1. Introduction
Mesenchymal stem/stromal cells (MSCs) are highly proliferative fibroblast-like cells that can be isolated from multiple tissue sources and possess potent regenerative, immunomodulatory, neuroprotective and proangiogenic properties [
1]. MSCs have been widely studied to establish reliable and effective cures to numerous diseases, predominantly through the secretion of bioactive factors [
2,
3,
4]. The MSC secretome can include free proteins but also contains extracellular vesicles (EVs), including exosomes, which are nanosized particles that are produced by budding from the endosomal membrane. EVs act as messengers of intercellular communication and can contain bioactive factors, including proteins, lipids and micro-RNAs [
5]. EVs have also been proposed as a mechanism by which MSCs provide therapeutic benefits and have immunoregulatory and neuroprotective properties that are achieved through multiple molecular mechanisms [
5,
6]. Cell-based therapies are limited by potential immune rejection of donor cells and also pose other safety concerns [
7]. Increasingly, studies have shown that MSC survival and integration within the host after transplantation are usually poor and that MSCs exert their therapeutic functions mainly via paracrine signaling mechanisms [
8]. Therefore, MSC-secreted EVs have promising cell-free-based regenerative therapeutic potential; however, the exact molecular mechanism by which MSCs and secreted-EVs exert immunoregulatory, neuroprotective and proangiogenic properties is poorly understood.
MSC-derived EVs have also been shown to readily cross the blood–brain barrier (BBB) and deliver therapeutic cargo to reduce the effects of neuropathological disease, including multiple sclerosis (MS) [
9]. MS is a heterogenous disease that is characterized by demyelination and inflammation caused by immune cell infiltration of the central nervous system (CNS) [
10]. MS results from an autoimmune response within the CNS that can lead to neuronal degradation, inflammation and loss of axonal conductivity and gliosis in grey and white matter [
11]. Myelin sheaths provide a supportive insulating layer for axons and are produced by oligodendrocytes. Oligodendrocyte progenitor cells (OPCs) are an immature cell subset that can differentiate into mature myelinating oligodendrocytes (OLs) [
12]. Loss of oligodendrocytes and accompanying demyelination is associated with progressive axonal degeneration and neurological decline [
13]. Current therapies for MS target the immune component of the disease but do not prevent the progressive axonal and neural degradation.
Cellular-based therapies utilizing MSCs are currently being used in clinical trials for the treatment of adult MS [
11,
14,
15]. The placenta has been suggested to be a unique source of MSCs that possess robust immunomodulatory properties and have been reported to be beneficial in murine models of graft versus host disease [
16,
17]. MSCs derived from the placenta (PMSCs) may be a more appropriate cell source for pediatric diseases because the placenta demonstrates “fetomaternal tolerance” during pregnancy, which is attributed to the expression of human leukocyte antigen-G (HLA-G), a non-classical major histocompatibility complex (MHC) class I molecule that inhibits natural killer cell (NK) killing [
18]. Unlike bone marrow-derived MSCs (BM-MSCs), PMSCs express HLA-G on their surface in response to interferon gamma (IFNγ) [
19], which is a key inflammatory mediator involved with the onset of MS [
10]. Therefore, the expression of HLA-G on PMSCs would make them a unique therapeutic cell source for the treatment of neurodegenerative diseases like MS. Currently, a clinical trial is underway using term PMSCs to treat adult MS and no paradoxical worsening of MS lesion counts has been noted [
20].
Our group has previously demonstrated that early-gestational chorionic villus-derived PMSCs can prevent hind limb paralysis in a surgically created lamb model of spina bifida (SB) [
21]. The neuroprotective properties of PMSCs occur through paracrine signaling mechanisms. Thorough characterization of PMSC-derived EVs has been performed and it was demonstrated that this unique source of EVs has potent neuroprotective properties and contains key proteins and RNAs that contribute to neuronal survival [
22]. The goal of the current study was to determine if PMSC-derived EVs provided regenerative effects in an experimental autoimmune encephalomyelitis (EAE) model of MS and their potential in promoting remyelination. It was hypothesized that PMSC-derived EV treatments would lead to motor function improvement in a dose-dependent manner by preventing oligodendroglia degradation and demyelination associated with MS pathology.
2. Materials and Methods
2.1. PMSC Isolation and Expansion
PMSCs for this study were previously isolated from the chorionic villus tissue of de-identified discarded second trimester human placenta by explant culture [
23]. Early passage (P2-P5) PMSCs were cultured in Dulbecco’s Modified Eagle Medium high glucose (DMEM; GE Life Sciences, Pittsburgh, PA, USA) supplemented with 5% fetal bovine serum (FBS; Atlanta Biologics, Flowery Branch, GA, USA), 20 ng/mL fibroblast growth factor basic (FGF basic; Advent Bio, Elk Grove Village, IL, USA), 20 ng/mL epidermal growth factor (EGF; Advent Bio) and 1% penicillin/streptomycin (P/S; ThermoFisher Scientific, Pittsburgh, PA, USA). Cells were cultured in T150 flasks (Corning Inc., Corning, NY, USA) at 37 °C, under 5% CO
2 as previously described [
23].
2.2. PMSC Phenotype
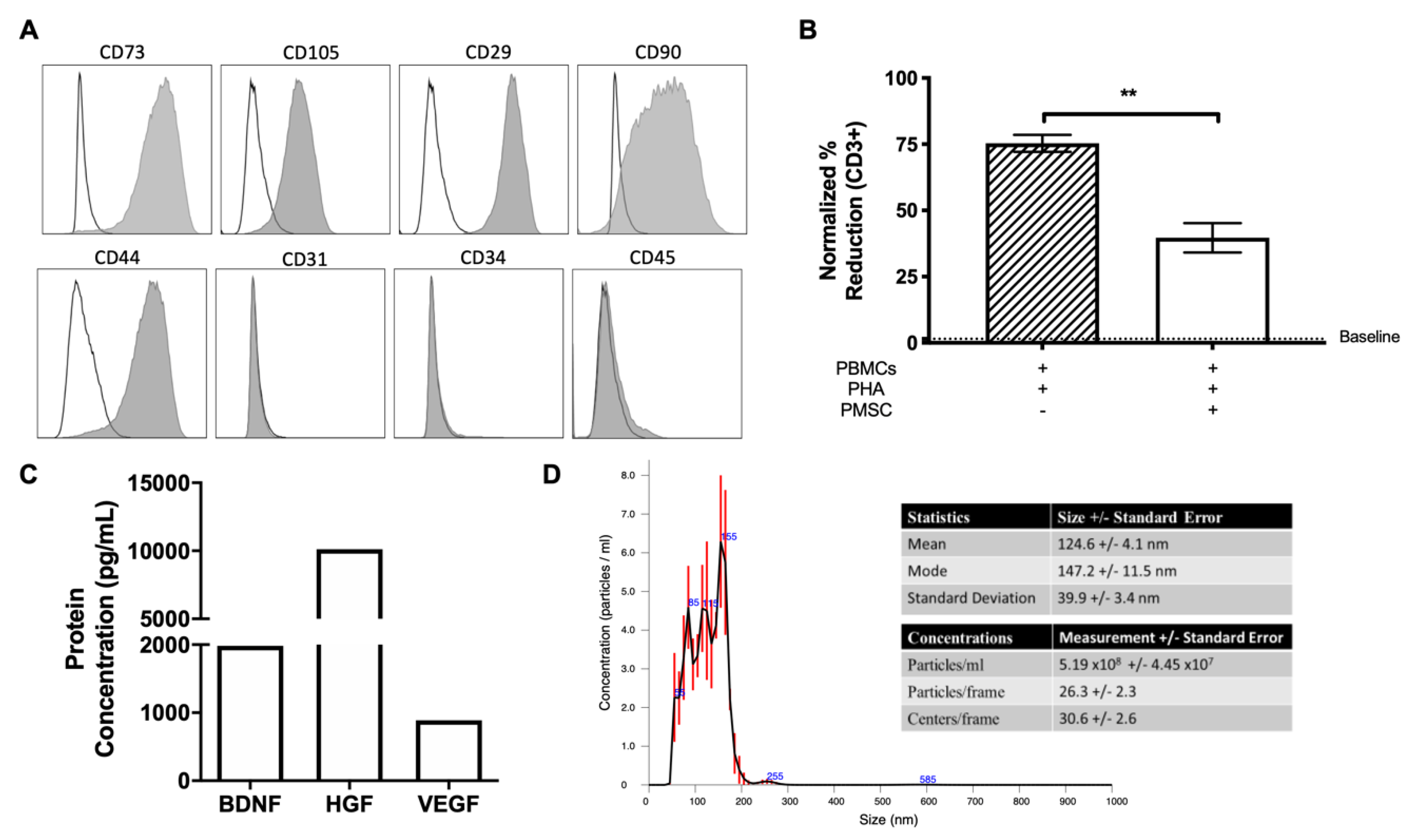
PMSCs were detached using Accutase (Thermo Fisher Scientific) for immunophenotype analysis via flow cytometry. Single suspension PMSCs were first labeled with LIVE/DEAD
® Fixable Aqua Dead Cell Stain Kit (Thermo Fischer Scientific) to detect dead cells. PMSCs were then washed and incubated with antibodies directed against CD44 (clone G44-26), CD90 (clone 5E10), CD73 (cloneAD2), CD29 (clone MAR4), CD34 (clone 563), CD31 (clone WM59), and CD45 (clone HI30). Appropriate isotype controls were used for each antibody as previously described [
23]. All antibodies were purchased from BD Bioscience (San Jose, CA, USA). All the samples were read by flow cytometry (Attune NxT Flow Cytometer, Thermo Fisher Scientific) and analyzed using FlowJo software (Tree Star Inc., Ashland, OR, USA).
2.3. Mixed Leukocyte Reaction (MLR)
To evaluate the immunosuppressive potential of PMSCs, mixed leukocyte reactions (MLRs) were performed exactly as previously described [
24]. Briefly, peripheral blood mononuclear cells (PBMCs) were isolated from whole blood by using an underlay method of Ficoll-Paque
TM PLUS (GE healthcare, Uppsala, Sweden) and gradient centrifugation. PBMCs were collected from the buffy coat layer and stimulated with 5 µg/mL phytohemagglutin (PHA, Millipore-Sigma, St. Louis, MO, USA). PMSCs were irradiated (10Gy, Varian 2100C linear accelerator, Varian Medical Systems Inc., Palo Alto, CA, USA) and kept on ice before experimental setup. PBMCs and PMSCs were co-cultured at a ratio of 5:1 in DMEM supplemented with 10% heat inactivated FBS, 1% P/S and L-tryptophan (600 μM, Millipore-Sigma).
After 3 days of co-culture, samples were treated with 1 mM Bromodeoxyuridine (BrdU, BD Biosciences). Twenty-four hours following BrdU treatment, leukocytes were collected and cells were stained for LIVE/DEAD® Fixable Aqua Dead Cell to identify live cells and CD3 (clone UCHT1, BD Bioscience) to identify lymphocyte populations. Cells were then stained for nuclear BrdU incorporation per manufacture directions (FITC BrdU Flow Kit, BD Biosciences) and read by flow cytometry (Attune NxT Flow Cytometer, Thermo Fisher Scientific). Flow cytometry data were analyzed using FlowJo software (Tree Star Inc.).
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
PMSCs were cultured for 24 h and supernatants were collected for protein quantification via ELISA as previously described [
23]. ELISAs for brain derived neurotrophic factor (BDNF; R&D Systems, Minneapolis, MN, USA), hepatocyte growth factor (HGF; R&D Systems) and vascular endothelial growth factor (VEGF; R&D Systems) were performed per the manufacturer’s instructions and read on a plate reader (SpectraMax Plate Readers, Molecular Devices, San Jose, CA, USA).
2.5. Neuroprotection Assay by Indirect Coculture
Neuroprotection assays were performed exactly as previously described [
22]. In brief, the SH-SY5Y neuroblastoma cell line was cultured for 24 h. PMSCs were indirectly cultured in hanging well inserts for 24 h. To assess the neuroprotective properties of PMSCs, SH-SY5Y cells were treated with 1 µM staurosporine to induce apoptosis. PMSC inserts were washed and co-cultured with apoptotic SH-SY5Y cells. After 96 h, cells were washed with 2 µM calcein AM (Thermo Fisher Scientific) and imaged using an Axio Observer D1 inverted microscope (Carl Zeiss). Images were processed with WimNeuron Image Analysis (Onimagin Technologies, Cordoba, Spain) for neurite outgrowth analysis.
2.6. PMSC-Derived Extracellular Vesicle (EV) Isolation
PMSC-derived EVs were isolated as previously described [
22]. In brief, EVs were first depleted from FBS by spinning FBS samples at 112,700 G using the L7 Ultracentrifuge (Beckman Coulter, Brea, CA, USA) and a SW28 rotor for 16 h at 4 °C. Supernatants were collected, aliquoted and stored at −20 °C. PMSCs at passage 4 were seeded at 20,000 cells/cm
2 in T175 flasks (Corning Inc.) in 20 mL of medium containing 5% EV-depleted FBS, 20 ng/mL FGF (Advent Bio), 20 ng/mL EGF (Advent Bio), and 1% P/S (Thermo Fischer Scientific) for 48 h at 37 °C, under 5% CO
2. Conditioned medium was collected and EVs were isolated by differential centrifugation exactly as previously described [
22]. After the final centrifugation step, EV pellets were resuspended in 10 µL of triple-filtered PBS (GE, Life Sciences) per T175 flask used for the generation of the conditioned medium. EVs were aliquoted and stored at −80 °C.
2.7. EV Characterization by Western Blot
To characterize EVs, a Western blotting analysis was performed exactly as previously described [
22]. In brief, EVs were treated with NuPAGE LDS Sample Buffer (Thermo Fisher Scientific) and heated to 90 °C. The samples were run, transferred, probed with 1:500 dilution of primary antibodies against Alg-2 interacting protein X (ALIX; rabbit polyclonal, Millipore-Sigma), tumor susceptibility gene (TSG101; clone T5701; Millipore-Sigma), CD9 (clone MM2-57; Millipore-Sigma), calnexin (Clone C5C9, Cell Signaling Technology, Danvers, MA, USA), and CD63 (clone TS63; Thermo Fisher Scientific) in 5% nonfat dry milk in 20 mM Tris-HCl (pH 7.4), 150 mM NaCl, and 0.5% Tween 20 (Millipore-Sigma). Blots were then probed against their respective secondary antibodies and developed using Chemidoc MP Imaging System (Bio-Rad, Hercules, CA, USA).
2.8. Characterization of EVs by Nanoparticle Tracking Analysis
Nanoparticle Tracking Analysis (NTA) was performed to quantify EV yield and size distribution. The Nano Sight LM10 Nanoparticle Analysis System (Malvern Panalytical Ltd., Malvern, UK) and the NTA 3.1 Analytical Software (Malvern Panalytical Ltd.) were used to characterize isolated EVs. A 5 µl aliquot of stored EVs was diluted in 995 µl of 0.2 µm triple-filtered Milli-Q water (Millipore-Sigma) and injected into the NTA. A 1:200 dilution is necessary to obtain a concentration between 2 × 10
8 and 20 × 10
8 particle/mL, which is the suggested optimal range for NTA [
25,
26]. Three-30 second videos were recorded and analyzed by the software. The scientific complementary metal-oxide-semiconductor (sCMOS) camera collected 739 frames at 22.0 °C. A 404 nm laser with a detection threshold of 5, determined the mean-square displacement of each nanoparticle based on its own Brownian motion. The analytical software determined EV size, mean, mode and standard deviation, as well as the number of particles per frame and milliliter of sample.
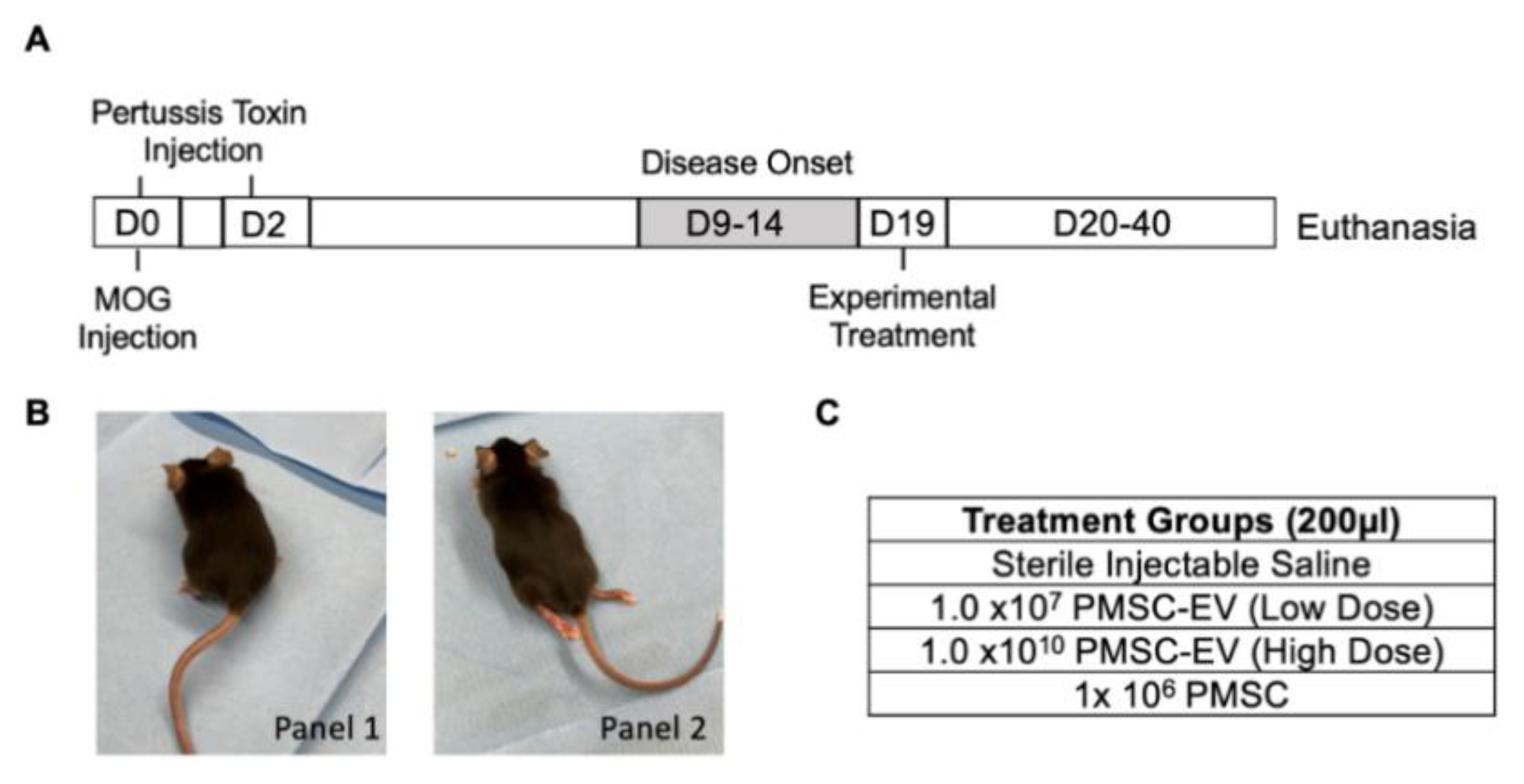
2.9. Experimental Autoimmune Encephalomyelitis (EAE) Induction in a Murine Model
In order to mimic the pathology of human MS, EAE was induced using a murine model as previously described [
27,
28]. Three-month old C57BL/6J mice (male and female) were immunized with myelin oligodendrocyte glycoprotein (MOG) peptide 35–55 to induce EAE (IACUC #19014). In brief, 300 μg of rodent MOG peptide (amino acids 35–55, New England Peptides, Gardner, MA, USA) in Complete Freund’s Adjuvant (CFA) containing 5 mg/mL killed Mycobacterium tuberculosis (Difco, Thermo Fischer Scientific) was administered into the subcutaneous flank of mice at day 0. At day 0, each mouse received two subcutaneous injections of the MOG solution as well as a 100 µl dose of 2 ng/µl pertussis toxins and virulence factors (List Biological Laboratories Inc., Campbell, CA, USA) diluted in sterile PBS (Thermo Fisher Scientific). Pertussis toxins and virulence factors were administered again on day 2. Pertussis toxins allow an increase in the blood–brain barrier permeability in order to facilitate the incursion of the different treatments into the CNS.
In order to monitor disease progression, mice were weighed and scored daily. Neurological deficits were assessed on a five-point scale (limp tail or waddling gait = 1; limp tail and waddling gait = 2; single limb paresis and ataxia = 2.5; double limb paresis = 3; single limb paralysis and paresis of second limb = 3.5; full paralysis of 2 limbs = 4; moribund = 4.5; and death = 5) [
27].
2.10. Experimental Treatment of EAE Animals
Daily motor quantification was performed. Mouse treatment groups were randomized in order to contain comparable numbers of males and females and an average score close to 3.5 to represent EAE onset. For this study, disease onset and treatment occurred on day 19 post-MOG immunization. On the day of treatment, PMSCs were detached from culture using TrypLE (Thermo Fisher Scientific) and washed twice with PBS (Thermo Fisher Scientific). PMSCs with dimensions of 1 × 106 were resuspended in 200 µl injectable saline and placed on ice prior to administration. Stored EVs were thawed and resuspended at either 1 × 107 (low dose) or 1 × 1010 (high dose) EVs in 200 µl of injectable saline.
Tail-vein injections were performed using 1 mL syringes connected to standard hypodermic needles (Covidien, Dublin, Republic of Ireland). EAE mouse scoring was repeated up to day 40 or 43 post-MOG immunization.
2.11. Tissue Preparation
Mice were euthanized by CO2 asphyxiation and were perfused with ice-cold PBS (Thermo Fisher Scientific). Lumbar spinal cords were harvested, post-fixed in 4% paraformaldehyde (PFA) at room temperature for 2 h, cryopreserved in 30% sucrose overnight, and embedded in OCT. Fourteen micrometer frozen transverse sections were cut on a Leica cryostat.
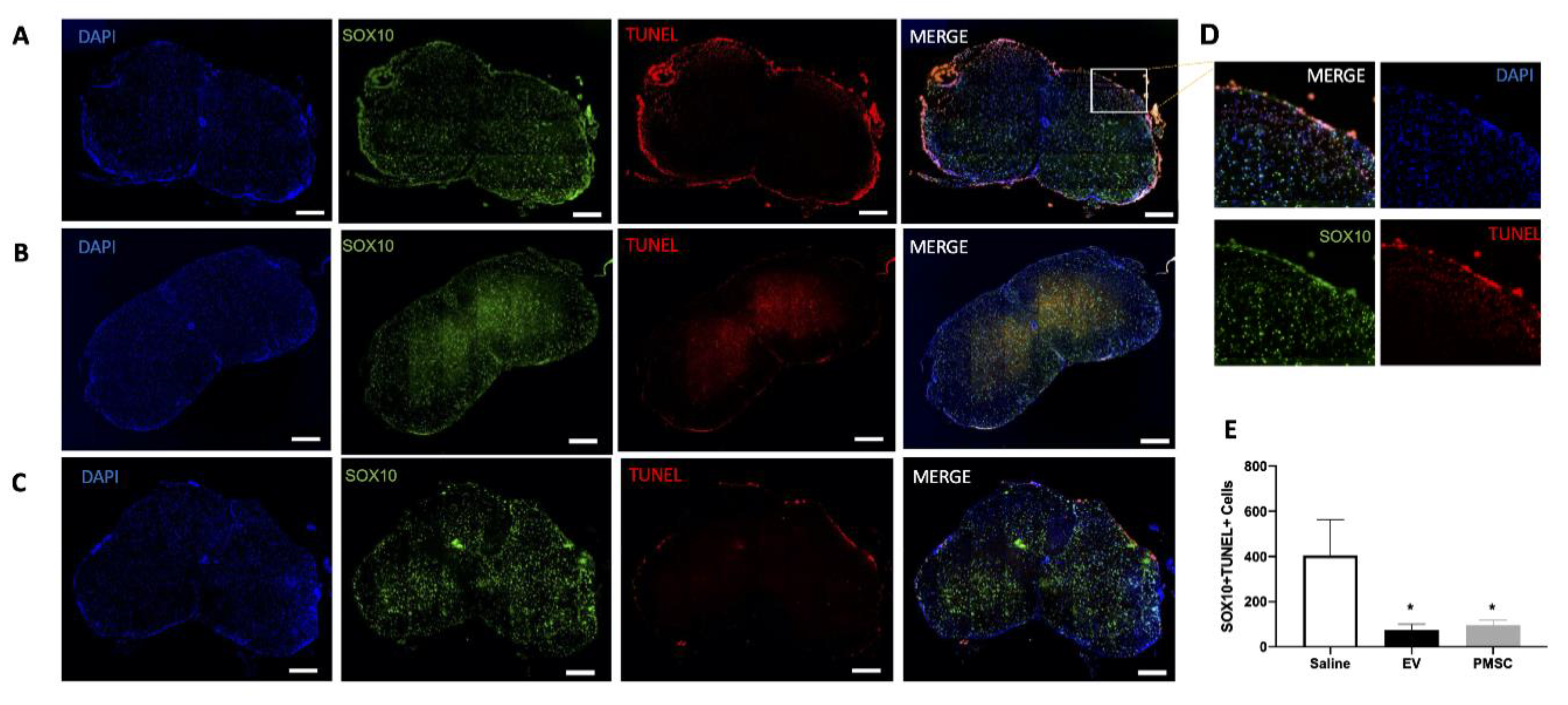
2.12. Quantification of Oligodendroglia Survival
Frozen sections were dried and blocked using PBS (Thermo Fisher Scientific) containing 0.1% Tween 20 (Millipore-Sigma) and 10% donkey serum (Thermo Fisher Scientific) for 1 h at room temperature. Sections were incubated with primary antibodies directed against SOX-10 (clone EP268, Millipore-Sigma) at 4 °C overnight, followed by 2 h incubation at room temperature with secondary antibody. TUNEL staining (Terminal deoxynucleotidyl transferase dUTP nick end labeling) was performed on samples to quantify DNA damaged within SOX10 populations. TUNEL was performed using the In-Situ Cell Death Detection Kit (TMR Red, Millipore-Sigma) per the manufacturer’s instructions. DAPI (Thermo Fisher Scientific) was used to label nuclei, and the sections were mounted with Permount (Thermo Fisher Scientific) for microscopic analysis. Samples were imaged using a Carl Zeiss Axio Observer D1 inverted microscope and analyzed using NIH ImageJ software.
2.13. Quantification of Myelin Loss
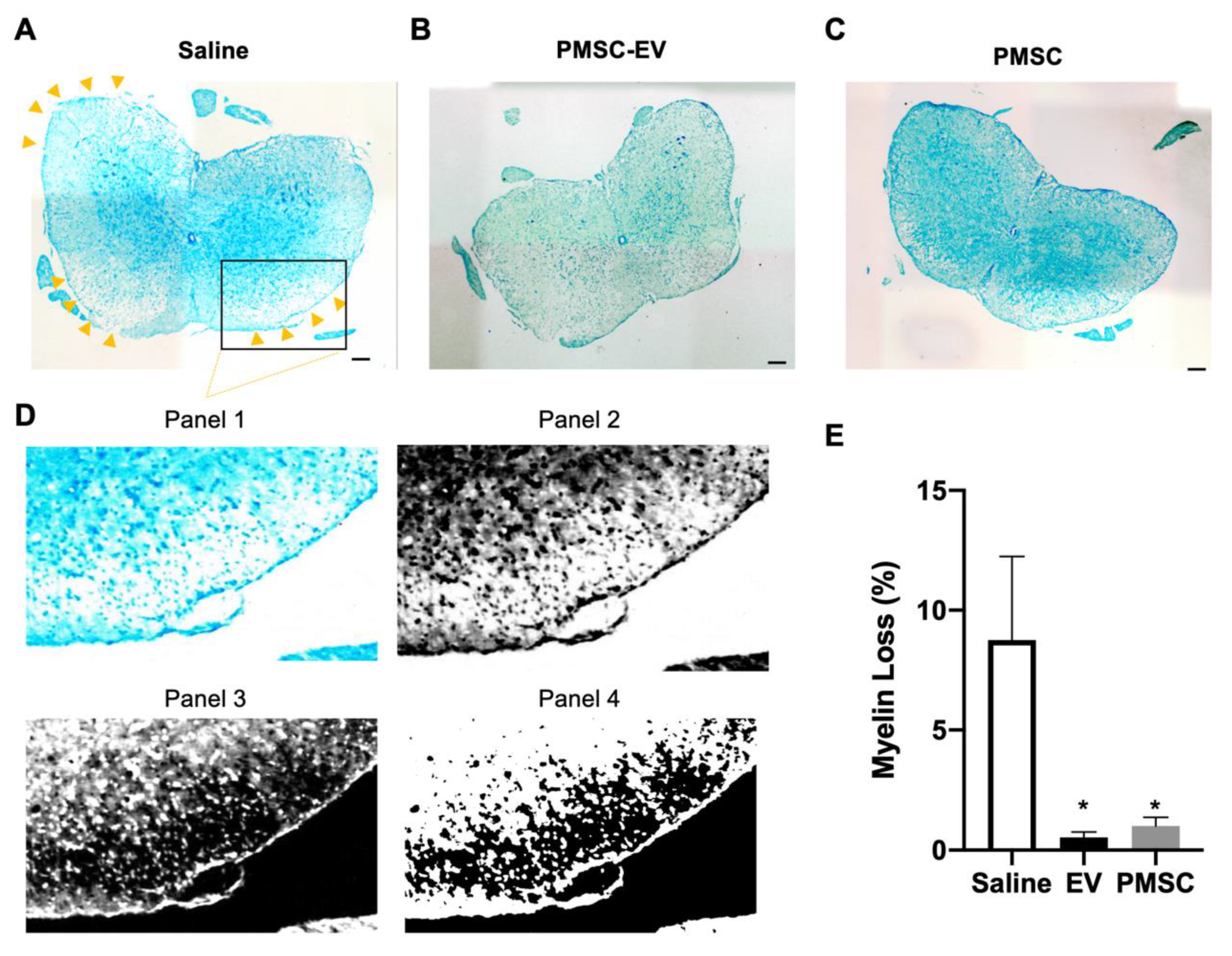
In order to quantify the loss of myelin in treated EAE mice, Luxol Fast Blue (LFB) was used to stain frozen sections. LFB is a copper phthalocyanine dye that binds to lipoproteins found within the myelin sheath. Frozen sections were dried and rehydrated using 95% EtOH. LFB staining was performed according to the manufacturer’s instructions (IHC World, Woodstock, MD, USA). Sections were mounted with Permount (Thermo Fisher Scientific) and samples were imaged using a Carl Zeiss Axio Observer D1 inverted microscope. Images were analyzed using NIH ImageJ software. Myelin stains blue using LFB, therefore, thresholds were standardized and lack of LFB staining was quantified to denote the percentage of myelin loss in each sample.
2.14. EV Functions on Oligodendrocyte Precursor Cell (OPC) Differentiation
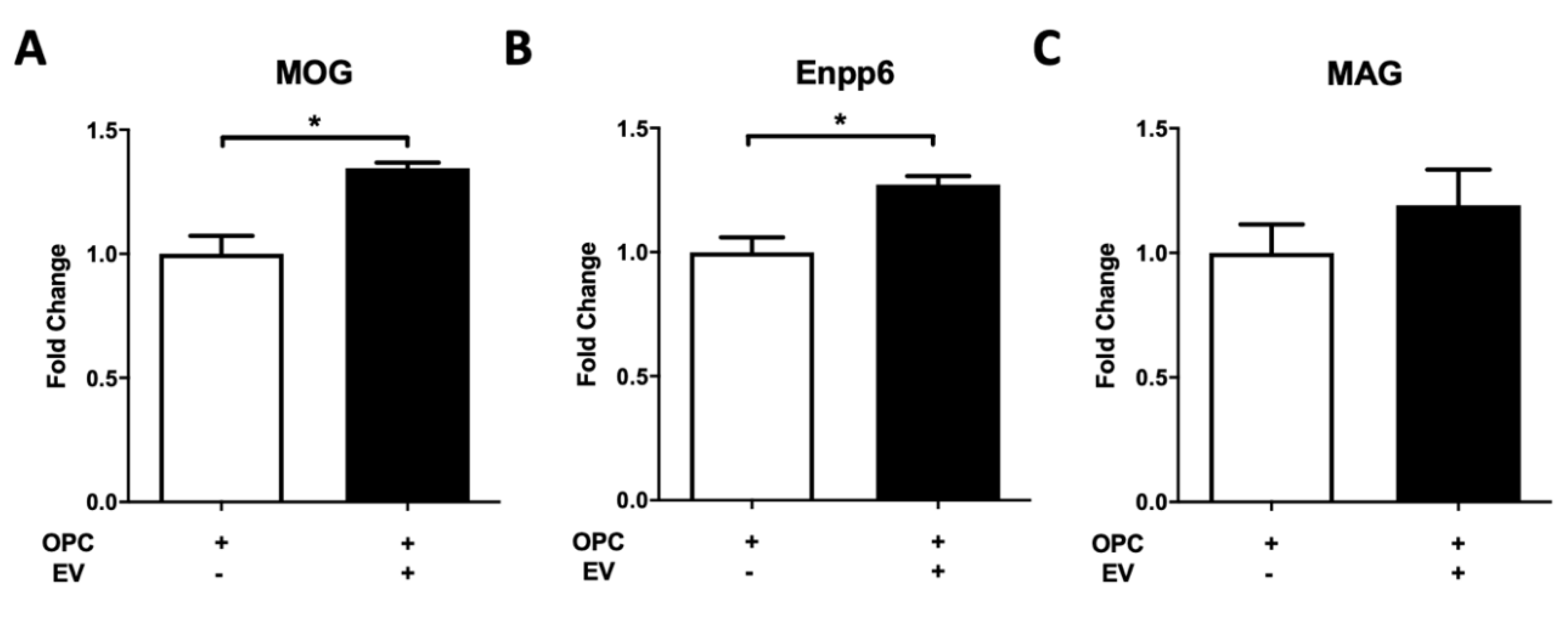
Primary OPC culture and differentiation were conducted according to our previous protocol [
29]. In brief, OPCs were isolated by immunopanning from neonatal mouse forebrain and maintained in growth medium for population expansion. To study the role of PMSC-EVs in OPC differentiation, 1 × 10
5 OPCs were switched to the chemically defined differentiation medium with or without PMSC-EVs (2000 × 10
5). RNA was isolated from differentiating OPCs 24 h after PMSC-EV treatment. RNA isolation was performed using the RNeasy Mini Kit’s manufacturer’s protocol (QIAGEN, Germantown, MD, USA), and RNA concentration was measured with the NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific). Sybr Green-based RT-qPCR was used to quantify the expression of OL differentiation markers oligodendrocyte-specific molecules (MOG), ectonucleotide pyrophosphatase/phosphodiesterase 6 (Enpp6) and myelin associated glycoprotein (MAG). Data were analyzed using the StepOnePlus Real-Time PCR System (Thermo Fisher Scientific) and processed by the V2.3 StepOne software. For quantification, the mRNA expression level of interested genes in each sample was normalized to the internal control, housekeeping gene
Hsp90 and fold change in gene expression was calculated based on the delta-delta Ct method as previously described [
29,
30].
2.15. Statistical Analyses
The results are expressed as mean and standard error. Imaging and in vitro data were analyzed using non-parametric Mann–Whitney-Wilcoxon t-tests (GraphPad Prism version 8.2.1 for macOS, La Jolla, CA, USA). Multiple comparisons were performed using a Kruskal–Wallis test, followed by Dunn’s post hoc correction to determine which groups were significantly different (GraphPad Prism version 8.2.1 for macOS). A p-value < 0.05 was considered significant.
4. Discussion
The goal of the present study was to investigate the use of a novel early-gestational chorionic villus-derived MSC source for the treatment of MS. Furthermore, we aimed to investigate whether PMSC-derived EVs confer therapeutic benefits and whether these particles can be used as a cell-free treatment for neurodegenerative disease. Comparable to findings from other studies using adult-derived MSCs [
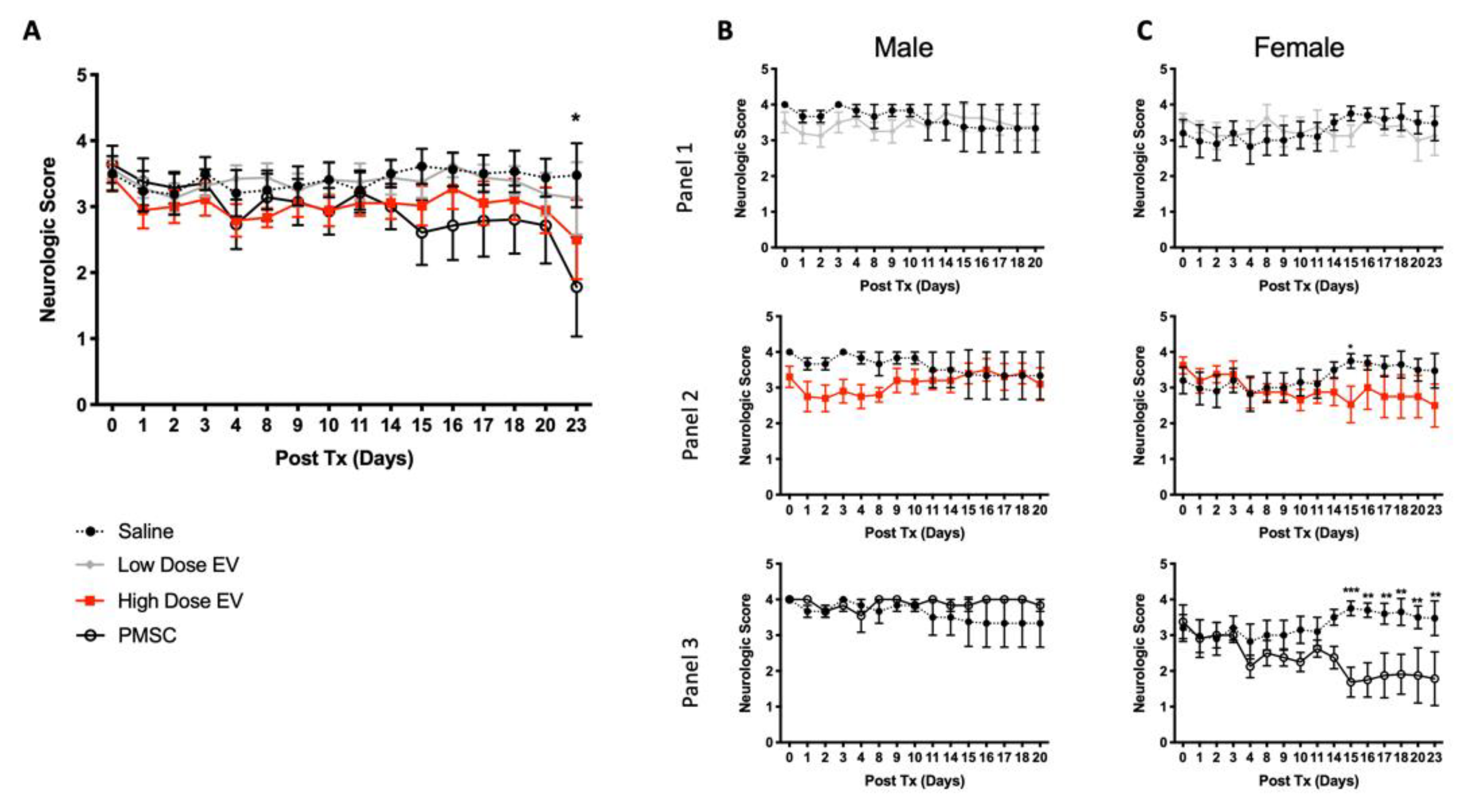
11], PMSCs achieved motor improvement in an EAE murine model of MS. Improvements in motor function were achieved in part through a neuroprotective mechanism. PMSC-treated animals displayed less DNA damage within oligodendrocyte populations and myelin was preserved in the spinal cords of these animals. Interestingly, in the treatment group, animals that did not show motor improvements, mild increases of DNA damage and demyelination were noted. Additionally, similar protective effects preserving oligodendroglia and myelination were also achieved by treatment with high-dose PMSC-derived EVs. These effects were not noted in the low dose PMSC-EV-treatment groups, suggesting that the PMSC secretory mechanism of action occurs through EV signaling in a dose-dependent manner.
These findings demonstrate that the clinical benefits of PMSCs can be achieved by treatment with PMSC-EVs alone. Our group has previously demonstrated that PMSCs secrete high levels of BDNF, HGF and VEGF as compared to BM-MSCs [
21]. Proteomic analysis of PMSC-EVs revealed the presence of HGF and VEGF in these nanoparticles. HGF, in particular, has been shown to be secreted in MSC-conditioned medium and can mediate motor recovery functions in an EAE model of MS [
30]. HGF is a potent angiogenic factor that has been shown to exert immunomodulatory effects through the stimulation of regulatory T cells, which, in turn, mediate autoimmune responses [
31]. These factors may play a role in the protective and regenerative properties of PMSCs. In this study, high-dose PMSC-EV treatments lead to comparable responses to PMSC treatments. This suggests that PMSC-mediated clinical improvements in the current EAE model occurs through an EV-meditated mechanism. The presence of these factors in PMSC-EVs also suggests that these mediators may play an important role in protective, regenerative and immunomodulatory properties of PMSCs. Additionally, PMSCs secrete higher levels of these proteins compared to BM-MSCs [
21], thus, this unique cell source may result in improved clinical outcomes compared to adult-derived MSC sources.
Our in vivo data demonstrate increased myelin present in spinal cord white matter of PMSC and PMSC-EV-treated EAE mice. In these animals, oligodendrocyte survival was increased as compared to saline-treated controls. PMSC and high-dose PMSC-EV treatments led to less DNA damage to oligodendroglia. Oligodendrocytes are myelin producing cells; therefore, increased levels of oligodendrocytes correspond to increased myelin levels. To elucidate whether PMSC-EVs promote oligodendrocyte maturation, differentiation assays were performed. In vitro data demonstrate that PMSC-EVs drive OPCs to express OL differentiation markers, which suggests that PMSC-EVs are promoting maturation of myelinating oligodendroglia. These findings suggest that PMSCs and PMSC-EVs have both protective and regenerative properties, exhibited by driving OPC differentiation. Additionally, interactions of oligodendrocytes and autoreactive T lymphocytes are not well understood, therefore, investigations of these interactions in response to PMSC and PMSC-EV therapy are warranted.
Limitations of using an EAE model have been widely noted, and although the model is reliable to induce MS-like symptoms, including inflammation and demyelination, variation in disease onset occurs even within genetically identical rodents [
32]. Variability in disease onset was observed in this study; however, a standardized average score within each experimental group was used prior to experimental treatment to account for these alterations. This variability likely affected motor function and recovery data and could explain why some animals did not respond to treatment with PMSCs or high-dose PMSC-EVs. Additionally, for the purposes of this study, both male and female mice were used; however, a timelier improvement was noted in female mice. Sex differences have been noted in EAE onset, which reflects higher incidences of MS in human female populations [
33]. These alterations may be due to effects of androgens on T lymphocytes, which are key mediators in the pathologic features of MS [
34]. Sex differences in response to EAE onset methodologies can occur in multiple mouse strains, and it has been shown that male mice do not result in robust Th1 responses and instead shift to anti-inflammatory Th2 responses as compared to females [
33]. Sex differences are not typically noted by C57BL/6 mice; however, in the current study, EAE onset by males and females differed, but to recapitulate MS patient populations, both sexes were included for analysis.
Myelin degeneration is a main pathologic feature of MS; however, autoimmune infiltration of Th1 pro-inflammatory cells into the CNS is a key contributor to the inflammatory and degenerative properties of MS [
35]. T regulatory cells modulate Th1 and Th2 responses and have impaired functions in MS patients [
36]. MSCs have been shown to induce T regulatory populations and drive shifts of Th1 to Th2 responses [
36]. The induction of T regulatory responses by MSCs is a mechanism by which these cells modulate immune responses and may have clinical utility for autoimmune diseases such as MS. Given that male mice have a stronger tendency towards Th1 responses in EAE [
33], this could explain the modest motor recovery responses observed in these animals to PMSC and PMSC-EV treatment. While the focus of the present study was on the myelin protective and regenerative properties of PMSCs and PMSC-EVs, future studies will investigate the immunoregulatory properties of these cells and nanovesicles within immune cell subsets.
Although PMSC and PMSC-EV treatment resulted in motor improvements following a single injection in mice in the acute phase of EAE, future studies are needed to address dosage timing and strategy. For this study, small volumes of diluted EVs and PMSCs were injected via tail veins of mice. However, it has been reported that PMSCs have poor homing and engraftment [
2]. Additionally, nanoparticle treatments pose technical challenges because injections need to be precise to ensure proper and complete administration of therapeutic doses. In future studies, tracking cells and EVs, as well as evaluation of different administration modalities will be performed. Biodistribution studies will provide key insights to further improve clinical outcomes for MS patients.
The findings from this study demonstrate that PMSC-EVs are a viable option for the treatment of neurodegenerative diseases and pose several advantages compared to cellular-based therapies. EVs provide many benefits, including immunotolerance, storage stability, heterogenous cargo and multiple therapeutic outcomes. This study provides key preliminary data that will facilitate future studies investigating the use of PMSC-EVs for the treatment of MS.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





