-Sylvie Gross, Pauline Arnaud, Nadine Hanna, Guillaume Jondeau, and Catherine Boileau. 2019. "Reference Expression Profile of Three FBN1 Transcript Isoforms and Their Association with Clinical Variability in Marfan Syndrome" Genes 10, no. 2: 128. https://doi.org/10.3390/genes10020128
APA StyleBenarroch, L., Aubart, M., Gross, M.-S., Arnaud, P., Hanna, N., Jondeau, G., & Boileau, C. (2019). Reference Expression Profile of Three FBN1 Transcript Isoforms and Their Association with Clinical Variability in Marfan Syndrome. Genes, 10(2), 128. https://doi.org/10.3390/genes10020128